
Genomic Epidemiology Identifies Azole Resistance Due to TR34/L98H in European Aspergillus fumigatus Causing COVID-19-Associated Pulmonary Aspergillosis
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. CAPA Definition
2.2. Fungal Isolates
Clinical Characterisation of the CAPA Isolates
2.3. Antifungal Susceptibility Testing
2.4. Genomic DNA Preparation and Whole Genome Sequencing
2.5. Bioinformatics Analysis
2.6. A. fumigatus and MAT Identification
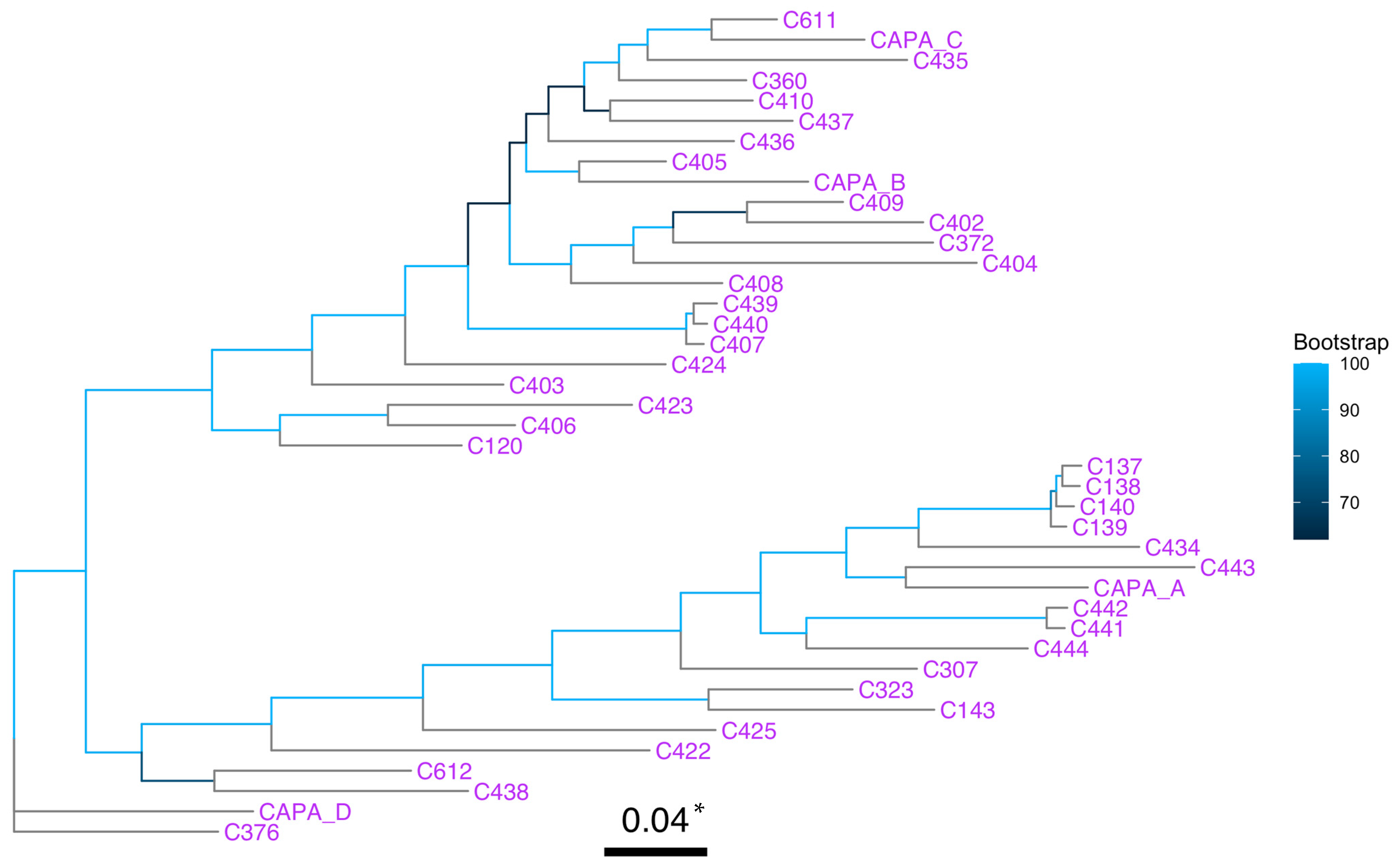
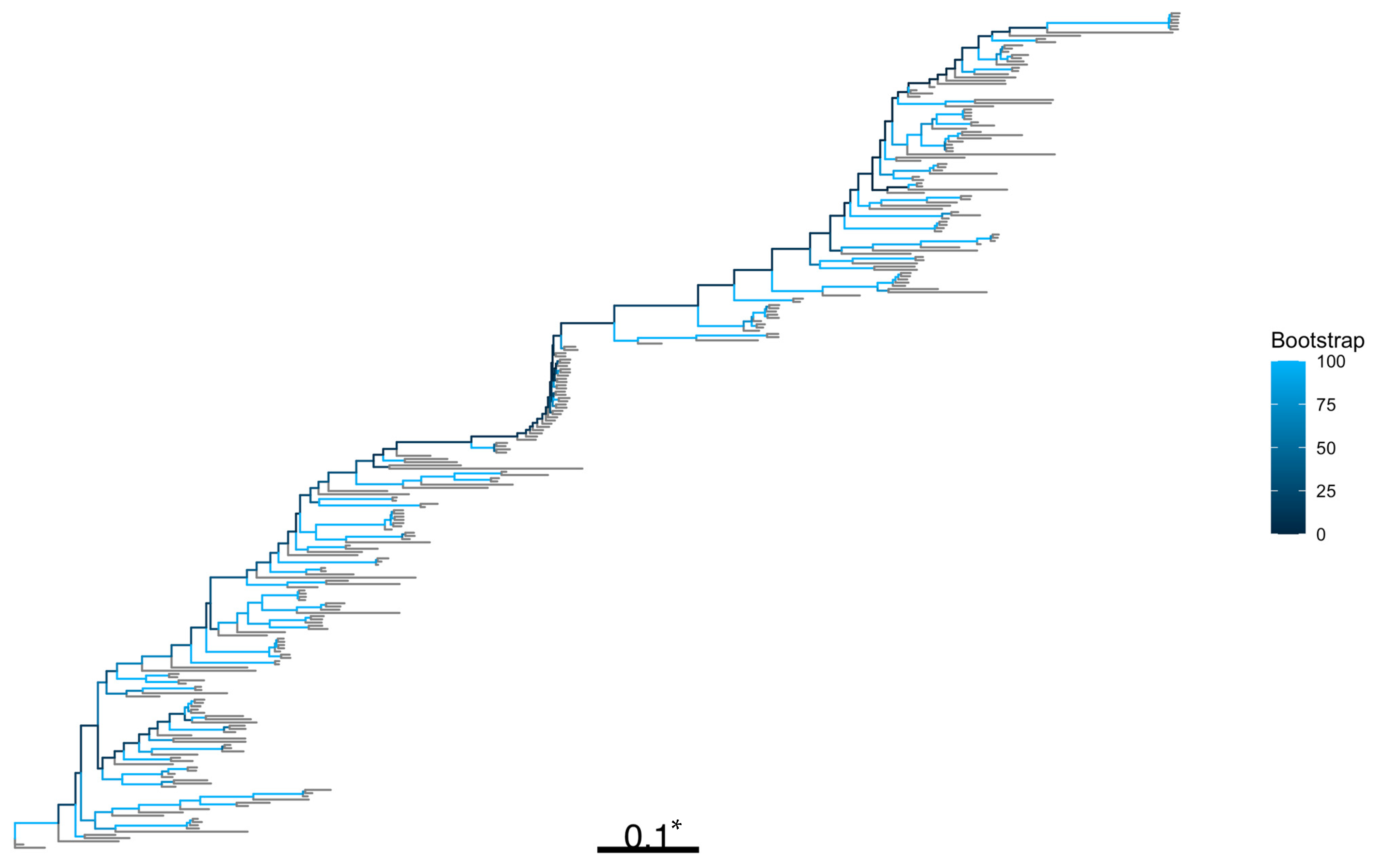
2.7. Phylogenetic and Spatial Analyses
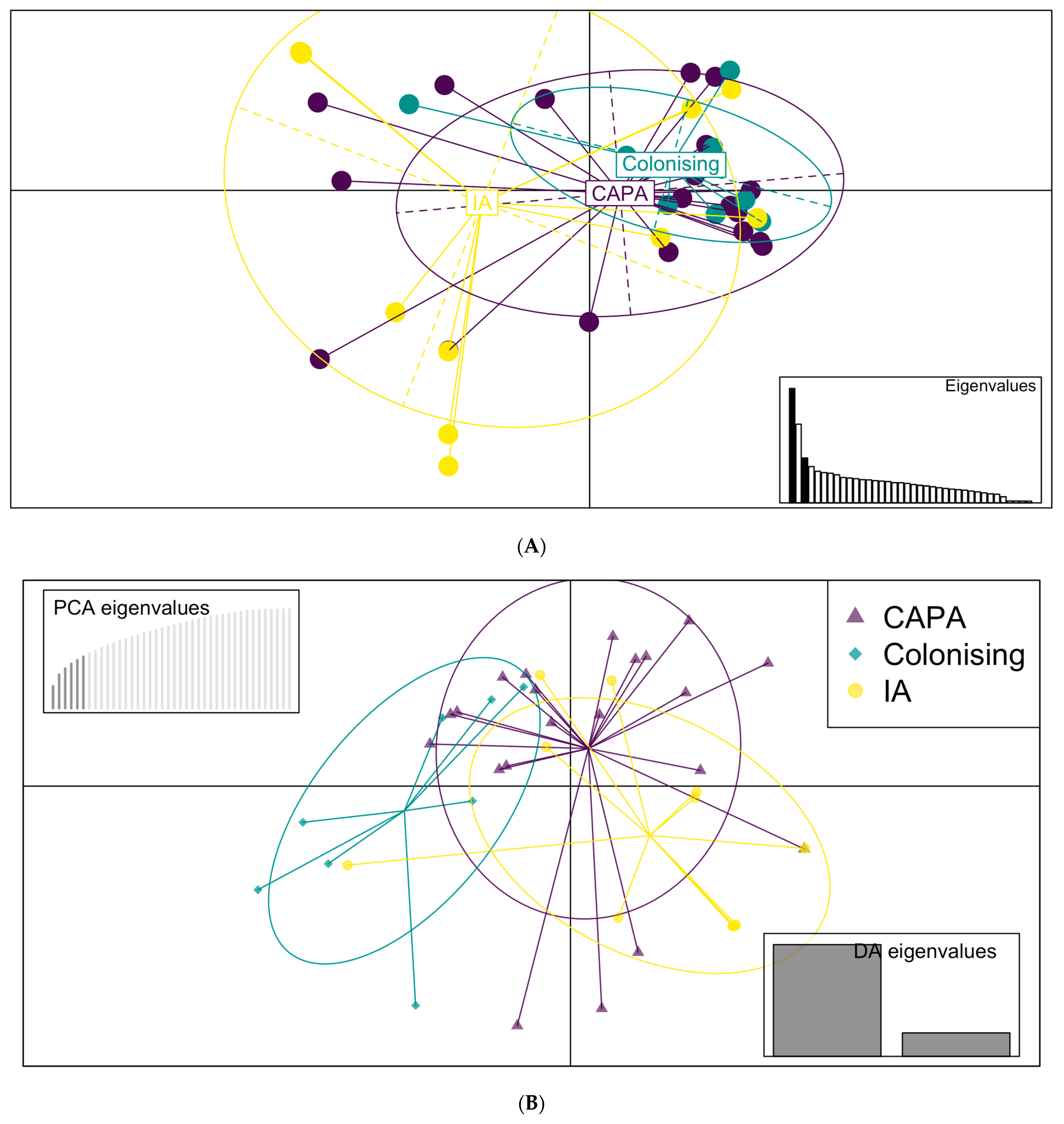
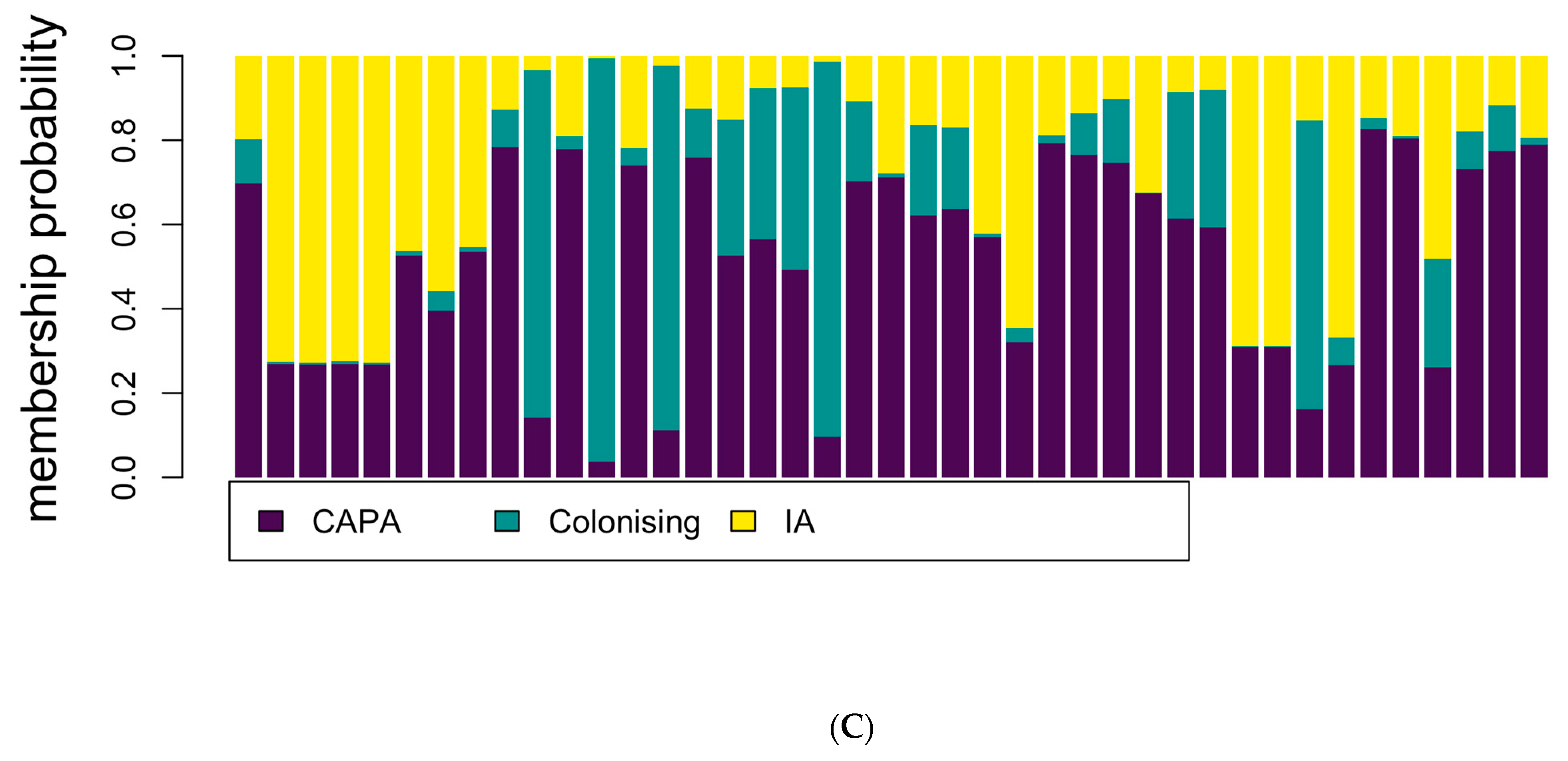
2.8. Statistical Analyses
3. Results
3.1. WGS of 21 CAPA Isolates
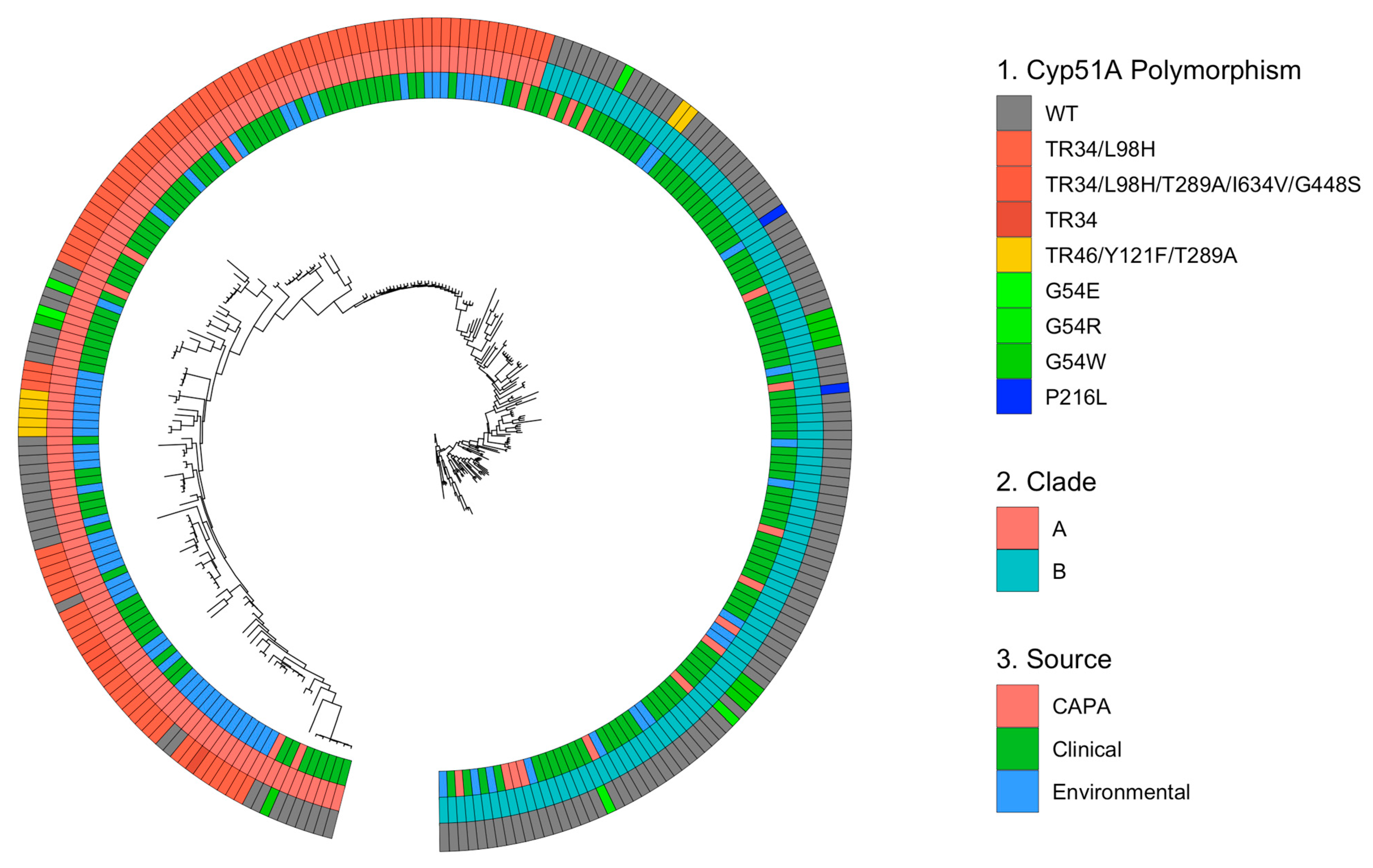
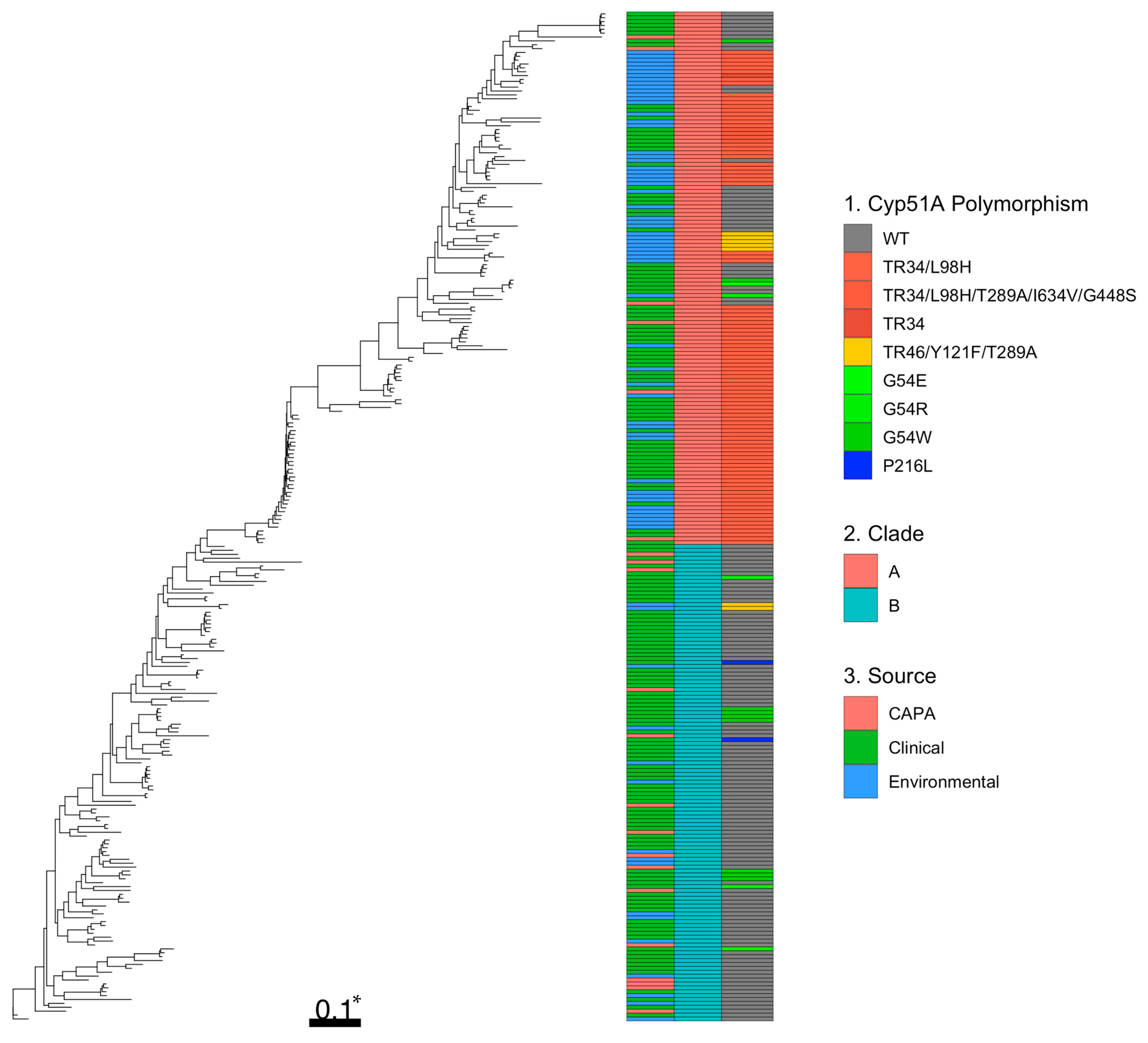
3.2. Azole Resistance within CAPA Isolates Primarily Centred on Known Polymorphisms within cyp51A
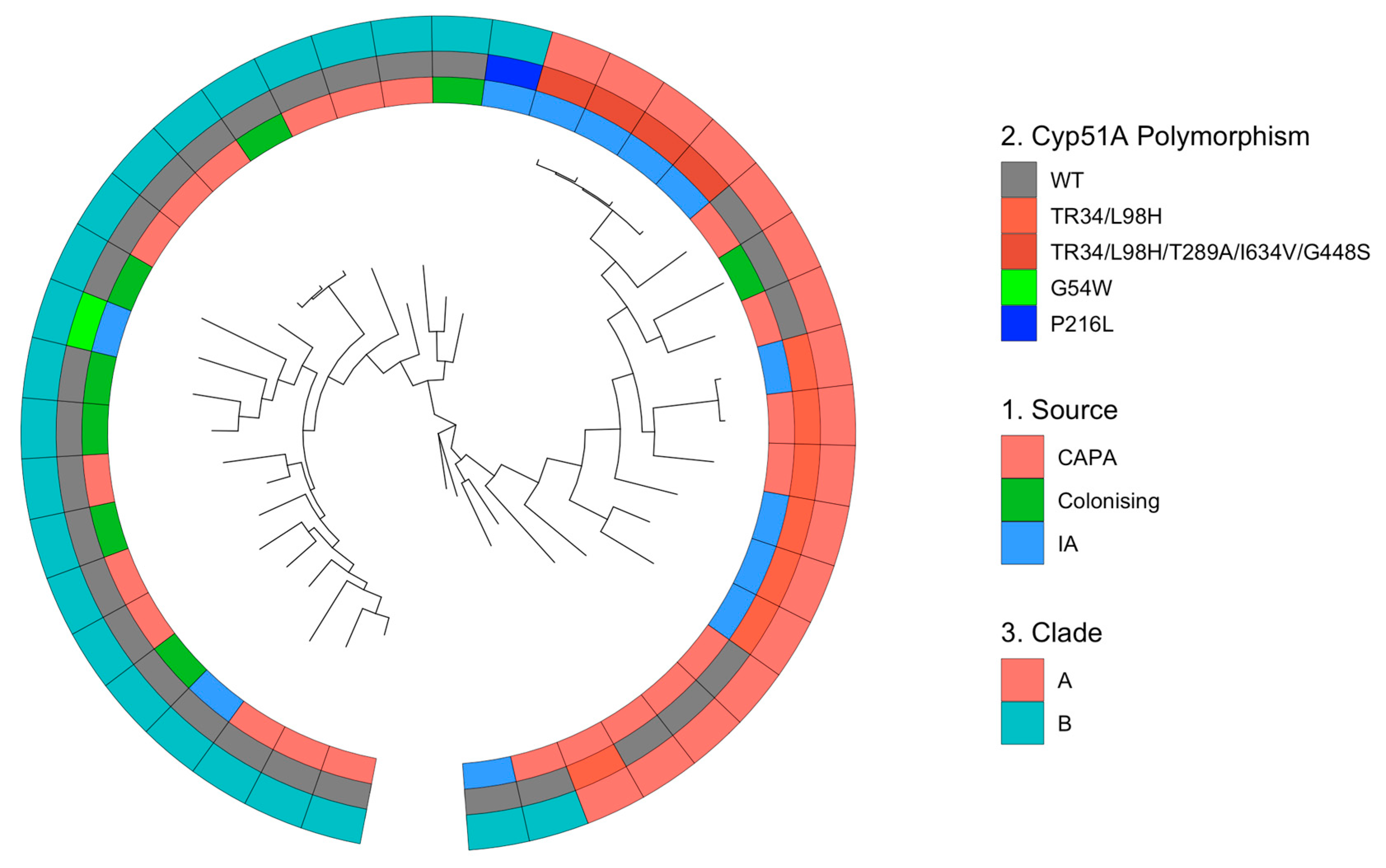
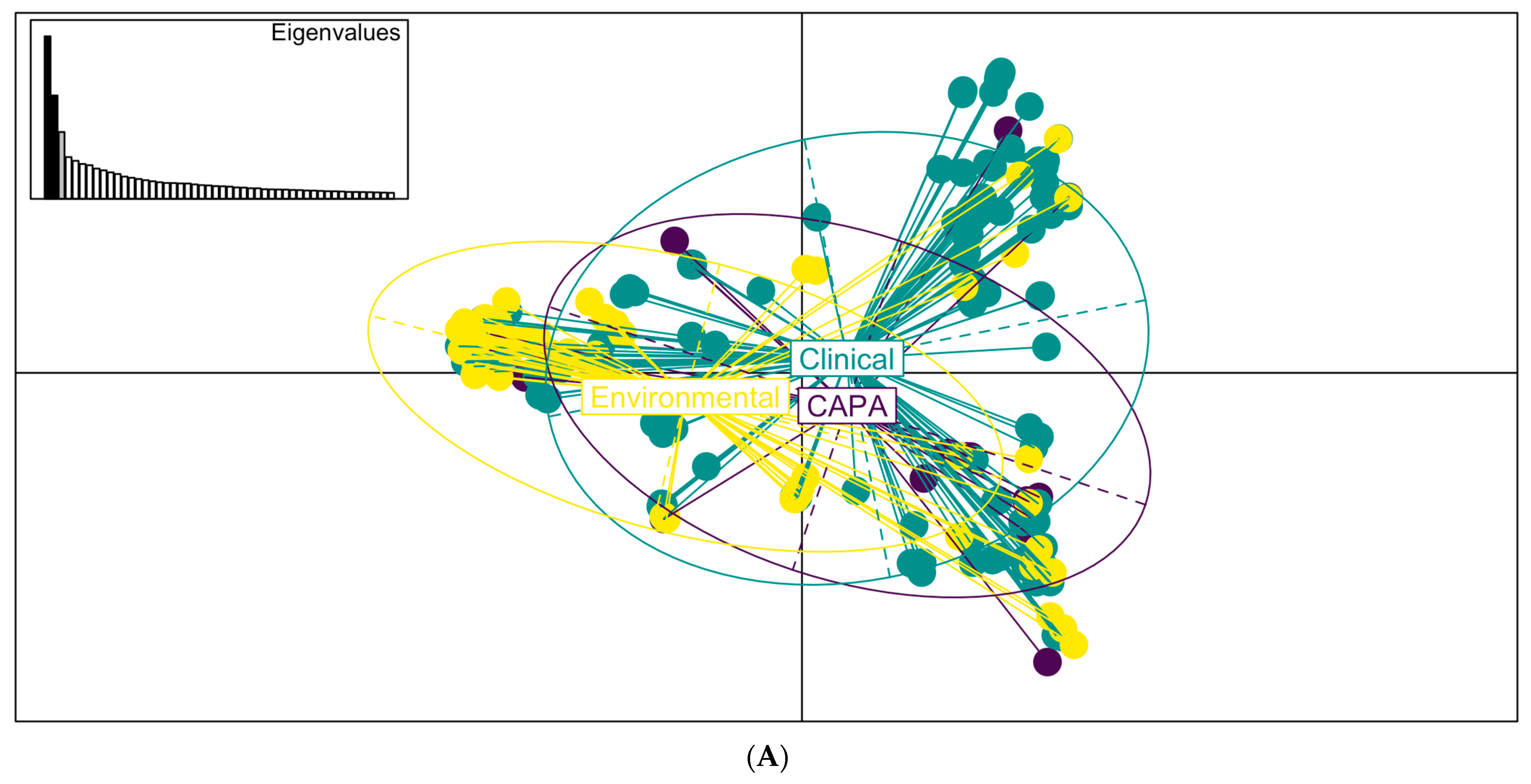
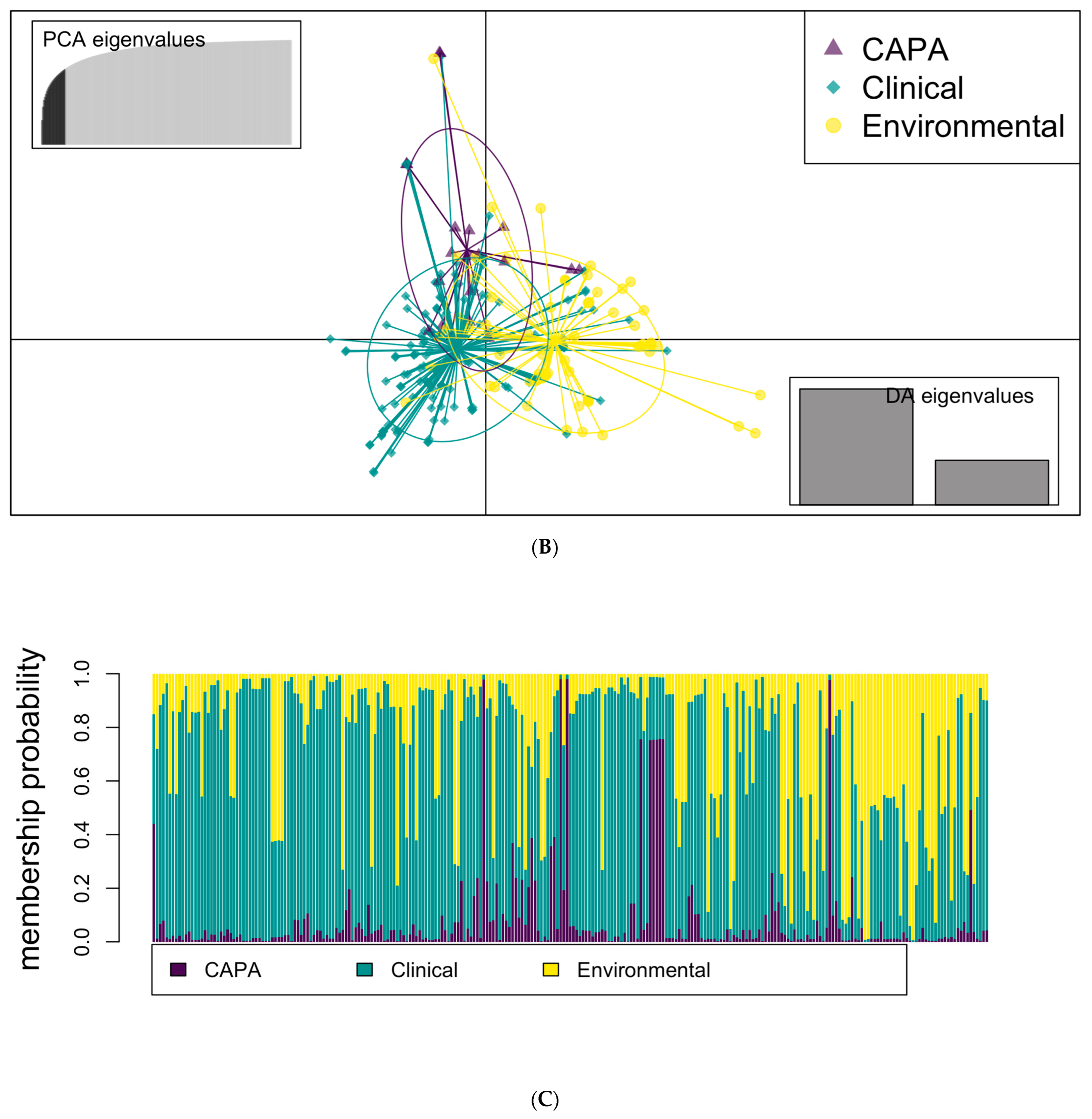
3.3. Phylogenetic and Spatial Analysis Shows CAPA Isolates Are Highly Related to Non-CAPA Clinical and Environmental A. fumigatus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Polymorphism | Reference |
|---|---|---|
| Afu4g06890—cyp51A | TR34 | [32] |
| TR34/L98H | [15,16,17,23,30,32,56,64,65,66,67,68,69,70,71,72,73] | |
| TR34/L98H/S297T/F495I | [15,18] | |
| TR34/L98H/T289A/I364V/G448S | [16,32] | |
| TR46/Y121F/T289A | [16,17,30,32,64,66,68,69,74] | |
| TR46/L98H/T289A/I364V/G448S | [16] | |
| (TR46)2/Y121F/M172V/T289A/G448S | [16] | |
| TR53 | [16] | |
| TR120 | [16] | |
| F46Y/M172V/N248T/D255E/E427K | [15,65,75,76] | |
| F46Y/M172V/E427K | [23] | |
| G138C | [77,78] | |
| Y121F | [18] | |
| Y431C | [75,77,78] | |
| G434C | [77,78] | |
| G432S | [18] | |
| M220L | [17,75] | |
| M220T | [30,79] | |
| M220R | [18,80] | |
| M220I | [18,31,79] | |
| M220K | [18,67] | |
| M220V | [81] | |
| N248K | [15,67,72,82] | |
| A284T | [30,75] | |
| H285Y | [75,77] | |
| G54E | [30,32,67,75] | |
| G54R | [18,30,31,32,72,75,80] | |
| G54W | [18,30,31,32,65,81] | |
| G54V | [30,80] | |
| G448S | [18,30,81] | |
| F219V | [30] | |
| F219I | [30,65,75] | |
| F219L | [18] | |
| F219C | [81] | |
| P216H | [30] | |
| P216S | [18] | |
| P216L | [32,65,75,77,80,81] | |
| P216A | [31] | |
| H147Y | [77] | |
| I242V | [16,82] | |
| I266N | [31] | |
| (TR130)3/D430G | [16] | |
| Afu3g09850—MshA | E812G | [28] |
| A45T | ||
| P329T | ||
| E467N | ||
| Afu6g05300—hapE | P88L | [83] |
| Afu7g01960—Unknown transcription factor | 167* | [84] |
| Afu2g03700—hmg1 | L273F | [81] |
| Afu2g03700—hmg1 | S305P/M220I | [81] |
| Afu4g06890—cyp51A | ||
| Afu2g03700—hmg1 | A640V/A9T | [31] |
| Afu4g06890—cyp51A | ||
| Afu2g03700—hmg1 | H237Y/N157S | [31] |
| Afu4g03630—erg6 | ||
| Afu2g03700—hmg1 | S269C/G54R | [31] |
| Afu4g06890 –cyp51A | ||
| Afu4g03630—erg6 | W320G | [83] |
| Afu4g04820—erg25 | W218 * | |
