
Survival Factor A (SvfA) Contributes to Aspergillus nidulans Pathogenicity
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Generation of Fungal Strains
2.2. Biofilm Formation Assay
2.3. Protease Activity
2.4. RNA Preparation, cDNA Synthesis, and Quantitative Real-Time PCR
2.5. Phagocytosis and Macrophage Killing Assay
2.6. Microscopy
2.7. Polysaccharide Analysis
2.8. Preparation of Protein Extracts and Immunoblot Analysis
2.9. Cytokine Measurements
2.10. Zebrafish Infection Assay by Conidial Microinjection
2.11. Mouse Strains
2.12. Fungal Aspiration and Lung Harvest
2.13. Fungal Burden Assay
2.14. RNA Extraction from Mice Lungs for Cytokine Analysis
2.15. Flow Cytometric Analysis of BAL Cells
2.16. Statistical Analysis
3. Results
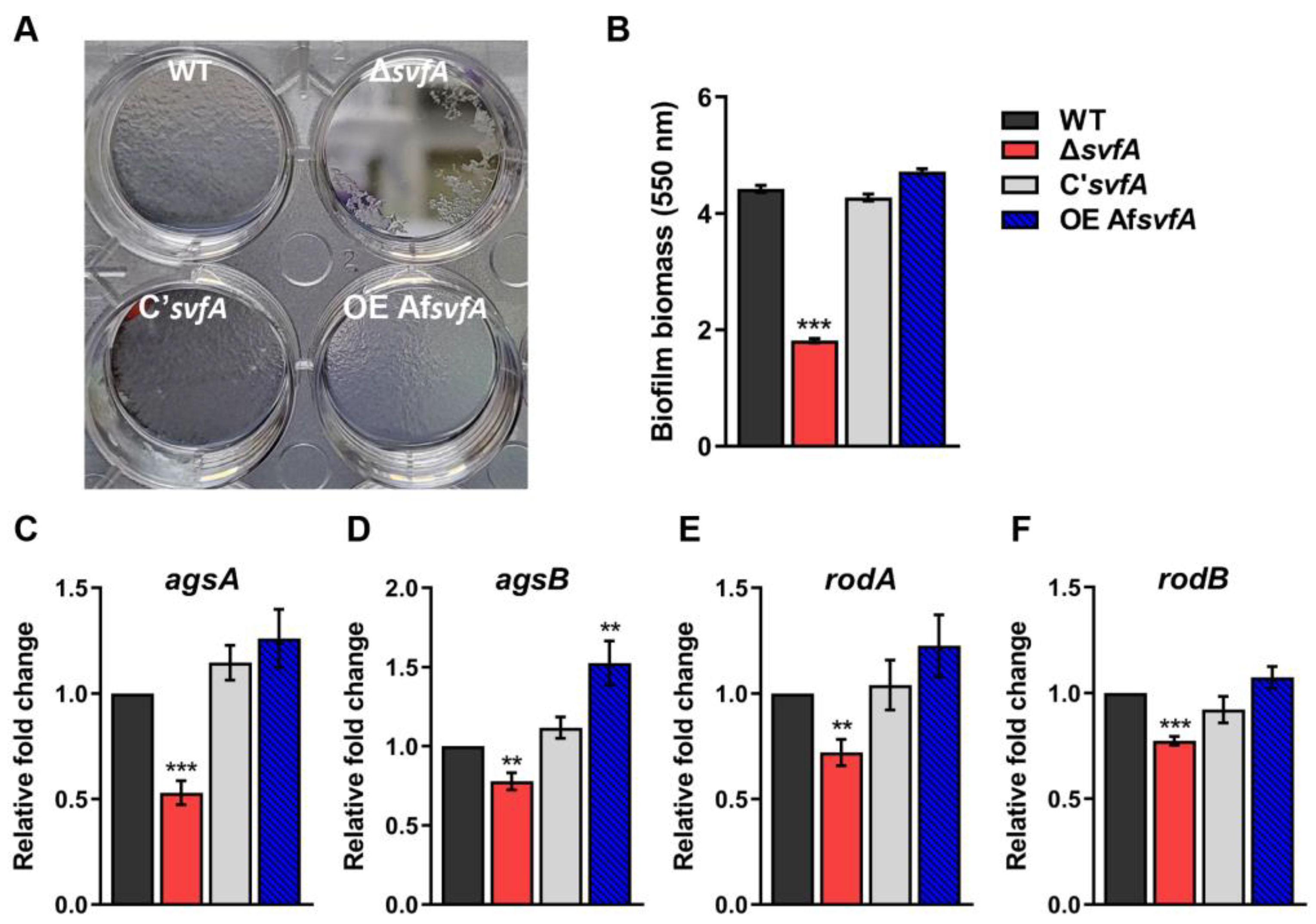
3.1. SvfA Was Involved in Biofilm Formation
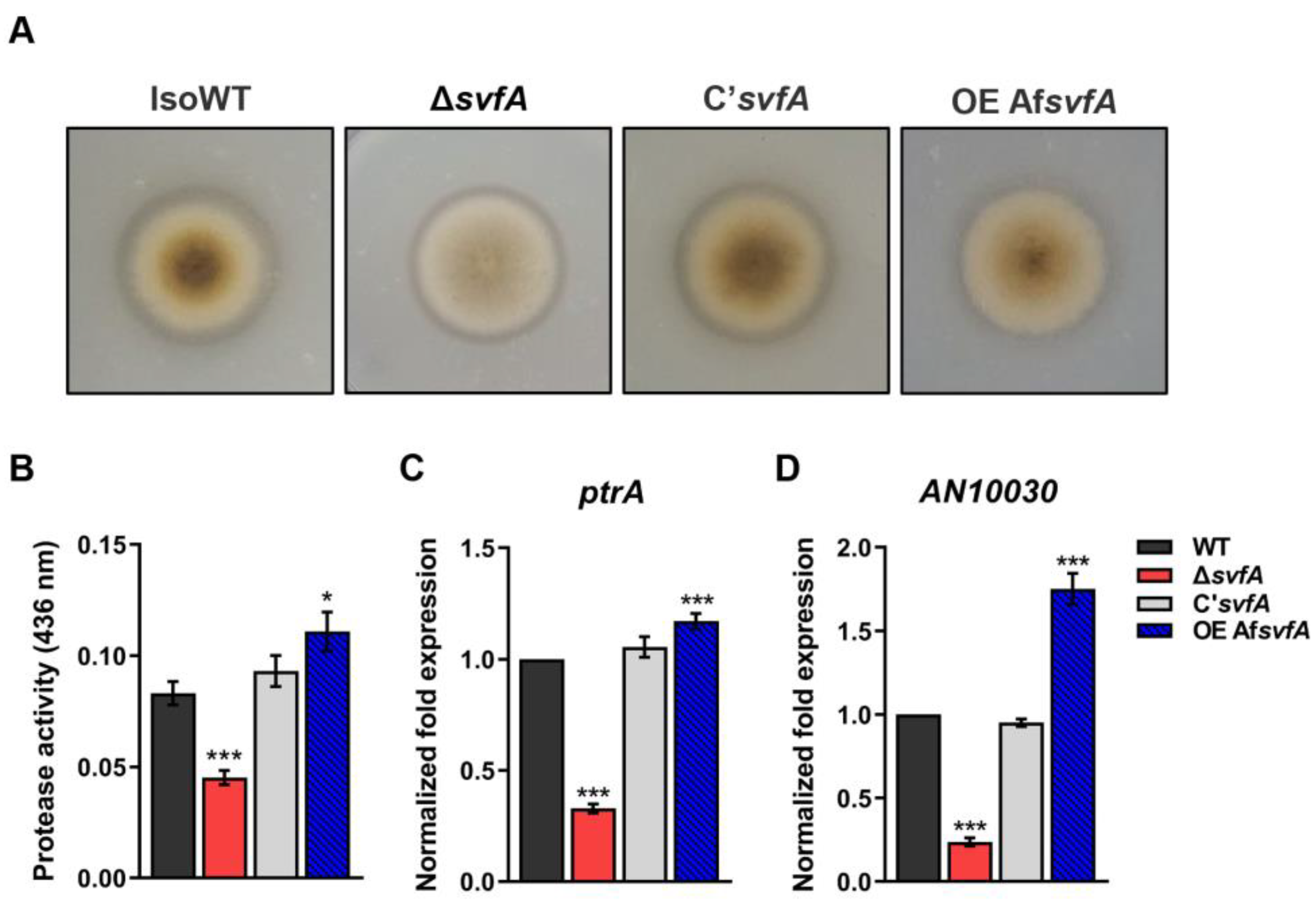
3.2. SvfA Modulated Protease Activity
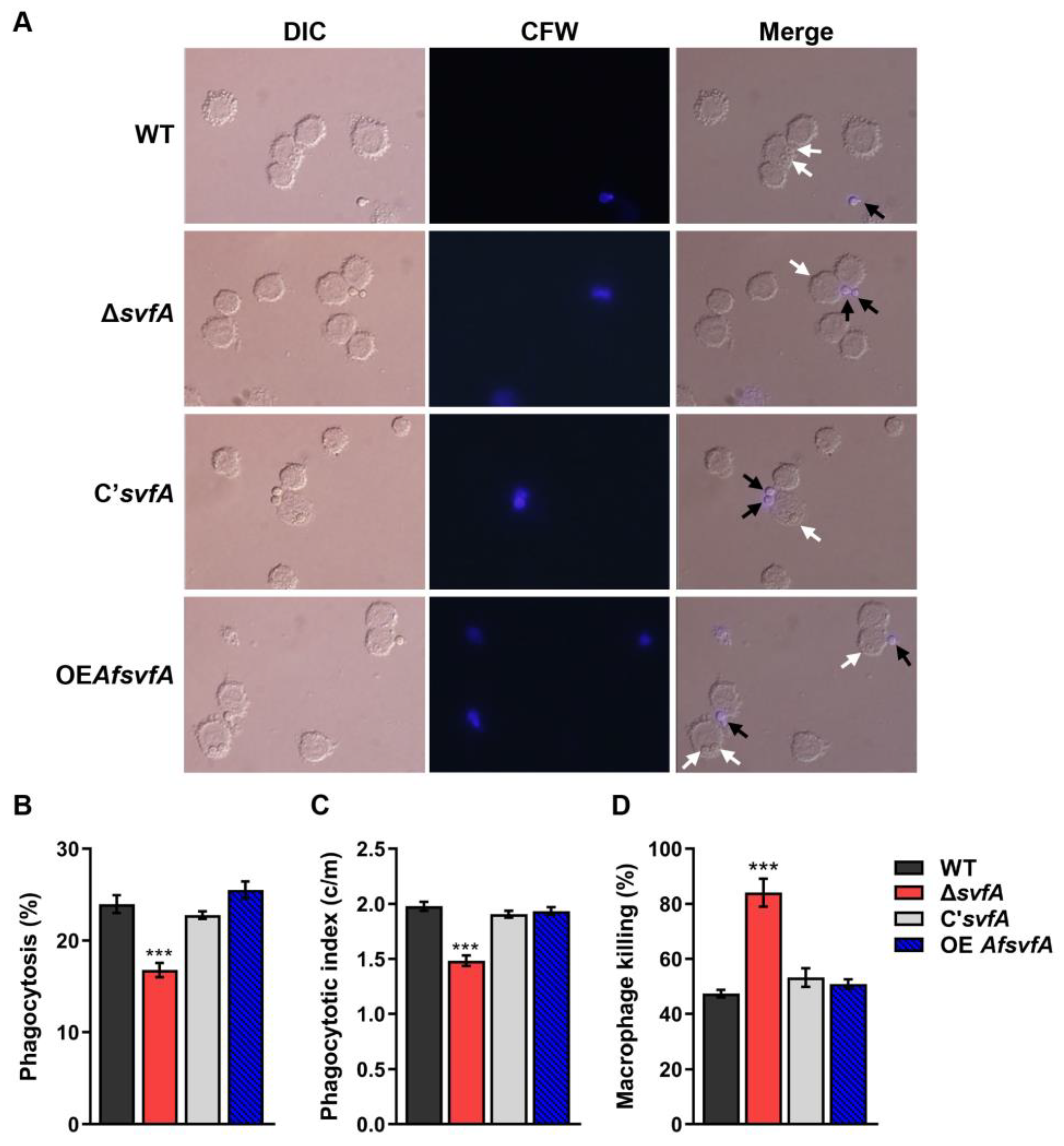
3.3. The Conidia of ΔsvfA were Susceptible to Killing by AMs
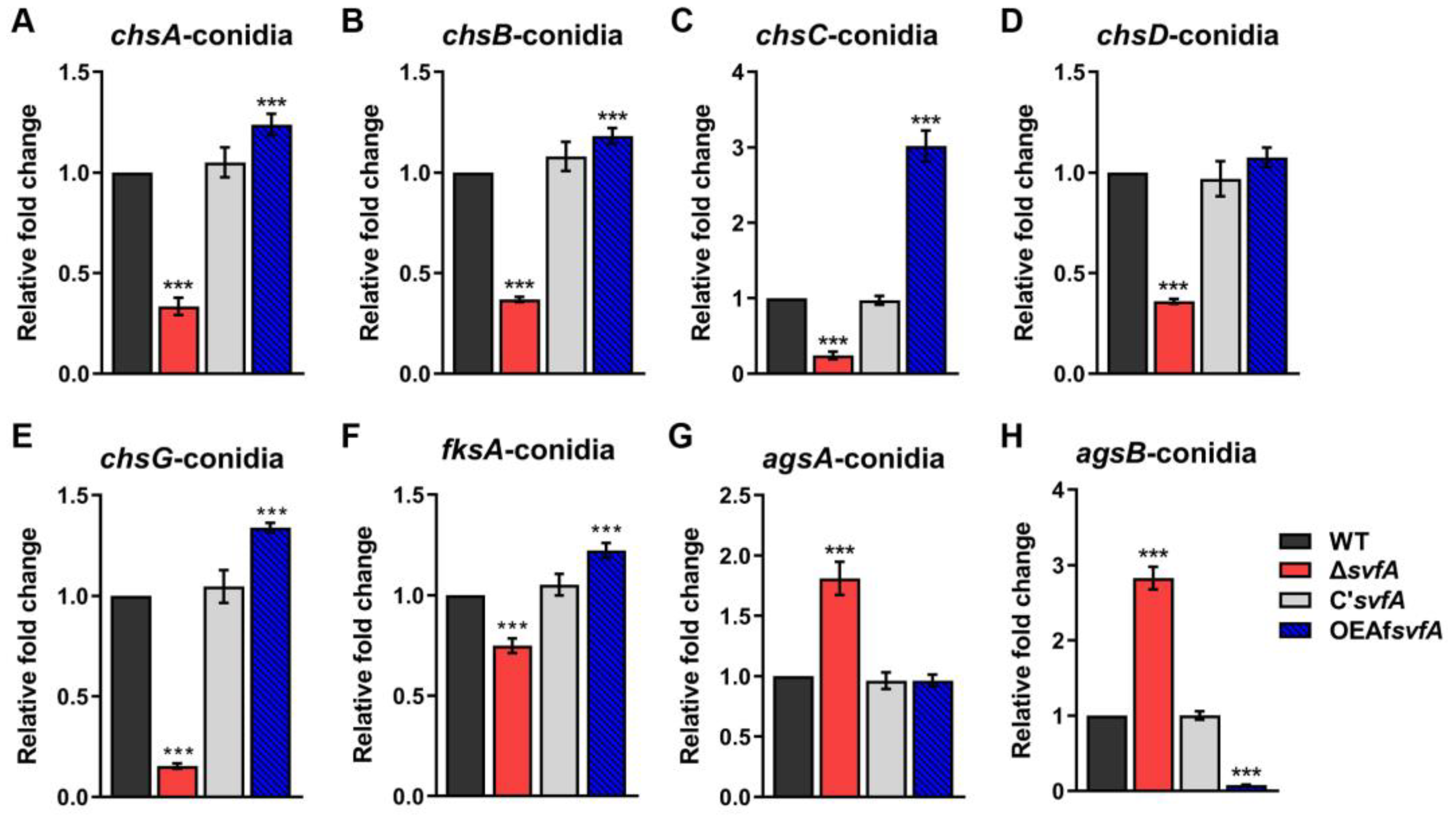
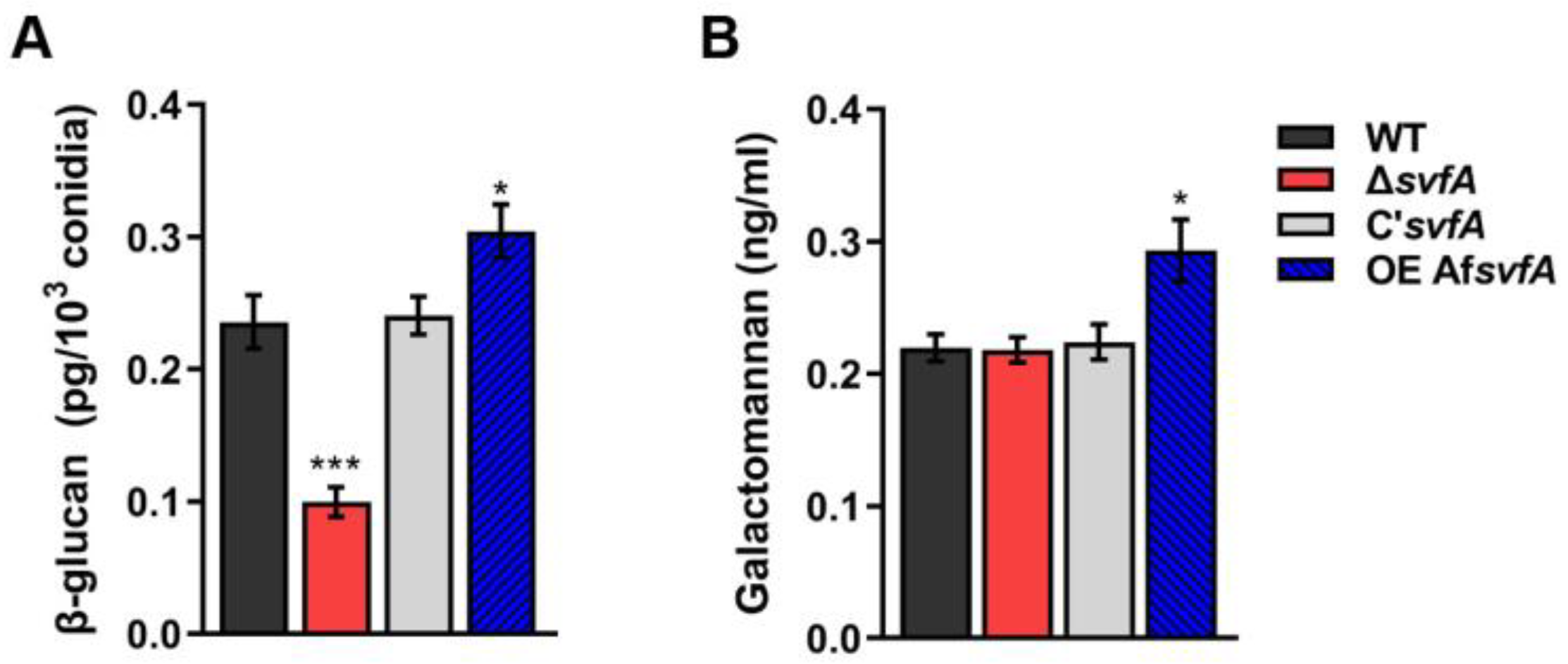
3.4. SvfA Affected the Content of Cell Wall PAMPs
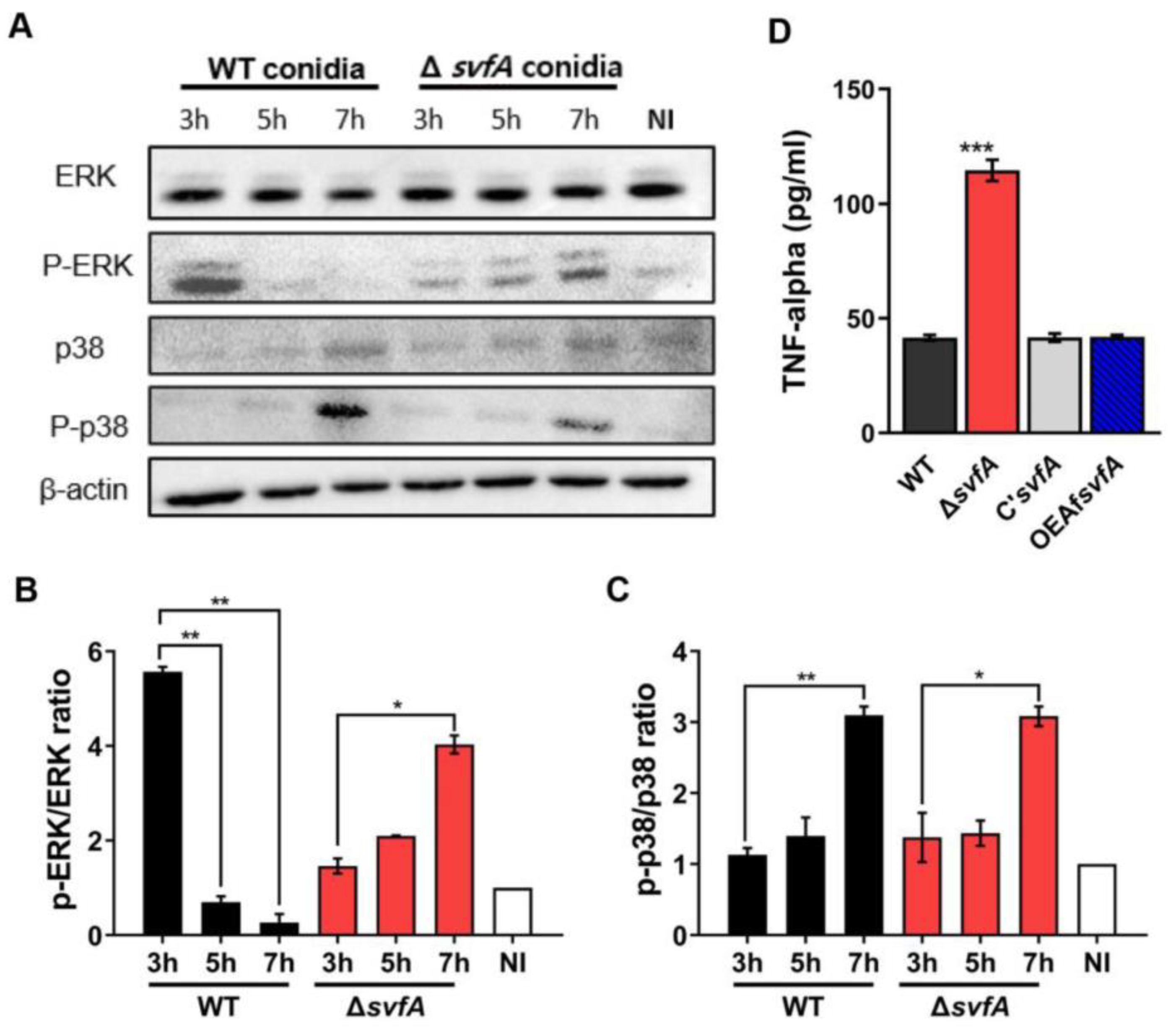
3.5. Macrophages Challenged with ΔsvfA Conidia Affected ERK Phosphorylation and Produced More TNF-α
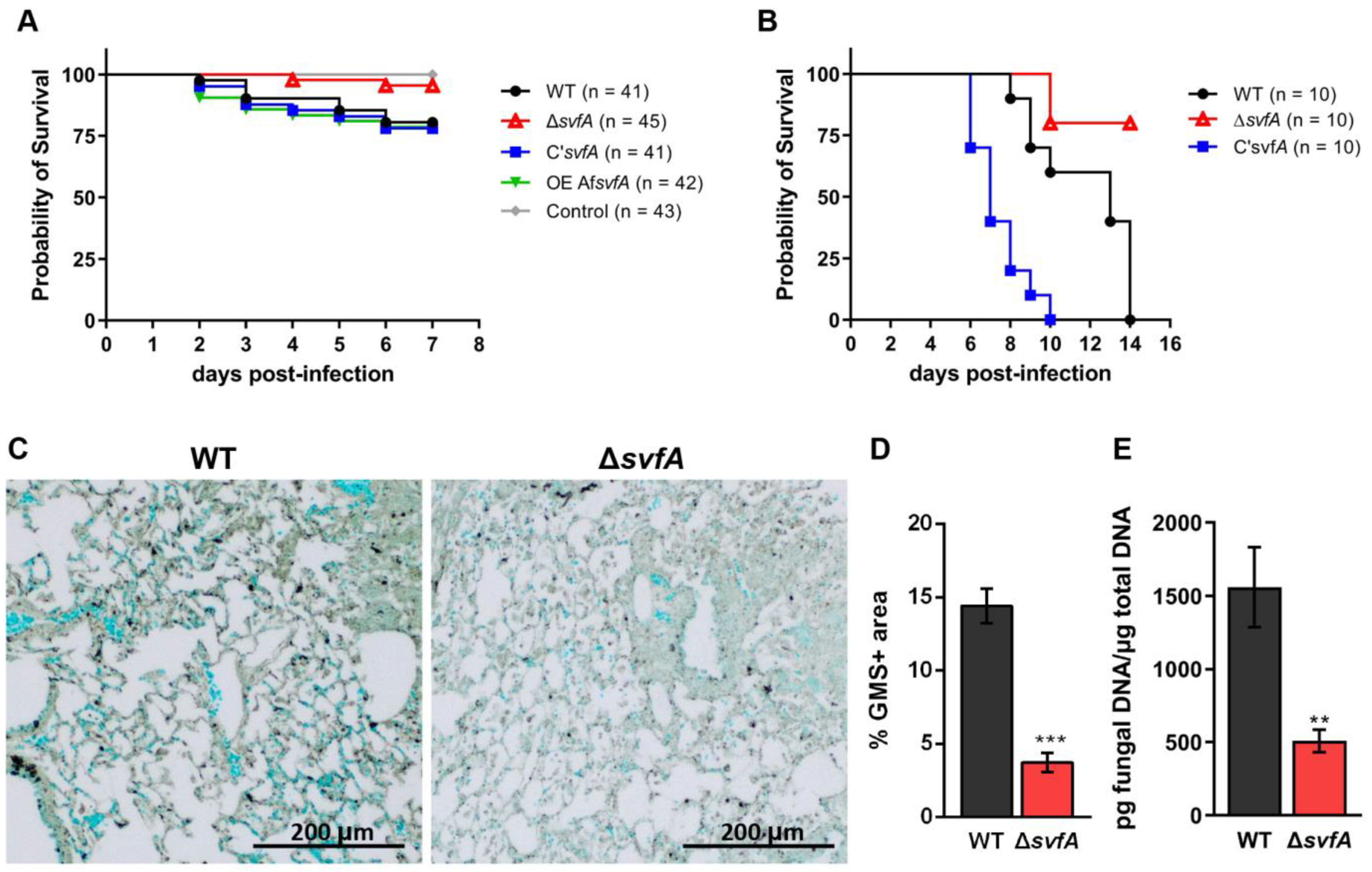
3.6. Lack of SvfA Attenuated the Virulence of A. nidulans in the T-Cell-Deficient Zebrafish and the CGD Mice Model
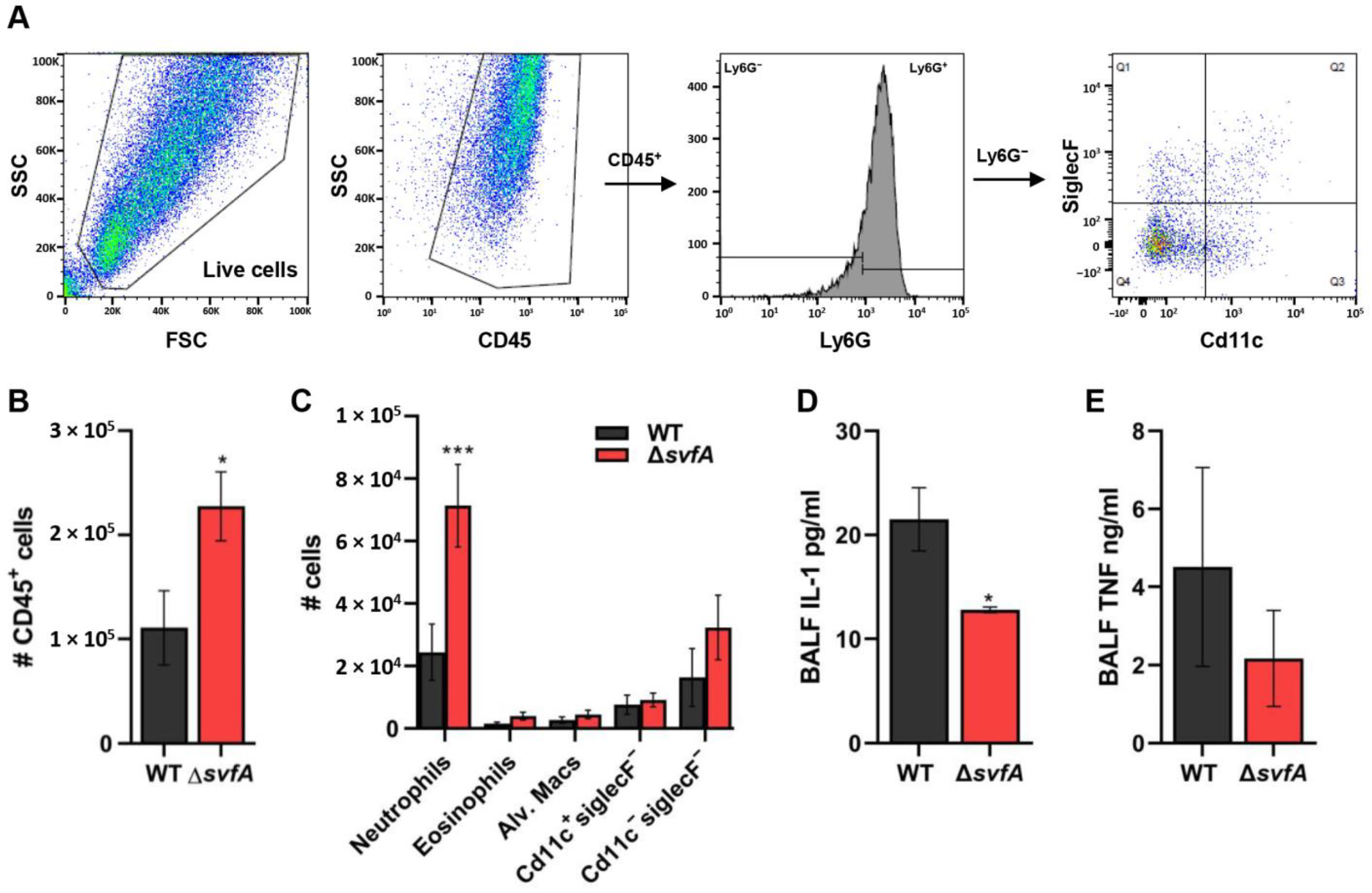
3.7. CGD Mice Infected with the ΔsvfA Conidia Exhibited Decreased Disease Pathology
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Souza, C.P.C.; Osmani, S.A. Mitotic Cell Cycle Control. In Cellular and Molecular Biology of Filamentous Fungi; ASM Press: Washington, DC, USA, 2010; pp. 61–80. [Google Scholar] [CrossRef]
- Fischer, R.; Zekert, N.; Takeshita, N. Polarized Growth in Fungi--Interplay between the Cytoskeleton, Positional Markers and Membrane Domains. Mol. Microbiol. 2008, 68, 813–826. [Google Scholar] [CrossRef] [PubMed]
- Bayram, Ö.; Braus, G.H. Coordination of Secondary Metabolism and Development in Fungi: The Velvet Family of Regulatory Proteins. FEMS Microbiol. Rev. 2012, 36, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Kasahara, S.; Jhingran, A.; Tosini, N.L.; Zhai, B.; Aufiero, M.A.; Mills, K.A.M.; Gjonbalaj, M.; Espinosa, V.; Rivera, A.; et al. During Aspergillus Infection, Monocyte-Derived DCs, Neutrophils, and Plasmacytoid DCs Enhance Innate Immune Defense through CXCR3-Dependent Crosstalk. Cell Host Microbe 2020, 28, 104–116.e4. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, V.; Jhingran, A.; Dutta, O.; Kasahara, S.; Donnelly, R.; Du, P.; Rosenfeld, J.; Leiner, I.; Chen, C.C.; Ron, Y.; et al. Inflammatory Monocytes Orchestrate Innate Antifungal Immunity in the Lung. PLoS Pathog. 2014, 10, e1003940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langfelder, K.; Jahn, B.; Gehringer, H.; Schmidt, A.; Wanner, G.; Brakhage, A.A. Identification of a Polyketide Synthase Gene (pksP) of Aspergillus fumigatus Involved in Conidial Pigment Biosynthesis and Virulence. Med. Microbiol. Immunol. 1998, 187, 79–89. [Google Scholar] [CrossRef]
