
Functional Analysis of the P-Type ATPases Apt2-4 from Cryptococcus neoformans by Heterologous Expression in Saccharomyces cerevisiae


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Plasmid Construction
2.3. Yeast Strains and Media
2.4. Functional Complementation Assays
2.5. Membrane Preparation and Gradient Fractionation
2.6. Co-Immunoprecipitation
2.7. NBD-lipid Uptake and Flow Cytometry
2.8. Sequence Alignment and Similarity Analysis
3. Results
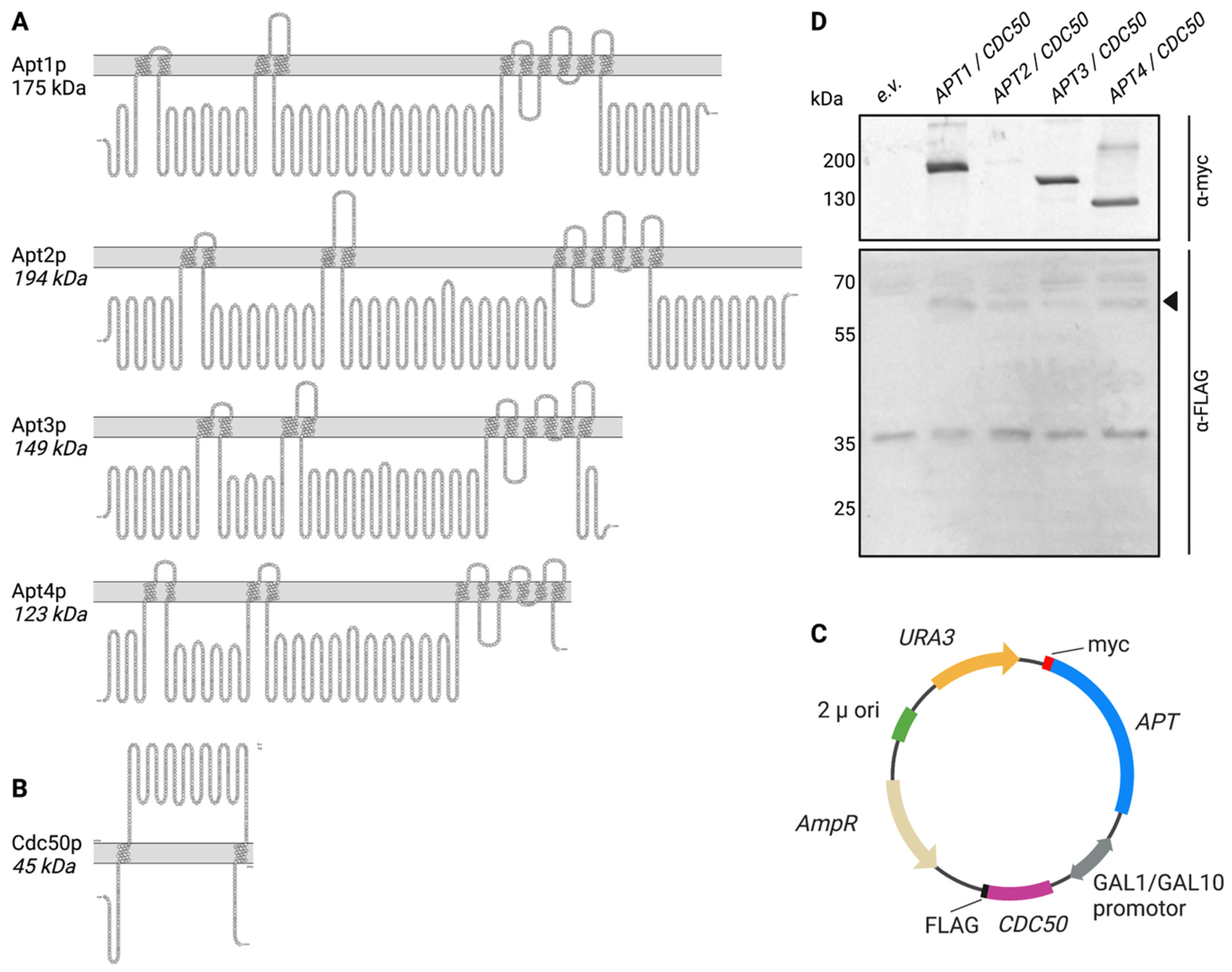
3.1. Expression of Heterologous APT1-4 and CDC50 in S. cerevisiae
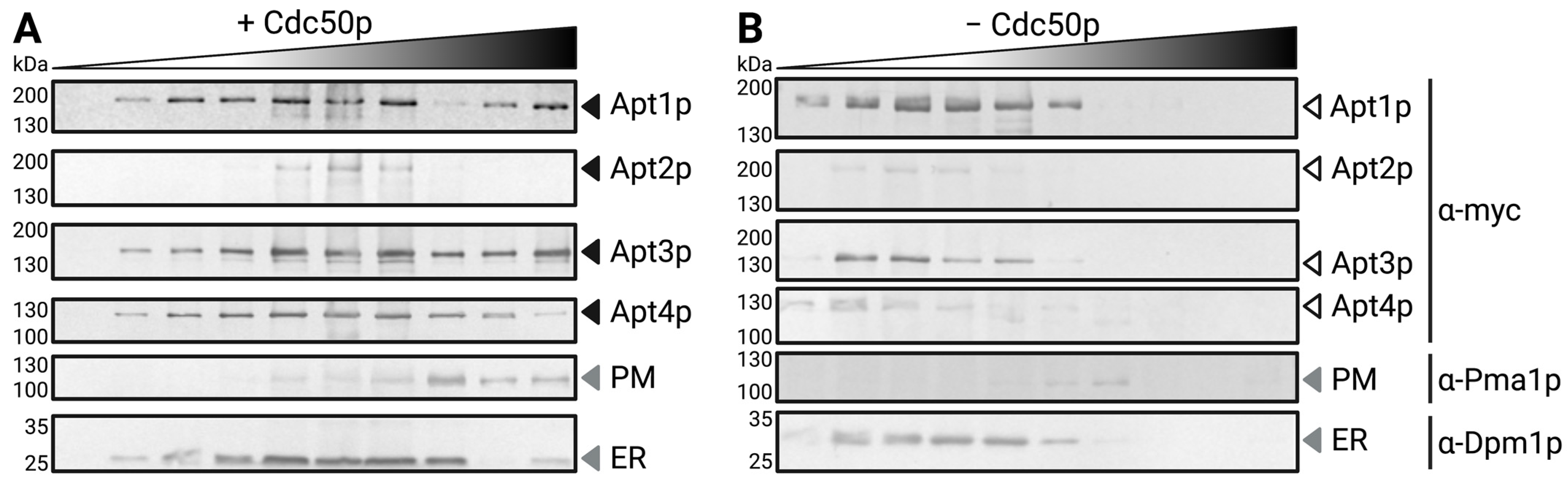
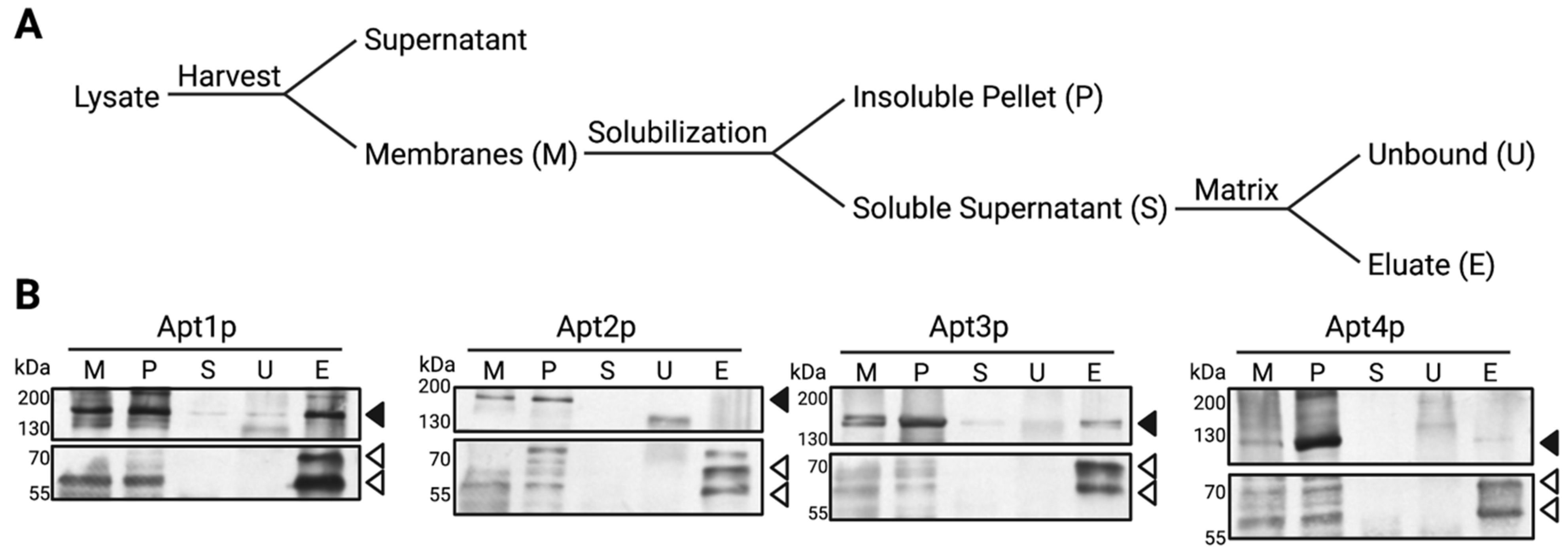
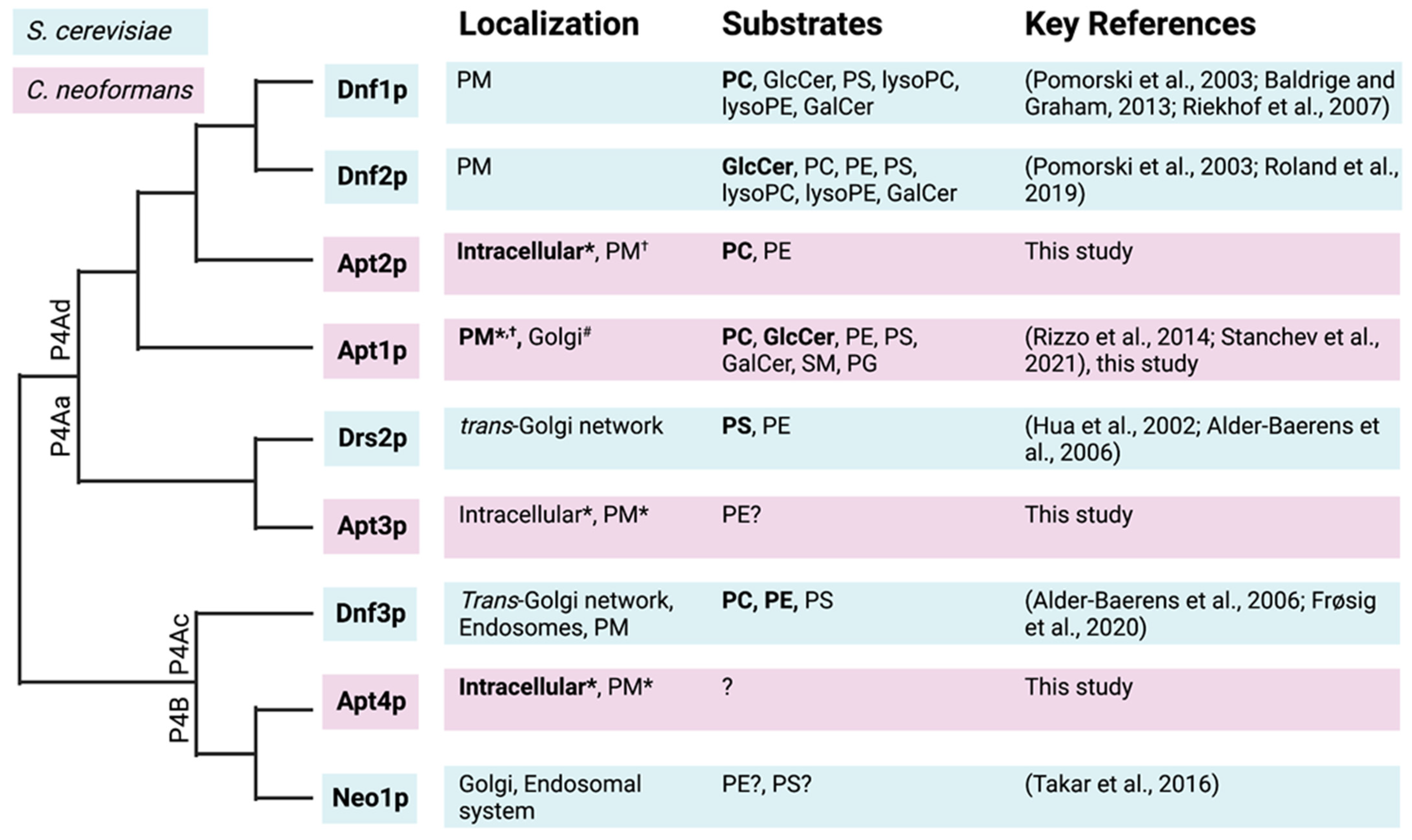
3.2. Subcellular Localization of Heterologous Apt1-4p and Interaction with Cdc50p
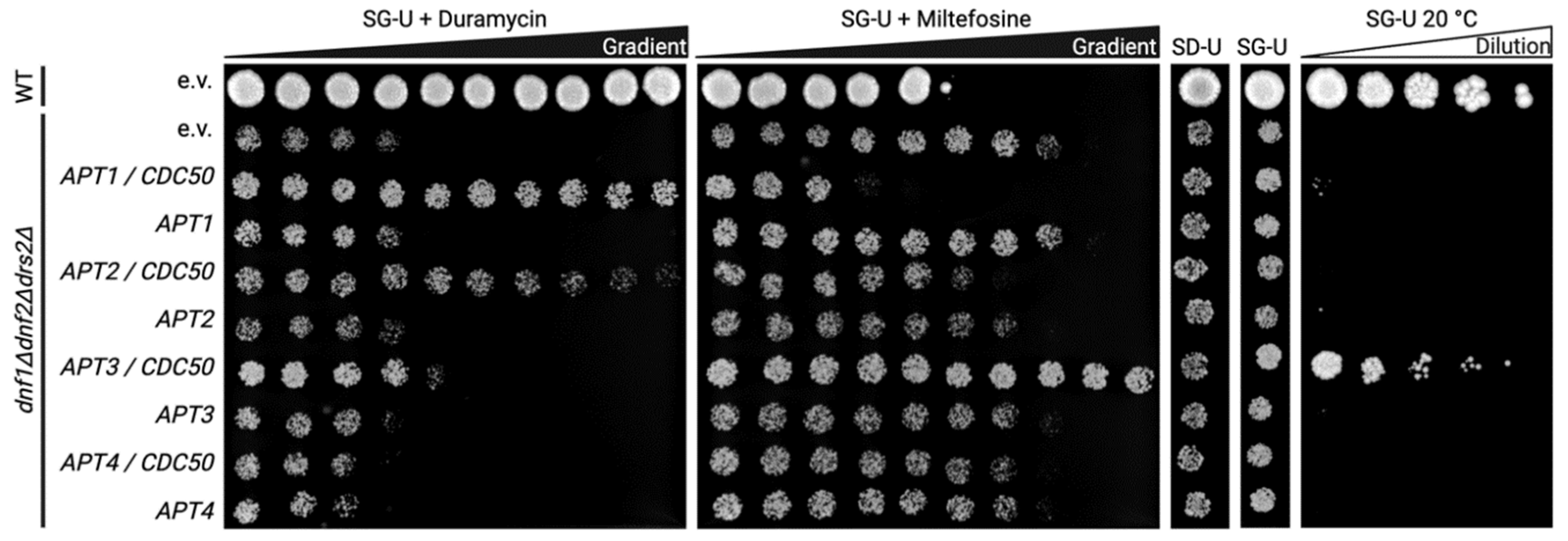
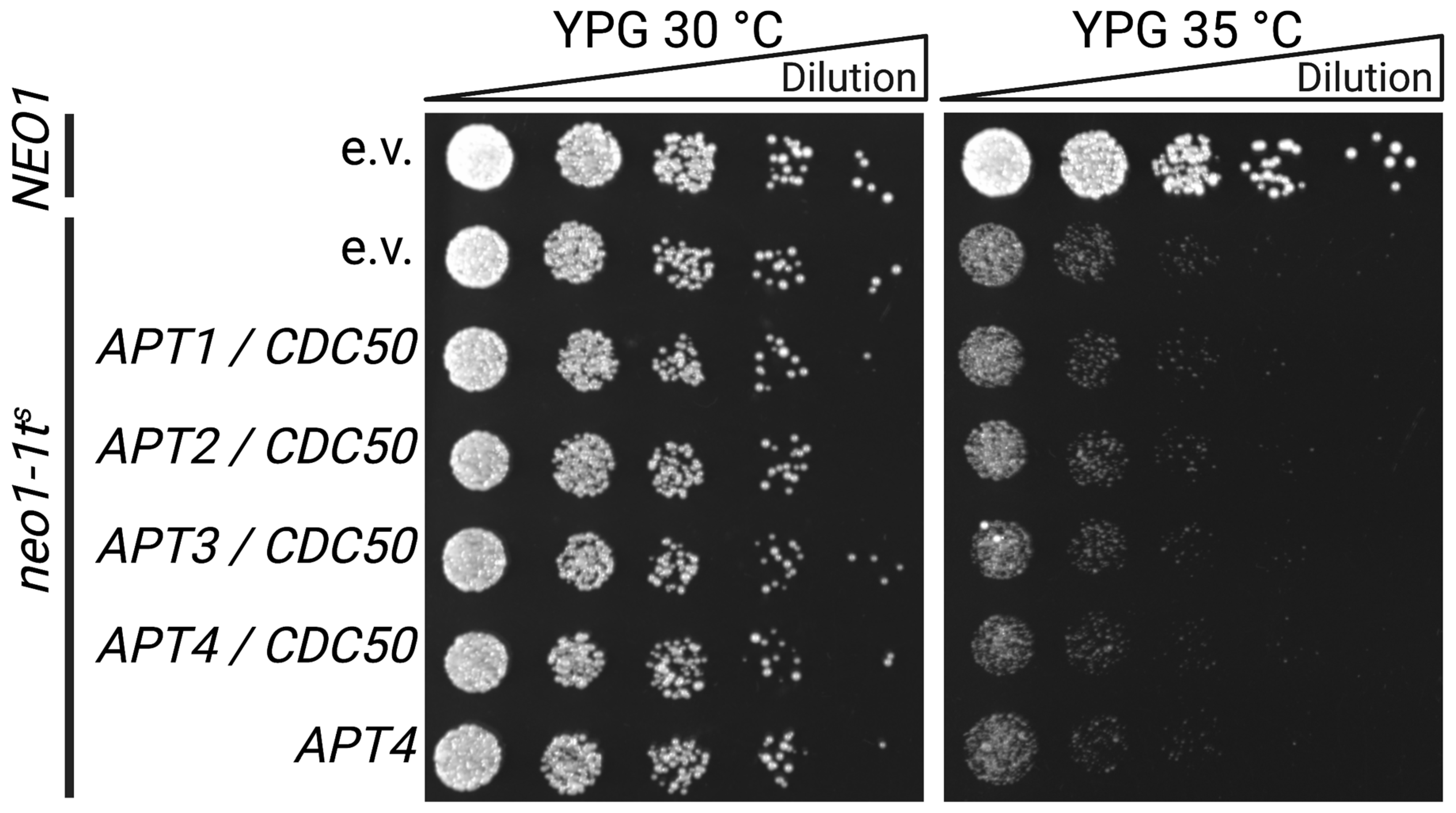
3.3. Functional Status of Heterologously Expressed Apt1-4p in S. cerevisiae
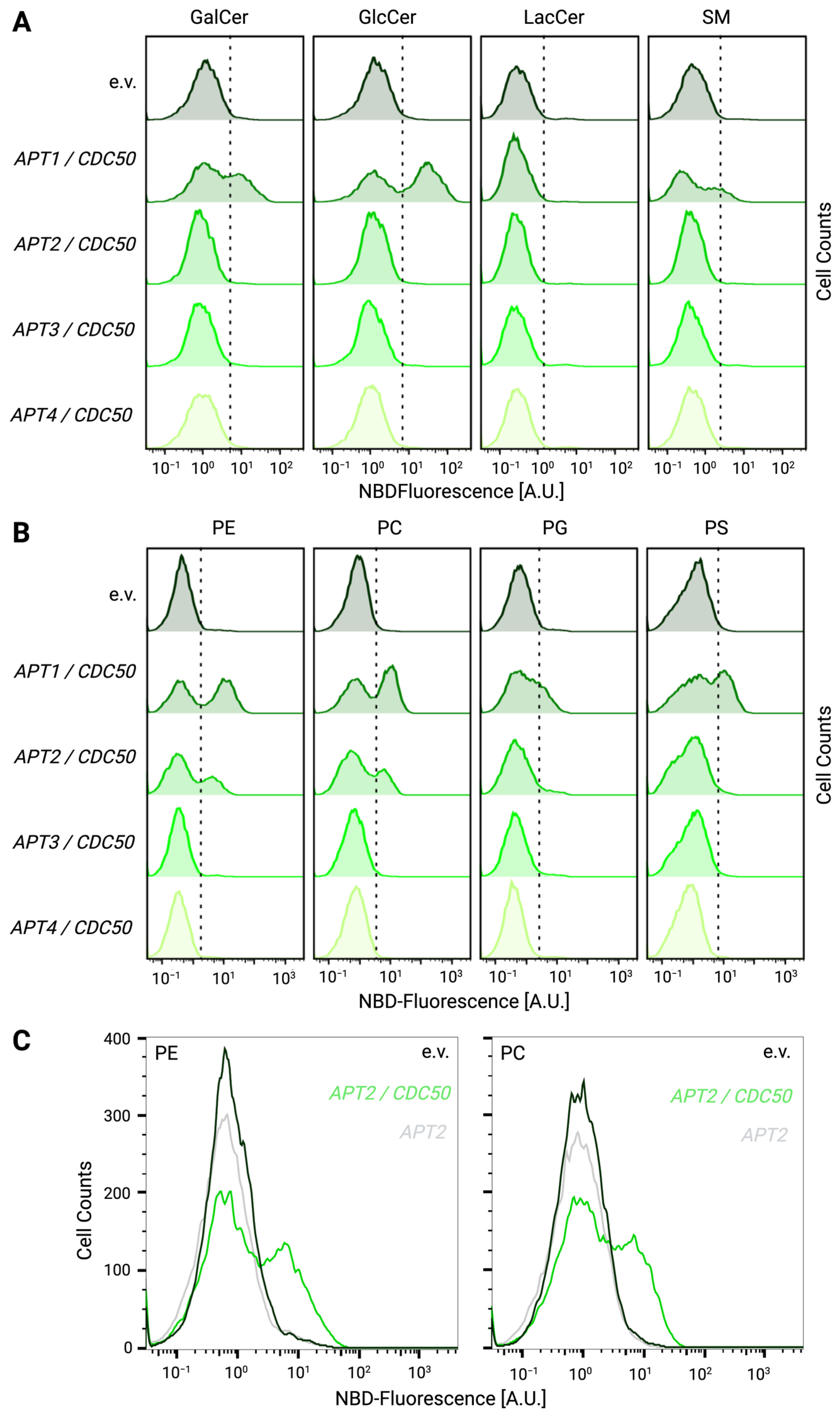
3.4. Heterologously Expressed Apt1p and Apt2p Support Lipid Flippase Activity
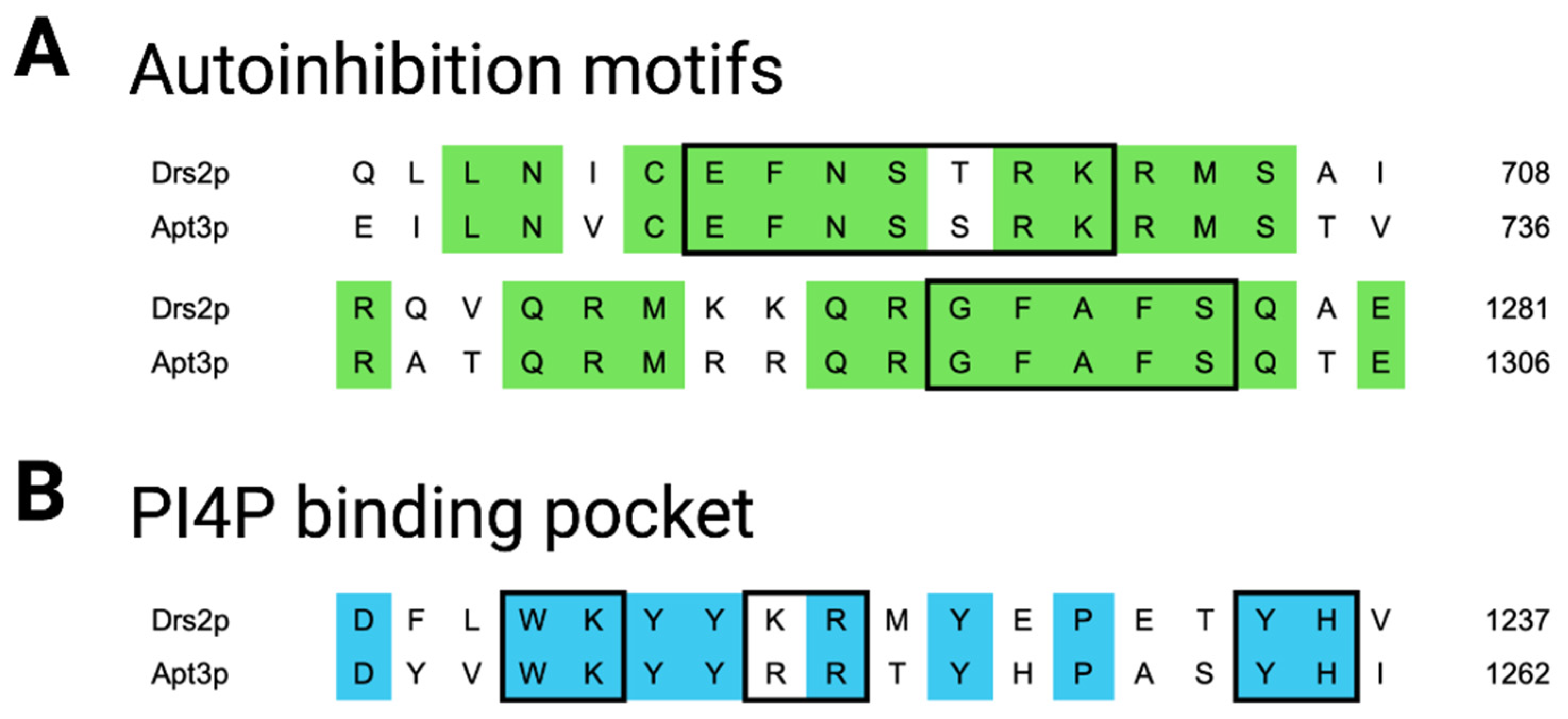
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global Burden of Disease of HIV-Associated Cryptococcal Meningitis: An Updated Analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef]
- Maziarz, E.K.; Perfect, J.R. Cryptococcosis. Infect. Dis. Clin. North Am. 2016, 30, 179–206. [Google Scholar] [CrossRef]
- World Health Organisation. WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action; Licence: CC BY-NC-SA 3.0 IGO; World Health Organisation: Geneva, Switzerland, 2022. [Google Scholar]
- Bongomin, F.; Oladele, R.O.; Gago, S.; Moore, C.B.; Richardson, M.D. A Systematic Review of Fluconazole Resistance in Clinical Isolates of Cryptococcus Species. Mycoses 2018, 61, 290–297. [Google Scholar] [CrossRef]
- Spadari, C.d.C.; Wirth, F.; Lopes, L.B.; Ishida, K. New Approaches for Cryptococcosis Treatment. Microorganisms 2020, 8, 613. [Google Scholar] [CrossRef]
