
Regulatory Ability of Lactiplantibacillus plantarum on Human Skin Health by Counteracting In Vitro Malassezia furfur Effects
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.2. Microorganisms and Culture Media
2.3. Phospholipase Assay
2.4. Cell Treatments
2.5. Real-Time PCR
2.6. ELISA Assay
2.7. Scratch Wound Healing Assay
2.8. Statistical Analysis
3. Results
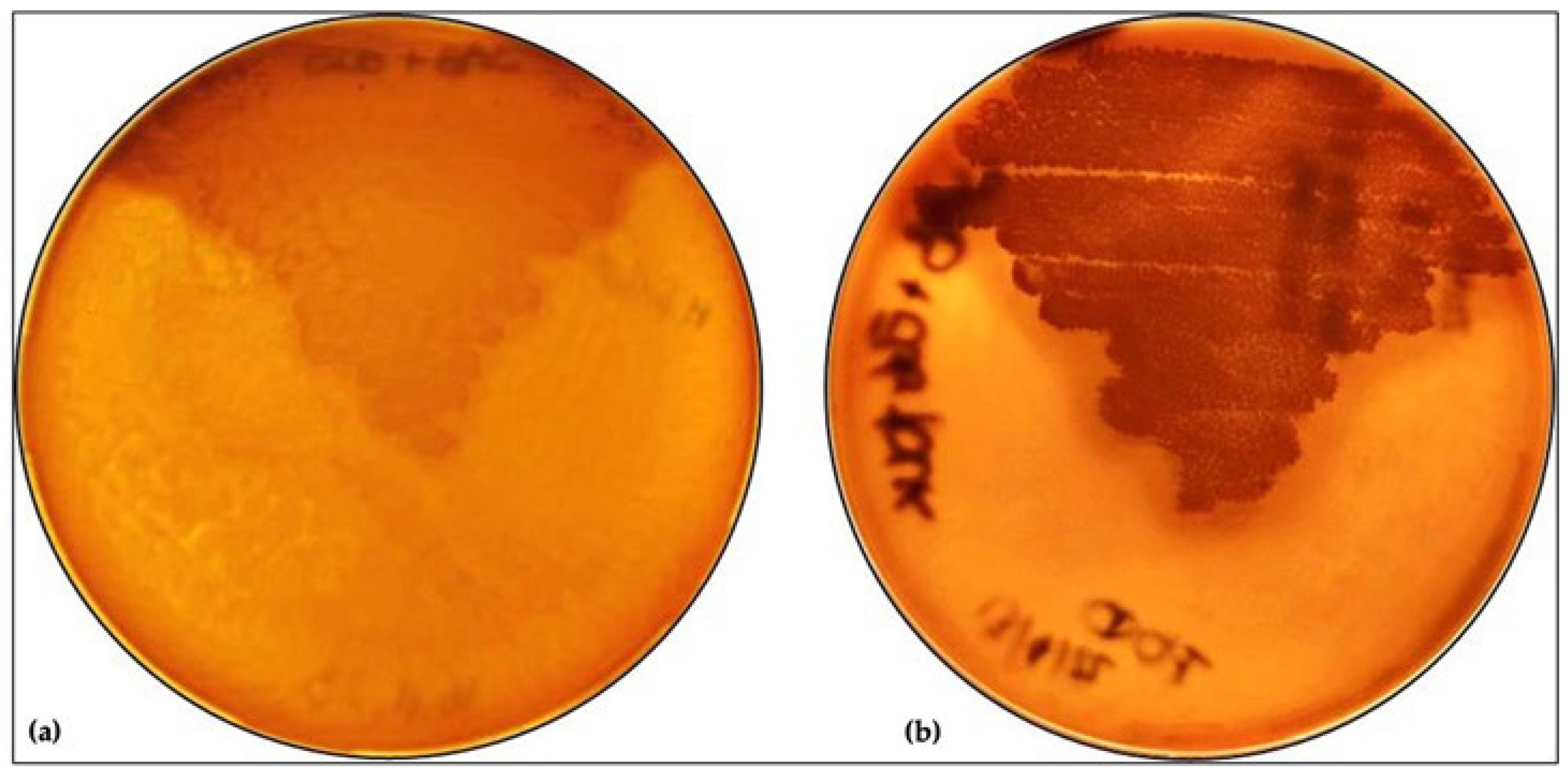
3.1. Effect of L. plantarum on M. furfur Phospholipase Activity
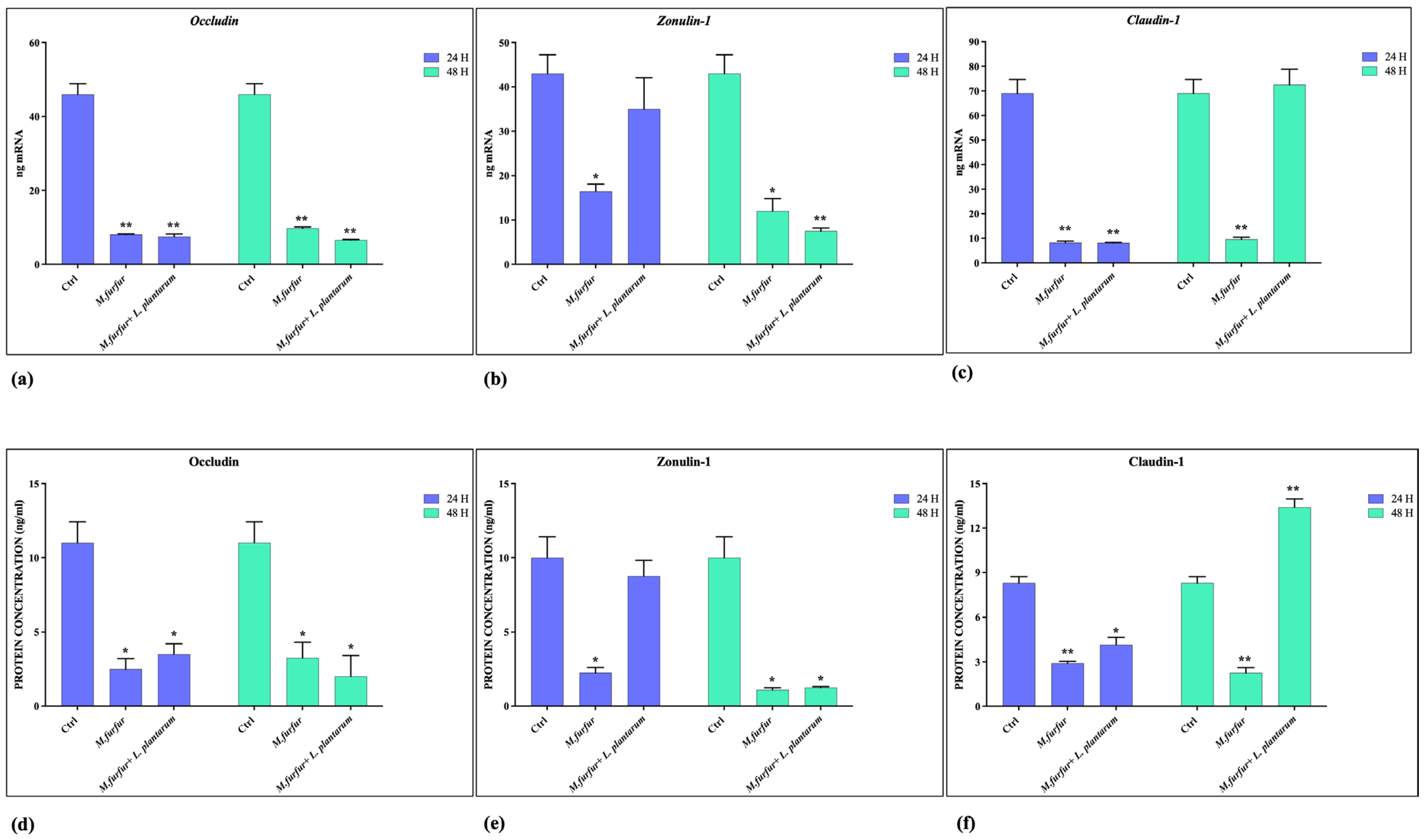
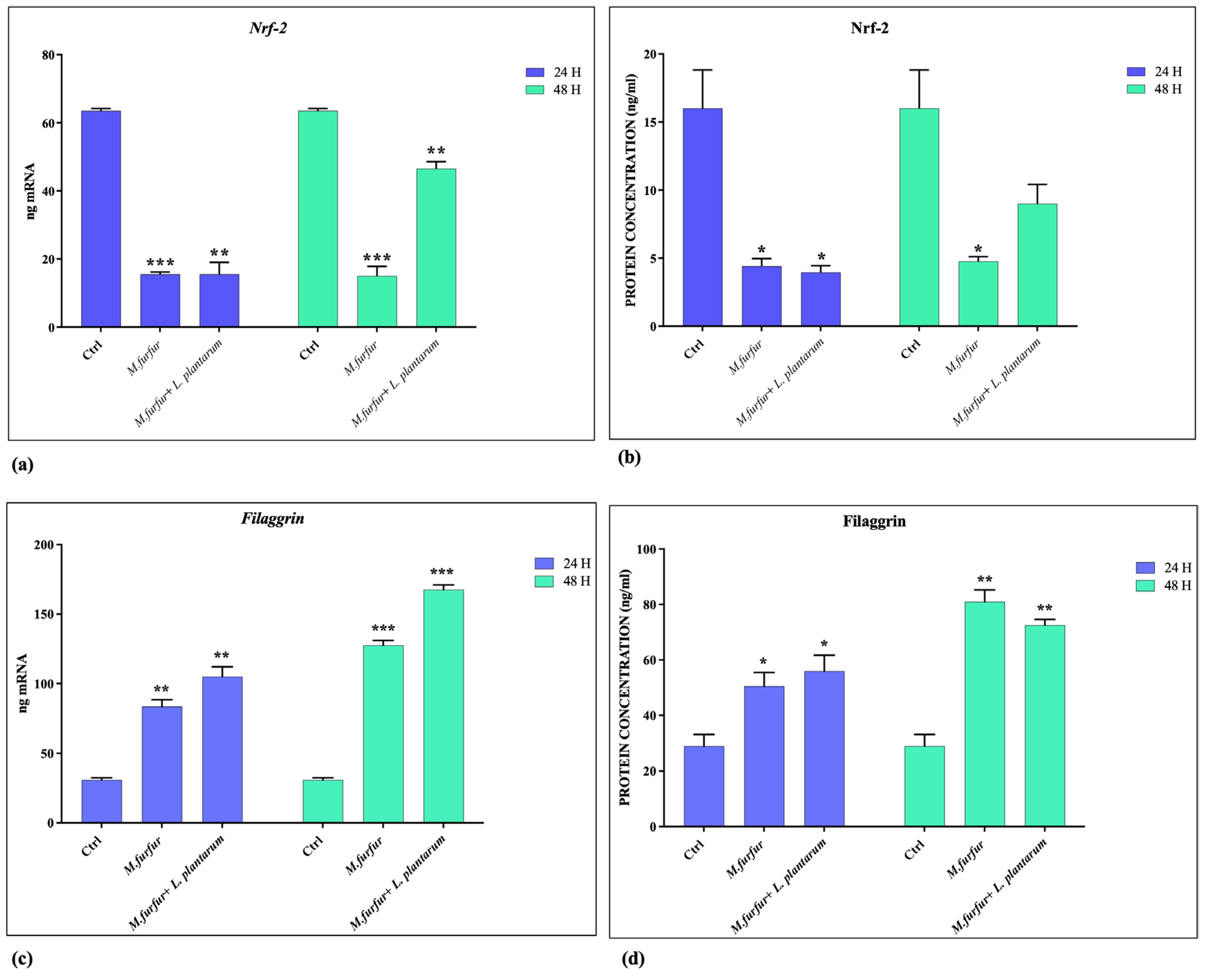
3.2. Effect of L. plantarum on Barrier Integrity
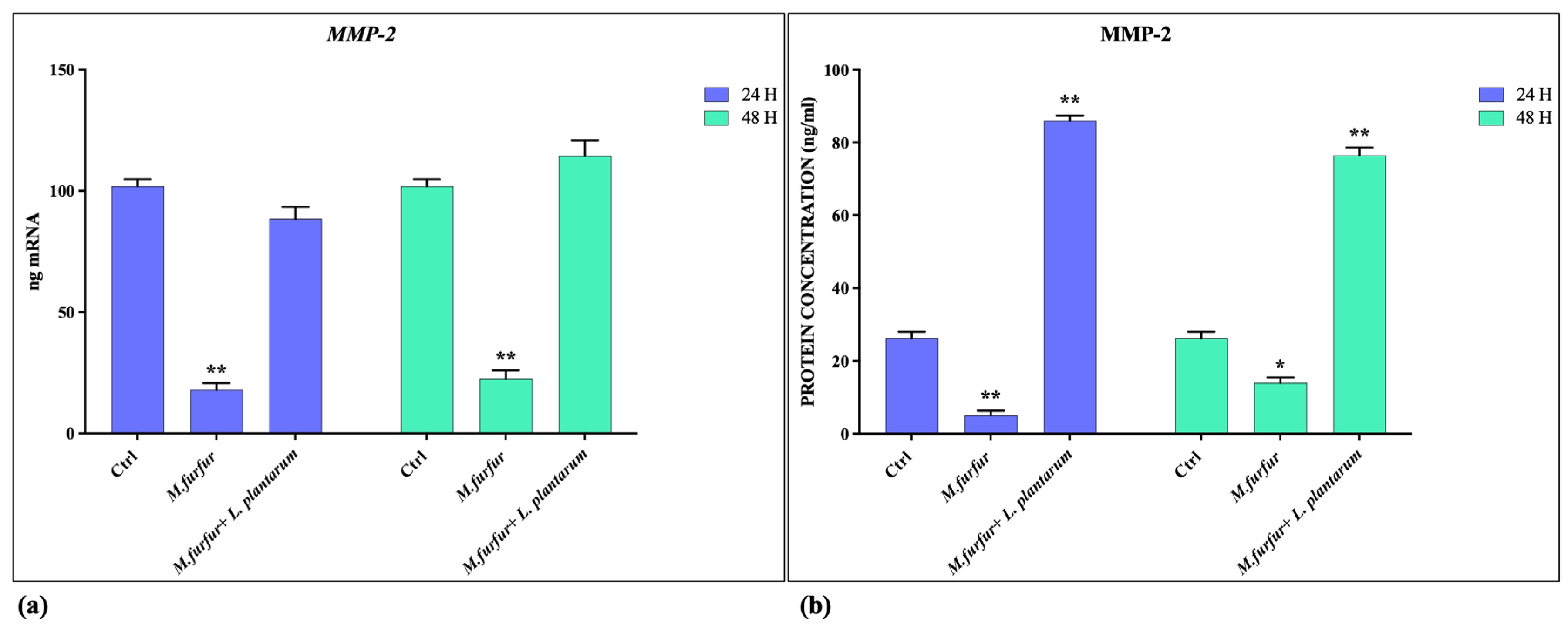
3.3. Tissue Repair Activity of L. plantarum
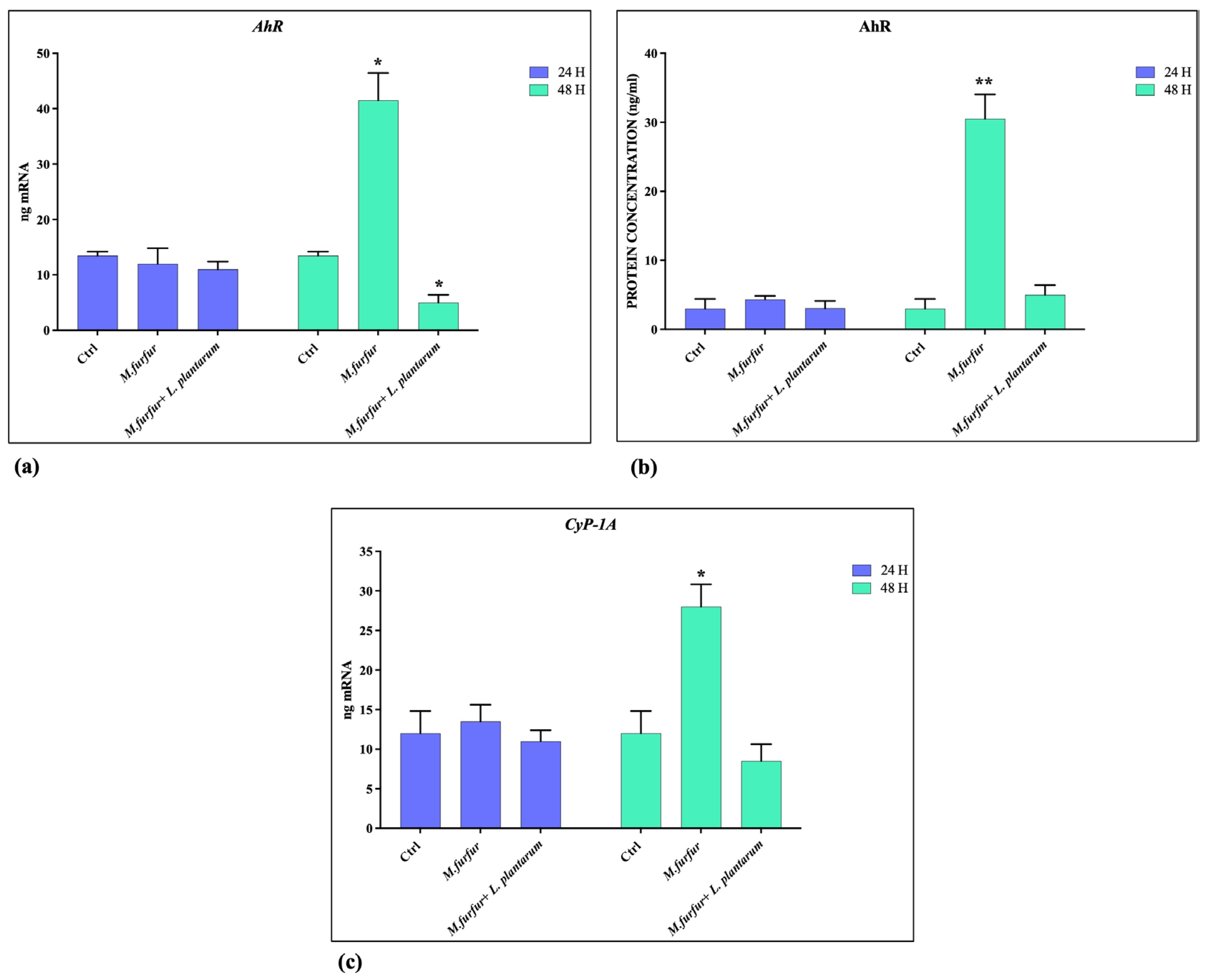
3.4. Activation of AhR Pathways
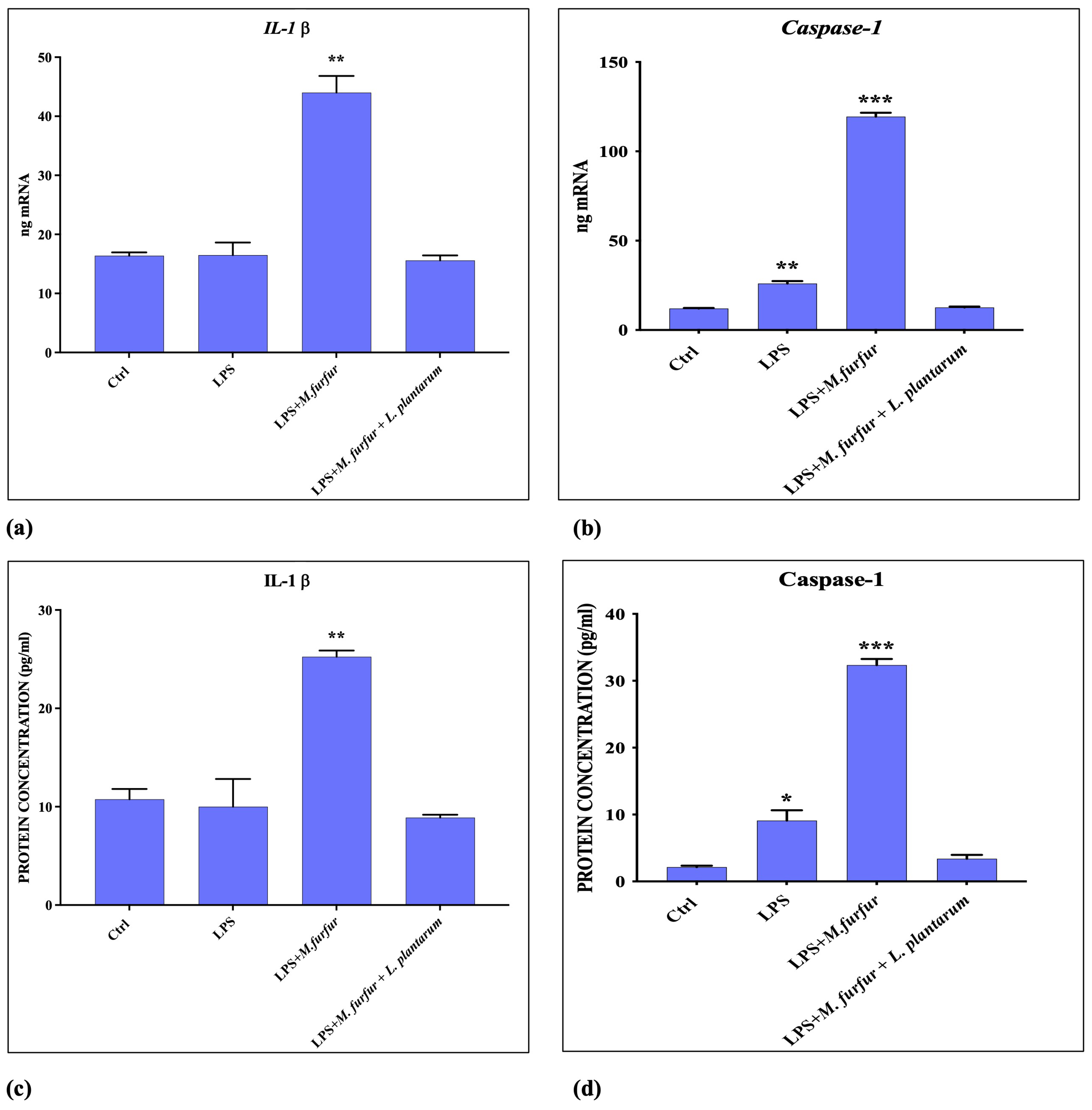
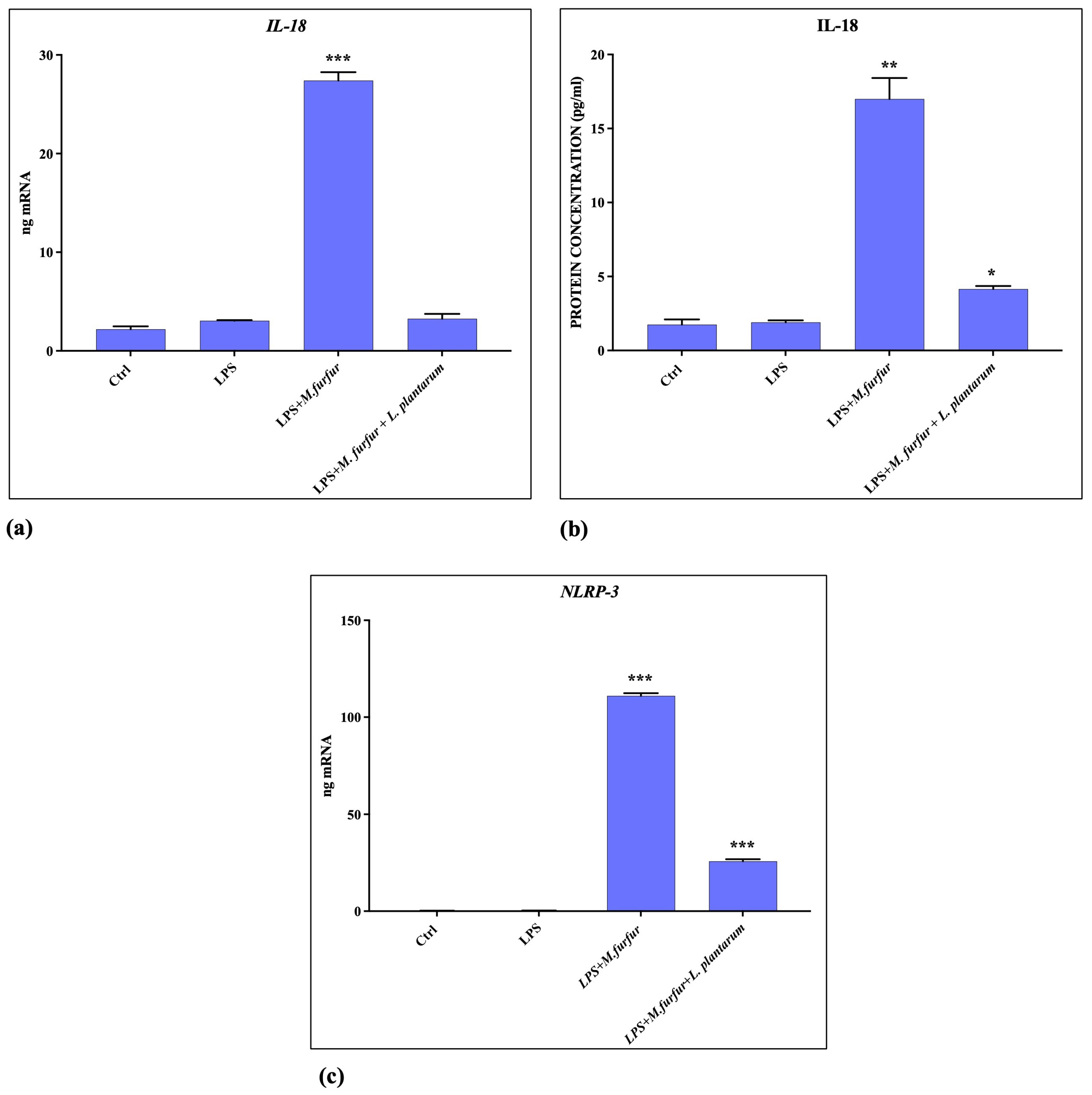
3.5. Inflammasome Activation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Braun, C.; Patra, V.; Lina, G.; Nicolas, J.F.; Vocanson, M.; Nosbaum, A. The role of skin dysbiosis in atopic dermatitis. Eur. J. Dermatol. 2022, 32, 439–444. [Google Scholar]
- Carmona-Cruz, S.; Orozco-Covarrubias, L.; Sáez-de-Ocariz, M. The Human Skin Microbiome in Selected Cutaneous Diseases. Front. Cell. Infect. Microbiol. 2022, 12, 834135. [Google Scholar] [CrossRef]
- Yan, F.; Polk, D.B. Probiotics and Probiotic-Derived Functional Factors-Mechanistic Insights into Applications for Intestinal Homeostasis. Front. Immunol. 2020, 11, 428. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, Z.; Tang, P.; Wu, Y.; Zhang, A.; Li, D.; Wang, C.Z.; Wan, J.Y.; Yao, H.; Yuan, C.S. Probiotics fortify intestinal barrier function: A systematic review and meta-analysis of randomized trials. Front. Immunol. 2023, 14, 1143548. [Google Scholar] [CrossRef]