- Henriet, S.S.V.; Verweij, P.E.; Warris, A. Aspergillus nidulans and Chronic Granulomatous Disease: A Unique Host–Pathogen Interaction. J. Infect. Dis. 2012, 206, 1128–1137. [Google Scholar] [CrossRef]
- Panday, A.; Sahoo, M.K.; Osorio, D.; Batra, S. NADPH Oxidases: An Overview from Structure to Innate Immunity-Associated Pathologies. Cell Mol. Immunol. 2015, 12, 5–23. [Google Scholar] [CrossRef] [Green Version]
- Anjani, G.; Vignesh, P.; Joshi, V.; Shandilya, J.K.; Bhattarai, D.; Sharma, J.; Rawat, A. Recent Advances in Chronic Granulomatous Disease. Genes Dis. 2019, 7, 84–92. [Google Scholar] [CrossRef]
- Reiko Minakami; Hideki Sumimoto Phagocytosis-Coupled Activation of the Superoxide-Producing Phagocyte Oxidase, a Member of the NADPH Oxidase (Nox) Family. Int. J. Hematol. 2006, 84, 193–198. [CrossRef]
- Dotis, J.; Roilides, E. Osteomyelitis Due to Aspergillus Spp. in Patients with Chronic Granulomatous Disease: Comparison of Aspergillus nidulans and Aspergillus fumigatus. Int. J. Infect. Dis. 2004, 8, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Åhlin, A.; de Boer, M.; Roos, D.; Leusen, J.; Smith, C.; Sundin, U.; Rabbani, H.; Palmblad, J.; Elinder, G. Prevalence, Genetics and Clinical Presentation of Chronic Granulomatous Disease in Sweden. Acta Paediatr. 1995, 84, 1386–1394. [Google Scholar] [CrossRef] [PubMed]
- Erwig, L.P.; Gow, N.A.R. Interactions of Fungal Pathogens with Phagocytes. Nat. Rev. Microbiol. 2016, 14, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Hatinguais, R.; Willment, J.A.; Brown, G.D. PAMPs of the Fungal Cell Wall and Mammalian PRRs. Curr. Top. Microbiol. Immunol. 2020, 425, 187–223. [Google Scholar] [CrossRef] [PubMed]
- Vega, K.; Kalkum, M. Chitin, Chitinase Responses, and Invasive Fungal Infections. Int. J. Microbiol. 2012, 2012, 920459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, K.M. MAP Kinase Activation in Macrophages. J. Leukoc. Biol. 2001, 69, 3–10. [Google Scholar] [CrossRef]
- Dubourdeau, M.; Athman, R.; Balloy, V.; Philippe, B.; Sengmanivong, L.; Chignard, M.; Philpott, D.J.; Latgé, J.P.; Ibrahim-Granet, O. Interaction of Aspergillus fumigatus with the Alveolar Macrophage. Med. Mycol. 2006, 44, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Loussert, C.; Schmitt, C.; Prevost, M.C.; Balloy, V.; Fadel, E.; Philippe, B.; Kauffmann-Lacroix, C.; Latgé, J.P.; Beauvais, A. In vivo Biofilm Composition of Aspergillus fumigatus. Cell Microbiol. 2010, 12, 405–410. [Google Scholar] [CrossRef]
