
Effects of Interaction of Protein Hydrolysate and Arbuscular Mycorrhizal Fungi Effects on Citrus Growth and Expressions of Stress-Responsive Genes (Aquaporins and SOSs) under Salt Stress
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Treatments
2.2. Sample Collection and Physiological Index Determination
2.3. RNA Extraction and Gene Expression Analysis
2.4. Data Analysis
3. Results
3.1. Effects of PH and Ri on Plant Growth of Citrus Exposed to Salt Stress
3.2. Effects of PHs and Ri on Leaves Mineral Element Content of Citrus Exposed to Salt Stress
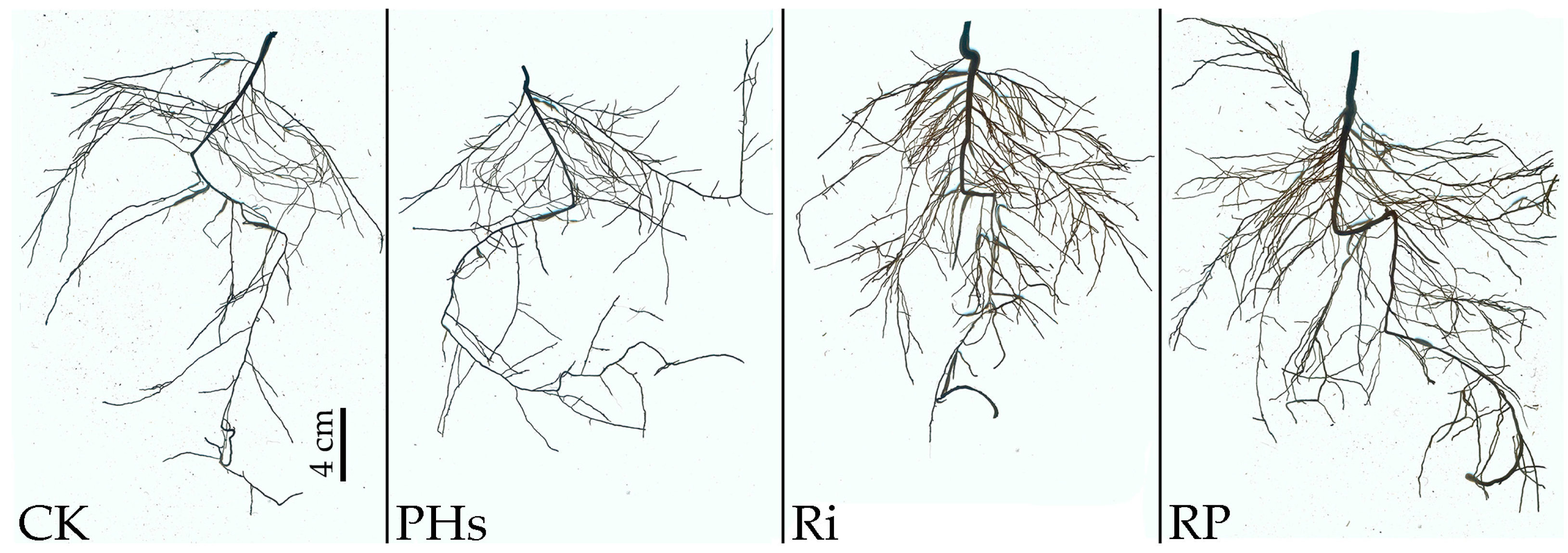
3.3. Effects of PHs and Ri on Root Structure of Citrus Exposed to Salt Stress
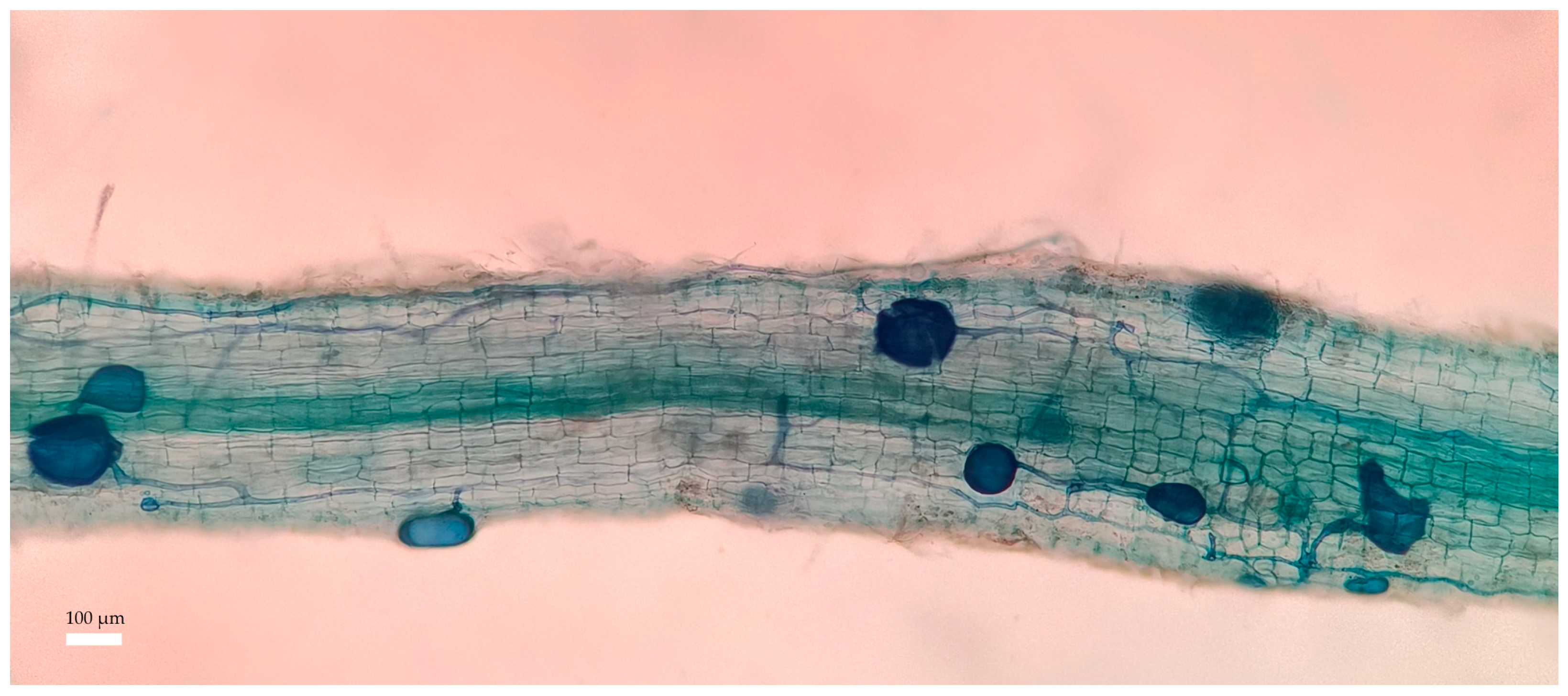
3.4. Effects of PHs on Root Colonization of Citrus Inoculated by Ri under Salt Stress
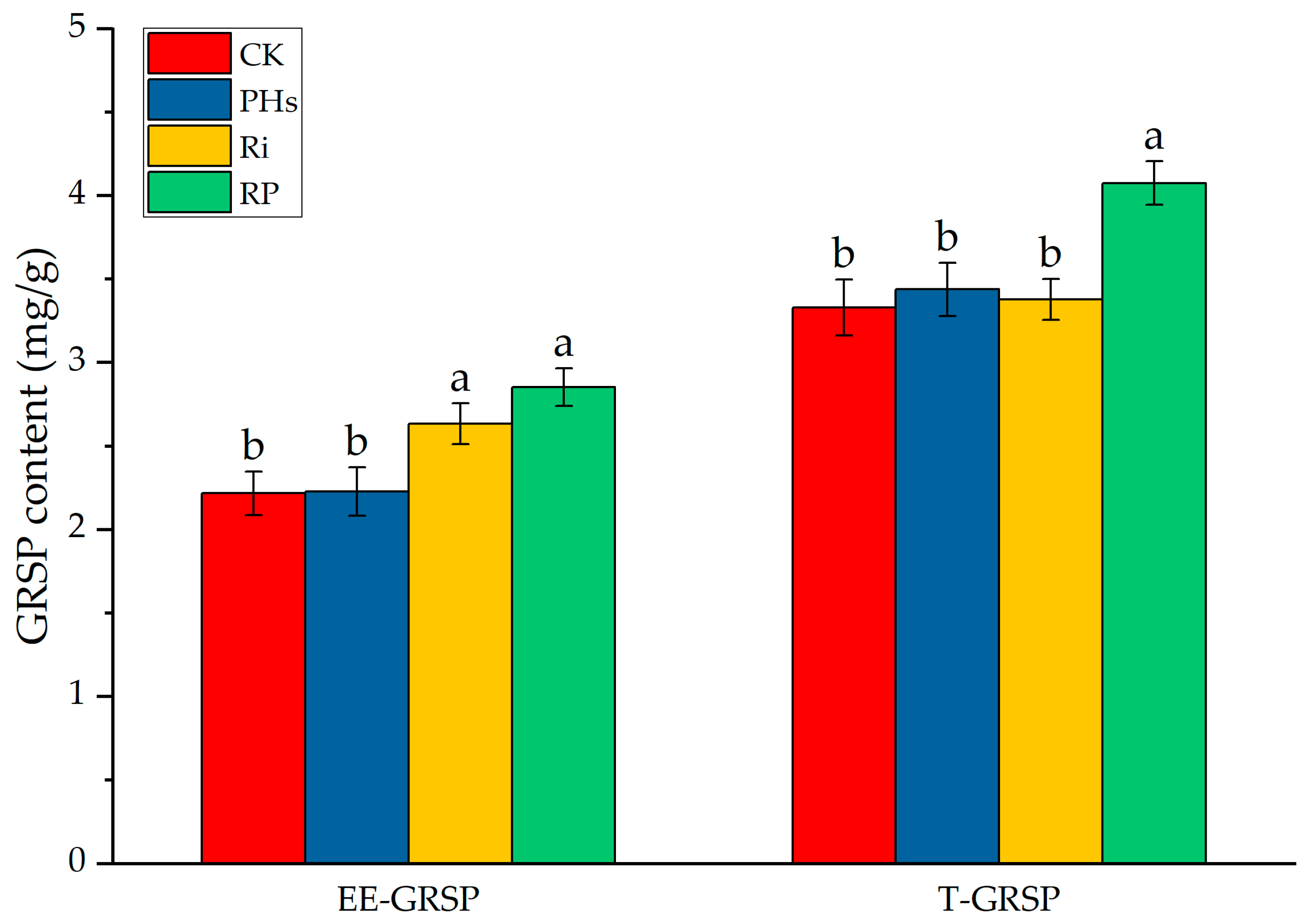
3.5. Effects of PHs and Ri on Rhizospheric GRSP Contents of Citrus Exposed to Salt Stress
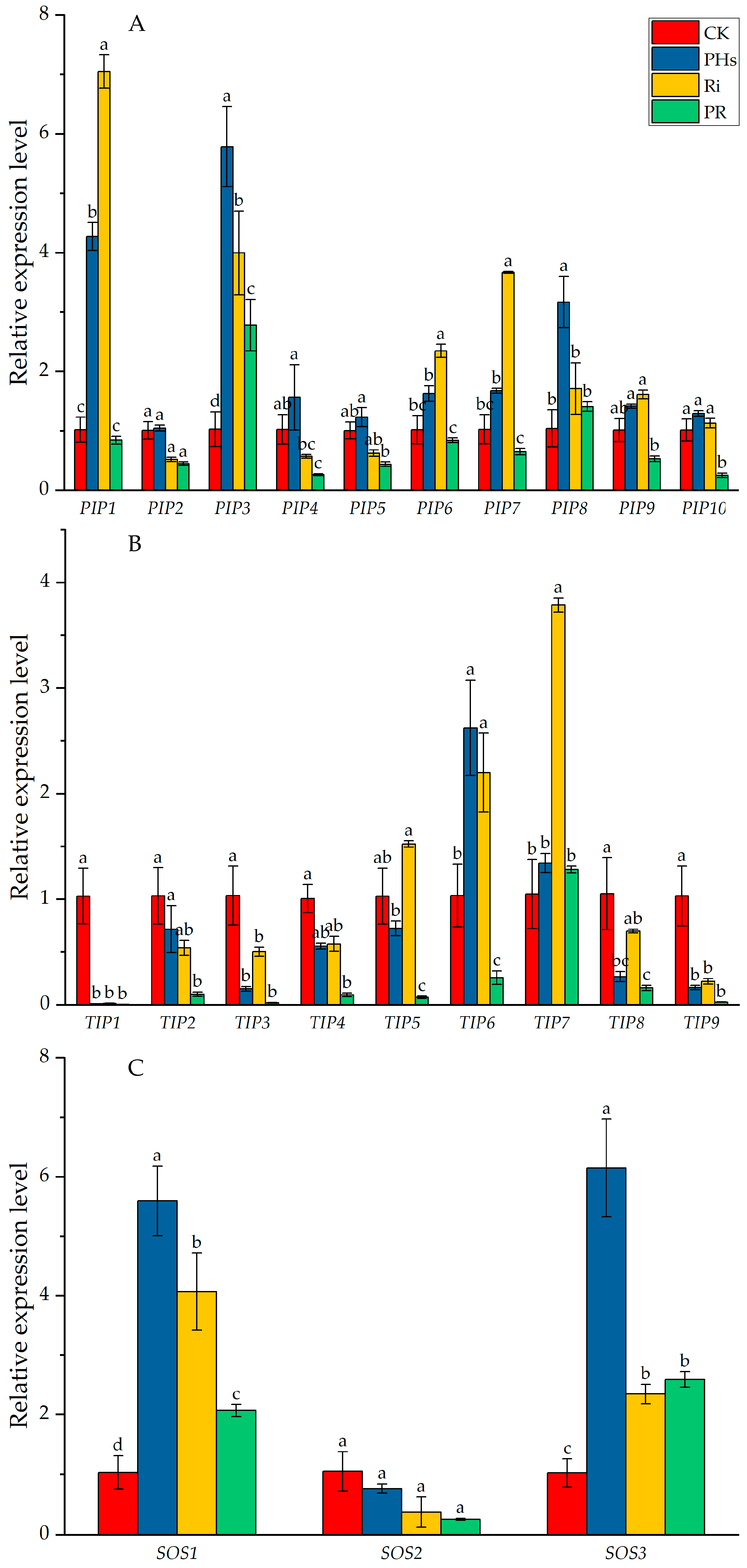
3.6. Effects of PHs and Ri on AQPs and SOSs Expressions in Root of Citrus Exposed to Salt Stress
4. Discussion
4.1. PH Application Enhanced the Tolerance of Citrus to Salt Stress by Improving Water Utilization and Sodium Effluxion
4.2. AMF Inoculation Enhanced the Tolerance of Citrus to Salt Stress by Improving Water Utilization and Sodium Effluxion
4.3. Interaction of PHs and AMF Enhanced the Tolerance of Citrus to Salt Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morinaga, K.; Ikeda, F. Photosynthetic characteristics and fruit productivity of Satsuma Mandarin (Citrus unshiu Marc.) tree under plastic greenhouse culture. J. Japan. Soc. Hort. Sci. 1991, 60, 61–69. [Google Scholar] [CrossRef]
- Sun, H.; Wei, C.; Xu, W.; Yang, J.; Wang, X.; Qiu, Y. Characteristics of salt contents in soils under greenhouse conditions in China. Environ. Sci. Pollut. Res. Int. 2019, 26, 3882–3892. [Google Scholar] [CrossRef] [PubMed]
- Haj-Amor, Z.; Araya, T.; Kim, D.; Bouri, S.; Lee, J.; Ghiloufi, W.; Yang, Y.; Kang, H.; Jhariya, M.K.; Banerjee, A.; et al. Soil salinity and its associated effects on soil microorganisms, greenhouse gas emissions, crop yield, biodiversity and desertification: A review. Sci. Total. Environ. 2022, 843, 156946. [Google Scholar] [CrossRef] [PubMed]
- Kourgialas, N. Drought and salinity in citriculture: Optimal practices to alleviate salinity and water stress. Agronomy 2021, 11, 1283. [Google Scholar] [CrossRef]