- Brandi, J.; Cheri, S.; Manfredi, M.; Di Carlo, C.; Vita Vanella, V.; Federici, F.; Bombiero, E.; Bazaj, A.; Rizzi, E.; Manna, L.; et al. Exploring the wound healing, anti-inflammatory, anti-pathogenic and proteomic effects of lactic acid bacteria on keratinocytes. Sci. Rep. 2020, 10, 11572. [Google Scholar] [CrossRef]
- Lombardi, F.; Palumbo, P.; Mattei, A.; Augello, F.R.; Cifone, M.G.; Giuliani, M.; Cinque, B. Soluble Fraction from Lysates of Selected Probiotic Strains Differently Influences Re-Epithelialization of HaCaT Scratched Monolayer through a Mechanism Involving Nitric Oxide Synthase 2. Biomolecules 2019, 9, 756. [Google Scholar] [CrossRef]
- Li, W.; Zhang, Z.W.; Luo, Y.; Liang, N.; Pi, X.X.; Fan, Y.M. Molecular epidemiology, in vitro susceptibility and exoenzyme screening of Malassezia clinical isolates. J. Med. Microbiol. 2020, 69, 436–442. [Google Scholar] [CrossRef]
- Hamdino, M.; Saudy, A.A.; El-Shahed, L.H.; Taha, M. Identification of Malassezia species isolated from some Malassezia associated skin diseases. J. Mycol. Med. 2022, 32, 101301. [Google Scholar] [CrossRef]
- Ashbee, H.R. Update on the genus Malassezia. Med. Mycol. 2007, 45, 287–303. [Google Scholar] [CrossRef]
- Baroni, A.; Orlando, M.; Donnarumma, G.; Farro, P.; Iovene, M.R.; Tufano, M.A.; Buommino, E. Toll-like receptor 2 (TLR2) mediates intracellular signaling in human keratinocytes in response to Malassezia furfur. Arch. Dermatol. Res. 2006, 297, 280–288. [Google Scholar] [CrossRef]
