
Antifungal Activity of Chitosan against Histoplasma capsulatum in Planktonic and Biofilm Forms: A Therapeutic Strategy in the Future?
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Low Molecular Weight Chitosan Samples
2.2. Fungal Culture
2.3. Preparation of Chitosan and Control Drugs
2.4. Antifungal Susceptibility Test in Planktonic Cells of H. capsulatum
2.5. Susceptibility Test of Biofilm Formation
2.6. Biomass Quantification Assay
2.7. Metabolic Activity Quantification Assay
2.8. Evaluation of the Morphology and Structure of H. capsulatum Biofilms
2.9. Pharmacological Interaction—Checkerboard
2.10. Statistical Analyses
3. Results
3.1. Antifungal Susceptibility
3.2. Pharmacological Interactions
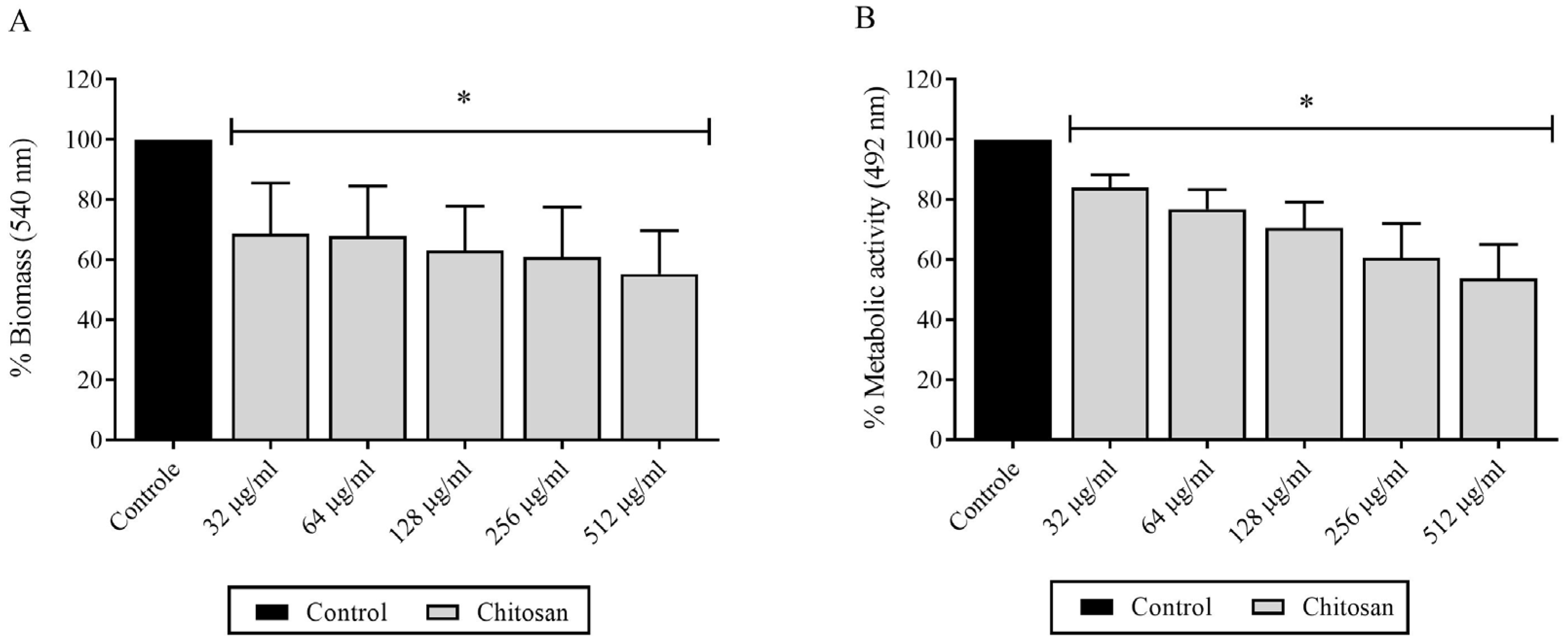
3.3. Evaluation of Chitosan Effects on H. capsulatum Biofilm Formation
3.4. Structural Analysis of Biofilms
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sil, A. Molecular Regulation of Histoplasma Dimorphism. Curr. Opin. Microbiol. 2019, 52, 151–157. [Google Scholar] [CrossRef]
- Gugnani, H.C.; Denning, D.W. Infection of Bats with Histoplasma Species. Med. Mycol. 2023, 61, myad080. [Google Scholar] [CrossRef] [PubMed]
- Adenis, A.A.; Aznar, C.; Couppié, P. Histoplasmosis in HIV-Infected Patients: A Review of New Developments and Remaining Gaps. Curr. Trop. Med. Rep. 2014, 1, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Limaye, A.P.; Connolly, P.A.; Sagar, M.; Fritsche, T.R.; Cookson, B.T.; Wheat, L.J.; Stamm, W.E. Transmission of Histoplasma Capsulatum by Organ Transplantation. N. Engl. J. Med. 2000, 343, 1163–1166. [Google Scholar] [CrossRef]
- Falci, D.R.; Dalla Lana, D.F.; Pasqualotto, A.C. The Era of Histoplasmosis in Brazilian Endemic Mycoses. Lancet Reg. Health Am. 2021, 3, 100037. [Google Scholar] [CrossRef]
- Kainz, K.; Bauer, M.A.; Madeo, F.; Carmona-Gutierrez, D. Fungal Infections in Humans: The Silent Crisis. Microb. Cell 2020, 7, 143–145. [Google Scholar] [CrossRef]
- Rodrigues, M.L.; Nosanchuk, J.D. Fungal Diseases as Neglected Pathogens: A Wake-up Call to Public Health Officials. PLoS Negl. Trop. Dis. 2020, 14, e0007964. [Google Scholar] [CrossRef]
- Berman, J.; Krysan, D.J. Drug Resistance and Tolerance in Fungi. Nat. Rev. Microbiol. 2020, 18, 319–331. [Google Scholar] [CrossRef]
- Garfoot, A.L.; Rappleye, C.A. Histoplasma Capsulatum Surmounts Obstacles to Intracellular Pathogenesis. FEBS J. 2016, 283, 619–633. [Google Scholar] [CrossRef] [PubMed]
- Pitangui, N.S.; Sardi, J.C.O.; Silva, J.F.; Benaducci, T.; Moraes da Silva, R.A.; Rodríguez-Arellanes, G.; Taylor, M.L.; Mendes-Giannini, M.J.S.; Fusco-Almeida, A.M. Adhesion of Histoplasma capsulatum to Pneumocytes and Biofilm Formation on an Abiotic Surface. Biofouling 2012, 28, 711–718. [Google Scholar] [CrossRef]
