
Phenotypic and Genomic Difference among Four Botryosphaeria Pathogens in Chinese Hickory Trunk Canker

Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin and Collection of Botryosphaeria Species
2.2. Phenotypic Characterization of Botryosphaeria Species
2.3. Comparison of Genomic Differences among Botryosphaeria Species
2.4. B. dothidea Rapid Identification Method with Species-Specific Primers
3. Results
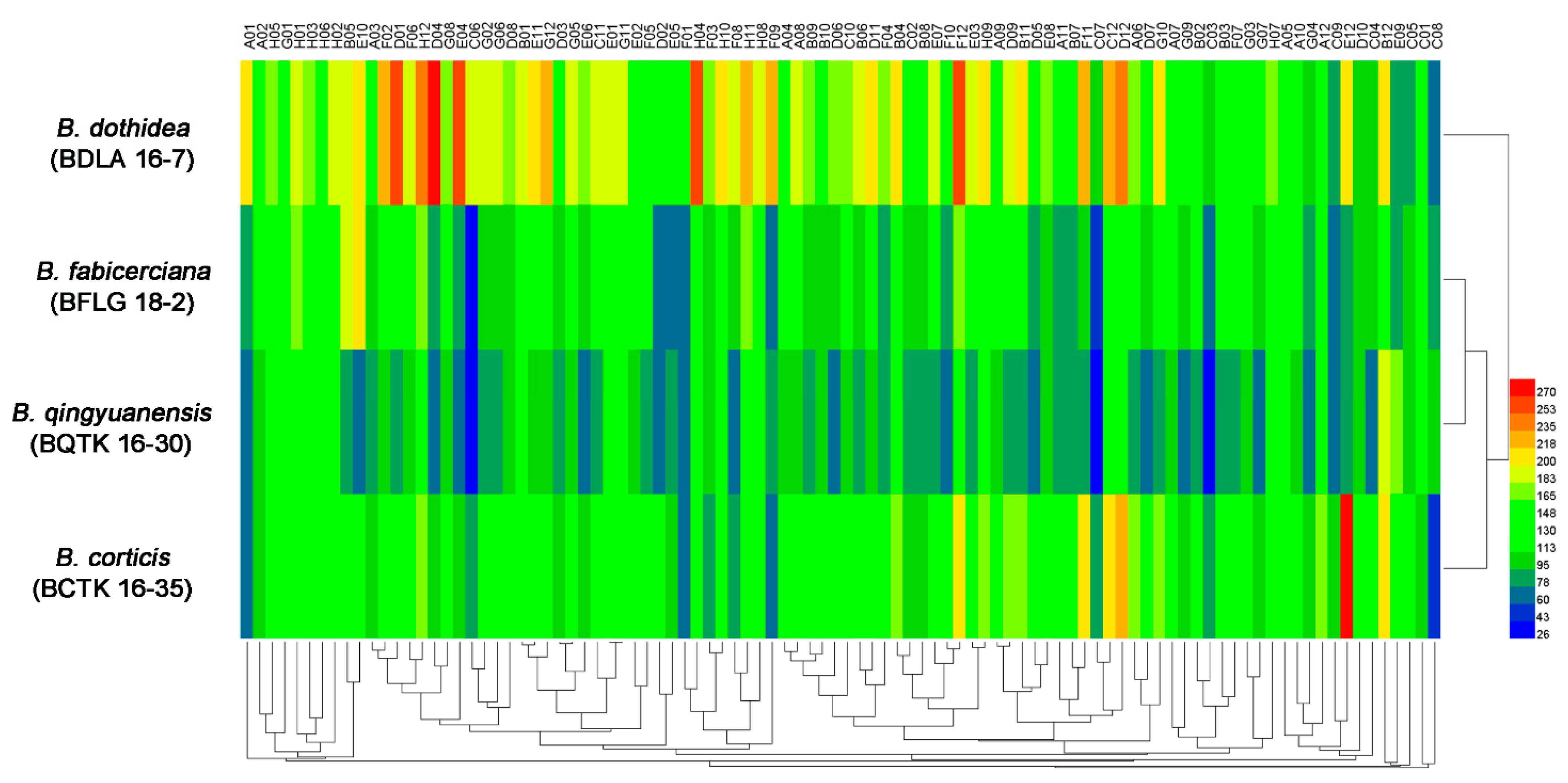
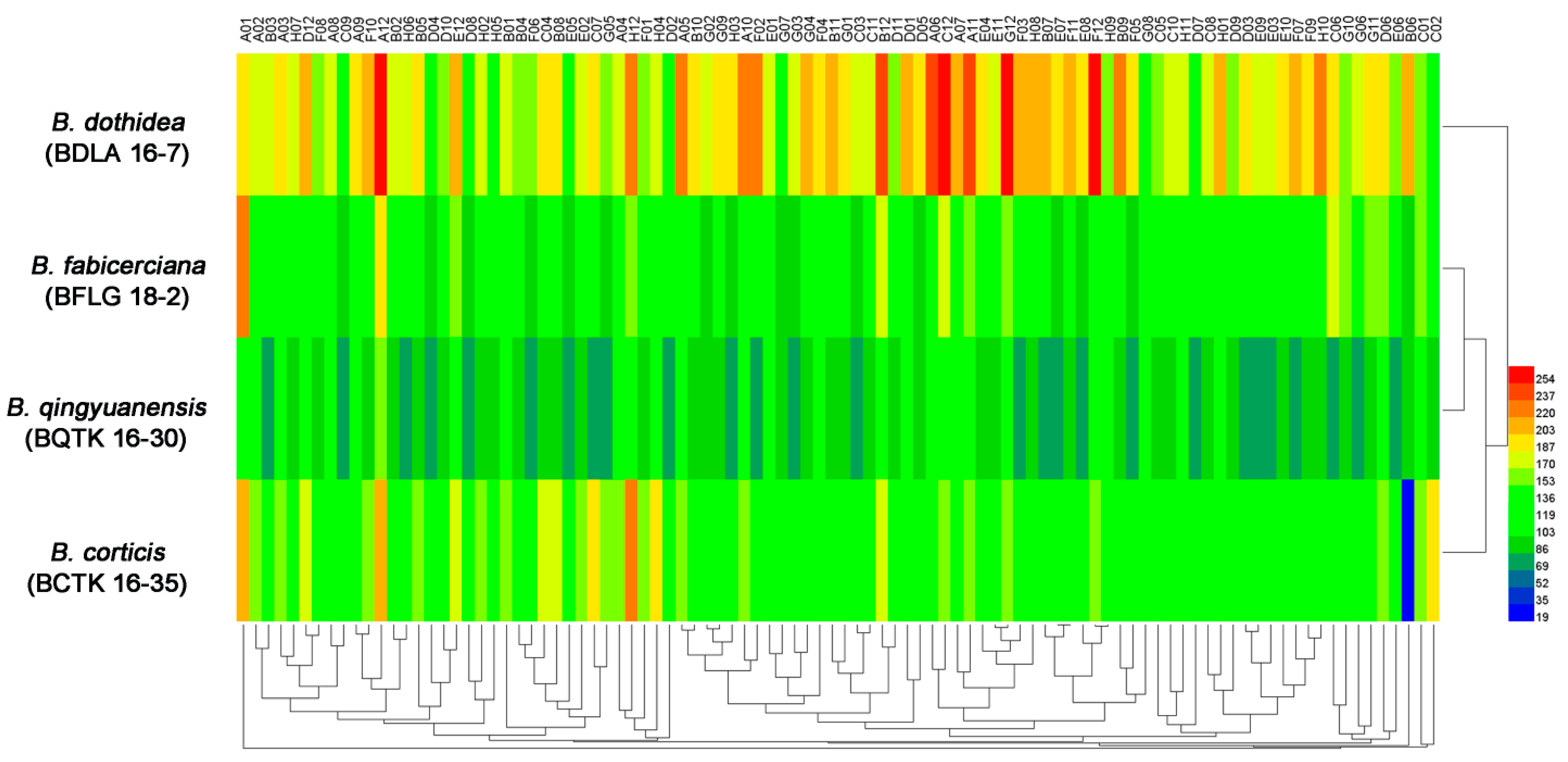
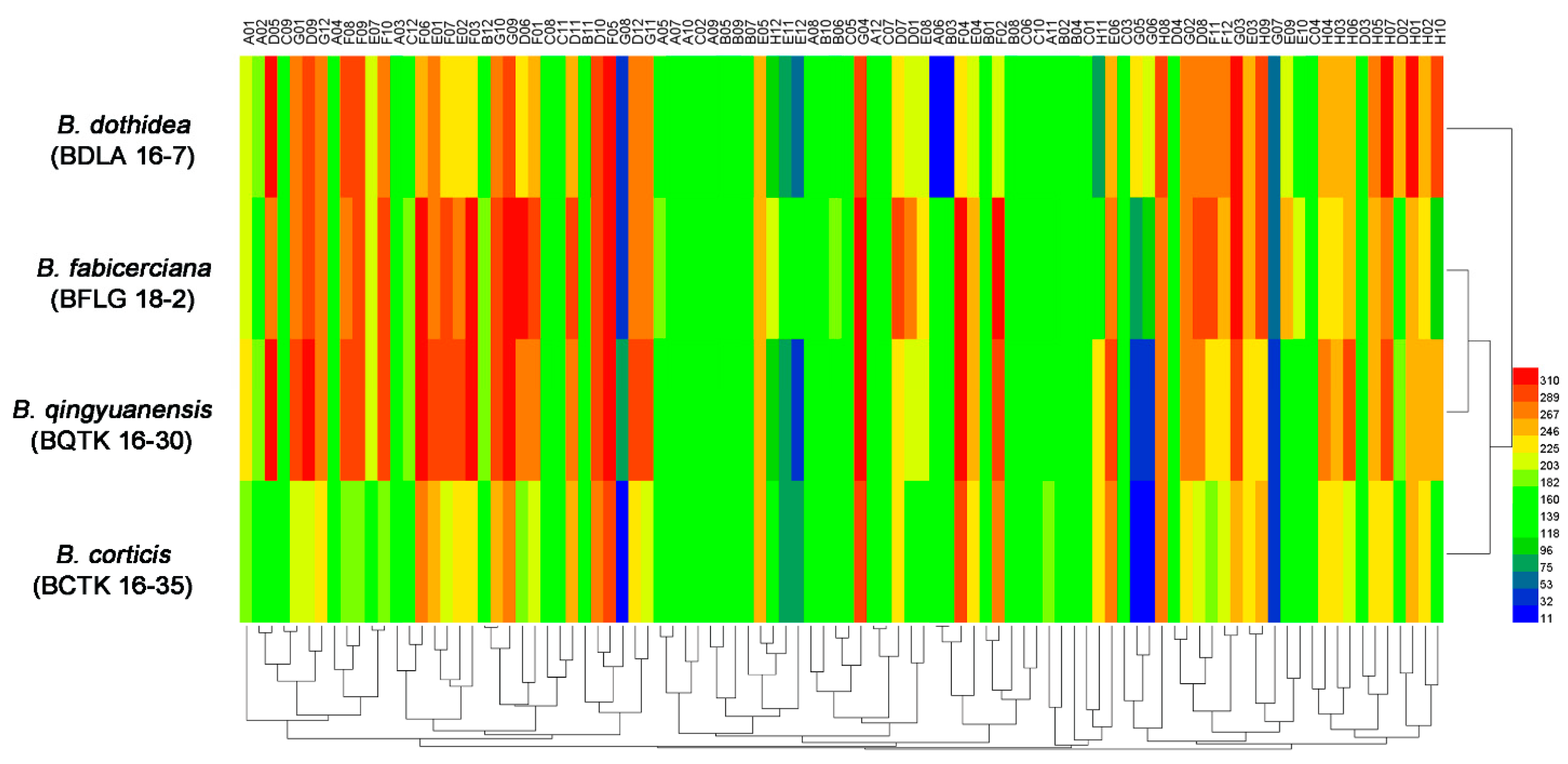
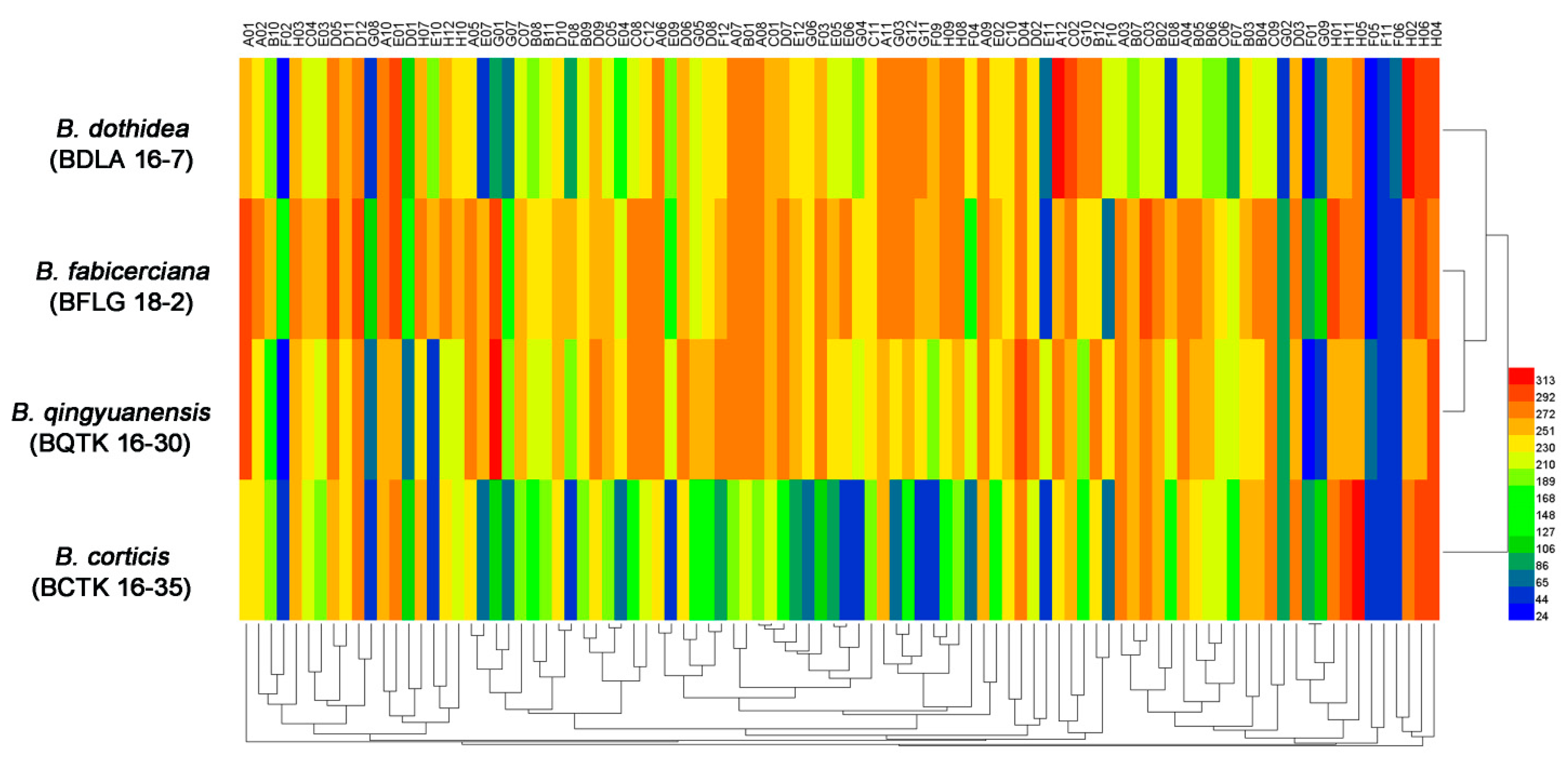
3.1. Phenotypic Characterization of Four Botryosphaeria Species
3.2. Comparison of Genomic Differences among Botryosphaeria Species
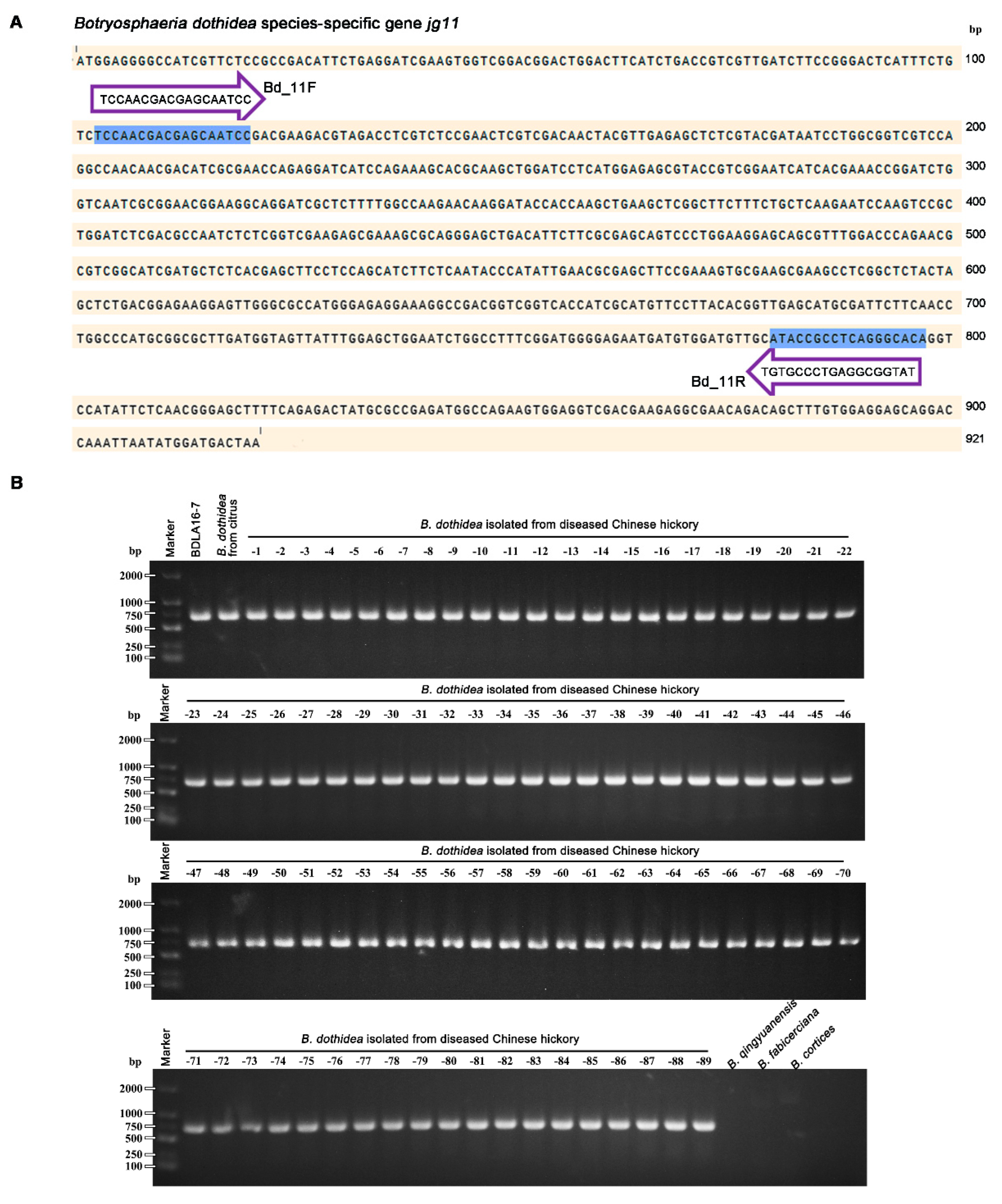
3.3. Design and Application of Primers for B. dothidea Molecular Identification
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, C.G.; Deng, X.Y.; Shi, F. Evaluation of the antioxidant activity of Chinese hickory Carya cathayensis kernel ethanol extraction. Afr. J. Biotechnol. 2008, 7, 2169–2173. [Google Scholar]
- Dai, J.D.; Wang, H.D.; Wang, Y.P.; Zhang, C.Q. Management of Chinese hickory (Carya cathayensis) trunk canker through effective fungicide application programs and baseline sensitivity of Botryosphaeria dothidea to trifloxystrobin. Australas Plant Path. 2017, 46, 75–82. [Google Scholar] [CrossRef]
- Wang, Q.W.; Zhang, C.Q. q-LAMP assays for the detection of Botryosphaeria dothidea causing Chinese hickory canker in trunk, water, and air samples. Plant Dis. 2019, 103, 3142–3149. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Mei, L.; Jiang, H.; Hu, G.; Wang, Y. Evaluation of Carya cathayensis resistance to Botryosphaeria trunk canker using grafting on pecan. Sci. Hortic. 2019, 248, 184–188. [Google Scholar] [CrossRef]