- McClelland, E.E.; Bernhardt, P.; Casadevall, A. Estimating the Relative Contributions of Virulence Factors for Pathogenic Microbes. Infect Immun. 2006, 74, 1500–1504. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.P.; Vestergaard, A.L.; Mikkelsen, S.A.; Mogensen, L.S.; Chalat, M.; Molday, R.S. P4-ATPases as Phospholipid Flippases—Structure, Function, and Enigmas. Front. Physiol. 2016, 7, 275. [Google Scholar] [CrossRef]
- Hanson, P.K.; Malone, L.; Birchmore, J.L.; Nichols, J.W. Lem3p Is Essential for the Uptake and Potency of Alkylphosphocholine Drugs, Edelfosine and Miltefosine*. J. Biol. Chem. 2003, 278, 36041–36050. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Liao, G.; Baker, G.M.; Wang, Y.; Lau, R.; Paderu, P.; Perlin, D.S.; Xue, C. Lipid Flippase Subunit Cdc50 Mediates Drug Resistance and Virulence in Cryptococcus neoformans. mBio 2016, 7, e00478-16. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, X.; Zhang, B.; Zeng, X.; Mao, H.; Xu, H.; Jiang, L.; Li, F. The Lipid Flippase Subunit Cdc50 Is Required for Antifungal Drug Resistance, Endocytosis, Hyphal Development and Virulence in Candida Albicans. FEMS Yeast Res. 2019, 19, foz033. [Google Scholar] [CrossRef] [PubMed]
- Cowell, A.N.; Istvan, E.S.; Lukens, A.K.; Gomez-Lorenzo, M.G.; Vanaerschot, M.; Sakata-Kato, T.; Flannery, E.L.; Magistrado, P.; Owen, E.; Abraham, M.; et al. Mapping the Malaria Parasite Druggable Genome by Using in Vitro Evolution and Chemogenomics. Science 2018, 359, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Azouaoui, H.; Montigny, C.; Dieudonné, T.; Champeil, P.; Jacquot, A.; Vázquez-Ibar, J.L.; Maréchal, P.L.; Ulstrup, J.; Ash, M.-R.; Lyons, J.A.; et al. High Phosphatidylinositol 4-Phosphate (PI4P)-Dependent ATPase Activity for the Drs2p-Cdc50p Flippase after Removal of Its N- and C-Terminal Extensions. J. Biol. Chem. 2017, 292, 7954–7970. [Google Scholar] [CrossRef]
- Timcenko, M.; Lyons, J.A.; Januliene, D.; Ulstrup, J.J.; Dieudonné, T.; Montigny, C.; Ash, M.-R.; Karlsen, J.L.; Boesen, T.; Kühlbrandt, W.; et al. Structure and Autoregulation of a P4-ATPase Lipid Flippase. Nature 2019, 571, 366–370. [Google Scholar] [CrossRef]