- Prior, L.D.; Grieve, A.M.; Bevington, K.B.; Slavich, P.G. Long-term effects of saline irrigation water on ‘Valencia’ orange trees: Relationships between growth and yield, and salt levels in soil and leaves. Aust. J. Agric. Res. 2007, 58, 349–358. [Google Scholar] [CrossRef]
- Colin, L.; Ruhnow, F.; Zhu, J.; Zhao, C.; Zhao, Y.; Persson, S. The cell biology of primary cell walls during salt stress. Plant Cell. 2023, 35, 201–217. [Google Scholar] [CrossRef]
- Ali, A.; Petrov, V.; Yun, D.; Gechev, T. Revisiting plant salt tolerance: Novel components of the SOS pathway. Trends Plant Sci. 2023, 28, 1060–1069. [Google Scholar] [CrossRef]
- Dai, W.; Wang, M.; Gong, X.; Liu, J.H. The transcription factor FcWRKY40 of Fortunella crassifolia functions positively in salt tolerance through modulation of ion homeostasis and proline biosynthesis by directly regulating SOS2 and P5CS1 homologs. N. Phytol. 2018, 219, 972–989. [Google Scholar] [CrossRef]
- Shahzad, B.; Shabala, L.; Zhou, M.; Venkataraman, G.; Solis, C.A.; Page, D.; Chen, Z.H.; Shabala, S. Comparing essentiality of SOS1-Mediated Na+ exclusion in salinity tolerance between cultivated and wild rice species. Int. J. Mol. Sci. 2022, 23, 9900. [Google Scholar] [CrossRef]
- van Zelm, E.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant. Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef]
- Maurel, C.; Boursiac, Y.; Luu, D.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.; Agarwal, P.; Shanware, A.; Sane, V.A. Heterologous expression of two jatropha aquaporins imparts drought and salt tolerance and improves seed viability in transgenic Arabidopsis thaliana. PLoS ONE 2015, 10, e128866. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ni, H.; Yan, W.; Xie, Y.; Liu, X.; Tan, X.; Zhang, L.; Zhang, S. Overexpression of an aquaporin protein from Aspergillus glaucus confers salt tolerance in transgenic soybean. Transgenic Res. 2021, 30, 727–737. [Google Scholar] [CrossRef]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar applications of protein hydrolysate, plant and seaweed extracts increase yield but differentially modulate fruit quality of greenhouse tomato. Hortscience 2017, 9, 1214–1220. [Google Scholar] [CrossRef]
- Amirkhani, M.; Netravali, A.N.; Huang, W.; Taylor, A.G. Investigation of soy protein-based biostimulant seed coating for broccoli seedling and plant growth enhancement. Hortscience 2016, 51, 1121–1126. [Google Scholar] [CrossRef]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Canaguier, R.; Colla, G. The effect of a plant-derived biostimulant on metabolic profiling and crop performance of lettuce grown under saline conditions. Sci. Hortic-Amst. 2015, 182, 124–133. [Google Scholar] [CrossRef]