- Brasch, J.; Morig, A.; Neumann, B.; Proksch, E. Expression of antimicrobial peptides and toll-like receptors is increased in tinea and pityriasis versicolor. Mycoses 2014, 57, 147–152. [Google Scholar] [CrossRef]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.J.; Wells, J.M. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef]
- Vera, L.; Caley, M.; O’Toole, E.A. Matrix metalloproteinases and epidermal wound repair. Cell Tissue Res. 2013, 351, 255–268. [Google Scholar]
- Oku, N.; Sasabe, E.; Ueta, E.; Yamamoto, T.; Osaki, T. Tight junction protein claudin-1 enhances the invasive activity of oral squamous cell carcinoma cells by promoting cleavage of laminin-5 gamma2 chain via matrix metalloproteinase (MMP)-2 and membrane-type MMP-1. Cancer Res. 2006, 66, 5251–5257. [Google Scholar] [CrossRef]
- Fusco, A.; Savio, V.; Cammarota, M.; Donnarumma, G.; Baroni, A. Decreased expression of Malassezia furfur virulence factors after Q-switched Nd:YAG laser irradiation. Eur. J. Dermatol. 2021, 17, 470–472. [Google Scholar] [CrossRef]
- Tee, C.B.; Sei, Y.; Kajiwara, S. Secreted Hydrolytic and Haemolytic Activities of Malassezia Clinical Strains. Mycopathologia 2019, 184, 227–238. [Google Scholar] [CrossRef]
- Sato, Y.; Fujimura, T.; Tanita, K.; Chunbing, L.; Matsushita, S.; Fujisawa, Y.; Otsuka, A.; Yamamoto, Y.; Hidaka, T.; Aiba, S. Malassezia-derived aryl hydrocarbon receptor ligands enhance the CCL20/Th17/soluble CD163 pathogenic axis in extra-mammary Paget’s disease. Exp. Dermatol. 2019, 28, 933–939. [Google Scholar] [CrossRef]
- Furue, M. Regulation of Filaggrin, Loricrin, and Involucrin by IL-4, IL-13, IL-17A, IL-22, AHR, and NRF2: Pathogenic Implications in Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 5382. [Google Scholar] [CrossRef]