- Brilhante, R.S.N.; de Lima, R.A.C.; de Farias Marques, F.J.; Silva, N.F.; Caetano, É.P.; de Souza Collares Maia Castelo-Branco, D.; de Jesus Pinheiro Gomes Bandeira, T.; Moreira, J.L.B.; de Aguiar Cordeiro, R.; Monteiro, A.J. Histoplasma capsulatum in Planktonic and Biofilm Forms: In Vitro Susceptibility to Amphotericin B, Itraconazole and Farnesol. J. Med. Microbiol. 2015, 64, 394–399. [Google Scholar] [CrossRef]
- Brilhante, R.S.N.; Sales, J.A.; da Silva, M.L.Q.; de Oliveira, J.S.; de Alencar Pereira, L.; Pereira-Neto, W.A.; de Aguiar Cordeiro, R.; Sidrim, J.J.C.; de Souza Collares Maia Castelo-Branco, D.; Rocha, M.F.G. Antifungal Susceptibility and Virulence of Candida Parapsilosis Species Complex: An Overview of Their Pathogenic Potential. J. Med. Microbiol. 2018, 67, 903–914. [Google Scholar] [CrossRef] [PubMed]
- de Cássia Orlandi Sardi, J.; de Souza Pitangui, N.; Voltan, A.R.; Braz, J.D.; Machado, M.P.; Fusco Almeida, A.M.; Mendes Giannini, M.J.S. In Vitro Paracoccidioides Brasiliensis Biofilm and Gene Expression of Adhesins and Hydrolytic Enzymes. Virulence 2015, 6, 642–651. [Google Scholar] [CrossRef]
- Costa-Orlandi, C.B.; Sardi, J.C.O.; Pitangui, N.S.; de Oliveira, H.C.; Scorzoni, L.; Galeane, M.C.; Medina-Alarcón, K.P.; Melo, W.C.M.A.; Marcelino, M.Y.; Braz, J.D. Fungal Biofilms and Polymicrobial Diseases. J. Fungi 2017, 3, 22. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Mishra, S.; Singh, S. Advanced Acuity in Microbial Biofilm Genesis, Development, Associated Clinical Infections and Control. J. Anti-Infect. 2017, 19, 20–31. [Google Scholar] [CrossRef]
- Sardi, J.D.C.O.; Pitangui, N.D.S.; Rodriguez-Arellanes, G.; Taylor, M.L.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. Highlights in Pathogenic Fungal Biofilms. Rev. Iberoam. Micol. 2014, 31, 22–29. [Google Scholar] [CrossRef]
- Brilhante, R.S.N.; Aguiar, F.R.M.; Silva, M.L.Q.; Oliveira, J.S.; Camargo, Z.P.; Rodrigues, A.M.; Pereira, V.S.; Serpa, R.; de Souza Collares Maia Castelo-Branco, D.; Correia, E.E.M.; et al. Antifungal Susceptibility of Sporothrix schenckii Complex Biofilms. Med. Mycol. 2018, 56, 297–306. [Google Scholar] [CrossRef]
- de Aguiar Cordeiro, R.; Serpa, R.; Alexandre, C.F.U.; de Farias Marques, F.J.; de Melo, C.V.S.; da Silva Franco, J.; de Jesus Evangelista, A.J.; de Camargo, Z.P.; Brilhante, R.S.N.; Rocha, M.F.G. Trichosporon Inkin Biofilms Produce Extracellular Proteases and Exhibit Resistance to Antifungals. J. Med. Microbiol. 2015, 64, 1277–1286. [Google Scholar] [CrossRef]
- Brilhante, R.S.N.; Correia, E.E.M.; de Melo Guedes, G.M.; Pereira, V.S.; de Oliveira, J.S.; Bandeira, S.P.; de Alencar, L.P.; de Andrade, A.R.C.; de Souza Collares Maia Castelo-Branco, D.; de Aguiar Cordeiro, R.; et al. Quantitative and Structural Analyses of the in Vitro and Ex Vivo Biofilm-Forming Ability of Dermatophytes. J. Med. Microbiol. 2017, 66, 1045–1052. [Google Scholar] [CrossRef]