- Zhuang, C.J.; Wang, Q.W.; Wu, Q.Q.; Qiu, Z.L.; Xu, B.C.; Zhang, C. Diversity of Botryosphaeriaceae species associated with Chinese hickory tree (Carya cathayensis) trunk cankers. Plant Dis. 2021, 105, 3869–3879. [Google Scholar] [CrossRef]
- Phillips, A.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef]
- Xu, S.E.; Yao, J.L.; Wu, F.; Mei, L.; Wang, Y.J. Evaluation of Paenibacillus polymyxa carboxymethylcellulose/poly (vinyl alcohol) formulation for control of Carya cathayensis canker caused by Botryosphaeria dothidea. Forest Pathol. 2018, 48, e12464. [Google Scholar] [CrossRef]
- Adesemoye, A.O.; Mayorquin, J.S.; Wang, D.H.; Twizeyimana, M.; Lynch, S.C.; Eskalen, A. Identification of species of Botryosphaeriaceae causing bot gummosis in citrus in California. Plant Dis. 2014, 98, 55–61. [Google Scholar] [CrossRef]
- Mayorquin, J.S.; Wang, D.H.; Twizeyimana, M.; Eskalen, A. Identification, distribution, and pathogenicity of Diatrypaceae and Botryosphaeriaceae associated with citrus branch canker in the southern California desert. Plant Dis. 2016, 100, 2402–2413. [Google Scholar] [CrossRef]
- Tang, W.; Ding, Z.; Zhou, Z.Q.; Wang, Y.Z.; Guo, L.Y. Phylogenetic and pathogenic analyses show that the causal agent of apple ring rot in China is Botryosphaeria dothidea. Plant Dis. 2012, 96, 486–496. [Google Scholar] [CrossRef]
- Ma, Z.; Boehm, E.W.; Luo, Y.; Michailides, T.J. Population structure of Botryosphaeria dothidea from pistachio and other hosts in California. Phytopathology 2001, 91, 665–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehl, J.; Slippers, B.; Roux, J.; Wingfield, M.J. Cankers and other diseases caused by the Botryosphaeriaceae. Infect. For. Dis. 2013, 298–317. [Google Scholar]