- Sorrentino, M.; De Diego, N.; Ugena, L.; Spíchal, L.; Lucini, L.; Miras-Moreno, B.; Zhang, L.; Rouphael, Y.; Colla, G.; Panzarová, K. Seed priming with protein hydrolysates improves Arabidopsis growth and stress tolerance to abiotic stresses. Front. Plant. Sci. 2021, 12, 626301. [Google Scholar] [CrossRef]
- Sorrentino, M.; Panzarová, K.; Spyroglou, I.; Spíchal, L.; Buffagni, V.; Ganugi, P.; Rouphael, Y.; Colla, G.; Lucini, L.; De Diego, N. Integration of phenomics and metabolomics datasets reveals different mode of action of biostimulants based on protein hydrolysates in Lactuca sativa L. and Solanum lycopersicum L. under salinity. Front. Plant. Sci. 2021, 12, 808711. [Google Scholar] [CrossRef]
- Liu, S.; Guo, X.; Feng, G.; Maimaitiaili, B.; Fan, J.; He, X. Indigenous arbuscular mycorrhizal fungi can alleviate salt stress and promote growth of cotton and maize in saline fields. Plant Soil. 2016, 398, 195–206. [Google Scholar] [CrossRef]
- Estrada, B.; Aroca, R.; Barea, J.M.; Ruiz-Lozano, J.M. Native arbuscular mycorrhizal fungi isolated from a saline habitat improved maize antioxidant systems and plant tolerance to salinity. Plant Sci. Int. J. Exp. Plant Biol. 2013, 201–202, 42–51. [Google Scholar] [CrossRef]
- Zou, Y. Glomalin: A potential soil conditioner for perennial fruits. Int. J. Agric. Biol. 2016, 18, 293–297. [Google Scholar] [CrossRef]
- Zou, Y.; Srivastava, A.K.; Wu, Q.; Huang, Y. Glomalin-related soil protein and water relations in mycorrhizal citrus (Citrus tangerina) during soil water deficit. Arch. Agron. Soil. Sci. 2013, 60, 1103–1114. [Google Scholar] [CrossRef]
- Hammer, E.C.; Rillig, M.C. The influence of different stresses on glomalin levels in an arbuscular mycorrhizal fungus—Salinity increases glomalin content. PLoS ONE 2011, 6, e28426. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.Y.; Li, Q.S.; Ding, W.Y.; Dong, L.W.; Deng, M.; Chen, J.H.; Tian, X.; Hashem, A.; Al, A.A.B.F.; Alenazi, M.M.; et al. Arbuscular mycorrhizal fungi inoculation impacts expression of aquaporins and salt overly sensitive genes and enhances tolerance of salt stress in tomato. Chem. Biol. Technol. Ag. 2023, 10, 5. [Google Scholar] [CrossRef]
- Liang, S.; Li, Q.; Liu, M.; Hashem, A.; Al-Arjani, A.F.; Alenazi, M.M.; Abd Allah, E.F.; Muthuramalingam, P.; Wu, Q. Mycorrhizal effects on growth and expressions of stress-responsive genes (aquaporins and SOSs) of tomato under salt stress. J. Fungi 2022, 8, 1305. [Google Scholar] [CrossRef]
