
Impacts of Lithology and Slope Position on Arbuscular Mycorrhizal Fungi Communities in a Karst Forest Soil
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Soil Sampling
2.3. Soil Physiochemical Properties Analysis
2.4. DNA Extraction and Amplicon Sequencing
2.5. Statistical Analysis
3. Results
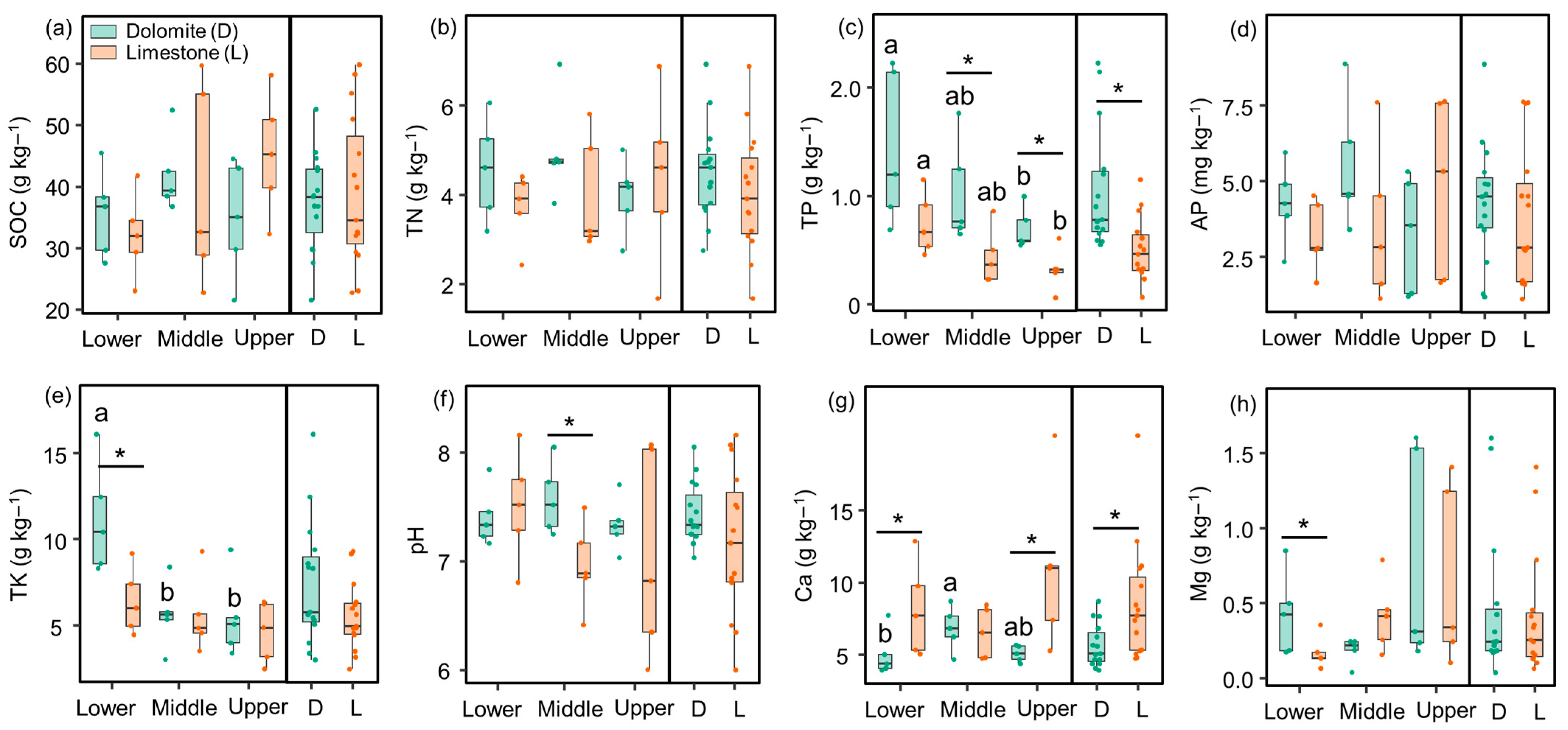
3.1. Change in Soil Properties
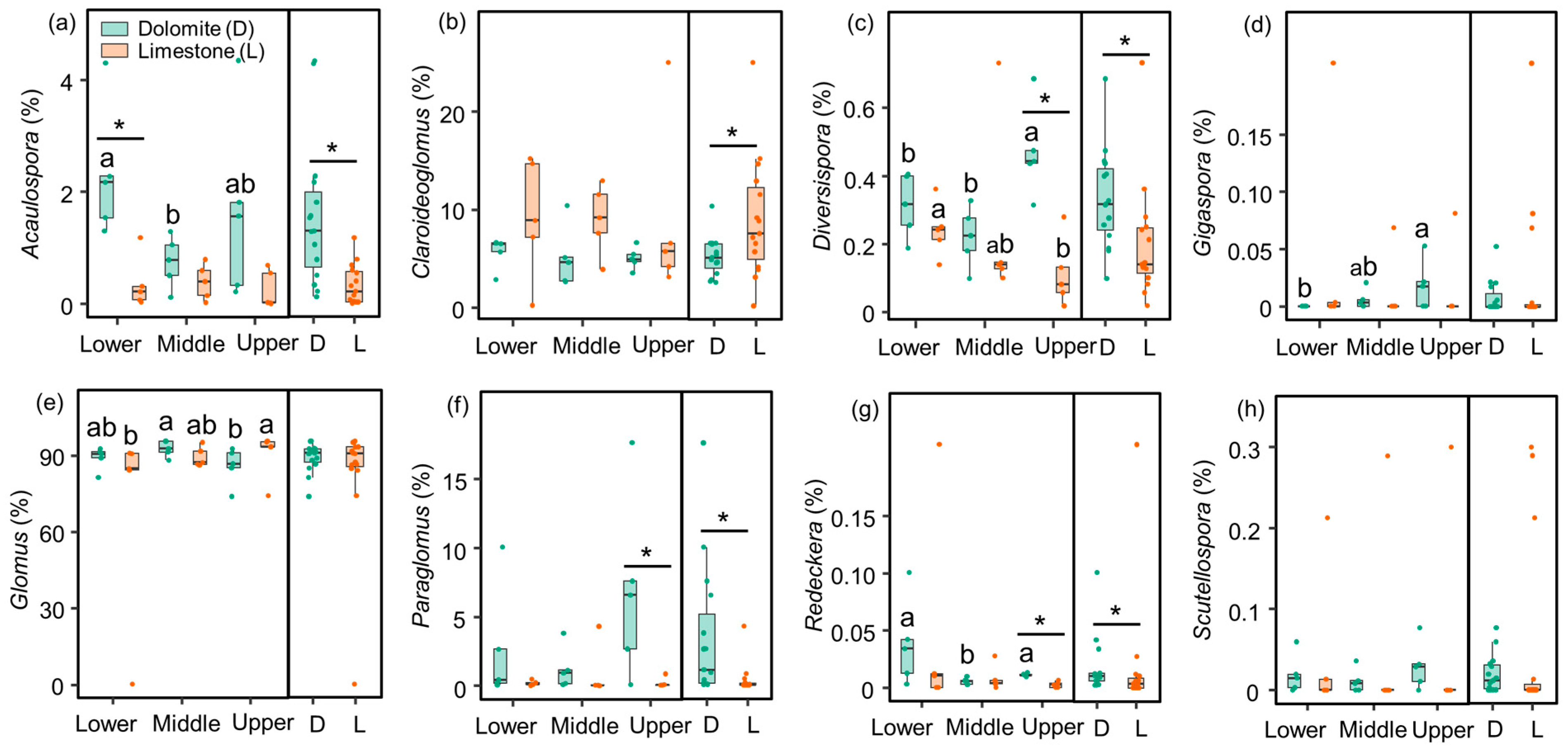
3.2. Characteristics of the Soil AMF Community
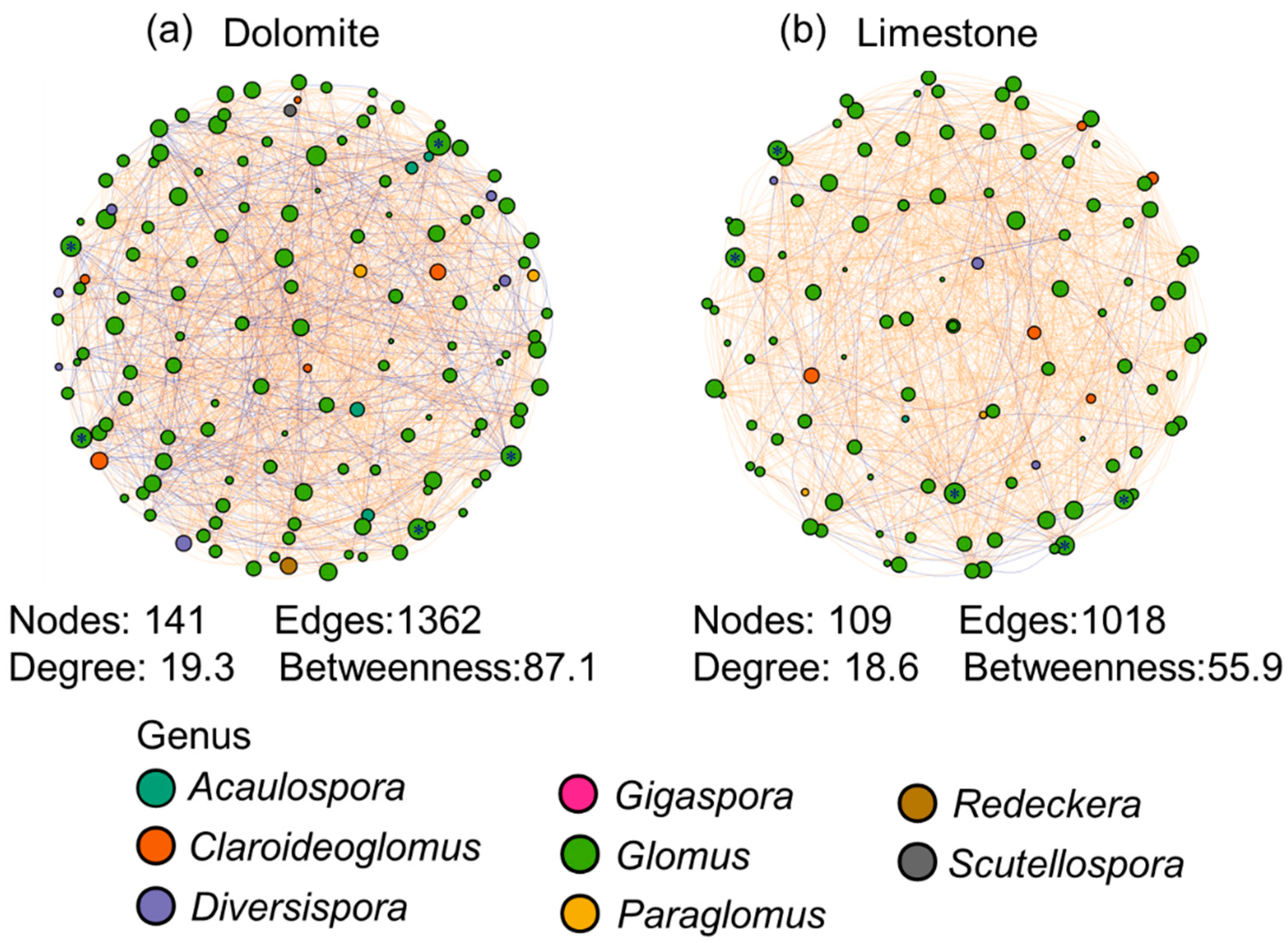
3.3. Co-Occurrence Network Analysis among AMF Taxa under Different Lithologies
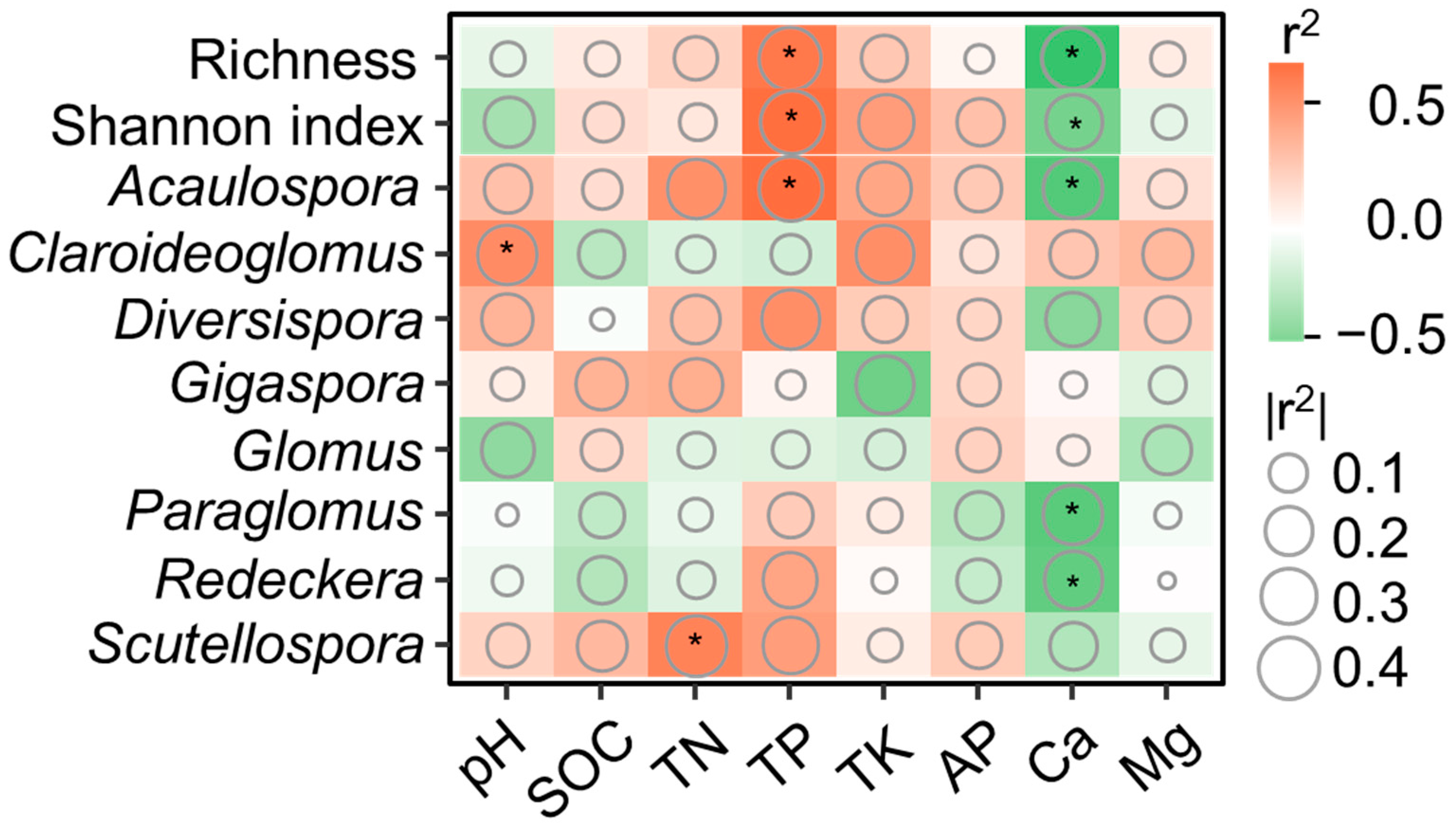
3.4. The Relationships between Soil Properties and AMF Communities
4. Discussion
4.1. Effect of Slope Position and Lithology on AMF Diversity and Community Composition
4.2. Stronger Interaction among AMF Taxa in the Dolomite Compared to Limestone
4.3. Implications for Future Management
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cao, J.; Yuan, D.; Zhang, C.; Jiang, Z. Karst ecosystem constrained by geological conditions in southwest China. Geology 2004, 32, 1–8. [Google Scholar]
- Wang, K.; Zhang, C.; Chen, H.; Yue, Y.; Zhang, W.; Zhang, M.; Qi, X.; Fu, Z. Karst landscapes of China: Patterns, ecosystem processes and services. Landsc. Ecol. 2019, 34, 2743–2763. [Google Scholar] [CrossRef]
- Li, S.; Liu, C.; Chen, J.; Wang, S. Karst ecosystem and environment: Characteristics, evolution processes, and sustainable development. Agric. Ecosyst. Environ. 2021, 306, 107173. [Google Scholar] [CrossRef]
- Jiang, Z.; Lian, Y.; Qin, X. Rocky desertification in Southwest China: Impacts, causes, and restoration. Earth-Sci. Rev. 2014, 132, 1–12. [Google Scholar] [CrossRef]
