
Lignin Promotes Mycelial Growth and Accumulation of Polyphenols and Ergosterol in Lentinula edodes
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Materials
2.2. Culture Condition
2.3. Determinations of Mycelium Biomass and Compositional Analysis
2.4. Phenolic Compounds Extraction and Determination
2.5. Enzyme Assays
2.6. Statistical Analysis
3. Results and Discussions
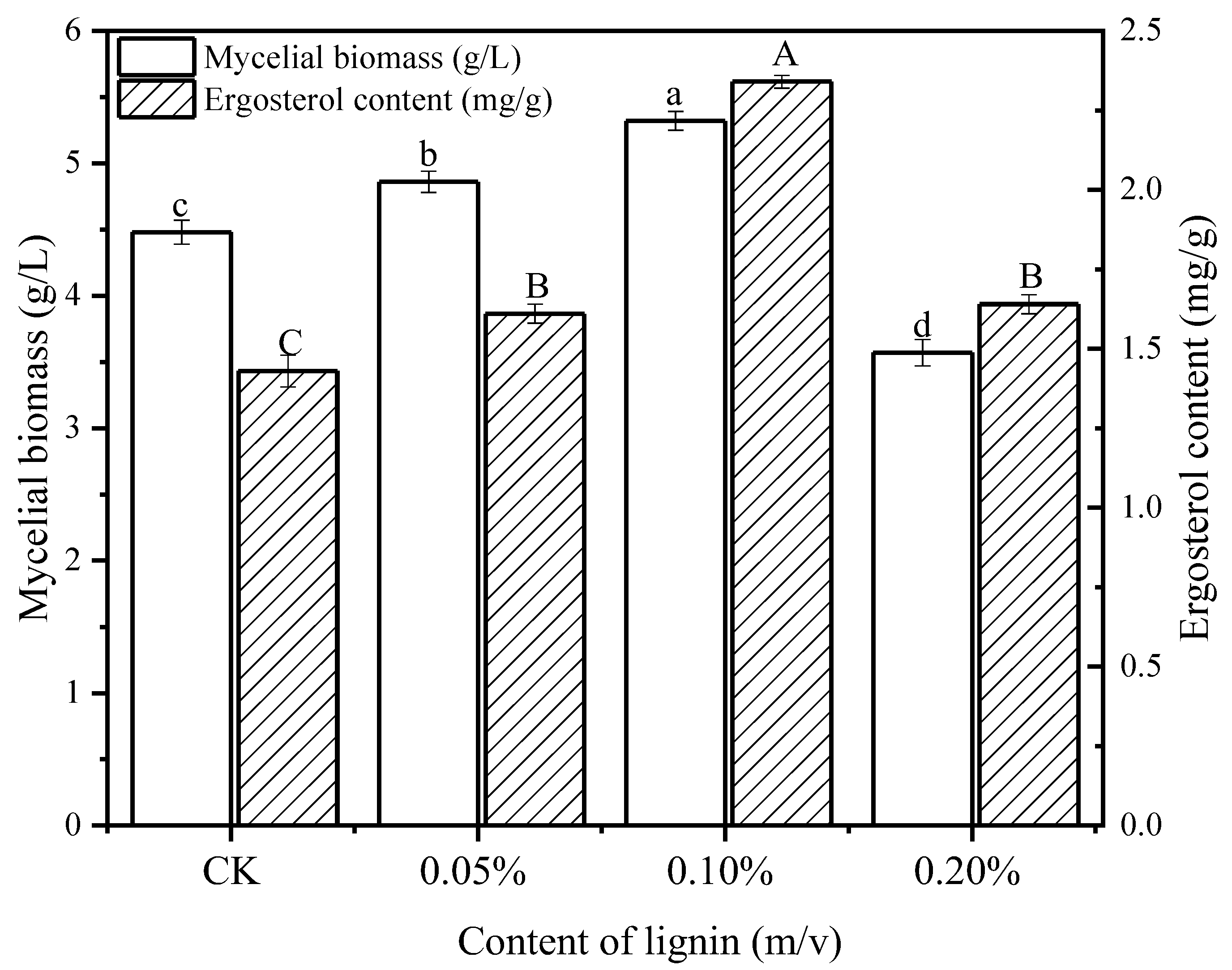
3.1. Effect of Lignin on Mycelia Growth
3.2. Effect of Lignin on Chemical Compositions of L. edodes Mycelium
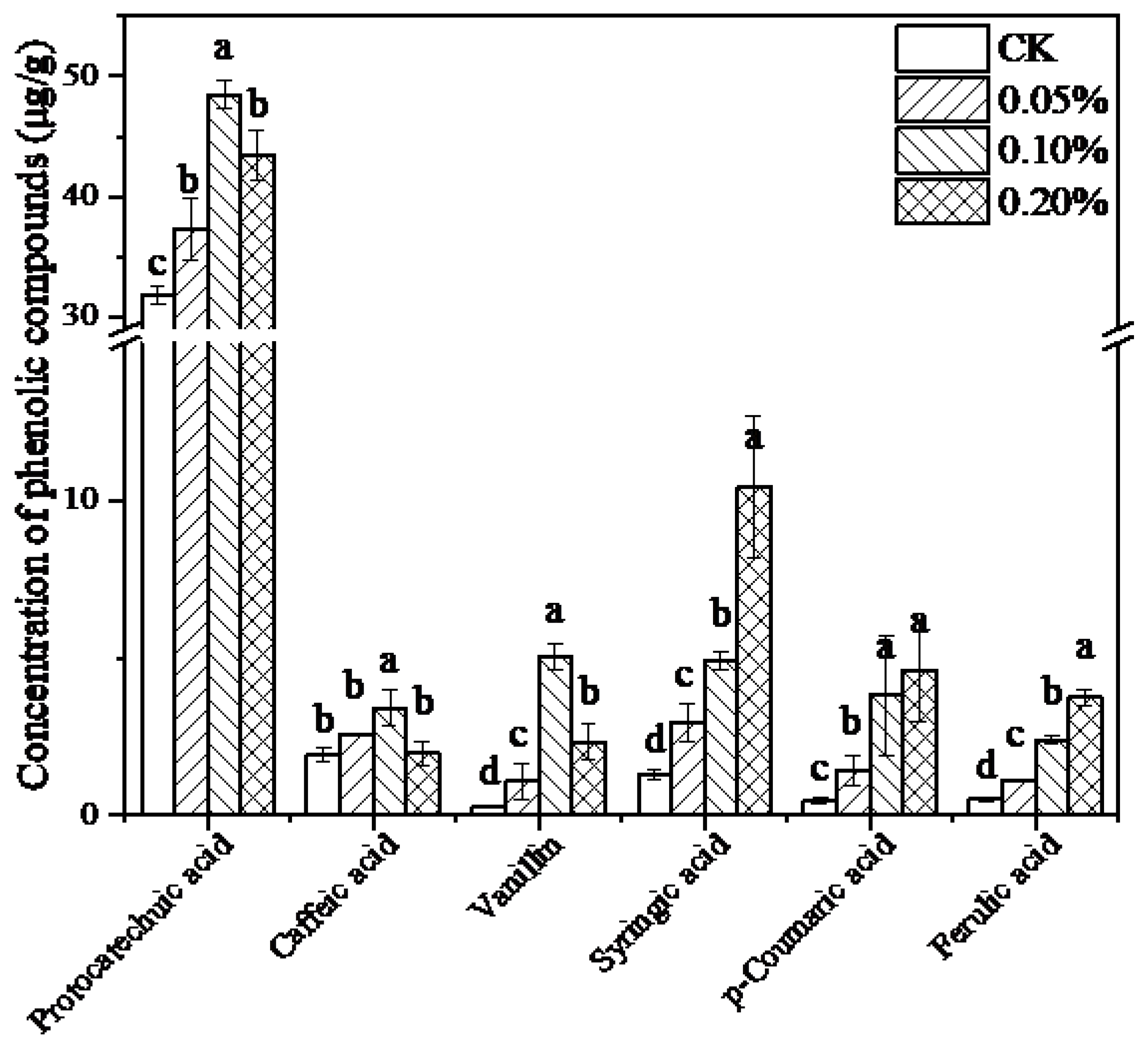
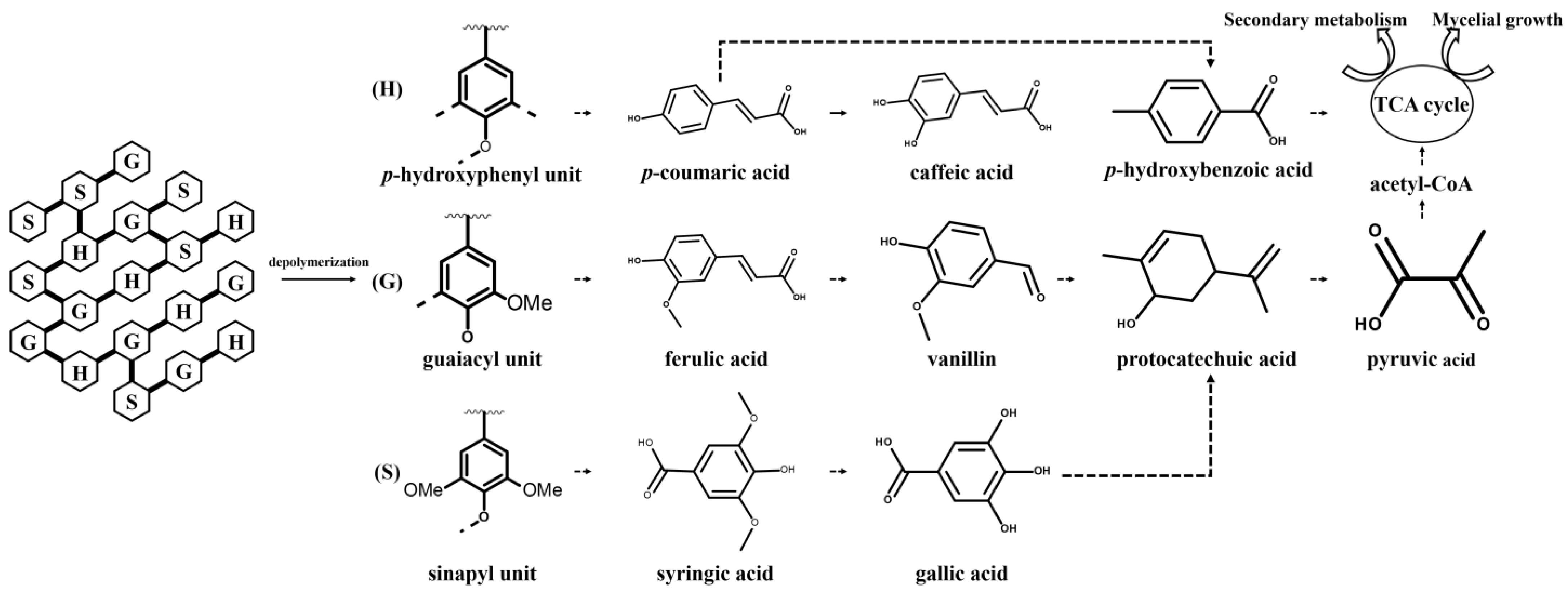
3.3. Effect of Lignin on Phenolic Compounds Metabolism in Mycelia of L. edodes
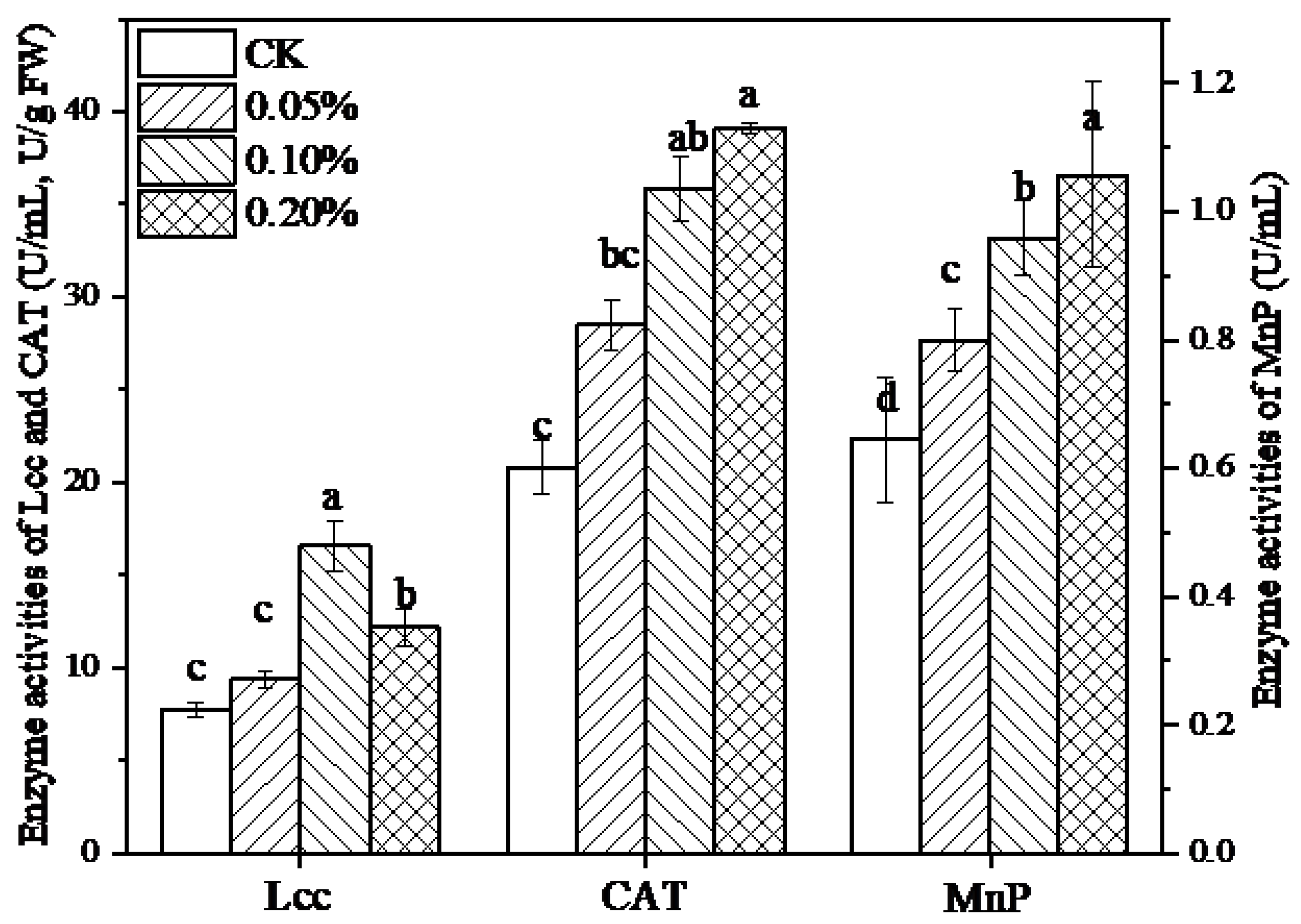
3.4. Effect of Lignin on Ligninolytic Enzymes in L. edodes Mycelia
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chio, C.; Sain, M.; Qin, W. Lignin utilization: A review of lignin depolymerization from various aspects. Renew. Sustain. Energy Rev. 2019, 107, 232–249. [Google Scholar] [CrossRef]
- Li, C.; Zhao, X.; Wang, A.; Huber, G.W.; Zhang, T. Catalytic transformation of lignin for the production of chemicals and fuels. Chem. Rev. 2015, 115, 11559–11624. [Google Scholar] [CrossRef]
- Weng, C.; Peng, X.; Han, Y. Depolymerization and conversion of lignin to value-added bioproducts by microbial and enzymatic catalysis. Biotechnol. Biofuels 2021, 14, 84. [Google Scholar] [CrossRef] [PubMed]
- Del Cerro, C.; Erickson, E.; Dong, T.; Wong, A.R.; Eder, E.K.; Purvine, S.O.; Mitchell, H.D.; Weitz, K.K.; Markillie, L.M.; Burnet, M.C.; et al. Intracellular pathways for lignin catabolism in white-rot fungi. Proc. Natl. Acad. Sci. USA 2021, 118, e2017381118. [Google Scholar] [CrossRef] [PubMed]
- Janusz, G.; Pawlik, A.; Sulej, J.; Świderska-Burek, U.; Jarosz-Wilkołazka, A.; Paszczyński, A. Lignin degradation: Microorganisms, enzymes involved, genomes analysis and evolution. FEMS Microbiol. Rev. 2017, 41, 941–962. [Google Scholar] [CrossRef] [PubMed]