- Marsberg, A.; Kemler, M.; Jami, F.; Nagel, J.H.; Postma-Smidt, A.; Naidoo, S.; Wingfield, M.J.; Crous, P.W.; Spatafora, J.W.; Hesse, C.N.; et al. Botryosphaeria dothidea: A latent pathogen of global importance to woody plant health. Mol. Plant Pathol. 2017, 18, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.; Di Lonardo, D.P.; Bodelier, P.L. Revisiting life strategy concepts in environmental microbial ecology. FEMS Microbiol. Ecol. 2017, 93, fix006. [Google Scholar] [CrossRef]
- Bochner, B.R. New technologies to assess genotype-phenotype relationships. Nat. Rev. Genet. 2003, 4, 309–314. [Google Scholar] [CrossRef]
- Bochner, B.R.; Gadzinski, P.; Panomitros, E. Phenotype microarrays for high-throughput phenotypic testing and assay of gene function. Genome Res. 2001, 11, 1246–1255. [Google Scholar] [CrossRef]
- Barbosa, C.; Mendes-Faia, A.; Mendes-Ferreira, A. The nitrogen source impacts major volatile compounds released by Saccharomyces cerevisiae during alcoholic fermentation. Int. J. Food Microbiol. 2012, 160, 87–93. [Google Scholar] [CrossRef]
- Shiraishi, K.; Oku, M.; Uchida, D.; Yurimoto, H.; Sakai, Y. Regulation of nitrate and methylamine metabolism by multiple nitrogen sources in the methylotrophic yeast Candida boidinii. Fems Yeast Res. 2015, 15, fov084. [Google Scholar] [CrossRef]
- Sanchez-Arreguin, J.A.; Hernandez-Onate, M.A.; Leon-Ramirez, C.G.; Ruiz-Herrera, J. Transcriptional analysis of the adaptation of Ustilago maydis during growth under nitrogen fixation conditions. J. Basic Microbiol. 2017, 57, 597–604. [Google Scholar] [CrossRef]
- Canas, P.; Morales, A.M.; Manso, J.; Romero, E.G.; Vinas, M.; Palomo, E.S. Chemical and sensory characrerization of the aroma of ‘chardonnay’ musts fermented with different nitrogen sources. Cienc Tec Vitivinic 2018, 33, 116–124. [Google Scholar]