- Buommino, E.; Baroni, A.; Papulino, C.; Nocera, F.P.; Coretti, L.; Donnarumma, G.; De Filippis, A.; De Martino, L. Malassezia pachydermatis up-regulates AhR related CYP1A1 gene and epidermal barrier markers in human keratinocytes. Med. Mycol. 2018, 56, 987–993. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.; Xia, P.; Zhang, R.; Wu, Y.; Xia, J.; Su, G.; Zhang, J.; Giesy, J.P.; Wang, Z.; et al. Activation of AhR-mediated toxicity pathway by emerging pollutants polychlorinated diphenyl sulfides. Chemosphere 2016, 144, 1754–1762. [Google Scholar] [CrossRef]
- Park, H.R.; Oh, J.H.; Lee, Y.J.; Park, S.H.; Lee, Y.W.; Lee, S.; Kang, H.; Kim, J.E. Inflammasome-mediated Inflammation by Malassezia in human keratinocytes: A comparative analysis with different strains. Mycoses 2021, 64, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Kistowska, M.; Fenini, G.; Jankovic, D.; Feldmeyer, L.; Kerl, K.; Bosshard, P.; Contassot, E.; French, L.E. Malassezia yeasts activate the NLRP3 inflammasome in antigen- presenting cells via Syk-kinase signalling. Exp. Dermatol. 2014, 23, 884–889. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A.J.; Limon, J.J.; Nguyen, C.; Prince, A.; Castro, A.; Underhill, D.M. Malassezia spp. induce inflammatory cytokines and activate NLRP3 inflammasomes in phagocytes. J. Leukoc. Biol. 2021, 109, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.H.; Chou, C.H.; Chiang, Y.J.; Lin, C.G.; Lee, C.H. Regulatory effects of Lactobacillus plantarum GMNL6 on human skin health by improving skin microbiome. Int. J. Med. Sci. 2021, 18, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Tsuji, G.; Hashimoto-Hachiya, A.; Furue, M. Galactomyces Ferment Filtrate Potentiates an Anti-Inflammaging System in Keratinocytes. J. Clin. Med. 2022, 11, 6338. [Google Scholar] [CrossRef] [PubMed]