- Bugg, T.D.H.; Ahmad, M.; Hardiman, E.M.; Rahmanpour, R. Pathways for degradation of lignin in bacteria and fungi. Nat. Prod. Rep. 2011, 28, 1883–1896. [Google Scholar] [CrossRef]
- Kunjapur, A.M.; Prather, K.L.J. Development of a vanillate biosensor for the vanillin biosynthesis pathway in E. Coli. ACS Synth. Biol. 2019, 8, 1958–1967. [Google Scholar] [CrossRef]
- Sonoki, T.; Takahashi, K.; Sugita, H.; Hatamura, M.; Azuma, Y.; Sato, T.; Suzuki, S.; Kamimura, N.; Masai, E. Glucose-Free cis,cis-Muconic Acid Production via New Metabolic Designs Corresponding to the Heterogeneity of Lignin. ACS Sustain. Chem. Eng. 2018, 6, 1256–1264. [Google Scholar] [CrossRef]
- Liu, L.; Liu, H.; Zhang, W.; Yao, M.; Li, B.; Liu, D.; Yuan, Y. Engineering the biosynthesis of caffeic acid in saccharomyces cerevisiae with heterologous enzyme combinations. Engineering 2019, 5, 287–295. [Google Scholar] [CrossRef]
- Yaguchi, A.; Franaszek, N.; O’Neill, K.; Lee, S.; Blenner, M. Identification of oleaginous yeasts that metabolize aromatic compounds. J. Ind. Microbiol. Biotechnol. 2020, 47, 801–813. [Google Scholar] [CrossRef]
- Chatterjee, A.; Delorenzo, D.M.; Carr, R.; Moon, T.S. Bioconversion of renewable feedstocks by Rhodococcus opacus. Curr. Opin. Biotech. 2019, 64, 10–16. [Google Scholar] [CrossRef]
- Sheng, K.; Wang, C.; Chen, B.; Kang, M.; Wang, M.; Liu, K.; Wang, M. Recent advances in polysaccharides from Lentinus edodes (Berk.): Isolation, structures and bioactivities. Food Chem. 2021, 358, 129883. [Google Scholar] [CrossRef]
- Tang, W.; Liu, C.; Liu, J.; Hu, L.; Huang, Y.; Yuan, L.; Liu, F.; Pan, S.; Chen, S.; Bian, S.; et al. Purification of polysaccharide from Lentinus edodes water extract by membrane separation and its chemical composition and structure characterization. Food Hydrocoll. 2020, 105, 105851. [Google Scholar] [CrossRef]
- Thetsrimuang, C.; Khammuang, S.; Chiablaem, K.; Srisomsap, C.; Sarnthima, R. Antioxidant properties and cytotoxicity of crude polysaccharides from Lentinus polychrous Lév. Food Chem. 2011, 128, 634–639. [Google Scholar] [CrossRef]