- Da Silva, L.G.; Martins, M.P.; Sanches, P.R.; Peres, N.; Martinez-Rossi, N.M.; Rossi, A. Saline stress affects the pH-dependent regulation of the transcription factor PacC in the dermatophyte Trichophyton interdigitale. Braz. J. Microbiol. 2020, 51, 1585–1591. [Google Scholar] [CrossRef] [PubMed]
- Fedoseeva, E.V.; Tereshina, V.M.; Danilova, O.A.; Ianutsevich, E.A.; Yakimenko, O.S.; Terekhova, V.A. Effect of humic acid on the composition of osmolytes and lipids in a melanin-containing phytopathogenic fungus Alternaria alternata. Environ. Res. 2021, 193, 110395. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.C.; Guo, H.; Cai, L.; Cai, L.T.; Guo, Y.S.; Ding, W. Effect of temperature on phenotype characterization of Ralstonia solanacearum from tobacco. Can. J. Plant Pathol. 2020, 42, 164–181. [Google Scholar] [CrossRef]
- Wang, H.C.; Li, L.C.; Cai, B.; Cai, L.T.; Chen, X.J.; Yu, Z.H.; Zhang, C.Q. Metabolic phenotype characterization of Botrytis cinerea, the causal agent of gray mold. Front. Microbiol. 2018, 9, 470. [Google Scholar] [CrossRef]
- Gusarov, I.; Shatalin, K.; Starodubtseva, M.; Nudler, E. Endogenous nitric oxide protects bacteria against a wide spectrum of antibiotics. Science 2009, 325, 1380–1384. [Google Scholar] [CrossRef]
- Chen, X.; Li, L.; Wang, H.; Huang, Y.; Zhang, C. Phenotypic fingerprints of Ralstonia solanacearum under various osmolytes and pH environments. Plant Pathol. J. 2016, 15, 102–107. [Google Scholar] [CrossRef]
- Wang, H.; Huang, Y.; Xia, H.; Wang, J.; Wang, M.; Zhang, C.; Lu, H. Phenotypic Analysis of Alternaria alternata, the Causal Agent of Tobacco Brown Spot. Plant Pathol. J. 2015, 14, 79–85. [Google Scholar] [CrossRef]