- Abbaspour, H.; Pour, F.S.N.; Abdel-Wahhab, M.A. Arbuscular mycorrhizal symbiosis regulates the physiological responses, ion distribution and relevant gene expression to trigger salt stress tolerance in pistachio. Physiol. Mol. Biol. Plants 2021, 27, 1765–1778. [Google Scholar] [CrossRef]
- Wu, Q.; Liu, C.; Zhang, D.; Zou, Y.; He, X.; Wu, Q. Mycorrhiza alters the profile of root hairs in trifoliate orange. Mycorrhiza 2016, 26, 237–247. [Google Scholar] [CrossRef]
- Li, Q.; Srivastava, A.K.; Zou, Y.; Wu, Q. Field inoculation responses of arbuscular mycorrhizal fungi versus endophytic fungi on sugar metabolism associated changes in fruit quality of Lane late navel orange. Sci. Hortic-Amst. 2023, 308, 111587. [Google Scholar] [CrossRef]
- Jia-Dong, H.; Tao, D.; Hui-Hui, W.; Ying-Ning, Z.; Qiang-Sheng, W.; Kamil, K. Mycorrhizas induce diverse responses of root TIP aquaporin gene expression to drought stress in trifoliate orange. Sci. Hortic-Amst. 2019, 243, 64–69. [Google Scholar] [CrossRef]
- Cheng, X.; Wu, H.; Zou, Y.; Wu, Q.; Kuča, K. Mycorrhizal response strategies of trifoliate orange under well-watered, salt stress, and waterlogging stress by regulating leaf aquaporin expression. Plant Physiol. Biochem. 2021, 162, 27–35. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, P.; Wu, Q.; Zou, Y.; Bao, Q.; Wu, Q. Arbuscular mycorrhizas improve plant growth and soil structure in trifoliate orange under salt stress. Arch. Acker Pflanzenbau Bodenkd. 2017, 63, 491–500. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N.; He, X.H. Contributions of arbuscular mycorrhizal fungi to growth, photosynthesis, root morphology and ionic balance of citrus seedlings under salt stress. Acta Physiol. Plant 2010, 32, 297–304. [Google Scholar] [CrossRef]
- Sykes, S.R. Chloride and sodium excluding capacities of citrus rootstock germplasm introduced to Australia from the People’s Republic of China. Sci. Hortic-Amst. 2011, 128, 443–449. [Google Scholar] [CrossRef]
- Wang, N.; Yan, T.; Fu, L.; Zhou, G.; Liu, Y.; Peng, S.A. Differences in boron distribution and forms in four citrus scion–rootstock combinations with contrasting boron efficiency under boron-deficient conditions. Trees 2014, 28, 1589–1598. [Google Scholar] [CrossRef]
- Wang, P.; Shu, B.; Wang, Y.; Zhang, D.J.; Liu, J.F.; Xia, R.X. Diversity of arbuscular mycorrhizal fungi in red tangerine (Citrus reticulata Blanco) rootstock rhizospheric soils from hillside citrus orchards. Pedobiologia 2013, 56, 161–167. [Google Scholar] [CrossRef]