- Roudbary, M.; Vahedi-Shahandashti, R.; dos Santos, A.L.S.; Roudbar Mohammadi, S.; Aslani, P.; Lass-Flörl, C.; Rodrigues, C.F. Biofilm Formation in Clinically Relevant Filamentous Fungi: A Therapeutic Challenge. Crit. Rev. Microbiol. 2022, 48, 197–221. [Google Scholar] [CrossRef]
- Azar, M.M.; Loyd, J.L.; Relich, R.F.; Wheat, L.J.; Hage, C.A. Current Concepts in the Epidemiology, Diagnosis, and Management of Histoplasmosis Syndromes. Semin. Respir. Crit. Care Med. 2020, 41, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.-L.; Dai, Q.-Q.; Li, G.-B. Deep Learning in Target Prediction and Drug Repositioning: Recent Advances and Challenges. Drug Discov. Today 2021, 27, 1796–1814. [Google Scholar] [CrossRef] [PubMed]
- Garcia, L.G.S.; de Melo Guedes, G.M.; Fonseca, X.M.Q.C.; Pereira-Neto, W.A.; Castelo-Branco, D.S.C.M.; Sidrim, J.J.C.; de Aguiar Cordeiro, R.; Rocha, M.F.G.; Vieira, R.S.; Brilhante, R.S.N. Antifungal Activity of Different Molecular Weight Chitosans against Planktonic Cells and Biofilm of Sporothrix brasiliensis. Int. J. Biol. Macromol. 2020, 143, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Kvasničková, E.; Paulíček, V.; Paldrychová, M.; Ježdík, R.; Maťátková, O.; Masák, J. Aspergillus fumigatus DBM 4057 Biofilm Formation Is Inhibited by Chitosan, in Contrast to Baicalein and Rhamnolipid. World J. Microbiol. Biotechnol. 2016, 32, 187. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Moya, F.; Suarez-Fernandez, M.; Lopez-Llorca, L.V. Molecular Mechanisms of Chitosan Interactions with Fungi and Plants. Int. J. Mol. Sci. 2019, 20, 332. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017; ISBN 1562388304. [Google Scholar]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, 2nd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020; ISBN 978-1-68440-083-6. [Google Scholar]
- Brilhante, R.S.N.; Lopes, R.G.P.; de Aguiar, L.; de Oliveira, J.S.; dos Santos Araújo, G.; Paixão, G.C.; de Aquino Pereira-Neto, W.; Freire, R.S.; Nunes, J.V.S.; de Lima, R.P.; et al. Inhibitory Effect of Proteinase K against Dermatophyte Biofilms: An Alternative for Increasing the Antifungal Effects of Terbinafine and Griseofulvin. Biofouling 2022, 38, 286–297. [Google Scholar] [CrossRef]
- Pierce, C.G.; Uppuluri, P.; Tristan, A.R.; Wormley, F.L., Jr.; Mowat, E.; Ramage, G.; Lopez-Ribot, J.L. A Simple and Reproducible 96-Well Plate-Based Method for the Formation of Fungal Biofilms and Its Application to Antifungal Susceptibility Testing. Nat. Protoc. 2008, 3, 1494. [Google Scholar] [CrossRef]