| Afu4g11240—aarA | F481S | |
| Afu5g07960—C2H2 | Y347 * | |
| Afu1g17440—ABC drug transporter | Y1149N | [32] |
| Afu1g17440—ABC drug transporter | Over expression | [85] |
| Afu2g14250—NctA | Loss-of-function mutations | [29] |
| Afu3g02340—NctB | Loss-of-function mutations | |
| Afu2g16260—microtubule bundle protein | Unknown | [27] |
| Afu4g04680—FGGY-family kinase | Unknown | |
| Afu4g04960—uncharacterized ORF | Unknown |
| Isolate ID | City of Origin | Source | Number of Aligned Reads (Millions) | Percentage of Reference Genome Covered (%) | Mean Depth of Coverage (X) |
|---|---|---|---|---|---|
| C1 | Leeds | Clinical | 54.6 | 97.1 | 189.3 |
| C2 | Leeds | Clinical | 50.8 | 96.0 | 175.4 |
| C3 | Leeds | Clinical | 45.5 | 96.4 | 157.3 |
| C4 | Leeds | Clinical | 45.7 | 98.2 | 158.8 |
| C5 | Leeds | Clinical | 46.6 | 99.6 | 167.8 |
| C6 | Leeds | Clinical | 52.6 | 97.4 | 185.2 |
| C34 | Dublin | Clinical | 6.6 | 97.1 | 34.6 |
| C35 | Dublin | Clinical | 6.1 | 97.5 | 31.6 |
| C36 | Dublin | Clinical | 6.5 | 97.3 | 33.6 |
| C37 | Dublin | Clinical | 6.6 | 98.5 | 34.5 |
| C38 | Dublin | Clinical | 6.6 | 98.6 | 34.7 |
| C39 | Dublin | Clinical | 7.1 | 97.2 | 37.1 |
| C40 | Dublin | Clinical | 7.1 | 97.7 | 37.1 |
| C41 | Dublin | Clinical | 6.9 | 98.0 | 36.4 |
| C42 | Dublin | Clinical | 7.3 | 98 | 38.3 |
| C43 | Dublin | Clinical | 7.7 | 97.8 | 40.4 |
| C44 | Dublin | Clinical | 6.9 | 97 | 34.7 |
| C45 | Dublin | Clinical | 6.7 | 97 | 34.5 |
| C46 | Dublin | Clinical | 6.1 | 97.6 | 31.8 |
| C47 | Dublin | Clinical | 1.9 | 99.6 | 33.1 |
| C48 | Dublin | Clinical | 6.5 | 95.3 | 33.8 |
| C49 | Dublin | Clinical | 7.1 | 96.3 | 36.5 |
| C50 | Dublin | Clinical | 7.1 | 96.4 | 36.8 |
| C51 | Dublin | Clinical | 7.6 | 97.8 | 40.1 |
| C52 | Dublin | Clinical | 8.4 | 98.1 | 44.1 |
| C53 | Dublin | Clinical | 7.5 | 96.8 | 38.5 |
| C54 | Dublin | Clinical | 7.5 | 98.2 | 39 |
| C55 | Dublin | Clinical | 7.0 | 97.1 | 36.2 |
| C56 | Dublin | Clinical | 6.6 | 97.2 | 34.1 |
| C57 | Dublin | Clinical | 8.0 | 97.1 | 41.3 |
| C58 | Dublin | Clinical | 7.3 | 97.1 | 37.7 |
| C59 | Dublin | Clinical | 7.6 | 97.7 | 39.5 |
| C60 | Dublin | Clinical | 6.1 | 97 | 32 |
| C61 | Dublin | Clinical | 7.3 | 97.5 | 38.3 |
| C62 | Dublin | Clinical | 6.5 | 97.3 | 34.3 |
| C63 | Dublin | Clinical | 6.9 | 97.5 | 36.3 |
| C64 | Dublin | Clinical | 7.0 | 96.8 | 36.7 |
| C65 | Dublin | Clinical | 7.1 | 96.8 | 37.1 |
| C66 | Dublin | Clinical | 7.1 | 97.3 | 37 |
| C67 | Dublin | Clinical | 7.3 | 97.25 | 38.4 |
| C68 | Dublin | Clinical | 6.6 | 97 | 34.3 |
| C69 | Dublin | Clinical | 6.3 | 97.3 | 32.9 |
| C70 | Dublin | Clinical | 7.0 | 97.4 | 36.1 |
| C71 | Dublin | Clinical | 6.4 | 96.9 | 33.3 |
| C72 | Dublin | Clinical | 6.5 | 97.6 | 32.6 |
| C73 | Dublin | Clinical | 7.4 | 97.7 | 39 |
| C74 | Dublin | Clinical | 7.3 | 98.1 | 38.3 |
| C75 | Dublin | Clinical | 7.7 | 97.6 | 40.3 |
| C76 | Dublin | Clinical | 8.5 | 97.6 | 44.2 |