- Jin, L.; Liu, Y.; Du, W.; Fu, L.; Hussain, S.B.; Peng, S. Physiological and transcriptional analysis reveals pathways involved in iron deficiency chlorosis in fragrant citrus. Tree Genet. Genomes 2017, 13, 51. [Google Scholar] [CrossRef]
- Ma, S.; Wang, T.; Ma, S. Effects of drip irrigation on root activity pattern, root-sourced signal characteristics and yield stability of winter wheat. Agr. Water Manag. 2022, 271, 107783. [Google Scholar] [CrossRef]
- Janos, D.P.; Garamszegi, S.; Beltran, B. Glomalin extraction and measurement. Soil Biol. Biochem. 2008, 40, 728–739. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Vives-Peris, V.; López-Climent, M.F.; Moliner-Sabater, M.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Morphological, physiological, and molecular scion traits are determinant for salt-stress tolerance of grafted citrus plants. Front. Plant. Sci. 2023, 14, 1145625. [Google Scholar] [CrossRef]
- Al-Yassin, A. Influence of salinity on citrus: A review paper. J. Cent. Eur. Agric. 2005, 5, 263–272. [Google Scholar]
- Poles, L.; Licciardello, C.; Distefano, G.; Nicolosi, E.; Gentile, A.; La Malfa, S. Recent advances of in vitro culture for the application of new breeding techniques in citrus. Plants 2020, 9, 938. [Google Scholar] [CrossRef] [PubMed]
- Nephali, L.; Piater, L.A.; Dubery, I.A.; Patterson, V.; Huyser, J.; Burgess, K.; Tugizimana, F. Biostimulants for plant growth and mitigation of abiotic stresses: A metabolomics perspective. Metabolites 2020, 10, 505. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Front. Plant. Sci. 2014, 5, 448. [Google Scholar] [CrossRef]
- Lu, Y.; Fricke, W. Salt Stress-Regulation of Root Water Uptake in a Whole-Plant and Diurnal Context. Int. J. Mol. Sci. 2023, 24, 8070. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.; Hao, J.; Su, Y.; Li, B.; Zhao, N.; Zhu, M.; Huang, Y.; Tian, B.; Shi, G.; Hua, J. Two aquaporin genes, GhPIP2;7 and GhTIP2;1, positively regulate the tolerance of upland cotton to salt and osmotic stresses. Front. Plant. Sci. 2021, 12, 780486. [Google Scholar] [CrossRef]
- Qiu, Q.; Guo, Y.; Quintero, F.J.; Pardo, J.M.; Schumaker, K.S.; Zhu, J. Regulation of vacuolar Na+/H+ exchange in Arabidopsis thaliana by the salt-overly-sensitive (SOS) pathway. J. Biol. Chem. 2004, 279, 207–215. [Google Scholar] [CrossRef]
- Shi, H.; Ishitani, M.; Kim, C.; Zhu, J.K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6896–6901. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, W.; Xie, Q.; Liu, N.; Liu, L.; Wang, D.; Zhang, X.; Yang, C.; Chen, X.; Tang, D.; et al. Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi. Science 2017, 356, 1172–1175. [Google Scholar] [CrossRef]