- Hu, G.; Kronstad, J.W. A Putative P-Type ATPase, Apt1, Is Involved in Stress Tolerance and Virulence in Cryptococcus neoformans. Eukaryot Cell 2010, 9, 74–83. [Google Scholar] [CrossRef]
- Rizzo, J.; Oliveira, D.L.; Joffe, L.S.; Hu, G.; Gazos-Lopes, F.; Fonseca, F.L.; Almeida, I.C.; Frases, S.; Kronstad, J.W.; Rodrigues, M.L. Role of the Apt1 Protein in Polysaccharide Secretion by Cryptococcus neoformans. Eukaryot Cell 2014, 13, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, J.; Colombo, A.C.; Zamith-Miranda, D.; Silva, V.K.A.; Allegood, J.C.; Casadevall, A.; Del Poeta, M.; Nosanchuk, J.D.; Kronstad, J.W.; Rodrigues, M.L. The Putative Flippase Apt1 Is Required for Intracellular Membrane Architecture and Biosynthesis of Polysaccharide and Lipids in Cryptococcus neoformans. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Caza, M.; Bakkeren, E.; Kretschmer, M.; Bairwa, G.; Reiner, E.; Kronstad, J. A P4-ATPase Subunit of the Cdc50 Family Plays a Role in Iron Acquisition and Virulence in Cryptococcus neoformans. Cell Microbiol 2017, 19, e12718. [Google Scholar] [CrossRef] [PubMed]
- Graham, T.R. Flippases and Vesicle-Mediated Protein Transport. Trends Cell Biol. 2004, 14, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Stanchev, L.D.; Rizzo, J.; Peschel, R.; Pazurek, L.A.; Bredegaard, L.; Veit, S.; Laerbusch, S.; Rodrigues, M.L.; López-Marqués, R.L.; Günther Pomorski, T. P-Type ATPase Apt1 of the Fungal Pathogen Cryptococcus neoformans Is a Lipid Flippase of Broad Substrate Specificity. J. Fungi 2021, 7, 843. [Google Scholar] [CrossRef] [PubMed]
- Bryde, S.; Hennrich, H.; Verhulst, P.M.; Devaux, P.F.; Lenoir, G.; Holthuis, J.C.M. CDC50 Proteins Are Critical Components of the Human Class-1 P4-ATPase Transport Machinery*. J. Biol. Chem. 2010, 285, 40562–40572. [Google Scholar] [CrossRef] [PubMed]
- Lenoir, G.; Williamson, P.; Puts, C.F.; Holthuis, J.C.M. Cdc50p Plays a Vital Role in the ATPase Reaction Cycle of the Putative Aminophospholipid Transporter Drs2p*♦. J. Biol. Chem. 2009, 284, 17956–17967. [Google Scholar] [CrossRef] [Green Version]