| C77 | Dublin | Clinical | 8.7 | 97.2 | 45 |
| C78 | Dublin | Clinical | 6.6 | 97.1 | 34.4 |
| C79 | Dublin | Clinical | 8.2 | 97.8 | 42.6 |
| C80 | Dublin | Clinical | 6.8 | 97.4 | 35.1 |
| C81 | Edinburgh | Clinical | 5.9 | 97.6 | 29.6 |
| C82 | Brighton | Clinical | 6.3 | 97.5 | 32 |
| C85 | London | Clinical | 5.8 | 97.2 | 29.2 |
| C86 | Edinburgh | Clinical | 5.7 | 97.7 | 29.1 |
| C87 | Edinburgh | Clinical | 5.6 | 97.5 | 28.6 |
| C88 | London | Clinical | 6.0 | 97.3 | 30.1 |
| C89 | London | Clinical | 6.5 | 97 | 33 |
| C90 | Cambridge | Clinical | 6.8 | 97.1 | 34 |
| C91 | Liverpool | Clinical | 6.1 | 97.4 | 30.4 |
| C92 | Cardiff | Clinical | 6.3 | 97.7 | 31.8 |
| C93 | Bristol | Clinical | 6.6 | 97.3 | 33.1 |
| C94 | London | Clinical | 6.5 | 97.8 | 32.9 |
| C95 | London | Clinical | 6.2 | 97.1 | 31.4 |
| C96 | Edinburgh | Clinical | 6.6 | 97.7 | 33.2 |
| C97 | London | Clinical | 6.2 | 97 | 31.2 |
| C99 | Cardiff | Clinical | 6.1 | 98.4 | 31.1 |
| C100 | Birmingham | Clinical | 6.3 | 98.4 | 32.3 |
| C101 | Cambridge | Clinical | 6.2 | 97.3 | 31.3 |
| C102 | Leicester | Clinical | 6.3 | 97.6 | 31.9 |
| C103 | London | Clinical | 6.3 | 98.5 | 32.5 |
| C104 | London | Clinical | 6.5 | 97.6 | 32.8 |
| C105 | London | Clinical | 6.0 | 97.5 | 30.6 |
| C106 | London | Clinical | 5.8 | 96.3 | 29.6 |
| C107 | London | Clinical | 5.6 | 97.9 | 28.5 |
| C108 | London | Clinical | 5.8 | 96.8 | 29.2 |
| C109 | London | Clinical | 5.8 | 97.6 | 29.7 |
| C110 | London | Clinical | 5.5 | 95.2 | 27.8 |
| C111 | London | Clinical | 5.6 | 97.7 | 28.5 |
| C112 | London | Clinical | 5.8 | 97.8 | 29.5 |
| C113 | London | Clinical | 6.0 | 97.7 | 30.6 |
| C114 | London | Clinical | 6.5 | 97.6 | 32.9 |
| C115 | London | Clinical | 5.9 | 97.8 | 30 |
| C116 | London | Clinical | 6.1 | 97.7 | 30.6 |
| C117 | London | Clinical | 6.4 | 97.7 | 32.5 |
| C118 | London | Clinical | 5.8 | 95.3 | 29.2 |
| C119 | London | Clinical | 5.9 | 95.3 | 29.8 |
| C120 | London | IA (ABPA) | 6.3 | 97 | 31.7 |
| C121 | London | Clinical | 6.3 | 97.5 | 32.2 |
| C122 | London | Clinical | 6.5 | 95.7 | 32.8 |
| C123 | London | Clinical | 6.4 | 97.7 | 32.3 |
| C124 | London | Clinical | 6.5 | 96.9 | 32.5 |
| C125 | London | Clinical | 6.4 | 97.6 | 32.4 |
| C126 | London | Clinical | 6.3 | 97.6 | 31.7 |
| C127 | London | Clinical | 6.2 | 97.0 | 31.7 |
| C128 | Leeds | Clinical | 6.9 | 96.8 | 34.8 |
| C129 | Leeds | Clinical | 7.2 | 96.6 | 35.9 |
| C130 | Leeds | Clinical | 7.3 | 96.5 | 36.6 |
| C131 | Leeds | Clinical | 6.9 | 96.4 | 24.6 |
| C132 | Leeds | Clinical | 7.2 | 96.3 | 35.9 |
| C133 | Leeds | Clinical | 6.8 | 96.2 | 33.9 |
| C134 | Leeds | Clinical | 7.0 | 96.7 | 34.8 |
| C135 | Leeds | Clinical | 6.9 | 96.7 | 34.6 |
| C136 | Leeds | Clinical | 7.0 | 96.6 | 35.1 |
| C137 | London | IA (NAf) | 7.0 | 97.9 | 35.7 |
| C138 | London | IA (NAf) | 6.4 | 97.9 | 32.5 |
| C139 | London | IA (NAf) | 6.3 | 98.2 | 32.3 |