- Ong, J.S.; Taylor, T.D.; Yong, C.C.; Khoo, B.Y.; Sasidharan, S.; Choi, S.B.; Ohno, H.; Liong, M.T. Lactobacillus plantarum USM8613 Aids in Wound Healing and Suppresses Staphylococcus aureus Infection at Wound Sites. Probiotics Antimicrob. Proteins 2020, 12, 125–137. [Google Scholar] [CrossRef]
- Lolou, V.; Panayiotidis, M.I. Functional Role of Probiotics and Prebiotics on Skin Health and Disease. Fermentation 2019, 5, 41. [Google Scholar] [CrossRef]
- Jeong, J.H.; Lee, C.Y.; Chung, D.K. Probiotic Lactic Acid Bacteria and Skin Health. Crit. Rev. Food. Sci. Nutr. 2016, 56, 2331–2337. [Google Scholar] [CrossRef]
- Jain, C.; Das, S.; Saha, R.; Ramachandran, V.G.; Bhattacharya, S.N.; Dar, S. Detection of phospholipase production by egg yolk-agar in Malassezia isolates from diseased and healthy human host. Int. J. Med. Sci. Public Health 2017, 6, 1–5. [Google Scholar] [CrossRef]
- Vlachos, C.; Gaitanis, G.; Alexopoulos, E.C.; Papadopoulou, C.; Bassukas, I.D. Phospholipase activity after β-endorphin exposure discriminates Malassezia strains isolated from healthy and seborrhoeic dermatitis skin. J. Eur. Acad. Dermatol. Venereol. 2013, 27, 1575–1578. [Google Scholar] [CrossRef]
- Cafarchia, C.; Gasser, R.B.; Latrofa, M.S.; Parisi, A.; Campbell, B.E.; Otranto, D. Genetic variants of Malassezia pachydermatis from canine skin: Body distribution and phospholipase activity. FEMS Yeast Res. 2008, 8, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Fusco, A.; Savio, V.; Cimini, D.; D’Ambrosio, S.; Chiaromonte, A.; Schiraldi, C.; Donnarumma, G. In Vitro Evaluation of the Most Active Probiotic Strains Able to Improve the Intestinal Barrier Functions and to Prevent Inflammatory Diseases of the Gastrointestinal System. Biomedicines 2023, 11, 865. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Oerlemans, E.F.M.; Claes, I.; Henkens, T.; Delanghe, L.; Wuyts, S.; Spacova, I.; van den Broek, M.F.L.; Tuyaerts, I.; Wittouck, S.; et al. Selective targeting of skin pathobionts and inflammation with topically applied lactobacilli. Cell Rep. Med. 2022, 15, 100521. [Google Scholar] [CrossRef] [PubMed]
- Vijaya Chandra, S.H.; Srinivas, R.; Dawson, T.L., Jr.; Common, J.E. Cutaneous Malassezia: Commensal, Pathogen, or Protector? Front. Cell. Infect. Microbiol. 2021, 10, 614446. [Google Scholar] [CrossRef] [PubMed]
- Angrisano, T.; Pero, R.; Paoletti, I.; Keller, S.; Lembo, L.; Baroni, A.; Chiariotti, L.; Lembo, F.; Donnarumma, G. Epigenetic regulation of IL-8 and β-defensin genes in human keratinocytes in response to Malassezia furfur. J. Investig. Dermatol. 2013, 133, 2101–2104. [Google Scholar] [CrossRef]
- Sparber, F.; Ruchti, F.; LeibundGut-Landmann, S. Host Immunity to Malassezia in Health and Disease. Front. Cell. Infect. Microbiol. 2020, 10, 198. [Google Scholar] [CrossRef]
- Sparber, F.; LeibundGut-Landmann, S. Host Responses to Malassezia spp. in the Mammalian Skin. Front. Immunol. 2017, 8, 1614. [Google Scholar] [CrossRef]
- Buommino, E.; Nocera, F.P.; Parisi, A.; Rizzo, A.; Donnarumma, G.; Mallardo, K.; Fiorito, F.; Baroni, A.; De Martino, L. Correlation between genetic variability and virulence factors in clinical strains of Malassezia pachydermatis of animal origin. New Microbiol. 2016, 39, 216–223. [Google Scholar]