- Segawa, K.; Kurata, S.; Nagata, S. The CDC50A Extracellular Domain Is Required for Forming a Functional Complex with and Chaperoning Phospholipid Flippases to the Plasma Membrane. J. Biol. Chem. 2018, 293, 2172–2182. [Google Scholar] [CrossRef]
- Sebastian, T.T.; Baldridge, R.D.; Xu, P.; Graham, T.R. Phospholipid Flippases: Building Asymmetric Membranes and Transport Vesicles. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2012, 1821, 1068–1077. [Google Scholar] [CrossRef]
- Tancer, R.J.; Wang, Y.; Pawar, S.; Xue, C.; Wiedman, G.R. Development of Antifungal Peptides against Cryptococcus neoformans; Leveraging Knowledge about the Cdc50Δ Mutant Susceptibility for Lead Compound Development. Microbiol. Spectr. 2022, 10, e0043922. [Google Scholar] [CrossRef]
- Takar, M.; Wu, Y.; Graham, T.R. The Essential Neo1 Protein from Budding Yeast Plays a Role in Establishing Aminophospholipid Asymmetry of the Plasma Membrane. J. Biol. Chem. 2016, 291, 15727–15739. [Google Scholar] [CrossRef]
- Hua, Z.; Fatheddin, P.; Graham, T.R. An Essential Subfamily of Drs2p-Related P-Type ATPases Is Required for Protein Trafficking between Golgi Complex and Endosomal/Vacuolar System. MBoC 2002, 13, 3162–3177. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. High-Efficiency Yeast Transformation Using the LiAc/SS Carrier DNA/PEG Method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.S.; Costa, S.; Günther-Pomorski, T.; López-Marqués, R.L. Cell-Based Lipid Flippase Assay Employing Fluorescent Lipid Derivatives. In P-Type ATPases: Methods and Protocols; Bublitz, M., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2016; pp. 371–382. ISBN 978-1-4939-3179-8. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and Sequence Analysis Tools Services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology Modelling of Protein Structures and Complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnhammer, E.L.L.; Eddy, S.R.; Birney, E.; Bateman, A.; Durbin, R. Pfam: Multiple Sequence Alignments and HMM-Profiles of Protein Domains. Nucleic Acids Res. 1998, 26, 320–322. [Google Scholar] [CrossRef]