- Bao, J.; Wu, Q.; Huang, J.; Zhang, C.Q. High-quality genome assembly and annotation resource of Botryosphaeria dothidea strain BDLA16−7, causing trunk canker disease on Chinese hickory. Plant Dis. 2022, 106, 1023–1026. [Google Scholar] [CrossRef]
- Xiao, X.E.; Wang, W.; Crous, P.W.; Wang, H.K.; Jiao, C.; Huang, F.; Pu, Z.X.; Zhu, Z.R.; Li, H.Y. Species of Botryosphaeriaceae associated with citrus branch diseases in China. Persoonia 2021, 47, 106–135. [Google Scholar] [CrossRef]
- Leigh, J.A.; Dodsworth, J.A. Nitrogen regulation in bacteria and archaea. Annu. Rev. Microbiol. 2007, 61, 349–377. [Google Scholar] [CrossRef]
- Shiraishi, K.; Oku, M.; Kawaguchi, K.; Uchida, D.; Yurimoto, H.; Sakai, Y. Yeast nitrogen utilization in the phyllosphere during plant lifespan under regulation of autophagy. Sci. Rep. 2015, 5, 9719. [Google Scholar] [CrossRef] [PubMed]
- Brefort, T.; Doehlemann, G.; Mendoza-Mendoza, A.; Reissmann, S.; Djamei, A.; Kahmann, R. Ustilago maydis as a pathogen. Annu. Rev. Phytopathol 2009, 47, 423–445. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, J.; Leon-Ramirez, C.; Vera-Nunez, A.; Sanchez-Arreguin, A.; Ruiz-Medrano, R.; Salgado-Lugo, H.; Sanchez-Segura, L.; Pena-Cabriales, J.J. A novel intracellular nitrogen-fixing symbiosis made by Ustilago maydis and Bacillus spp. New Phytol. 2015, 207, 769–777. [Google Scholar] [CrossRef]
- Shabala, S.; Shabala, L. Ion transport and osmotic adjustment in plants and bacteria. Biomol. Concepts 2011, 2, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Li, C.Z. Effects of cobalt and DFMA on polyamine content and membrane-lipid peroxidation in wheat seedling under osmotic stresses. Agric. Res. Arid. Areas 2010, 28, 136. [Google Scholar]
- Malla, C.F.; Mireles, N.A.; Ramirez, A.S.; Poveda, J.B.; Tavio, M.M. Aspirin, sodium benzoate and sodium salicylate reverse resistance to colistin in Enterobacteriaceae and Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2020, 75, 3568–3575. [Google Scholar] [CrossRef] [PubMed]