- Li, D.; Wen, L.; Yang, L.; Luo, P.; Xiao, K.; Chen, H.; Zhang, W.; He, X.; Chen, H.; Wang, K. Dynamics of soil organic carbon and nitrogen following agricultural abandonment in a karst region. J. Geophys. Res. Biogeosci. 2017, 122, 230–242. [Google Scholar] [CrossRef]
- Wen, L.; Li, D.; Chen, H.; Wang, K. Dynamics of soil organic carbon in density fractions during post-agricultural succession over two lithology types, southwest China. J. Environ. Manag. 2017, 201, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Chen, M.; He, X.; Nie, Y.; Jiang, N.; Zhang, W.; Hu, P.; Wang, K. Soil nutrients and vegetation along a karst slope gradient affect arbuscular mycorrhizal fungi colonization of roots rather than bulk soil AMF diversity. Plant Soil 2023, 489, 139–154. [Google Scholar] [CrossRef]
- Xu, X.; Wang, X.; Cleary, M.; Wang, P.; Rnnberg, J. Slope position rather than thinning intensity affects arbuscular mycorrhizal fungi (AMF) community in Chinese fir plantations. Forests 2020, 11, 273. [Google Scholar] [CrossRef]
- Zhu, J.; Jansen-Willems, A.; Müller, C.; Drsch, P. Topographic differences in nitrogen cycling mediate nitrogen retention in a subtropical, N-saturated forest catchment. Soil Biol. Biochem. 2021, 159, 108303. [Google Scholar] [CrossRef]
- Nieuwenhuyse, A.; Van Breemen, N. Quantitative aspects of weathering and neoformation in selected Costa Rican volcanic soils. Soil Sci. Soc. Am. J. 1997, 61, 1450–1458. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. (Eds.) Mycorrhizal Symbiosis, 3rd ed.; Elservier: Oxford, UK, 2008; pp. 273–281. [Google Scholar]
- Cotton, E.T.A. Arbuscular mycorrhizal fungal communities and global change: An uncertain future. FEMS Microbiol. Ecol. 2018, 94, fiy179. [Google Scholar] [CrossRef] [PubMed]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Read, D.J.; Perez-Moreno, J. Mycorrhizas and nutrient cycling in ecosystems—A journey towards relevance? New Phytol. 