- Omasits, U.; Ahrens, C.H.; Müller, S.; Wollscheid, B. Protter: Interactive Protein Feature Visualization and Integration with Experimental Proteomic Data. Bioinformatics 2014, 30, 884–886. [Google Scholar] [CrossRef]
- Gupta, R.; Brunak, S. Prediction of Glycosylation across the Human Proteome and the Correlation to Protein Function. Pac. Symp. Biocomput. 2002, 7, 310–322. [Google Scholar]
- Jain, B.K.; Wagner, A.S.; Reynolds, T.B.; Graham, T.R. Lipid Transport by Candida albicans Dnf2 Is Required for Hyphal Growth and Virulence. Infect Immun. 2022, 90, e0041622. [Google Scholar] [CrossRef]
- Palmgren, M.G.; Nissen, P. P-Type ATPases. Annu. Rev. Biophys. 2011, 40, 243–266. [Google Scholar] [CrossRef]
- Bai, L.; Jain, B.K.; You, Q.; Duan, H.D.; Takar, M.; Graham, T.R.; Li, H. Structural Basis of the P4B ATPase Lipid Flippase Activity. Nat. Commun. 2021, 12, 5963. [Google Scholar] [CrossRef]
- Coleman, J.A.; Vestergaard, A.L.; Molday, R.S.; Vilsen, B.; Peter Andersen, J. Critical Role of a Transmembrane Lysine in Aminophospholipid Transport by Mammalian Photoreceptor P4-ATPase ATP8A2. Proc. Natl. Acad. Sci. USA 2012, 109, 1449–1454. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, K.; Hayakawa, T.; Murate, M.; Makino, A.; Ito, K.; Fujisawa, T.; Kobayashi, T. Curvature-Dependent Recognition of Ethanolamine Phospholipids by Duramycin and Cinnamycin. Biophys. J. 2007, 93, 1608–1619. [Google Scholar] [CrossRef]
- Rzeźnicka, I.I.; Sovago, M.; Backus, E.H.G.; Bonn, M.; Yamada, T.; Kobayashi, T.; Kawai, M. Duramycin-Induced Destabilization of a Phosphatidylethanolamine Monolayer at the Air−Water Interface Observed by Vibrational Sum-Frequency Generation Spectroscopy. Langmuir 2010, 26, 16055–16062. [Google Scholar] [CrossRef] [PubMed]
- Romanos, M.A.; Scorer, C.A.; Clare, J.J. Foreign Gene Expression in Yeast: A Review. Yeast 1992, 8, 423–488. [Google Scholar] [CrossRef]
- Bai, L.; You, Q.; Jain, B.K.; Duan, H.D.; Kovach, A.; Graham, T.R.; Li, H. Transport Mechanism of P4 ATPase Phosphatidylcholine Flippases. Elife 2020, 9, e62163. [Google Scholar] [CrossRef]