- Almeida-Paes, R.; Figueiredo-Carvalho, M.H.G.; Brito-Santos, F.; Almeida-Silva, F.; Oliveira, M.M.E.; Zancope-Oliveira, R.M. Melanins Protect Sporothrix brasiliensis and Sporothrix schenckii from the Antifungal Effects of Terbinafine. PLoS ONE 2016, 11, 1–13. [Google Scholar] [CrossRef]
- Odds, F.C. Synergy, Antagonism, and What the Chequerboard Puts between Them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef]
- Madu, U.L.; Ogundeji, A.O.; Folorunso, O.S.; Albertyn, J.; Pohl, C.H.; Sebolai, O.M. The Repurposing of the Antimalaria Drug, Primaquine, as a Photosensitizer to Inactivate Cryptococcal Cells. Photochem 2021, 1, 275–286. [Google Scholar] [CrossRef]
- Pearce, A.N.; Chen, D.; Edmeades, L.R.; Cadelis, M.M.; Troudi, A.; Brunel, J.M.; Bourguet-Kondracki, M.-L.; Copp, B.R. Repurposing Primaquine as a Polyamine Conjugate to Become an Antibiotic Adjuvant. Bioorg. Med. Chem. 2021, 38, 116110. [Google Scholar] [CrossRef]
- Brilhante, R.S.N.; de Melo Guedes, G.M.; Riello, G.B.; Ribeiro, J.F.; Alencar, L.P.; Bandeira, S.P.; Castelo-Branco, D.S.C.M.; Oliveira, J.S.; Freire, J.M.M.; de Mesquita, J.R.L. RYP1 Gene as a Target for Molecular Diagnosis of Histoplasmosis. J. Microbiol. Methods 2016, 130, 112–114. [Google Scholar] [CrossRef] [PubMed]
- Bongomin, F.; Kwizera, R.; Baluku, J.B.; Asio, L.G.; Otu, A.A. Treatment of Histoplasmosis. In Histoplasma and Histoplasmosis; IntechOpen: London, UK, 2020; p. 69. [Google Scholar]
- Brilhante, R.S.N.; de Melo Guedes, G.M.; da Silva, M.L.Q.; Castelo-Branco, D.S.C.M.; de Aguiar Cordeiro, R.; Sidrim, J.J.C.; Rocha, M.F.G. A Proposal for Antifungal Epidemiological Cut-off Values against Histoplasma capsulatum Var. Capsulatum Based on the Susceptibility of Isolates from HIV-Infected Patients with Disseminated Histoplasmosis in Northeast Brazil. Int. J. Antimicrob. Agents 2018, 52, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, C.; Ferreira, N.; Lourenço, L. Production of Low Molecular Weight Chitosan and Chitooligosaccharides (COS): A Review. Polymers 2021, 13, 2466. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, H.S.; Yadav, P.N. Anticancer Activity of Chitosan, Chitosan Derivatives, and Their Mechanism of Action. Int. J. Biomater. 2018, 2018, 2952085. [Google Scholar] [CrossRef] [PubMed]
- Grande-Tovar, C.D.; Chaves-López, C.; Serio, A.; Rossi, C.; Paparella, A. Chitosan Coatings Enriched with Essential Oils: Effects on Fungi Involved in Fruit Decay and Mechanisms of Action. Trends Food Sci. Technol. 2018, 78, 61–71. [Google Scholar] [CrossRef]