| C140 | London | IA (NAf) | 6.2 | 98.2 | 31.9 |
| C141 | London | Clinical | 6.3 | 96.8 | 31.9 |
| C142 | London | Clinical | 6.3 | 97.1 | 31.9 |
| C143 | London | IA (Trauma) | 6.3 | 96.7 | 31.8 |
| C144 | London | Clinical | 6.6 | 98.4 | 33.8 |
| C145 | London | Clinical | 6.6 | 97.9 | 34.2 |
| C146 | London | Clinical | 6.4 | 96.8 | 32.4 |
| C147 | London | Clinical | 6.4 | 98.2 | 33.2 |
| C148 | London | Clinical | 6.6 | 97.9 | 32.8 |
| C149 | London | Clinical | 6.5 | 96.6 | 33.3 |
| C150 | London | Clinical | 6.5 | 97.7 | 32.6 |
| C151 | London | Clinical | 6.4 | 97.2 | 32.7 |
| C152 | London | Clinical | 6.4 | 97.2 | 36.8 |
| C153 | London | Clinical | 7.3 | 96.5 | 37.3 |
| C154 | London | Clinical | 7.4 | 97.4 | 37.5 |
| C155 | London | Clinical | 7.3 | 98.4 | 39.7 |
| C156 | London | Clinical | 7.7 | 98.4 | 35.6 |
| C157 | London | Clinical | 7.1 | 97.2 | 35 |
| C158 | London | Clinical | 7.0 | 97.7 | 39 |
| C159 | London | Clinical | 7.6 | 98.1 | 36 |
| C160 | London | Clinical | 7.2 | 97.2 | 38.2 |
| C161 | London | Clinical | 7.5 | 98.0 | 29.2 |
| C162 | London | Clinical | 5.8 | 95.6 | 33.6 |
| C163 | London | Clinical | 6.6 | 97.7 | 4 |
| C164 | London | Clinical | 7.2 | 98.2 | 36 |
| C165 | London | Clinical | 6.8 | 96.6 | 34.3 |
| C166 | London | Clinical | 6.8 | 94.7 | 34.5 |
| C167 | London | Clinical | 6.8 | 97.2 | 34.2 |
| C168 | London | Clinical | 7.0 | 98.1 | 36 |
| C169 | London | Clinical | 6.7 | 97.4 | 34.3 |
| C170 | London | Clinical | 7.1 | 98.24 | 36.7 |
| C171 | London | Clinical | 6.6 | 97.8 | 33.2 |
| C172 | London | Clinical | 6.5 | 96.9 | 33.2 |
| C173 | London | Clinical | 6.8 | 96.6 | 34.1 |
| C191 | Dublin | Clinical | 9.1 | 97.0 | 46.5 |
| C192 | Dublin | Clinical | 9.9 | 97.2 | 49.9 |
| C193 | Dublin | Clinical | 8.9 | 97.6 | 45.2 |
| C194 | Dublin | Environmental | 8.9 | 97.2 | 45.5 |
| C195 | Dublin | Environmental | 9.3 | 96.2 | 47 |
| C196 | Dublin | Clinical | 10.3 | 96.1 | 51.7 |
| C197 | Dublin | Environmental | 8.7 | 96.3 | 44.1 |
| C198 | Dublin | Clinical | 9.1 | 96.0 | 45.8 |
| C199 | Dublin | Clinical | 8.4 | 97.7 | 42.3 |
| C200 | Dublin | Clinical | 9.2 | 96.8 | 46.7 |
| C307 | UK | IA (IPA) | 5.7 | 97.3 | 21.6 |
| C323 | UK | IA (ABPA and asthma) | 6.1 | 97.3 | 23.5 |
| C360 | UK | IA (ABPA) | 2.5 | 97.7 | 8.9 |
| C372 | UK | IA (CPA) | 7.1 | 98.2 | 29.5 |
| C376 | UK | IA (ABPA and asthma) | 6.9 | 98.0 | 27.8 |
| C402 | Netherlands | COVID—Colonising | 31.3 | 97.5 | 139.1 |
| C404 | Netherlands | COVID—Colonising | 49.4 | 98.3 | 216.4 |
| C405 | Netherlands | COVID—Colonising | 26.1 | 98.3 | 117.0 |
| C406 | Netherlands | COVID—Colonising | 28.3 | 97.1 | 124.2 |
| C407 | Netherlands | COVID—Colonising | 37.7 | 98.6 | 166.3 |
| C409 | Netherlands | COVID—Colonising | 33.1 | 97.9 | 147.3 |
| C410 | Netherlands | COVID—Colonising | 29.9 | 98.3 | 132.0 |
| C426 | Ireland | Environmental | 10.7 | 97.7 | 43.0 |