- Bai, L.; Kovach, A.; You, Q.; Hsu, H.-C.; Zhao, G.; Li, H. Autoinhibition and Activation Mechanisms of the Eukaryotic Lipid Flippase Drs2p-Cdc50p. Nat. Commun. 2019, 10, 4142. [Google Scholar] [CrossRef]
- Cheng, M.-T.; Chen, Y.; Chen, Z.-P.; Liu, X.; Zhang, Z.; Chen, Y.; Hou, W.-T.; Zhou, C.-Z. Structural Insights into the Activation of Autoinhibited Human Lipid Flippase ATP8B1 upon Substrate Binding. Proc. Natl. Acad. Sci. USA 2022, 119, e2118656119. [Google Scholar] [CrossRef] [PubMed]
- Dieudonné, T.; Herrera, S.A.; Laursen, M.J.; Lejeune, M.; Stock, C.; Slimani, K.; Jaxel, C.; Lyons, J.A.; Montigny, C.; Pomorski, T.G.; et al. Autoinhibition and Regulation by Phosphoinositides of ATP8B1, a Human Lipid Flippase Associated with Intrahepatic Cholestatic Disorders. Elife 2022, 11, e75272. [Google Scholar] [CrossRef]
- He, Y.; Xu, J.; Wu, X.; Li, L. Structures of a P4-ATPase Lipid Flippase in Lipid Bilayers. Protein Cell 2020, 11, 458–463. [Google Scholar] [CrossRef]
- Xu, J.; He, Y.; Wu, X.; Li, L. Conformational Changes of a Phosphatidylcholine Flippase in Lipid Membranes. Cell Rep. 2022, 38, 110518. [Google Scholar] [CrossRef] [PubMed]
- Iyoshi, S.; Cheng, J.; Tatematsu, T.; Takatori, S.; Taki, M.; Yamamoto, Y.; Salic, A.; Fujimoto, T. Asymmetrical Distribution of Choline Phospholipids Revealed by Click Chemistry and Freeze-Fracture Electron Microscopy. ACS Chem. Biol. 2014, 9, 2217–2222. [Google Scholar] [CrossRef]
- Rawyler, A.; Van der Schaft, P.H.; Roelofsen, B.; Op den Kamp, J.A.F. Phospholipid Localization in the Plasma Membrane of Friend Erythroleukemic Cells and Mouse Erythrocytes. Biochemistry 1985, 24, 1777–1783. [Google Scholar] [CrossRef] [PubMed]
- Palmgren, M.; Østerberg, J.T.; Nintemann, S.J.; Poulsen, L.R.; López-Marqués, R.L. Evolution and a Revised Nomenclature of P4 ATPases, a Eukaryotic Family of Lipid Flippases. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2019, 1861, 1135–1151. [Google Scholar] [CrossRef]
- Pomorski, T.; Lombardi, R.; Riezman, H.; Devaux, P.F.; van Meer, G.; Holthuis, J.C.M. Drs2p-Related P-Type ATPases Dnf1p and Dnf2p Are Required for Phospholipid Translocation across the Yeast Plasma Membrane and Serve a Role in Endocytosis. Mol. Biol. Cell 2003, 14, 1240–1254. [Google Scholar] [CrossRef]