2010, 157, 475–492. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, J.; Moosdorf, N. The new global lithological map database GLiM: A representation of rock properties at the Earth surface. Geochem. Geophys. Geosyst. 2012, 13. [Google Scholar] [CrossRef]
- Liu, Z.; Yuan, D.; Dreybrodt, W. Comparative study of dissolution rate-determining mechanisms of limestone and dolomite. Environ. Geol. 2005, 49, 274–279. [Google Scholar] [CrossRef]
- Taylor, H.L.; Duivestein, I.J.K.; Farkas, J.; Dietzel, M.; Dosseto, A. Lithium isotopes in dolostone as a palaeo-environmental proxy–an experimental approach. Clim. Past. 2019, 15, 635–646. [Google Scholar] [CrossRef]
- Xiao, D.; He, X.; Zhang, W.; Cheng, M.; Hu, P.; Wang, K. Diazotroph and arbuscular mycorrhizal fungal diversity and community composition responses to karst and non-karst soils. Appl. Soil Ecol. 2022, 170, 104227. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.; Klironomos, J.N.; Ursic, M.; Moutoglis, P.; Streitwolf-Engel, R.; Boller, T.; Wiemken, A.; Sanders, I.R. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 1998, 396, 69–72. [Google Scholar] [CrossRef]
- Weintraub, S.R.; Taylor, P.G.; Porder, S.; Cleveland, C.C.; Asner, G.P.; Townsend, A.R. Topographic controls on soil nitrogen availability in a lowland tropical forest. Ecology 2015, 96, 1561–1574. [Google Scholar] [CrossRef]
- Tian, Q.; Wang, D.; Li, D.; Huang, L.; Wang, M.; Liao, C.; Liu, F. Variation of soil carbon accumulation across a topographic gradient in a humid subtropical mountain forest. Biogeochemistry 2020, 149, 337–354. [Google Scholar] [CrossRef]
- Fairbanks, D.; Shepard, C.; Murphy, M.; Rasmussen, C.; Chorover, J.; Rich, V.; Gallery, R. Depth and topographic controls on microbial activity in a recently burned sub-alpine catchment. Soil Biol. Biochem. 2020, 148, 107844. [Google Scholar] [CrossRef]