- Liu, Q.; Cui, X.; Song, Z.; Kong, W.; Ng, T.B. Coating shiitake mushrooms (Lentinus edodes) with a polysaccharide from Oudemansiella radicata improves product quality and flavor during postharvest storage. Food Chem. 2021, 352, 129357. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Gong, Y.; Cai, Y.; Liu, W.; Zhou, Y.; Xiao, Y.; Xu, Z.; Liu, Y.; Lei, X.; Wang, G.; et al. Genome Sequence of the Edible Cultivated Mushroom Lentinula edodes (Shiitake) Reveals Insights into Lignocellulose Degradation. PLoS ONE 2016, 11, e160336. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, Y.; Nakade, K.; Sato, S.; Yoshida, K.; Konno, N. Lentinula edodes Genome Survey and Postharvest Transcriptome Analysis. Appl. Environ. Microbiol. 2017, 83, 2916–2990. [Google Scholar] [CrossRef]
- Wong, K.; Cheung, M.; Au, C.; Kwan, H. A novel Lentinula edodes laccase and its comparative enzymology suggest Guaiacol-Based laccase engineering for bioremediation. PLoS ONE 2013, 8, e66426. [Google Scholar] [CrossRef]
- De Oliveira Júnior, S.D.; Dos Santos Gouvêa, P.R.; de Aguiar, L.V.B.; Pessoa, V.A.; Dos Santos Cruz Costa, C.L.; Chevreuil, L.R.; Dedo Britonascimento, L.B.; Dos Santos, E.S.; Sales-Campos, C. Production of Lignocellulolytic Enzymes and Phenolic Compounds by Lentinus strigosus from the Amazon Using Solid-State Fermentation (SSF) of Guarana (Paullinia cupana) Residue. Appl. Biochem. Biotech. 2022, 194, 2882–2900. [Google Scholar] [CrossRef]
- Gaitan-Hernandez, R.; Zavaleta, M.; Aquino-Bolanos, E.N. Productivity, Physicochemical Changes, and Antioxidant Activity of Shiitake Culinary-Medicinal Mushroom Lentinus edodes (Agaricomycetes) Cultivated on Lignocellulosic Residues. Int. J. Med. Mushrooms 2017, 19, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.H.; Xia, X.; Lin, C.X.; Tong, D.S.; Beltramini, J. Catalytic conversion of lignocellulosic biomass to fine chemicals and fuels. Chem. Soc. Rev. 2011, 40, 5588. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Jia, X.; Yin, L.; Cheng, Y.; Miao, Y.; Zhang, X. The Effect of Hemicellulose and Lignin on Properties of Polysaccharides in Lentinus edodes and their Antioxidant Evaluation. Molecules 2019, 24, 1834. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Gong, Y.; Liu, W.; Hu, Y.; Chen, L.; Yan, L.; Zhou, Y.; Bian, Y. Comparative secretomic analysis of lignocellulose degradation by Lentinula edodes grown on microcrystalline cellulose, lignosulfonate and glucose. J. Proteom. 2017, 163, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Niemenmaa, O.; Galkin, S.; Hatakka, A. Ergosterol contents of some wood-rotting basidiomycete fungi grown in liquid and solid culture conditions. Int. Biodeter. Biodegr. 2008, 62, 125–134. [Google Scholar] [CrossRef]
- Feki, F.; Klisurova, D.; Masmoudi, M.A.; Choura, S.; Denev, P.; Trendafilova, A.; Chamkha, M.; Sayadi, S. Optimization of microwave assisted extraction of simmondsins and polyphenols from Jojoba (Simmondsia chinensis) seed cake using Box-Behnken statistical design. Food Chem. 2021, 356, 129670. [Google Scholar] [CrossRef]
- Gbylik-Sikorska, M.; Gajda, A.; Nowacka-Kozak, E.; Posyniak, A. Simultaneous determination of 45 antibacterial compounds in mushrooms—Agaricus bisporus by ultra-high performance liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2019, 1587, 111–118. [Google Scholar] [CrossRef]
- Teshome, E.; Melaku, A. Selection and optimization of lignocellulosic substrate for laccase production from Pleurotus species. Int. J. Biotechnol. Mol. Biol. Res. 2017, 8, 38–48. [Google Scholar] [CrossRef]
- Lechner, B.E.; Papinutti, V.L. Production of lignocellulosic enzymes during growth and fruiting of the edible fungus Lentinus tigrinus on wheat straw. Process Biochem. 2006, 41, 594–598. [Google Scholar] [CrossRef]
- Zhu, Z.; Liu, X.; Tang, Y.; Dong, F.; Sun, H.; Chen, L.; Zhang, Y. Effects of cultural medium on the formation and antitumor activity of polysaccharides by Cordyceps gunnii. J. Biosci. Bioeng. 2016, 122, 494–498. [Google Scholar] [CrossRef]
- Kim, H.O.; Lim, J.M.; Joo, J.H.; Kim, S.W.; Hwang, H.J.; Choi, J.W.; Yun, J.W. Optimization of submerged culture condition for the production of mycelial biomass and exopolysaccharides by Agrocybe cylindracea. Bioresour. Technol. 2005, 96, 1175–1182. [Google Scholar] [CrossRef]