- Baldridge, R.D.; Graham, T.R. Two-Gate Mechanism for Phospholipid Selection and Transport by Type IV P-Type ATPases. Proc. Natl. Acad. Sci. USA 2013, 110, E358–E367. [Google Scholar] [CrossRef] [Green Version]
- Riekhof, W.R.; Wu, J.; Gijón, M.A.; Zarini, S.; Murphy, R.C.; Voelker, D.R. Lysophosphatidylcholine Metabolism in Saccharomyces cerevisiae: The Role of P-Type ATPases in Transport and a Broad Specificity Acyltransferase in Acylation. J. Biol. Chem. 2007, 282, 36853–36861. [Google Scholar] [CrossRef]
- Roland, B.P.; Naito, T.; Best, J.T.; Arnaiz-Yépez, C.; Takatsu, H.; Yu, R.J.; Shin, H.-W.; Graham, T.R. Yeast and Human P4-ATPases Transport Glycosphingolipids Using Conserved Structural Motifs. J. Biol. Chem. 2019, 294, 1794–1806. [Google Scholar] [CrossRef]
- Alder-Baerens, N.; Lisman, Q.; Luong, L.; Pomorski, T.; Holthuis, J.C.M. Loss of P4 ATPases Drs2p and Dnf3p Disrupts Aminophospholipid Transport and Asymmetry in Yeast Post-Golgi Secretory Vesicles. Mol. Biol. Cell 2006, 17, 1632–1642. [Google Scholar] [CrossRef]
- Frøsig, M.M.; Costa, S.R.; Liesche, J.; Østerberg, J.T.; Hanisch, S.; Nintemann, S.; Sørensen, H.; Palmgren, M.; Pomorski, T.G.; López-Marqués, R.L. Pseudohyphal Growth in Saccharomyces Cerevisiae Involves Protein Kinase-Regulated Lipid Flippases. J. Cell Sci. 2020, 133, jcs235994. [Google Scholar] [CrossRef] [PubMed]
- Baldridge, R.D.; Xu, P.; Graham, T.R. Type IV P-Type ATPases Distinguish Mono- versus Diacyl Phosphatidylserine Using a Cytofacial Exit Gate in the Membrane Domain. J. Biol. Chem. 2013, 288, 19516–19527. [Google Scholar] [CrossRef] [PubMed]
- Lamy, A.; Macarini-Bruzaferro, E.; Dieudonné, T.; Perálvarez-Marín, A.; Lenoir, G.; Montigny, C.; Le Maire, M.; Vázquez-Ibar, J.L. ATP2, The Essential P4-ATPase of Malaria Parasites, Catalyzes Lipid-Stimulated ATP Hydrolysis in Complex with a Cdc50 β-Subunit. Emerg. Microbes Infect. 2021, 10, 132–147. [Google Scholar] [CrossRef]