- Seibert, J.; Stendahl, J.; Sørensen, R. Topographical influences on soil properties in boreal forests. Geoderma 2007, 141, 139–148. [Google Scholar] [CrossRef]
- Hall, S.J.; McNicol, G.; Natake, T.; Silver, W.L. Large fluxes and rapid turnover of mineral-associated carbon across topographic gradients in a humid tropical forest: Insights from paired 14C analysis. Biogeosciences 2015, 12, 2471–2487. [Google Scholar] [CrossRef]
- Johnson, K.D.; Scatena, F.N.; Silver, W.L. Atypical soil carbon distribution across a tropical steepland forest catena. Catena 2011, 87, 391–397. [Google Scholar] [CrossRef]
- Xiao, D.; Hong, T.; Chen, M.; He, X.; Wang, K. Assessing the effect of slope position on the community assemblage of soil diazotrophs and root arbuscular mycorrhizal fungi. J. Fungi 2023, 9, 394. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Che, R.; Liu, X.; Tan, Y.; Yang, R.; Zhang, W.; He, X.; Xu, Z.; Wang, K. Arbuscular mycorrhizal fungi abundance was sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems. Biol. Fertil. Soils 2019, 55, 457–469. [Google Scholar] [CrossRef]
- Zhao, Y.; Hu, X.; Pan, P. Positive feedback relationship between shrub encroachment and arbuscular mycorrhizal fungi in the Inner Mongolia grassland of northern China. Appl. Soil Ecol. 2022, 177, 104525. [Google Scholar] [CrossRef]
- Taylor, L.L.; Leake, J.R.; Quirk, J.; Hardy, K.; Banwart, S.A.; Beerling, D.J. Biological weathering and the long-term carbon cycle: Integrating mycorrhizal evolution and function into the current paradigm. Geobiology 2009, 7, 171–191. [Google Scholar] [CrossRef]
- Pastore, G.; Weig, A.R.; Vazquez, E.; Spohn, M. Weathering of calcareous bedrocks is strongly affected by the activity of soil microorganisms. Geoderma 2022, 405, 115408. [Google Scholar] [CrossRef]