- Chandrasekaran, M. Arbuscular mycorrhizal fungi mediated alleviation of drought stress via non-enzymatic antioxidants: A meta-analysis. Plants 2022, 11, 2488. [Google Scholar] [CrossRef]
- Wu, N.; Li, Z.; Wu, F.; Zhen, L. Sex-specific photosynthetic capacity and Na+ homeostasis in Populus euphratica exposed to NaCl stress and AMF inoculation. Front. Plant. Sci. 2022, 13, 1066954. [Google Scholar] [CrossRef] [PubMed]
- Ortíz, J.; Sanhueza, C.; Romero-Munar, A.; Hidalgo-Castellanos, J.; Castro, C.; Bascuñán-Godoy, L.; Coba de la Peña, T.; López-Gómez, M.; Florez-Sarasa, I.; Del-Saz, N.F. In vivo metabolic regulation of alternative oxidase under nutrient deficiency-interaction with arbuscular mycorrhizal fungi and rhizobium bacteria. Int. J. Mol. Sci. 2020, 21, 4201. [Google Scholar] [CrossRef] [PubMed]
- Riaz, M.; Kamran, M.; Fang, Y.; Wang, Q.; Cao, H.; Yang, G.; Deng, L.; Wang, Y.; Zhou, Y.; Anastopoulos, I.; et al. Arbuscular mycorrhizal fungi-induced mitigation of heavy metal phytotoxicity in metal contaminated soils: A critical review. J. Hazard. Mater. 2021, 402, 123919. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Sun, X.; Tian, R.; Li, K.; Ni, M.; Ying, J.; Xu, L.; Liu, L.; Wang, Y. Genome-wide characterization of the aquaporin gene family in radish and functional analysis of RsPIP2-6 involved in salt stress. Front. Plant. Sci. 2022, 13, 860742. [Google Scholar] [CrossRef]
- Wang, J.; Yang, L.; Chai, S.; Ren, Y.; Guan, M.; Ma, F.; Liu, J. An aquaporin gene MdPIP1;2 from Malus domestica confers salt tolerance in transgenic Arabidopsis. J. Plant. Physiol. 2022, 273, 153711. [Google Scholar] [CrossRef]
- Yaghobi, M.; Heidari, P. Genome-wide analysis of aquaporin gene family in Triticum turgidum and its expression profile in response to salt stress. Genes 2023, 14, 202. [Google Scholar] [CrossRef]
- Ayadi, M.; Brini, F.; Masmoudi, K. Overexpression of a wheat aquaporin gene, TdPIP2;1, enhances salt and drought tolerance in transgenic durum wheat cv. Maali. Int. J. Mol. Sci. 2019, 20, 2389. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, J.; Jia, C.; Hu, W.; Song, S.; Xu, B.; Jin, Z. Overexpression of a banana aquaporin gene MaPIP1;1 enhances tolerance to multiple abiotic stresses in transgenic banana and analysis of its interacting transcription factor. Front. Plant. Sci. 2021, 12, 699230. [Google Scholar] [CrossRef]
- Tang, H.; Yu, Q.; Li, Z.; Liu, F.; Su, W.; Zhang, C.; Ling, H.; Luo, J.; Su, Y.; Que, Y. A PIP-mediated osmotic stress signaling cascade plays a positive role in the salt tolerance of sugarcane. BMC Plant. Biol. 2021, 21, 589. [Google Scholar] [CrossRef]