- Beauvais, A.; Latgé, J.-P. Aspergillus Biofilm in vitro and in vivo. Microbiol. Spectr. 2015, 3, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Kernien, J.F.; Snarr, B.D.; Sheppard, D.C.; Nett, J.E. The Interface between Fungal Biofilms and Innate Immunity. Front. Immunol. 2018, 8, 1968. [Google Scholar] [CrossRef]
- Lee, M.J.; Liu, H.; Barker, B.M.; Snarr, B.D.; Gravelat, F.N.; al Abdallah, Q.; Gavino, C.; Baistrocchi, S.R.; Ostapska, H.; Xiao, T.; et al. The Fungal Exopolysaccharide Galactosaminogalactan Mediates Virulence by Enhancing Resistance to Neutrophil Extracellular Traps. PLoS Pathog. 2015, 11, e1005187. [Google Scholar] [CrossRef] [Green Version]
- Mowat, E.; Butcher, J.; Lang, S.; Williams, C.; Ramage, G. Development of a Simple Model for Studying the Effects of Antifungal Agents on Multicellular Communities of Aspergillus fumigatus. J. Med. Microbiol. 2007, 56, 1205–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van de Veerdonk, F.L.; Gresnigt, M.S.; Romani, L.; Netea, M.G.; Latgé, J.P. Aspergillus fumigatus Morphology and Dynamic Host Interactions. Nat. Rev. Microbiol. 2017, 15, 661–674. [Google Scholar] [CrossRef]
- Redes, J.L.; Basu, T.; Ram-Mohan, S.; Ghosh, C.C.; Chan, E.C.; Sek, A.C.; Zhao, M.; Krishnan, R.; Rosenberg, H.F.; Druey, K.M. Aspergillus fumigatus-Secreted Alkaline Protease 1 Mediates Airways Hyperresponsiveness in Severe Asthma. Immunohorizons 2019, 3, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Balenga, N.A.; Klichinsky, M.; Xie, Z.; Chan, E.C.; Zhao, M.; Jude, J.; Laviolette, M.; Panettieri, R.A.; Druey, K.M. A Fungal Protease Allergen Provokes Airway Hyper-Responsiveness in Asthma. Nat. Commun. 2015, 6, 6763. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.-Y.; Kang, E.-H.; Park, Y.-H.; Kook, J.-H.; Park, H.-M. Survival Factor SvfA Plays Multiple Roles in Differentiation and Is Essential for Completion of Sexual Development in Aspergillus nidulans. Sci. Rep. 2020, 10, 5586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vander Heiden, M.G.; Choy, J.S.; VanderWeele, D.J.; Brace, J.L.; Harris, M.H.; Bauer, D.E.; Prange, B.; Kronll, S.J.; Thompson, C.B.; Rudin, C.M. Bcl-x(L) Complements Saccharomyces cerevisiae Genes That Facilitate the Switch from Glycolytic to Oxidative Metabolism. J. Biol. Chem. 2002, 277, 44870–44876. [Google Scholar] [CrossRef] [Green Version]
- Brace, J.L.; VanderWeele, D.J.; Rudin, C.M. Svf1 Inhibits Reactive Oxygen Species Generation and Promotes Survival under Conditions of Oxidative Stress in Saccharomyces cerevisiae. Yeast 2005, 22, 641–652. [Google Scholar] [CrossRef]
- Aharoni-Simon, M.; Shumiatcher, R.; Yeung, A.; Shih, A.Z.L.; Dolinsky, V.W.; Doucette, C.A.; Luciani, D.S. Bcl-2 Regulates Reactive Oxygen Species Signaling and a Redox-Sensitive Mitochondrial Proton Leak in Mouse Pancreatic β-Cells. Endocrinology 2016, 157, 2270–2281. [Google Scholar] [CrossRef] [Green Version]