- Fernández-Gallego, N.; Sánchez-Madrid, F.; Cibrian, D. Role of AHR Ligands in Skin Homeostasis and Cutaneous Inflammation. Cells 2021, 10, 3176. [Google Scholar] [CrossRef]
- Antushevich, H. Interplays between inflammasomes and viruses, bacteria (pathogenic and probiotic), yeasts and parasites. Immunol. Lett. 2020, 228, 1–14. [Google Scholar] [CrossRef]
- Lee, Y.B.; Byun, E.J.; Kim, H.S. Potential Role of the Microbiome in Acne: A Comprehensive Review. J. Clin. Med. 2019, 8, 987. [Google Scholar] [CrossRef] [PubMed]
- França, K. Topical Probiotics in Dermatological Therapy and Skincare: A Concise Review. Dermatol. Ther. 2021, 11, 71–77. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer’s Sequence | Conditions | Product (bp) |
|---|---|---|---|
| AhR | 5′-ACCTACGCCAGTCGCAA-3′ 5′-CTGACGCTGAGCCTAAGAAC-3′ | 30″ at 95 °C, 30″ at 60 °C, 30″ at 72 °C for 40 cycles | 200 |
| CyP1A1 | 5′-TCCAGAGACAACAGGTAAAACA-3′ 5′-AGGAAGGGCAGAGGAATGTGAT-3′ | 15″ at 95 °C, 35″ at 60 °C, 35″ at 72 °C for 45 cycles | 371 |
| Occludin | 5′-TCAGGGAATATCCACCTATCACTTCAG-3′ 5′-CATCAGCAGCAGCCATGTACTCTTCAC-3′ | 10″ at 95 °C, 45″ at 60 °C for 40 cycles | 188 |
| Zonulin-1 | 5′-AGGGGCAGTGGTGGTTTTCTGTTCTTTC-3′ 5′-GCAGAGGTCAAAGTTCAAGGCTCAAGAGG-3′ | 10″ at 95 °C, 45″ at 60 °C for 40 cycles | 217 |
| Claudin-1 | 5′-CTGGGAGGTGCCCTACTTTG-3′ 5′-ACACGTAGTCTTTCCCGCTG-3′ | 1″ at 95 °C, 30″ at 60 °C, 20″ at 72 °C for 40 cycles | 128 |
| NrF2 | 5′-ACCACCCACAACTTACTGCC-3′ 5′-GCCATAGGAGTATGGGGGAT-3′ | 5″ at 95 °C, 2″ at 60 °C, 5″ at 72 °C for 40 cycles | 121 |