- Zhelifonova, V.P.; Antipova, T.V.; Kozlovskii, A.G. Effect of potassium sorbate, sodium benzoate, and sodium nitrite on biosynthesis of cyclopiazonic and mycophenolic acids and citrinin by fungi of the Penicillium genus. Appl. Biochem. Micro. 2017, 53, 711–714. [Google Scholar] [CrossRef]
- Yardimci, B.K.; Sahin, S.C.; Sever, N.I.; Ozek, N.S. Biochemical effects of sodium benzoate, potassium sorbate and sodium nitrite on food spoilage yeast Saccharomyces cerevisiae. Biologia 2022, 77, 547–557. [Google Scholar] [CrossRef]
- Dong, Y.L.; Ding, Z.F.; Song, L.X.; Zhang, D.S.; Xie, C.J.; Zhang, S.J.; Feng, L.; Liu, H.L.; Pang, Q.X. Sodium benzoate delays the development of Drosophila melanogaster larvae and alters commensal microbiota in adult Flies. Front. Microbiol. 2022, 13, 911928. [Google Scholar] [CrossRef]
- Jiang, L.; Pan, H. Functions of CaPhm7 in the regulation of ion homeostasis, drug tolerance, filamentation and virulence in Candida albicans. BMC Microbiol. 2018, 18, 49. [Google Scholar] [CrossRef]
- Medrano-Soto, A.; Moreno-Hagelsieb, G.; McLaughlin, D.; Ye, Z.S.; Hendargo, K.J.; Saier, M.J. Bioinformatic characterization of the anoctamin superfamily of Ca2+-activated ion channels and lipid scramblases. PLoS ONE 2018, 13, e0192851. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, T.R.; Segorbe, D.; Prusky, D.; Di Pietro, A. How alkalinization drives fungal pathogenicity. PLoS Pathog. 2017, 13, e1006621. [Google Scholar] [CrossRef] [PubMed]
- Cyert, M.S.; Philpott, C.C. Regulation of cation balance in Saccharomyces cerevisiae. Genetics 2013, 193, 677–713. [Google Scholar] [CrossRef]