- Brace, J.L.; Lester, R.L.; Dickson, R.C.; Rudin, C.M. SVF1 Regulates Cell Survival by Affecting Sphingolipid Metabolism in Saccharomyces cerevisiae. Genetics 2007, 175, 65–76. [Google Scholar] [CrossRef]
- Li, T.; Jung, B.; Park, S.Y.; Lee, J. Survival Factor Gene FgSvf1 Is Required for Normal Growth and Stress Resistance in Fusarium graminearum. Plant Pathol. J. 2019, 35, 393–405. [Google Scholar] [CrossRef]
- Yu, Y.; Du, J.; Wang, Y.; Zhang, M.; Huang, Z.; Cai, J.; Fang, A.; Yang, Y.; Qing, L.; Bi, C.; et al. Survival Factor 1 Contributes to the Oxidative Stress Response and Is Required for Full Virulence of Sclerotinia Sclerotiorum. Mol. Plant Pathol. 2019, 20, 895–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, P.; Ma, D.; Gao, S.; Zhang, Y.; Bae, Y.K.; Liang, G.; Gao, S.; Choi, J.H.; Kim, C.H.; Wang, L.; et al. Generation of foxn1/Casper Mutant Zebrafish for Allograft and Xenograft of Normal and Malignant Cells. Stem Cell Rep. 2020, 15, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.-Y.; Kim, Y.J.; Woo, S.A.; Jeong, J.W.; Lee, Y.-R.; Kim, C.-H.; Park, H.-M. The LAMMER Kinase, LkhA, Affects Aspergillus fumigatus Pathogenicity by Modulating Reproduction and Biosynthesis of Cell Wall PAMPs. Front. Cell Infect. Microbiol. 2021, 11, 756206. [Google Scholar] [CrossRef] [PubMed]
- Shemesh, E.; Hanf, B.; Hagag, S.; Attias, S.; Shadkchan, Y.; Fichtman, B.; Harel, A.; Krüger, T.; Brakhage, A.A.; Kniemeyer, O.; et al. Phenotypic and Proteomic Analysis of the Aspergillus fumigatus ΔPrtT, ΔXprG and ΔXprG/ΔPrtT Protease-Deficient Mutants. Front. Microbiol. 2017, 8, 2490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, M.C.; de Godoy, K.F.; de Castro, P.A.; Hori, J.I.; Pedro Bom, V.L.; Brown, N.A.; da Cunha, A.F.; Goldman, G.H.; Malavazi, I.; Yu, J.H. The Aspergillus fumigatus PkcAG579R Mutant Is Defective in the Activation of the Cell Wall Integrity Pathway but Is Dispensable for Virulence in a Neutropenic Mouse Infection Model. PLoS ONE 2015, 10, e0135195. [Google Scholar] [CrossRef] [Green Version]
- Park, D.S.; Yu, Y.M.; Kim, Y.J.; Maeng, P.J. Negative Regulation of the Vacuole-Mediated Resistance to K+ Stress by a Novel C2H2 Zinc Finger Transcription Factor Encoded by AslA in Aspergillus nidulans. J. Microbiol. 2015, 53, 100–110. [Google Scholar] [CrossRef]
- Matsunaga, K.; Klein, T.W.; Friedman, H.; Yamamoto, Y. Alveolar Macrophage Cell Line MH-S Is Valuable as an in vitro Model for Legionella pneumophila Infection. Am. J. Respir. Cell Mol. Biol. 2001, 24, 326–331. [Google Scholar] [CrossRef]
- Liu, C.; Wang, M.; Sun, W.; Cai, F.; Geng, S.; Su, X.; Shi, Y. PU.1 Serves a Critical Role in the Innate Defense against Aspergillus fumigatus via Dendritic Cell-Associated C-Type Lectin Receptor-1 and Toll-like Receptors-2 and 4 in THP-1-Derived Macrophages. Mol. Med. Rep. 2017, 15, 4084–4092. [Google Scholar] [CrossRef] [Green Version]