- Bogdan, V.I.; Sergeeva, Y.E.; Lunin, V.V.; Perminova, I.V.; Konstantinov, A.I.; Zinchenko, G.E.; Bogdan, K.V. Bioconversion of phenolic monomers of lignin and Lignin-Containing substrates by the basidiomycete Lentinus tigrinus. Appl. Biochem. Microbiol. 2018, 54, 198–205. [Google Scholar] [CrossRef]
- Vikman, M.; Karjomaa, S.; Kapanen, A.; Wallenius, K.; Itävaara, M. The influence of lignin content and temperature on the biodegradation of lignocellulose in composting conditions. Appl. Microbiol. Biotechnol. 2002, 59, 591–598. [Google Scholar] [PubMed]
- Qiu, W.; Liu, J. Fermenting and Lignin Degradability of a White-Rot Fungus Coriolopsis trogii Using Industrial Lignin as Substrate. Appl. Biochem. Biotechnol. 2022, 194, 5220–5235. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, Y.; Okada, K.; Kawai, G.; Motai, H. Efficient Production of Mycelium of Lentinus edodes by a Continuous Culture and the Effect of Lignin on Growth. J. Ferment. Bioeng. 1993, 1, 45–48. [Google Scholar] [CrossRef]
- Tsujiyama, S.; Muraoka, T.; Takada, N. Biodegradation of 2,4-dichlorophenol by shiitake mushroom (Lentinula edodes) using vanillin as an activator. Biotechnol. Lett. 2013, 35, 1079–1083. [Google Scholar] [CrossRef]
- Iivashechkin, A.A.; Sergeeva, I.; Lunin, V.V.; Bogdan, V.I.; Mysiakina, I.S.; Feofilova, E.P. Influence of lignin and oxygen on the growth and the lipid formation of the fungus Lentinus tigrinus. Prikl. Biokhim. Mikrobiol. 2014, 50, 318–323. [Google Scholar] [CrossRef]
- Mccue, P.P.; Shetty, K. A model for the involvement of lignin degradation enzymes in phenolic antioxidant mobilization from whole soybean during solid-state bioprocessing by Lentinus edodes. Process Biochem. 2005, 40, 1143–1150. [Google Scholar] [CrossRef]
- Koselny, K.; Mutlu, N.; Minard, A.Y.; Kumar, A.; Krysan, D.J.; Wellington, M. A Genome-Wide screen of deletion mutants in the filamentous saccharomyces cerevisiae background identifies ergosterol as a direct trigger of macrophage pyroptosis. mBio 2018, 9, e01204-18. [Google Scholar] [CrossRef]