- Weemstra, M.; Peay, K.G.; Davies, S.J.; Mohamad, M.; Itoh, A.; Tan, S.; Russo, S.E. Lithological constraints on resource economies shape the mycorrhizal composition of a Bornean rain forest. New Phytol. 2020, 228, 253–268. [Google Scholar] [CrossRef]
- Wang, M.; Chen, H.; Zhang, W.; Wang, K. Soil nutrients and stoichiometric ratios as affected by land use and lithology at county scale in a karst area, southwest China. Sci. Total Environ. 2018, 619–620, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Higo, M.; Sato, R.; Serizawa, A.; Takahashi, Y.; Gunji, K.; Tatewaki, Y.; Isobe, K. Can phosphorus application and cover cropping alter arbuscular mycorrhizal fungal communities and soybean performance after a five-year phosphorus-unfertilized crop rotational system? PeerJ 2018, 6, e4606. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Yuan, Y.; Zhang, Q.; Tang, J.; Liu, Y.; Chen, X. Changes in soil organic carbon, total nitrogen, and abundance of arbuscular mycorrhizal fungi along a large-scale aridity gradient. Catena 2011, 87, 70–77. [Google Scholar] [CrossRef]
- Ngosong, C.; Gabriel, E.; Ruess, L. Collembola grazing on arbuscular mycorrhiza fungi modulates nutrient allocation in plants. Pedobiologia 2014, 57, 171–179. [Google Scholar] [CrossRef]
- Iyamuremye, F. Organic amendments and phosphorus sorption by soils. Adv. Agron. 1996, 56, 139–185. [Google Scholar]
- Chen, H.; Li, D.; Xiao, K.; Wang, K.; Treseder, K. Soil microbial processes and resource limitation in karst and non-karst forests. Funct. Ecol. 2018, 32, 1400–1409. [Google Scholar] [CrossRef]