- Dubourdeau, M.; Athman, R.; Balloy, V.; Huerre, M.; Chignard, M.; Philpott, D.J.; Latgé, J.-P.; Ibrahim-Granet, O. Aspergillus fumigatus Induces Innate Immune Responses in Alveolar Macrophages through the MAPK Pathway Independently of TLR2 and TLR4. J. Immunol. 2006, 177, 3994–4001. [Google Scholar] [CrossRef]
- O’Dea, E.M.; Amarsaikhan, N.; Li, H.; Downey, J.; Steele, E.; van Dyken, S.J.; Locksley, R.M.; Templeton, S.P. Eosinophils Are Recruited in Response to Chitin Exposure and Enhance Th2-Mediated Immune Pathology in Aspergillus fumigatus Infection. Infect. Immun. 2014, 82, 3199–3205. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, S.; Amarsaikhan, N.; Maupin, A.J.; Schlosser, A.; Füchtbauer, E.-M.; Holmskov, U.; Moeller, J.B.; Templeton, S.P. FIBCD1 Deficiency Decreases Disease Severity in a Murine Model of Invasive Pulmonary Aspergillosis. Immunohorizons 2021, 5, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Amarsaikhan, N.; Sands, E.M.; Shah, A.; Abdolrasouli, A.; Reed, A.; Slaven, J.E.; Armstrong-James, D.; Templeton, S.P. Caspofungin Increases Fungal Chitin and Eosinophil and γδ T Cell-Dependent Pathology in Invasive Aspergillosis. J. Immunol. 2017, 199, 624–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauvais, A.; Schmidt, C.; Guadagnini, S.; Roux, P.; Perret, E.; Henry, C.; Paris, S.; Mallet, A.; Prévost, M.-C.; Latgé, J.P. An Extracellular Matrix Glues Together the Aerial-Grown Hyphae of Aspergillus fumigatus. Cell. Microbiol. 2007, 9, 1588–1600. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Parsania, C.; Tan, K.; Todd, R.B.; Wong, K.H. Co-Option of an Extracellular Protease for Transcriptional Control of Nutrient Degradation in the Fungus Aspergillus nidulans. Commun. Biol. 2021, 4, 1409. [Google Scholar] [CrossRef] [PubMed]
- Sriranganadane, D.; Waridel, P.; Salamin, K.; Reichard, U.; Grouzmann, E.; Neuhaus, J.M.; Quadroni, M.; Monod, M. Aspergillus Protein Degradation Pathways with Different Secreted Protease Sets at Neutral and Acidic pH. J. Proteome Res. 2010, 9, 3511–3519. [Google Scholar] [CrossRef] [PubMed]
- Castro-Ochoa, D.; Peña-Montes, C.; Farrés, A. Evaluation of Strategies to Improve the Production of Alkaline Protease PrtA from Aspergillus nidulans. Appl. Biochem. Biotechnol. 2013, 169, 1672–1682. [Google Scholar] [CrossRef] [PubMed]
- Stajich, J.E.; Harris, T.; Brunk, B.P.; Brestelli, J.; Fischer, S.; Harb, O.S.; Kissinger, J.C.; Li, W.; Nayak, V.; Pinney, D.F.; et al. FungiDB: An Integrated Functional Genomics Database for Fungi. Nucleic Acids Res. 2012, 40, D675–D681. [Google Scholar] [CrossRef]
- Antl, M.; Huber, B.; Stoecker, K.; Ebrecht, D.; Steinmetz, I.; Eberl, L. Killing of Aspergillus fumigatus by Alveolar Macrophages Is Mediated by Reactive Oxidant Intermediates. Cell. Microbiol. 2003, 5, 343–351. [Google Scholar] [CrossRef]