| C427 | Ireland | Environmental | 11.4 | 97.6 | 44.9 |
| C428 | Ireland | Environmental | 10.5 | 97.8 | 41.6 |
| C429 | Ireland | Environmental | 9.7 | 97.3 | 38.1 |
| C430 | Ireland | Environmental | 11.7 | 97.2 | 44.8 |
| C431 | Ireland | Environmental | 11.6 | 97.8 | 44.7 |
| C432 | Ireland | Environmental | 9.1 | 97.7 | 37.1 |
| C433 | Ireland | Environmental | 1.0 | 97.8 | 4.5 |
| C434 | Ireland | Clinical | 10.6 | 96.6 | 41.9 |
| C442 | Ireland | IA (IPA) | 11.9 | 97.8 | 45.6 |
| C443 | Ireland | COVID—Colonising | 9.7 | 97.5 | 39.2 |
| U1 | Dublin | Clinical | 6.2 | 97.9 | 32.3 |
| U2 | Dublin | Clinical | 7.4 | 97.63 | 38.5 |
| U3 | Dublin | Clinical | 7.5 | 97.1 | 39.2 |
| E9 | Wales | Environmental | 6.1 | 97.6 | 31.8 |
| E10 | Wales | Environmental | 7.2 | 97.8 | 38 |
| E11 | Wales | Environmental | 6.5 | 97.7 | 32.7 |
| E12 | Wales | Environmental | 6.2 | 96.6 | 32.2 |
| E13 | Wales | Environmental | 6.7 | 97.4 | 34.7 |
| E14 | Wales | Environmental | 7.3 | 98.0 | 38.4 |
| E15 | Wales | Environmental | 6.4 | 96.9 | 33.5 |
| E16 | Wales | Environmental | 6.8 | 97.5 | 35.5 |
| E17 | Wales | Environmental | 7.0 | 97.1 | 36.5 |
| E18 | Wales | Environmental | 7.4 | 97.0 | 38.6 |
| E19 | Wales | Environmental | 6.9 | 97.5 | 35.9 |
| E20 | Wales | Environmental | 7.2 | 97.0 | 37 |
| E21 | Wales | Environmental | 7.6 | 95.6 | 39.6 |
| E22 | Wales | Environmental | 7.4 | 97.8 | 37.1 |
| E23 | Wales | Environmental | 6.8 | 96.4 | 35.1 |
| E24 | Wales | Environmental | 7.5 | 97.2 | 39 |
| E25 | Wales | Environmental | 6.2 | 99.6 | 33.3 |
| E26 | Wales | Environmental | 6.4 | 96.3 | 33.4 |
| E27 | Wales | Environmental | 7.0 | 97.7 | 36.5 |
| E28 | Wales | Environmental | 6.9 | 97.7 | 35.6 |
| E29 | Wales | Environmental | 0.8 | 99.7 | 46.5 |
| E30 | Wales | Environmental | 7.4 | 97.4 | 38.5 |
| E31 | Wales | Environmental | 6.1 | 97.7 | 32 |
| E32 | Wales | Environmental | 7.0 | 98.6 | 37 |
| E33 | Wales | Environmental | 5.3 | 98.2 | 26.6 |
| E34 | Wales | Environmental | 6.2 | 98.2 | 32.6 |
| E35 | Wales | Environmental | 5.7 | 97.6 | 30.2 |
| E36 | Wales | Environmental | 5.8 | 97.3 | 30.4 |
| E37 | Wales | Environmental | 5.9 | 97.0 | 30.5 |
| E38 | Wales | Environmental | 6.4 | 97.6 | 33.2 |
| E39 | Dublin | Environmental | 8.1 | 97.2 | 42.5 |
| E40 | Dublin | Environmental | 8.1 | 96.7 | 41.6 |
| E41 | Dublin | Environmental | 7.8 | 97.1 | 40.6 |
| E42 | Dublin | Environmental | 7.1 | 97.5 | 36.7 |
| E43 | Dublin | Environmental | 7.2 | 96.6 | 37.1 |
| E44 | Dublin | Environmental | 8.0 | 98.1 | 39.4 |
| E45 | Dublin | Environmental | 7.4 | 96.1 | 37.9 |
| E46 | Dublin | Environmental | 7.3 | 97.9 | 37.7 |
| E48 | Dublin | Environmental | 6.8 | 97.8 | 35.6 |
| E49 | Dublin | Environmental | 7.3 | 97.5 | 38.1 |
| E50 | Dublin | Environmental | 6.6 | 97.2 | 34.5 |
| E52 | Dublin | Environmental | 7.3 | 97.2 | 38 |
| E53 | Dublin | Environmental | 7.6 | 97.8 | 39.2 |
| E81 | Dublin | Environmental | 6.6 | 98.2 | 31.5 |