- Tardi-Ovadia, R.; Linker, R.; Tsror, L.L. Direct estimation of local pH change at infection sites of fungi in potato tubers. Phytopathology 2017, 107, 132–137. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Li, G.; Jiang, D.; Chen, W. Sclerotinia sclerotiorum: An evaluation of virulence theories. Annu. Rev. Phytopathol 2018, 56, 311–338. [Google Scholar] [CrossRef]
- Davis, D.A. How human pathogenic fungi sense and adapt to pH: The link to virulence. Curr. Opin. Microbiol. 2009, 12, 365–370. [Google Scholar] [CrossRef]
- Davis, D.; Edwards, J.J.; Mitchell, A.P.; Ibrahim, A.S. Candida albicans RIM101 pH response pathway is required for host-pathogen interactions. Infect Immun. 2000, 68, 5953–5959. [Google Scholar] [CrossRef] [PubMed]
- Ortoneda, M.; Guarro, J.; Madrid, M.P.; Caracuel, Z.; Roncero, M.I.; Mayayo, E.; Di Pietro, A. Fusarium oxysporum as a multihost model for the genetic dissection of fungal virulence in plants and mammals. Infect Immun. 2004, 72, 1760–1766. [Google Scholar] [CrossRef]
- Bertuzzi, M.; Schrettl, M.; Alcazar-Fuoli, L.; Cairns, T.C.; Munoz, A.; Walker, L.A.; Herbst, S.; Safari, M.; Cheverton, A.M.; Chen, D.; et al. Correction: The pH-Responsive PacC transcription factor of Aspergillus fumigatus governs epithelial entry and tissue invasion during pulmonary aspergillosis. PLoS Pathog. 2015, 11, e1004943. [Google Scholar] [CrossRef]
- Fan, X.; Zhang, J.; Yang, L.; Wu, M.D.; Chen, W.D.; Li, G.Q. Development of PCR-based assays for detecting and differentiating three species of Botrytis infecting broad bean. Plant Dis. 2015, 99, 691–698. [Google Scholar] [CrossRef]
- Ni, H.F.; Yang, H.R.; Chen, R.S.; Hung, T.H.; Liou, R.F. A nested multiplex PCR for species-specific identification and detection of Botryosphaeriaceae species on mango. Eur. J. Plant Pathol. 2012, 133, 819–828. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Amino Acid Nitrogen Substrates (%) | Biosynthetic Pathways (%) | Osmotic and Ionic Conditions (%) | pH Condition (%) |