- Patin, E.C.; Thompson, A.; Orr, S.J. Pattern Recognition Receptors in Fungal Immunity. Semin. Cell Dev. Biol. 2019, 89, 24–33. [Google Scholar] [CrossRef]
- Lenardon, M.D.; Munro, C.A.; Gow, N.A.R.R. Chitin Synthesis and Fungal Pathogenesis. Curr. Opin. Microbiol. 2010, 13, 416–423. [Google Scholar] [CrossRef] [Green Version]
- Yoshimi, A.; Miyazawa, K.; Abe, K. Function and Biosynthesis of Cell Wall α-1,3-Glucan in Fungi. J. Fungi 2017, 3, 63. [Google Scholar] [CrossRef] [PubMed]
- Park, B.-C.; Park, Y.-H.; Yi, S.; Choi, Y.K.; Kang, E.-H.; Park, H.-M. Transcriptional Regulation of fksA, a β-1,3-Glucan Synthase Gene, by the APSES Protein StuA during Aspergillus nidulans Development. J. Microbiol. 2014, 52, 940–947. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Gómez, D.; Domínguez-Soto, A.; Ancochea, J.; Jimenez-Heffernan, J.A.; Leal, J.A.; Corbí, A.L. Dendritic Cell-Specific Intercellular Adhesion Molecule 3-Grabbing Nonintegrin Mediates Binding and Internalization of Aspergillus fumigatus Conidia by Dendritic Cells and Macrophages. J. Immunol. 2004, 173, 5635–5643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akira, S.; Takeda, K. Toll-like Receptor Signaling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Ulevitch, R.J. Limiting Inflammatory Responses during Activation of Innate Immunity. Nat. Immunol. 2005, 6, 1198–1205. [Google Scholar] [CrossRef]
- Hohl, T.M.; van Epps, H.L.; Rivera, A.; Morgan, L.A.; Chen, P.L.; Feldmesser, M.; Pamer, E.G. Aspergillus fumigatus Triggers Inflammatory Responses by Stage-Specific β-Glucan Display. PLoS Pathog. 2005, 1, e30. [Google Scholar] [CrossRef] [Green Version]
- Rosowski, E.E.; Knox, B.P.; Archambault, L.S.; Huttenlocher, A.; Keller, N.P.; Wheeler, R.T.; Davis, J.M. The Zebrafish as a Model Host for Invasive Fungal Infections. J. Fungi 2018, 4, 136. [Google Scholar] [CrossRef] [Green Version]
- Alexis Niño-Vega, G.; Gonçalves, T.; Anne Hall, R.; Trevijano-Contador, N.; Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J. The Fungal Cell Wall: Candida, Cryptococcus, and Aspergillus Species. Front. Microbiol. 2020, 10, 2993. [Google Scholar] [CrossRef]
- Munro, C. Candida albicans Cell Wall Mediated Virulence. In Pathogenic Yeasts; Springer: Berlin/Heidelberg, Germany, 2010; pp. 69–95. [Google Scholar]
- Valiante, V.; Macheleidt, J.; Föge, M.; Brakhage, A.A. The Aspergillus fumigatus Cell Wall Integrity Signaling Pathway: Drug Target, Compensatory Pathways and Virulence. Front. Microbiol. 2015, 6, 325. [Google Scholar] [CrossRef]
- Latgé, J.P.; Beauvais, A.; Chamilos, G. The Cell Wall of the Human Fungal Pathogen Aspergillus fumigatus: Biosynthesis, Organization, Immune Response, and Virulence. Annu. Rev. Microbiol. 2017, 71, 99–116. [Google Scholar] [CrossRef]