| E82 | Dublin | Environmental | 7.0 | 98.2 | 33.1 |
| E83 | Dublin | Environmental | 8.1 | 98.3 | 38.1 |
| E84 | Dublin | Environmental | 7.8 | 98.0 | 36.8 |
| E107 | Nottingham | Environmental | 9.7 | 98.0 | 44.1 |
| E133 | Nottingham | Environmental | 9.4 | 98.2 | 43.3 |
| E136 | Yorkshire | Environmental | 8.1 | 96.1 | 37.1 |
| E190 | Didcot | Environmental | 8.5 | 96.5 | 43.3 |
| E191 | London | Environmental | 9.9 | 97.3 | 50.7 |
| E192 | London | Environmental | 9.0 | 97.3 | 45.5 |
| E193 | London | Environmental | 9.1 | 97.9 | 46.9 |
| E194 | London | Environmental | 8.9 | 97.4 | 45.4 |
| E195 | London | Environmental | 8.6 | 96.6 | 43.9 |
| E201 | Aberdeen | Environmental | 1.6 | 99.7 | 48.9 |
| E202 | Aberdeen | Environmental | 10.0 | 97.8 | 50.5 |
| E203 | Aberdeen | Environmental | 10.0 | 97.1 | 50.8 |
| E204 | Aberdeen | Environmental | 10.2 | 97.9 | 52.3 |
| E205 | Aberdeen | Environmental | 9.6 | 94.5 | 49.9 |
| E206 | Aberdeen | Environmental | 9.3 | 97.5 | 47.1 |
| Number of Isolate Types (%) | χ2-test p-Value | |||
|---|---|---|---|---|
| CAPA (n = 21) | Clinical (n = 167) | Environmental (n = 73) | ||
| Clade | ||||
| A | 6 (28.6) | 75 (44.9) | 56 (76.7) | 0.002 * |
| B | 15 (71.4) | 92 (55.1) | 17 (23.3) | 0.001 * |
| MAT idiomorph | ||||
| MAT1-1 | 11 (52.4) | 73 (43.7) | 18 (24.7) | 0.056 |
| MAT1-2 | 10 (47.6) | 94 (56.3) | 55 (75.3) | 0.158 |
| Cyp51A genotype | ||||
| TR34/L98H | 3 (14.3) | 47 (28.1) | 42 (57.5) | <0.001 * |
| TR34 | 0 | 0 | 1 (1.4) | 0.041 * |
| TR34/L98H/T289A/I364V/G448S | 0 | 4 (2.4) | 0 | 0.091 |
| TR46/Y121F/T289A | 0 | 0 | 7 (9.6) | <0.001 * |
| P216L | 0 | 2 (1.2) | 0 | 0.093 |
| G54W | 0 | 9 (5.4) | 0 | 0.030 * |
| G54E | 0 | 1 (0.6) | 0 | 0.041 * |
| G54R | 0 | 3 (1.8) | 1 (1.4) | 0.295 |
| WT | 18 (85.7) | 101 (60. 5) | 22 (30.1) | 0.002 * |
| Resistance to >= 1 azole | ||||
| Yes | 3 (14.3) | 76 (45.5) | 43 (58.9) | 0.026 * |
| No | 18 (85.7) | 88 (52.7) | 19 (26.0) | <0.001 * |
| Test not performed | 0 | 3 (1.8) | 11 (15.1) | <0.001 * |
| Isolate ID | ITR MIC (mg/L) | VOR MIC (mg/L) | POS MIC (mg/L) | TR | Gene of A. fumigatus Amino Acid Substitution | |||
|---|---|---|---|---|---|---|---|---|
| Afu4g06890 | Afu1g17440 | AFu7g01960 | Afu4g03630 | |||||
| C438* | >16 | 4 | 2 | TR34 | L98H | E581A/Y1149N | WT | WT |
| C441* | 8 | 4 | 0.5 | TR34 | L98H | E581A/Y1149N | WT | WT |
| C444 | >32 | 2 | 1 | TR34 | L98H | E581A/Y1149N | I149V/ P160S | WT |
| C72 | 0.12 | 0.03 | 8 | WT | WT | G166A | I149V/ P160S | A281V |
| C99 | 1 | 2 | ND | WT | WT | G166A | I149V/ P160S | A281V |
| C100 | 8 | 1 | ND | WT | WT | N1073H | I149V/ P160S | WT |
| C109 | >16 | 0.25 | 0.125 | WT | WT | G166A | I149V/ P160S | WT |
| C110 | >16 | 2 | 1 | WT | WT | G166A | I149V/ P160S | WT |
| C111 | 8 | 2 | 0.5 | WT | WT | G166A | I149V/ P160S | A281V |
| C118 | >16 | 2 | 2 | WT | WT | G166A | I149V/ P160S | WT |
| C119 | >16 | 2 | 2 | WT | WT | G166A | I149V/ P160S | WT |