| MMP-2 | 5′-TGACGGTAAGGACGGACTC-3′ 5′-TGGAAGCGGATTGGAAACT-3′ | 5″ at 94 °C, 7″ at 57 °C, 14″ at 72 °C for 40 cycles | 342 |
| Caspase-1 | 5′-GCCTGTTCCTGTGATGTGGAG-3′ 5′-TGCCCACAGACATTCATACAGTTTC-3′ | 15″ at 95 °C, 1′ at 60 °C for 40 cycles | 165 |
| IL-1 β | 5′-GCATCCAGCTACGAATCTCC-3′ 5′-CCACATTCAGCACAGGACTC-3′ | 5″ at 95 °C, 9″ at 58 °C, 19″ at 72 °C for 40 cycles | 708 |
| IL-18 | 5′-GATTACTTTGGCAAGCTTGAA-3′ 5′-GCTTTCGTTTTGAACAGTGAA-3′ | 5″ at 95 °C, 6″ at 53 °C, 12″ at 72 °C for 40 cycles | 470 |
| NLRP-3 | 5′-GATCTTCGCTGCGATCAACA-3′ 5′-GGGATTCGAAACACGTGCATTA-3′ | 5″ at 95 °C, 34″ at 60 °C for 40 cycles | 93 |
| β-ACTIN | 5′-CGTGGGCCGCCCTAGGCACCA-3′ 5′-TTGGCCTTAGGGTTCAGGGGGG-3′ | 243 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fusco, A.; Perfetto, B.; Savio, V.; Chiaromonte, A.; Torelli, G.; Donnarumma, G.; Baroni, A. Regulatory Ability of Lactiplantibacillus plantarum on Human Skin Health by Counteracting In Vitro Malassezia furfur Effects. J. Fungi 2023, 9, 1153. https://doi.org/10.3390/jof9121153
Fusco A, Perfetto B, Savio V, Chiaromonte A, Torelli G, Donnarumma G, Baroni A. Regulatory Ability of Lactiplantibacillus plantarum on Human Skin Health by Counteracting In Vitro Malassezia furfur Effects. Journal of Fungi. 2023; 9(12):1153. https://doi.org/10.3390/jof9121153
Chicago/Turabian StyleFusco, Alessandra, Brunella Perfetto, Vittoria Savio, Adriana Chiaromonte, Giovanna Torelli, Giovanna Donnarumma, and Adone Baroni. 2023. "Regulatory Ability of Lactiplantibacillus plantarum on Human Skin Health by Counteracting In Vitro Malassezia furfur Effects" Journal of Fungi 9, no. 12: 1153. https://doi.org/10.3390/jof9121153