- Liu, J.; Xia, J.; Nie, K.; Wang, F.; Deng, L. Outline of the biosynthesis and regulation of ergosterol in yeast. World J. Microbiol. Biotechnol. 2019, 35, 98. [Google Scholar] [CrossRef]
- Blaga, A.C.; Ciobanu, C.; Caşcaval, D.; Galaction, A. Enhancement of ergosterol production by Saccharomyces cerevisiae in batch and fed-batch fermentation processes using n-dodecane as oxygen-vector. Biochem. Eng. J. 2018, 131, 70–76. [Google Scholar] [CrossRef]
- Silva, R.R.; Corso, C.R.; Matheus, D.R. Effect of culture conditions on the biomass determination by ergosterol of Lentinus crinitus and Psilocybe castanella. World J. Microbiol. Biotechnol. 2010, 26, 841–846. [Google Scholar] [CrossRef]
- Lu, X.; Wang, C.; Li, Y.; Liu, P. Improved production and antioxidant activity of exopolysaccharides by submerged culture of Lentinula edodes by the addition of lignocellulose. J. Biosci. Bioeng. 2022, 134, 162–166. [Google Scholar] [CrossRef]
- Kozarski, M.; Klaus, A.; Nikšić, M.; Vrvić, M.M.; Todorović, N.; Jakovljević, D.; Van Griensven, L.J.L.D. Antioxidative activities and chemical characterization of polysaccharide extracts from the widely used mushrooms Ganoderma applanatum, Ganoderma lucidum, Lentinus edodes and Trametes versicolor. J. Food Compos. Anal. 2012, 26, 144–153. [Google Scholar] [CrossRef]
- Li, F.; Cui, S.H.; Zha, X.Q.; Bansal, V.; Jiang, Y.L.; Asghar, M.N.; Wang, J.H.; Pan, L.H.; Xu, B.F.; Luo, J.P. Structure and bioactivity of a polysaccharide extracted from protocorm-like bodies of Dendrobium huoshanense. Int. J. Biol. Macromol. Struct. Funct. Interact. 2015, 72, 664–672. [Google Scholar] [CrossRef]
- Karnaouri, A.; Muraleedharan, M.N.; Dimarogona, M.; Topakas, E.; Rova, U.; Sandgren, M.; Christakopoulos, P. Recombinant expression of thermostable processive MtEG5 endoglucanase and its synergism with MtLPMO from Myceliophthora thermophila during the hydrolysis of lignocellulosic substrates. Biotechnol. Biofuels 2017, 10, 126. [Google Scholar] [CrossRef]