- Wang, X.; Cai, H.; Li, Y.; Zhu, Y.; Ji, W.; Bai, X.; Zhu, D.; Sun, X. Ectopic overexpression of a novel Glycine soja stress-induced plasma membrane intrinsic protein increases sensitivity to salt and dehydration in transgenic Arabidopsis thaliana plants. J. Plant. Res. 2015, 128, 103–113. [Google Scholar] [CrossRef]
- Ferguson, L.; Grattan, S.R. How salinity damages citrus: Osmotic effects and specific ion toxicities. Horttechnology 2005, 15, 95–99. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant. Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Hernández, J.M.; Benítez-García, I.; Mazorra-Manzano, M.A.; Ramírez-Suárez, J.C.; Sánchez, E. Strategies for production, characterization and application of protein-based biostimulants in agriculture: A review. Chil. J. Agr. Res. 2020, 80, 274–289. [Google Scholar] [CrossRef]
- Schmidt, C.S.; Mrnka, L.; Frantík, T.; Bárnet, M.; Vecová, E.B. Impact of protein hydrolysate biostimulants on growth of barley and wheat and their interaction with symbionts and pathogens. Agr. Food Sci. 2020, 29, 222–238. [Google Scholar] [CrossRef]
- Metwally, R.A.; Soliman, S.A.; Abdel Latef, A.A.H.; Abdelhameed, R.E. The individual and interactive role of arbuscular mycorrhizal fungi and Trichoderma viride on growth, protein content, amino acids fractionation, and phosphatases enzyme activities of onion plants amended with fish waste. Ecotox Environ. Safe 2021, 214, 112072. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height (cm) | Stem Diameter (mm) | Leaf Number | Shoot Dry Weight (g) |
|---|---|---|---|---|
| CK | 8.7 ± 0.44 c | 1.6 ± 0.12 c | 6.8 ± 0.32 c | 0.61 ± 0.06 c |
| PHs | 12.2 ± 0.27 b | 2.2 ± 0.06 b | 10.8 ± 0.19 b | 1.08 ± 0.07 b |
| Ri | 20.1 ± 3.33 a | 2.9 ± 0.10 a | 17.6 ± 1.50 a | 2.88 ± 0.43 a |
| PHs + Ri | 23.9 ± 3.18 a | 2.8 ± 0.38 a | 18.3 ± 1.56 a | 3.54 ± 0.59 a |
| Statistical significance | ||||
| PHs | * | * | *** | * |
| Ri | *** | *** | *** | *** |
| PHs×Ri | ns | ** | * | ns |
| Treatments | N (g/kg DW) | P (g/kg DW) | K (g/kg DW) | Ca (g/kg DW) | Mg (g/kg DW) | Na (g/kg DW) |
|---|---|---|---|---|---|---|
| CK | 21.86 ± 1.12 b | 1.35 ± 0.03 c | 13.43 ± 0.55 b | 35.52 ± 1.01 c | 4.19 ± 0.58 b | 5.83 ± 0.56 a |
| PHs | 29.48 ± 1.42 a | 2.16 ± 0.15 b | 16.07 ± 0.31 a | 41.91 ± 0.87 b | 6.50 ± 0.36 a | 1.45 ± 0.14 b |
| Ri | 27.59 ± 1.02 a | 2.26 ± 0.13 b | 8.42 ± 0.49 c | 45.54 ± 1.34 a | 3.61 ± 0.42 b | 1.22 ± 0.19 bc |