- Liu, Z.; Raj, S.; van Rhijn, N.; Fraczek, M.; Michel, J.P.; Sismeiro, O.; Legendre, R.; Varet, H.; Fontaine, T.; Bromley, M.; et al. Functional Genomic and Biochemical Analysis Reveals Pleiotropic Effect of Congo Red on Aspergillus fumigatus. mBio 2021, 12, e00863-21. [Google Scholar] [CrossRef] [PubMed]
- Wiederhold, N.P.; Kontoyiannis, D.P.; Chi, J.; Prince, R.A.; Tam, V.H.; Lewis, R.E. Pharmacodynamics of Caspofungin in a Murine Model of Invasive Pulmonary Aspergillosis: Evidence of Concentration-Dependent Activity. J. Infect. Dis. 2004, 190, 1464–1471. [Google Scholar] [CrossRef] [PubMed]
- Beauvais, A.; Bozza, S.; Kniemeyer, O.; Formosa, C.; Balloy, V.; Henry, C.; Roberson, R.W.; Dague, E.; Chignard, M.; Brakhage, A.A.; et al. Deletion of the α-(1,3)-Glucan Synthase Genes Induces a Restructuring of the Conidial Cell Wall Responsible for the Avirulence of Aspergillus fumigatus. PLoS Pathog. 2013, 9, e1003716. [Google Scholar] [CrossRef]
- Rappleye, C.A.; Eissenberg, L.G.; Goldman, W.E. Histoplasma Capsulatum α-(1,3)-Glucan Blocks Innate Immune Recognition by the β-Glucan Receptor. Proc. Natl. Acad. Sci. USA 2007, 104, 1366–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, J.; Orth, K. Rise of a Cereal Killer: The Biology of Magnaporthe Oryzae Biotrophic Growth. Trends Microbiol. 2018, 26, 582–597. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Singh, S. Biofilm Formation by Aspergillus fumigatus. Med. Mycol. 2014, 52, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Beauvais, A.; Maubon, D.; Park, S.; Morelle, W.; Tanguy, M.; Huerre, M.; Perlin, D.S.; Latgé, J.P. Two α(1-3) Glucan Synthases with Different Functions in Aspergillus fumigatus. Appl. Environ. Microbiol. 2005, 71, 1531–1538. [Google Scholar] [CrossRef] [Green Version]
- Maubon, D.; Park, S.; Tanguy, M.; Huerre, M.; Schmitt, C.; Prévost, M.C.; Perlin, D.S.; Latgé, J.P.; Beauvais, A. AGS3, an α(1-3)Glucan Synthase Gene Family Member of Aspergillus fumigatus, Modulates Mycelium Growth in the Lung of Experimentally Infected Mice. Fungal Genet. Biol. 2006, 43, 366–375. [Google Scholar] [CrossRef]
- Yoshimi, A.; Sano, M.; Inaba, A.; Kokubun, Y.; Fujioka, T.; Mizutani, O.; Hagiwara, D.; Fujikawa, T.; Nishimura, M.; Yano, S.; et al. Functional Analysis of the α-1,3-Glucan Synthase Genes agsA and agsB in Aspergillus nidulans: AgsB Is the Major α-1,3-Glucan Synthase in This Fungus. PLoS ONE 2013, 8, e54893. [Google Scholar] [CrossRef]
- Gibbons, J.G.; Beauvais, A.; Beau, R.; McGary, K.L.; Latgé, J.P.; Rokas, A. Global Transcriptome Changes Underlying Colony Growth in the Opportunistic Human Pathogen Aspergillus fumigatus. Eukaryot. Cell 2012, 11, 68–78. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Liang, H.; Zen, K. Molecular Mechanisms That Influence the Macrophage M1-M2 Polarization Balance. Front Immunol 2014, 5, 614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinekamp, T.; Thywißen, A.; Macheleidt, J.; Keller, S.; Valiante, V.; Brakhage, A.A. Aspergillus fumigatus Melanins: Interference with the Host Endocytosis Pathway and Impact on Virulence. Front. Microbiol. 2012, 3, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastos, R.W.; Valero, C.; Silva, L.P.; Schoen, T.; Drott, M.; Brauer, V.; Silva-Rocha, R.; Lind, A.; Steenwyk, J.L.; Rokas, A.; et al. Functional Characterization of Clinical Isolates of the Opportunistic Fungal Pathogen Aspergillus nidulans. mSphere 2020, 5, e00153-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghezzi, P.; Dinarello, C.A.; Bianchi, M.; Rosandich, M.E.; Repine, J.E.; White, C.W. Hypoxia Increases Production of Interleukin-1 and Tumor Necrosis Factor by Human Mononuclear Cells. Cytokine 1991, 3, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Grahl, N.; Puttikamonkul, S.; Macdonald, J.M.; Gamcsik, M.P.; Ngo, L.Y.; Hohl, T.M.; Cramer, R.A. In vivo Hypoxia and a Fungal Alcohol Dehydrogenase Influence the Pathogenesis of Invasive Pulmonary Aspergillosis. PLoS Pathog. 2011, 7, e1002145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mircescu, M.M.; Lipuma, L.; van Rooijen, N.; Pamer, E.G.; Hohl, T.M. Essential Role for Neutrophils but Not Alveolar Macrophages at Early Time Points Following Aspergillus fumigatus Infection. J. Infect. Dis. 2009, 200, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Morgenstern, D.E.; Gifford, M.A.C.; Li, L.L.; Doerschuk, C.M.; Dinauer, M.C. Absence of Respiratory Burst in X-Linked Chronic Granulomatous Disease Mice Leads to Abnormalities in Both Host Defense and Inflammatory Response to Aspergillus fumigatus. J. Exp. Med. 1997, 185, 207–218. [Google Scholar] [CrossRef]
- Hattori, H.; Subramanian, K.K.; Sakai, J.; Jia, Y.; Li, Y.; Porter, T.F.; Loison, F.; Sarraj, B.; Kasorn, A.; Jo, H.; et al. Small-Molecule Screen Identifies Reactive Oxygen Species as Key Regulators of Neutrophil Chemotaxis. Proc. Natl. Acad. Sci. USA 2010, 107, 3546–3551. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, J.-Y.; Jung, Y.-E.; Hwang, H.-E.; Kim, C.-H.; Basaran-Akgul, N.; Goli, S.H.; Templeton, S.P.; Park, H.-M. Survival Factor A (SvfA) Contributes to Aspergillus nidulans Pathogenicity. J. Fungi 2023, 9, 143. https://doi.org/10.3390/jof9020143
Lim J-Y, Jung Y-E, Hwang H-E, Kim C-H, Basaran-Akgul N, Goli SH, Templeton SP, Park H-M. Survival Factor A (SvfA) Contributes to Aspergillus nidulans Pathogenicity. Journal of Fungi. 2023; 9(2):143. https://doi.org/10.3390/jof9020143
Chicago/Turabian StyleLim, Joo-Yeon, Ye-Eun Jung, Hye-Eun Hwang, Cheol-Hee Kim, Nese Basaran-Akgul, Sri Harshini Goli, Steven P. Templeton, and Hee-Moon Park. 2023. "Survival Factor A (SvfA) Contributes to Aspergillus nidulans Pathogenicity" Journal of Fungi 9, no. 2: 143. https://doi.org/10.3390/jof9020143