- Kulikov, S.N.; Lisovskaya, S.A.; Zelenikhin, P.V.; Bezrodnykh, E.A.; Shakirova, D.R.; Blagodatskikh, I.V.; Tikhonov, V.E. Antifungal Activity of Oligochitosans (Short Chain Chitosans) against Some Candida Species and Clinical Isolates of Candida albicans: Molecular Weight–Activity Relationship. Eur. J. Med. Chem. 2014, 74, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Garrido-Maestu, A.; Jeong, K.C. Application, Mode of Action, and in vivo Activity of Chitosan and Its Micro-and Nanoparticles as Antimicrobial Agents: A Review. Carbohydr. Polym. 2017, 176, 257–265. [Google Scholar] [CrossRef]
- Poznanski, P.; Hameed, A.; Orczyk, W. Chitosan and Chitosan Nanoparticles: Parameters Enhancing Antifungal Activity. Molecules 2023, 28, 2996. [Google Scholar] [CrossRef]
- Lopez-Moya, F.; Colom-Valiente, M.F.; Martinez-Peinado, P.; Martinez-Lopez, J.E.; Puelles, E.; Sempere-Ortells, J.M.; Lopez-Llorca, L.V. Carbon and Nitrogen Limitation Increase Chitosan Antifungal Activity in Neurospora crassa and Fungal Human Pathogens. Fungal Biol. 2015, 119, 154–169. [Google Scholar] [CrossRef]
- Valdez, A.F.; Miranda, D.Z.; Guimarães, A.J.; Nimrichter, L.; Nosanchuk, J.D. Pathogenicity & Virulence of Histoplasma capsulatum-A Multifaceted Organism Adapted to Intracellular Environments. Virulence 2022, 13, 2137987. [Google Scholar]
- Martínez-Álvarez, J.A.; Pérez-García, L.A.; Mellado-Mojica, E.; López, M.G.; Martínez-Duncker, I.; Lópes-Bezerra, L.M.; Mora-Montes, H.M. Sporothrix schenckii Sensu stricto and Sporothrix brasiliensis Are Differentially Recognized by Human Peripheral Blood Mononuclear Cells. Front. Microbiol. 2017, 8, 843. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Arango, A.C.; Rueda, C.; Román, E.; Quintin, J.; Terrón, M.C.; Luque, D.; Netea, M.G.; Pla, J.; Zaragoza, O. Cell Wall Changes in Amphotericin B-Resistant Strains from Candida tropicalis and Relationship with the Immune Responses Elicited by the Host. Antimicrob. Agents Chemother. 2016, 60, 2326–2335. [Google Scholar] [CrossRef] [PubMed]
- Lo, W.-H.; Deng, F.-S.; Chang, C.-J.; Lin, C.-H. Synergistic Antifungal Activity of Chitosan with Fluconazole against Candida albicans, Candida tropicalis, and Fluconazole-Resistant Strains. Molecules 2020, 25, 5114. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.D.; MacDougall, C.; Ostrosky-Zeichner, L.; Perfect, J.R.; Rex, J.H. Combination Antifungal Therapy. Antimicrob. Agents Chemother. 2004, 48, 693–715. [Google Scholar] [CrossRef] [PubMed]
- Srimaneepong, V.; Thanamee, T.; Wattanasirmkit, K.; Muangsawat, S.; Matangkasombut, O. Efficacy of Low-molecular Weight Chitosan against Candida albicans Biofilm on Polymethyl Methacrylate Resin. Aust. Dent. J. 2021, 66, 262–269. [Google Scholar] [CrossRef]
- Atai, Z.; Atai, M.; Amini, J. In Vivo Study of Antifungal Effects of Low-Molecular-Weight Chitosan against Candida albicans. J. Oral Sci. 2017, 59, 425–430. [Google Scholar] [CrossRef]
- Riddell IV, J.; Wheat, L.J. Central Nervous System Infection with Histoplasma capsulatum. J. Fungi 2019, 5, 70. [Google Scholar] [CrossRef]