| PHs + Ri | 28.97 ± 0.99 a | 3.11 ± 0.19 a | 7.72 ± 0.60 c | 43.29 ± 1.11 ab | 3.44 ± 0.27 b | 1.18 ± 0.22 c |
| Statistical significance | ||||||
| PHs | *** | *** | * | *** | * | *** |
| Ri | ** | *** | *** | *** | *** | *** |
| PHs×Ri | ** | ns | *** | *** | ** | *** |
| Treatments | Lateral Roots Number | Root Volume (mL) | Total Root Length (cm) | Root Projection Area (cm2) | Root Surface Area (cm2) | Root Activity (mg/g∙h) |
|---|---|---|---|---|---|---|
| CK | 97.8 ± 6.71 d | 1.0 ± 0.14 b | 306.88 ± 16.63 d | 2.61 ± 0.21 b | 8.19 ± 0.65 b | 0.17 ± 0.02 d |
| PHs | 118.5 ± 5.99 c | 1.3 ± 0.19 b | 357.10 ± 16.97 c | 3.04 ± 0.46 b | 9.01 ± 1.86 b | 0.26 ± 0.02 c |
| Ri | 183.9 ± 15.66 b | 3.0 ± 0.41 a | 675.88 ± 33.56 b | 5.86 ± 0.67 a | 18.41 ± 2.11 a | 0.33 ± 0.02 b |
| PHs + Ri | 214.8 ± 15.99 a | 3.5 ± 0.45 a | 765.17 ± 46.11 a | 6.34 ± 1.30 a | 19.92 ± 4.08 a | 0.40 ± 0.01 a |
| Statistical significance | ||||||
| PHs | ** | ns | * | ns | ns | *** |
| Ri | *** | *** | *** | *** | *** | *** |
| PHs×Ri | ns | ns | ns | ns | ns | ns |
| Treatments | Hyphae Colonization Rate (%) | Arbuscule Colonization Rate (%) | Vesicle Colonization Rate (%) | Total Colonization Rate (%) | Spore Density (per/10 g) | Hyphal Density (cm/g) |
|---|---|---|---|---|---|---|
| CK | / | / | / | / | / | / |
| PHs | / | / | / | / | / | / |
| Ri | 32.69 ± 3.91 b | 13.26 ± 2.99 a | 5.55 ± 2.86 a | 35.18 ± 2.97 b | 59.75 ± 13.35 a | 19.02 ± 1.13 b |
| PHs + Ri | 39.69 ± 2.61 a | 14.80 ± 1.74 a | 6.53 ± 2.87 a | 42.52 ± 2.50 a | 60.25 ± 5.80 a | 24.40 ± 2.01 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Q.; Jin, L.; Wang, P.; Liu, F.; Huang, B.; Wen, M.; Wu, S. Effects of Interaction of Protein Hydrolysate and Arbuscular Mycorrhizal Fungi Effects on Citrus Growth and Expressions of Stress-Responsive Genes (Aquaporins and SOSs) under Salt Stress. J. Fungi 2023, 9, 983. https://doi.org/10.3390/jof9100983
Lu Q, Jin L, Wang P, Liu F, Huang B, Wen M, Wu S. Effects of Interaction of Protein Hydrolysate and Arbuscular Mycorrhizal Fungi Effects on Citrus Growth and Expressions of Stress-Responsive Genes (Aquaporins and SOSs) under Salt Stress. Journal of Fungi. 2023; 9(10):983. https://doi.org/10.3390/jof9100983
Chicago/Turabian StyleLu, Qi, Longfei Jin, Peng Wang, Feng Liu, Bei Huang, Mingxia Wen, and Shaohui Wu. 2023. "Effects of Interaction of Protein Hydrolysate and Arbuscular Mycorrhizal Fungi Effects on Citrus Growth and Expressions of Stress-Responsive Genes (Aquaporins and SOSs) under Salt Stress" Journal of Fungi 9, no. 10: 983. https://doi.org/10.3390/jof9100983