|---|---|---|---|---|
| B. dothidea (BDLA 16−7) | 96.8 | 100 | 93.8 | 89.6 |
| B. fabicerciana (BFLG 18−2) | 89.5 | 100 | 93.8 | 68.8 |
| B. qingyuanensis (BQTK 16−30) | 88.4 | 98.9 | 93.8 | 89.6 |
| B. corticis (BCTK 16−35) | 86.3 | 100 | 93.8 | 68.8 |
| Gene Name | KEGG Ortholog(KO) | Thrshld | Score | E-Value | KO Definition |
|---|---|---|---|---|---|
| jg9527 | K23322 | 48.03 | 74.2 | 6.8 × 10−23 | aspirochlorine biosynthesis cytochrome P450 monooxygenase |
| jg3332 | K22993 | 508.27 | 561.6 | 6.3 × 10−171 | bifunctional Delta-12/omega-3 fatty acid desaturase |
| jg10959 | K23504 | 91.30 | 266.6 | 2.9 × 10−81 | protein SERAC1 |
| jg10977 | K21989 | 150.87 | 173.2 | 5.8 × 10−53 | calcium permeable stress-gated cation channel |
| g8479 | K01183 | 118.77 | 159.1 | 1.4 × 10−48 | chitinase |
| jg3335 | K01687 | 579.77 | 924.5 | 4 × 10−280 | dihydroxy-acid dehydratase |
| jg3334 | K06867 | 98.20 | 108.3 | 4.4 × 10−33 | uncharacterized protein |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, T.; Zhang, Y.; Yan, C.; Zhang, C. Phenotypic and Genomic Difference among Four Botryosphaeria Pathogens in Chinese Hickory Trunk Canker. J. Fungi 2023, 9, 204. https://doi.org/10.3390/jof9020204
Ma T, Zhang Y, Yan C, Zhang C. Phenotypic and Genomic Difference among Four Botryosphaeria Pathogens in Chinese Hickory Trunk Canker. Journal of Fungi. 2023; 9(2):204. https://doi.org/10.3390/jof9020204
Chicago/Turabian StyleMa, Tianling, Yu Zhang, Chenyi Yan, and Chuanqing Zhang. 2023. "Phenotypic and Genomic Difference among Four Botryosphaeria Pathogens in Chinese Hickory Trunk Canker" Journal of Fungi 9, no. 2: 204. https://doi.org/10.3390/jof9020204