- Kaiser, M.; Ellerbrock, R.; Wulf, M.; Dultz, S.; Hierath, C.; Sommer, M. The influence of mineral characteristics on organic matter content, composition, and stability of topsoils under long-term arable and forest land use. J. Geophys. Res.-Biogeosci. 2012, 117. [Google Scholar] [CrossRef]
- Li, D.; Wen, L.; Zhang, W.; Yang, L.; Xiao, K.; Chen, H.; Wang, K. Afforestation effects on soil organic carbon and nitrogen pools modulated by lithology. For. Ecol. Manag. 2017, 400, 85–92. [Google Scholar] [CrossRef]
- Morriën, E.; Hannula, S.E.; Snoek, L.B.; Helmsing, N.R.; Zweers, H.; de Hollander, M.; Soto, R.L.; Bouffaud, M.-L.; Buée, M.; Dimmers, W.; et al. Soil networks become more connected and take up more carbon as nature restoration progresses. Nat. Commun. 2017, 8, 14349. [Google Scholar] [CrossRef]
- Fan, K.; Cardona, C.; Li, Y.; Shi, Y.; Xiang, X.; Shen, C.; Wang, H.; Gilbert, J.A.; Chu, H. Rhizosphere-associated bacterial network structure and spatial distribution differ significantly from bulk soil in wheat crop fields. Soil Biol. Biochem. 2017, 113, 275–284. [Google Scholar] [CrossRef]
- Wagg, C.; Schlaeppi, K.; Banerjee, S.; Kuramae, E.E.; Heijden, M. Fungal-bacterial diversity and microbiome complexity predict ecosystem functioning. Nat. Commun. 2019, 10, 4841. [Google Scholar] [CrossRef]
- Xun, W.; Liu, Y.; Li, W.; Ren, Y.; Xiong, W.; Xu, Z.; Zhang, N.; Miao, Y.; Shen, Q.; Zhang, R. Specialized metabolic functions of keystone taxa sustain soil microbiome stability. Microbiome 2021, 9, 35. [Google Scholar] [CrossRef]
- Xiao, D.; He, X.; Zhang, W.; Hu, P.; Sun, M.; Wang, K. Comparison of bacterial and fungal diversity and network connectivity in karst and non-karst forests in southwest China. Sci. Total Environ. 2022, 822, 153179. [Google Scholar] [CrossRef]
- Nie, Y.; Chen, H.; Wang, K.; Ding, Y. Rooting characteristics of two widely distributed woody plant species growing in different karst habitats of southwest China. Plant Ecol. 2014, 215, 1099–1109. [Google Scholar] [CrossRef]