- Natarajan, P.; Liu, K.; Patil, D.V.; Sciorra, V.A.; Jackson, C.L.; Graham, T.R. Regulation of a Golgi Flippase by Phosphoinositides and an ArfGEF. Nat. Cell Biol. 2009, 11, 1421–1426. [Google Scholar] [CrossRef]
- Zhou, X.; Sebastian, T.T.; Graham, T.R. Auto-Inhibition of Drs2p, a Yeast Phospholipid Flippase, by Its Carboxyl-Terminal Tail*. J. Biol. Chem. 2013, 288, 31807–31815. [Google Scholar] [CrossRef] [PubMed]
- Takatsu, H.; Baba, K.; Shima, T.; Umino, H.; Kato, U.; Umeda, M.; Nakayama, K.; Shin, H.-W. ATP9B, a P4-ATPase (a Putative Aminophospholipid Translocase), Localizes to the Trans-Golgi Network in a CDC50 Protein-Independent Manner. J. Biol. Chem. 2011, 286, 38159–38167. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Fujimura-Kamada, K.; Furuta, N.; Kato, U.; Umeda, M.; Tanaka, K. Cdc50p, a Protein Required for Polarized Growth, Associates with the Drs2p P-Type ATPase Implicated in Phospholipid Translocation in Saccharomyces cerevisiae. Mol. Biol. Cell 2004, 15, 3418–3432. [Google Scholar] [CrossRef] [Green Version]
- Wicky, S.; Schwarz, H.; Singer-Krüger, B. Molecular Interactions of Yeast Neo1p, an Essential Member of the Drs2 Family of Aminophospholipid Translocases, and Its Role in Membrane Trafficking within the Endomembrane System. Mol. Cell. Biol. 2004, 24, 7402–7418. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veit, S.; Laerbusch, S.; López-Marqués, R.L.; Günther Pomorski, T. Functional Analysis of the P-Type ATPases Apt2-4 from Cryptococcus neoformans by Heterologous Expression in Saccharomyces cerevisiae. J. Fungi 2023, 9, 202. https://doi.org/10.3390/jof9020202
Veit S, Laerbusch S, López-Marqués RL, Günther Pomorski T. Functional Analysis of the P-Type ATPases Apt2-4 from Cryptococcus neoformans by Heterologous Expression in Saccharomyces cerevisiae. Journal of Fungi. 2023; 9(2):202. https://doi.org/10.3390/jof9020202
Chicago/Turabian StyleVeit, Sarina, Sabine Laerbusch, Rosa L. López-Marqués, and Thomas Günther Pomorski. 2023. "Functional Analysis of the P-Type ATPases Apt2-4 from Cryptococcus neoformans by Heterologous Expression in Saccharomyces cerevisiae" Journal of Fungi 9, no. 2: 202. https://doi.org/10.3390/jof9020202