- Horwath, M.C.; Fecher, R.A.; Deepe, G.S., Jr. Histoplasma capsulatum, Lung Infection and Immunity. Future Microbiol. 2015, 10, 967–975. [Google Scholar] [CrossRef]
- Arce Miranda, J.E.; Sotomayor, C.E.; Albesa, I.; Paraje, M.G. Oxidative and Nitrosative Stress in Staphylococcus Aureus Biofilm. FEMS Microbiol. Lett. 2011, 315, 23–29. [Google Scholar] [CrossRef]
- Arce Miranda, J.E.; Baronetti, J.L.; Sotomayor, C.E.; Paraje, M.G. Oxidative and Nitrosative Stress Responses during Macrophage–Candida Albicans Biofilm Interaction. Med. Mycol. 2019, 57, 101–113. [Google Scholar] [CrossRef]
- Pais, P.; Vagueiro, S.; Mil-Homens, D.; Pimenta, A.I.; Viana, R.; Okamoto, M.; Chibana, H.; Fialho, A.M.; Teixeira, M.C. A New Regulator in the Crossroads of Oxidative Stress Resistance and Virulence in Candida Glabrata: The Transcription Factor CgTog1. Virulence 2020, 11, 1522–1538. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-Y.; Chen, H.-F.; Yeh, Y.-C.; Xue, Y.-P.; Lan, C.-Y. The Transcription Factor Sfp1 Regulates the Oxidative Stress Response in Candida Albicans. Microorganisms 2019, 7, 131. [Google Scholar] [CrossRef] [PubMed]
- Caceres, I.; El Khoury, R.; Bailly, S.; Oswald, I.P.; Puel, O.; Bailly, J.-D. Piperine Inhibits Aflatoxin B1 Production in Aspergillus Flavus by Modulating Fungal Oxidative Stress Response. Fungal Genet. Biol. 2017, 107, 77–85. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Strains | Minimum Inhibitory Concentration (µg/mL) | ||
|---|---|---|---|
| (CEMM Code) | AMB | ITC | CHI |
| CEMM 03-2-088 | 2 | 1 | 64 |
| CEMM 03-2-090 | 0.5 | 0.25 | 32 |
| CEMM 03-3-003 | 0.25 | 1 | 64 |
| CEMM 03-3-026 | 2 | 1 | 64 |
| CEMM 03-3-033 | 2 | 0.25 | 64 |
| CEMM 03-3-034 | 0.5 | 0.25 | 32 |
| CEMM 03-3-036 | 2 | 1 | 64 |
| CEMM 03-3-037 | 2 | 1 | 64 |
| CEMM 03-3-038 | 2 | 1 | 128 |
| CEMM 03-3-039 | 0.5 | 0.03 | 64 |
| CEMM 03-3-015 | 1 | 0.25 | 32 |
| CEMM 03-3-072 | 2 | 1 | 64 |
| CEMM 03-3-100 | 0.5 | 0.03 | 64 |
| CEMM 03-3-024 | 0.5 | 0.03 | 128 |
| CEMM 03-3-049 | 0.35 | 1 | 32 |
| CEMM 03-3-066 | 2 | 0.25 | 32 |
| CEMM 03-3-052 | 1 | 0.25 | 32 |
| CEMM 03-3-070 | 0.5 | 1 | 64 |
| CEMM 03-3-040 | 1 | 1 | 64 |
| CEMM 03-3-053 | 0.5 | 0.03 | 32 |
| Strains | Minimum Inhibitory Concentration (µg/mL) | ||
|---|---|---|---|
| (CEMM Code) | AMB | ITC | CHI |
| CEMM 03-2-088 | 0.25 | 0.5 | 8 |
| CEMM 03-2-090 | 0.03 | 0.25 | 32 |
| CEMM 03-3-003 | 0.5 | 0.5 | 64 |
| CEMM 03-3-026 | 0.03 | 0.5 | 64 |
| CEMM 03-3-037 | 0.03 | 0.5 | 8 |
| CEMM 03-3-038 | 0.25 | 0.25 | 64 |
| CEMM 03-3-039 | 0.5 | 0.5 | 32 |
| CEMM 03-3-015 | 0.03 | 0.25 | 16 |
| CEMM 03-3-072 | 0.5 | 0.5 | 16 |
| CEMM 03-3-100 | 0.03 | 0.5 | 32 |
| CEMM 03-3-033 | 0.25 | 0.25 | 16 |
| CEMM 03-3-034 | 0.25 | 0.25 | 32 |