| C121 | >16 | 2 | 1 | WT | WT | G166A | I149V/ P160S | A281V |
| C130 | 2 | 0.125 | 0.03 | WT | WT | H222R/R645C | I149V/ P160S | WT |
| C144 | 2 | 2 | 1 | WT | WT | E581A/Y1149N | I149V/ P160S | WT |
| C146 | 2 | 1 | 0.125 | WT | WT | V1439L | I149V/ P160S | WT |
| C147 | 2 | 0.5 | 0.125 | WT | WT | G166A | I149V/ P160S | WT |
| C150 | 2 | 0.25 | 0.03 | WT | WT | G166A | I149V/ P160S | WT |
| C3 | 1 | 0.25 | 0.06 | WT | WT | WT | A87S | WT |
| C4 | 1 | 0.5 | 0.25 | WT | WT | WT | A87S | WT |
| C5 | 0.5 | 0.125 | 0.06 | WT | WT | WT | P160S | WT |
| C6 | 0.5 | 0.5 | 0.06 | WT | WT | E581A/S715G/Y1149N | WT | WT |
| C36 | <0.03 | 0.06 | 0.03 | WT | WT | N1073H | I149V/P160S | WT |
| C37 | 0.06 | 0.06 | 0.06 | WT | WT | T743S/Y1149N | WT | WT |
| C39 | 0.25 | 0.125 | 0.06 | WT | WT | N1073H | I149V/P160S | A281V |
| C58 | 0.12 | 0.12 | 0.015 | WT | WT | G166A | I149V/P160S | A281V |
| C59 | 0.06 | 0.06 | 0.03 | WT | WT | G166A | I149V/P160S | A281V |
| C73 | 0.03 | 0.12 | 0.015 | WT | WT | G166A | I149V/P160S | A281V |
| C74 | 0.03 | 0.12 | 0.015 | WT | WT | G166A | I149V/P160S | WT |
| C75 | 0.03 | 0.12 | 0.015 | WT | WT | G166A | I149V/P160S | WT |
| C76 | 0.25 | 0.25 | 0.12 | WT | WT | G166A | I149V/P160S | WT |
| C77 | 0.25 | 0.25 | 0.06 | WT | WT | G166A | I149V/P160S | WT |
| C95 | ND | 0.25 | ND | WT | WT | G166A/V1439L | I149V/P160S | WT |
| C148 | 1 | 0.25 | 0.06 | WT | WT | N1073H | I149V/P160S | A281V |
| C154 | 0.06 | 0.06 | 0.03 | WT | WT | G166A | I149V/P160S | WT |
| C158 | 0.06 | 0.06 | 0.03 | WT | WT | G166A | I149V/P160S | WT |
| C161 | 0.06 | 0.06 | 0.03 | WT | WT | G166A | I149V/P160S | A281V |
| C169 | 0.06 | 0.06 | 0.03 | WT | WT | G166A | I149V/P160S | A281V |
| C360 | 0.06 | 0.3 | 0.015 | WT | WT | G166A/N1073H | I149V/P160S | A281V |
| C376 | <0.015 | <0.015 | <0.015 | WT | WT | G166A | I149V/P160S | WT |
| C402 | ND | ND | ND | WT | WT | N1073 | I149V/P160S | WT |
| C403 | 0.25 | 0.25 | 0.063 | WT | WT | R645C/H222R | WT | WT |
| C404 | 0.25 | 0.5 | 0.063 | WT | WT | N1073 | P160S | WT |
| C405 | 0.25 | 0.25 | 0.063 | WT | WT | G166A | P160S | WT |
| C406 | 0.5 | 0.5 | 0.125 | WT | WT | G166A | I149V/P160S | WT |
| C407 | 0.25 | 0.5 | 0.063 | WT | WT | N1073 | I149V/P160S | WT |
| C408 | 0.125 | 0.5 | 0.031 | WT | WT | N1073 | I149V/P160S | WT |
| C409 | 0.5 | 0.25 | 0.125 | WT | WT | N1073 | I149V/P160S | WT |
| C410 | 0.5 | 0.5 | 0.063 | WT | WT | N1073 | I149V/P160S | WT |
| C442 | ND | ND | ND | TR34 | L98H | E581A/Y1149N | WT | WT |
| Isolate ID | ITR MIC (mg/L) | VOR MIC (mg/L) | POS MIC (mg/L) | TEB Score | Resistance Marker | Clade Membership | Mating Type |
|---|---|---|---|---|---|---|---|
| C1 | >16 | 1 | 0.5 | 3 | TR34/L98H | A | MAT1-1 |
| C2 | >16 | 1 | 0.5 | 2 | TR34/L98H | A | MAT1-1 |
| C3 | 1 | 0.25 | 0.06 | ND | WT | B | MAT1-2 |
| C4 | 1 | 0.5 | 0.25 | 1 | WT | A | MAT1-1 |
| C5 | 0.5 | 0.125 | 0.06 | ND | WT | B | MAT1-2 |