- Pan, F.; Liang, Y.; Wang, K.; Zhang, W. Responses of fine root functional traits to soil nutrient limitations in a Karst ecosystem of southwest China. Forests 2018, 9, 743. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Lithology | Slope Position | Lithology × Slope Position |
|---|---|---|---|
| SOC | 0.21 | 1.47 | 1.48 |
| TN | 1.05 | 0.25 | 0.95 |
| TP | 14.2 ** | 5.34 * | 0.39 |
| AP | 0.44 | 0.37 | 1.85 |
| TK | 5.15 * | 7.86 ** | 3.08 |
| pH | 2.02 | 0.68 | 1.20 |
| Ca | 7.46 * | 0.77 | 2.83 |
| Mg | 0.08 | 3.34 | 0.87 |
| Richness | 16.3 ** | 0.31 | 1.86 |
| Shannon index | 6.90 * | 0.28 | 1.24 |
| Acaulospora | 14.3 ** | 1.84 | 2.05 |
| Claroideoglomus | 4.45 * | 0.02 | 0.01 |
| Diversispora | 6.46 * | 0.42 | 4.81 * |
| Gigaspora | 1.02 | 0.18 | 0.72 |
| Glomus | 0.89 | 1.23 | 1.23 |
| Paraglomus | 6.55 * | 1.59 | 2.26 |
| Redeckera | <0.01 | 2.53 | 0.11 |
| Scutellospora | 1.17 | 0.06 | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; He, X.; Xiao, D.; Chen, M.; Cheng, M.; Wang, Z. Impacts of Lithology and Slope Position on Arbuscular Mycorrhizal Fungi Communities in a Karst Forest Soil. J. Fungi 2023, 9, 1133. https://doi.org/10.3390/jof9121133
Zhao J, He X, Xiao D, Chen M, Cheng M, Wang Z. Impacts of Lithology and Slope Position on Arbuscular Mycorrhizal Fungi Communities in a Karst Forest Soil. Journal of Fungi. 2023; 9(12):1133. https://doi.org/10.3390/jof9121133
Chicago/Turabian StyleZhao, Jin, Xunyang He, Dan Xiao, Meifeng Chen, Ming Cheng, and Zhongcheng Wang. 2023. "Impacts of Lithology and Slope Position on Arbuscular Mycorrhizal Fungi Communities in a Karst Forest Soil" Journal of Fungi 9, no. 12: 1133. https://doi.org/10.3390/jof9121133