| CEMM 03-3-036 | 0.25 | 0.25 | 32 |
| CEMM Code | Isolated Drugs (µg/mL) | Combined Drugs (µg/mL) | FICI | ||||
|---|---|---|---|---|---|---|---|
| CHI | AMB | ITC | CHI/AMB | CHI/ITC | CHI/AMB | CHI/ITC | |
| CEMM 03-2-088 | 64 | 2 | 1 | 16/0.5 | 16/0.5 | 0,75 I | 1.25 I |
| CEMM 03-2-090 | 128 | 0.5 | 0.25 | 64/2 | 32/1 | 1.5 I | 0.75 I |
| CEMM 03-3-003 | 64 | 0.25 | 1 | 16/0.25 | 16/0.25 | 0.5 I | 0.75 I |
| CEMM 03-3-026 | 64 | 2 | 1 | 32/0.0125 | 32/0.125 | 0.625 I | 1.5 I |
| CEMM 03-3-033 | 64 | 2 | 4 | 32/1 | 16/0.25 | 1.5 I | 0.75 I |
| CEMM 03-3-034 | 64 | 0.5 | 0.25 | 16/2 | 16/0.125 | 1.25 I | 0.03 S |
| CEMM 03-3-036 | 64 | 2 | 1 | 16/2 | 16/0.25 | 1.25 I | 0.03 S |
| CEMM 03-3-037 | 32 | 2 | 1 | 32/1 | 32/0.25 | 2.0 I | 1.25 I |
| CEMM Code | Isolated Drugs (µg/mL) | Combined Drugs (µg/mL) | FICI | |||
|---|---|---|---|---|---|---|
| CHI | AMB | CHI/AMB | CHI/ITC | CHI/AMB | CHI/ITC | |
| CEMM 03-2-088 | 32 | 0.25 | 4/0.03125 | 2/0.0078125 | 0.125 S | 0.3125 S |
| CEMM 03-2-090 | 32 | 0.125 | 4/0.01625 | 2/0.0078125 | 0.125 S | 0.3125 S |
| CEMM 03-3-003 | 32 | 0.5 | 2/0.03125 | 2/0.0078125 | 0.125 S | 0.3125 S |
| CEMM 03-3-026 | 64 | 0.5 | 2/0.03125 | 2/0.0078125 | 0.125 S | 0.3125 S |
| CEMM 03-3-033 | 64 | 0.5 | 2/0.03125 | 2/0.0078125 | 0.125 S | 0.3125 S |
| CEMM 03-3-034 | 64 | 0.5 | 2/0.03125 | 2/0.0078125 | 0.125 S | 0.3125 S |
| CEMM 03-3-036 | 32 | 0.5 | 2/0.03125 | 2/0.0078125 | 0.125 S | 0.3125 S |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brilhante, R.S.N.; Costa, A.d.C.; Mesquita, J.R.L.d.; dos Santos Araújo, G.; Freire, R.S.; Nunes, J.V.S.; Nobre, A.F.D.; Fernandes, M.R.; Rocha, M.F.G.; Pereira Neto, W.d.A.; et al. Antifungal Activity of Chitosan against Histoplasma capsulatum in Planktonic and Biofilm Forms: A Therapeutic Strategy in the Future? J. Fungi 2023, 9, 1201. https://doi.org/10.3390/jof9121201
Brilhante RSN, Costa AdC, Mesquita JRLd, dos Santos Araújo G, Freire RS, Nunes JVS, Nobre AFD, Fernandes MR, Rocha MFG, Pereira Neto WdA, et al. Antifungal Activity of Chitosan against Histoplasma capsulatum in Planktonic and Biofilm Forms: A Therapeutic Strategy in the Future? Journal of Fungi. 2023; 9(12):1201. https://doi.org/10.3390/jof9121201
Chicago/Turabian StyleBrilhante, Raimunda Sâmia Nogueira, Anderson da Cunha Costa, Jacó Ricarte Lima de Mesquita, Gessica dos Santos Araújo, Rosemeyre Souza Freire, João Victor Serra Nunes, Augusto Feynman Dias Nobre, Mirele Rodrigues Fernandes, Marcos Fábio Gadelha Rocha, Waldemiro de Aquino Pereira Neto, and et al. 2023. "Antifungal Activity of Chitosan against Histoplasma capsulatum in Planktonic and Biofilm Forms: A Therapeutic Strategy in the Future?" Journal of Fungi 9, no. 12: 1201. https://doi.org/10.3390/jof9121201