- Carneiro, A.A.J.; Ferreira, I.C.F.R.; Dueñas, M.; Barros, L.; Da Silva, R.; Gomes, E.; Santos-Buelga, C. Chemical composition and antioxidant activity of dried powder formulations of Agaricus blazei and Lentinus edodes. Food Chem. 2013, 138, 2168–2173. [Google Scholar] [CrossRef]
- Wu, X.J.; Hansen, C. Effects of Whey Permeate-Based Medium on the Proximate Composition of Lentinus edodes in the Submerged Culture. J. Food Sci. 2006, 71, M174–M179. [Google Scholar] [CrossRef]
- Kim, M.; Seguin, P.; Ahn, J.; Kim, J.; Chun, S.; Kim, E.; Seo, S.; Kang, E.; Kim, S.; Park, Y.; et al. Phenolic compound concentration and antioxidant activities of edible and medicinal mushrooms from Korea. J. Agric. Food Chem. 2008, 56, 7265–7270. [Google Scholar] [CrossRef]
- Li, X.; Zheng, Y. Biotransformation of lignin: Mechanisms, applications and future work. Biotechnol. Prog. 2019, 36, e2922. [Google Scholar] [CrossRef] [PubMed]
- Yaguchi, A.L.; Lee, S.J.; Blenner, M.A. Synthetic Biology towards Engineering Microbial Lignin Biotransformation. Trends Biotechnol. 2021, 39, 1037–1064. [Google Scholar] [CrossRef]
- Tomizawa, S.; Chuah, J.; Matsumoto, K.; Doi, Y.; Numata, K. Understanding the limitations in the biosynthesis of polyhydroxyalkanoate (PHA) from lignin derivatives. ACS Sustain. Chem. Eng. 2014, 2, 1106–1113. [Google Scholar] [CrossRef]
- Johnson, C.W.; Beckham, G.T. Aromatic catabolic pathway selection for optimal production of pyruvate and lactate from lignin. Metab. Eng. 2015, 28, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Lau, B.F.; Abdullah, N. Bioprospecting of Lentinus squarrosulus Mont., An underutilized wild edible mushroom, as a potential source of functional ingredients: A review. Trends Food Sci. Technol. 2017, 61, S0924224416302898. [Google Scholar] [CrossRef]
- Xue, Y.; Zhang, Y.; Grace, S.; He, Q. Functional expression of an Arabidopsis p450 enzyme, p-coumarate-3-hydroxylase, in the cyanobacterium Synechocystis PCC 6803 for the biosynthesis of caffeic acid. J. Appl. Phycol. 2014, 26, 219–226. [Google Scholar] [CrossRef]
- Elisashvili, V.; Kachlishvili, E.; Khardziani, T.; Agathos, S.N. Effect of aromatic compounds on the production of laccase and manganese peroxidase by white-rot basidiomycetes. J. Ind. Microbiol. Biotechnol. 2010, 37, 1091–1096. [Google Scholar] [CrossRef]
- Shutova, V.V.; Revin, V.V.; Makushina, I. The effect of copper ions on the production of laccase by the fungus Lentinus (Panus) tigrinus. Prikl. Biokhim. Mikrobiol. 2008, 44, 683–687. [Google Scholar] [CrossRef]
- Elisashvili, V.; Penninckx, M.; Kachlishvili, E.; Tsiklauri, N.; Metreveli, E.; Kharziani, T.; Kvesitadze, G. Lentinus edodes and Pleurotus species lignocellulolytic enzymes activity in submerged and solid-state fermentation of lignocellulosic wastes of different composition. Bioresour. Technol. 2008, 99, 457–462. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lignin | WSP Extracts (g/100 g) | Crude Protein (w/w %) | Ash (w/w %) | Total Phenolics (mg GAE/g DW) |
|---|---|---|---|---|
| 0 | 2.11 ± 0.14 d | 25.84 ± 0.35 c | 3.78 ± 0.07 d | 1.45 ± 0.03 d |
| 0.05% | 4.14 ± 0.35 b | 27.63 ± 0.26 b | 4.89 ± 0.09 c | 2.28 ± 0.05 c |
| 0.10% | 6.44 ± 0.33 a | 29.24 ± 0.09 a | 5.21 ± 0.11 b | 3.87 ± 0.14 b |
| 0.20% | 2.94 ± 0.28 c | 26.17 ± 0.18 c | 6.14 ± 0.17 a | 5.62 ± 0.21 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, F.; Wang, H.; Chen, Q.; Pang, X.; Jing, H.; Yin, L.; Zhang, X. Lignin Promotes Mycelial Growth and Accumulation of Polyphenols and Ergosterol in Lentinula edodes. J. Fungi 2023, 9, 237. https://doi.org/10.3390/jof9020237
Wu F, Wang H, Chen Q, Pang X, Jing H, Yin L, Zhang X. Lignin Promotes Mycelial Growth and Accumulation of Polyphenols and Ergosterol in Lentinula edodes. Journal of Fungi. 2023; 9(2):237. https://doi.org/10.3390/jof9020237
Chicago/Turabian StyleWu, Feifei, Heqin Wang, Qiufeng Chen, Xiao Pang, Hao Jing, Lijun Yin, and Xiuqing Zhang. 2023. "Lignin Promotes Mycelial Growth and Accumulation of Polyphenols and Ergosterol in Lentinula edodes" Journal of Fungi 9, no. 2: 237. https://doi.org/10.3390/jof9020237