| C6 | 0.5 | 0.5 | 0.06 | 1 | WT | A | MAT1-1 |
| C34 | >16 | 0.5 | 0.5 | 1 | TR34/L98H | A | MAT1-2 |
| C35 | >16 | 1 | 0.5 | ND | TR34/L98H | A | MAT1-2 |
| C36 | <0.03 | 0.06 | 0.03 | ND | WT | B | MAT1-1 |
| C37 | 0.06 | 0.06 | 0.06 | ND | WT | A | MAT1-1 |
| C38 | 0.06 | 0.25 | 0.06 | 1 | WT | A | MAT1-1 |
| C39 | 0.25 | 0.125 | 0.06 | ND | WT | B | MAT1-1 |
| C40 | 0.125 | 0.25 | 0.06 | ND | WT | A | MAT1-1 |
| C41 | 0.06 | 0.25 | 0.06 | ND | WT | B | MAT1-2 |
| C42 | 0.06 | 0.25 | 0.03 | ND | WT | B | MAT1-1 |
| C43 | 0.25 | 0.25 | 0.25 | ND | WT | B | MAT1-1 |
| C44 | 0.25 | 0.25 | 0.12 | ND | WT | B | MAT1-2 |
| C45 | 0.03 | 0.12 | 0.015 | ND | WT | B | MAT1-2 |
| C46 | 0.03 | 0.12 | 0.015 | ND | WT | B | MAT1-2 |
| C47 | 0.03 | 0.12 | 0.015 | ND | WT | B | MAT1-2 |
| C48 | 0.03 | 0.12 | 0.015 | ND | WT | B | MAT1-2 |
| C49 | 0.03 | 0.015 | 0.015 | ND | WT | B | MAT1-2 |
| C50 | 0.03 | 0.015 | 0.015 | ND | WT | B | MAT1-1 |
| C51 | 0.12 | 0.25 | 0.06 | ND | WT | B | MAT1-1 |
| C52 | 0.25 | 0.12 | 0.06 | ND | WT | B | MAT1-1 |
| C53 | 0.06 | 0.25 | 0.03 | ND | WT | B | MAT1-1 |
| C54 | <0.015 | 0.12 | 0.03 | ND | WT | B | MAT1-2 |
| C55 | 0.25 | 0.12 | 0.06 | ND | WT | B | MAT1-2 |
| C56 | 0.06 | 0.25 | 0.03 | ND | WT | B | MAT1-2 |
| C57 | 0.06 | 0.06 | 0.03 | ND | WT | B | MAT1-2 |
| C58 | 0.12 | 0.12 | 0.015 | ND | WT | B | MAT1-1 |
| C59 | 0.06 | 0.06 | 0.03 | ND | WT | B | MAT1-1 |
| C60 | 0.03 | 0.12 | 0.015 | ND | WT | A | MAT1-1 |
| C61 | 0.25 | 0.12 | 0.06 | ND | WT | A | MAT1-1 |
| C62 | 0.12 | 0.12 | 0.03 | ND | WT | A | MAT1-1 |
| C63 | 0.03 | 0.25 | 0.015 | ND | WT | A | MAT1-1 |
| C64 | 0.03 | 0.25 | 0.015 | ND | WT | A | MAT1-1 |
| C65 | 0.03 | 0.12 | 0.015 | ND | WT | A | MAT1-1 |
| C66 | 0.25 | 0.25 | 0.12 | ND | WT | A | MAT1-1 |
| C67 | 0.03 | 0.12 | 0.015 | ND | WT | A | MAT1-1 |
| C68 | 0.03 | 0.12 | 0.015 | ND | WT | A | MAT1-1 |
| C69 | 0.25 | 0.25 | 0.06 | ND | TR34/L98H | A | MAT1-2 |
| C70 | 1 | 0.5 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| C71 | 1 | 1 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| C72 | 0.12 | 0.03 | 8 | ND | WT | B | MAT1-1 |
| C73 | 0.03 | 0.12 | 0.015 | ND | WT | B | MAT1-1 |
| C74 | 0.03 | 0.12 | 0.015 | ND | WT | B | MAT1-2 |
| C75 | 0.03 | 0.12 | 0.015 | ND | WT | B | MAT1-2 |
| C76 | 0.25 | 0.25 | 0.12 | ND | WT | A | MAT1-1 |
| C77 | 0.25 | 0.25 | 0.06 | ND | WT | B | MAT1-1 |
| C78 | 4 | 2 | 1 | ND | TR34/L98H | A | MAT1-2 |
| C79 | 16 | 2 | 0.5 | ND | TR34/L98H | A | MAT1-2 |
| C80 | 4 | 4 | 0.5 | ND | TR34/L98H | A | MAT1-2 |
| C81 | 2 | 2 | 0.5 | ND | TR34/L98H | A | MAT1-1 |
| C82 | 16 | 4 | ND | ND | TR34/L98H | A | MAT1-2 |
| C85 | 1 | 2 | ND | ND | TR34/L98H | A | MAT1-2 |
| C86 | >16 | 0.5 | ND | ND | TR34/L98H | A | MAT1-1 |
| C87 | >16 | 4 | ND | 1 | TR34/L98H | A | MAT1-2 |
| C88 | 2 | 4 | ND | 1 | TR34/L98H | A | MAT1-2 |
| C89 | 1 | 4 | ND | 3 | TR34/L98H | A | MAT1-2 |
| C90 | 16 | 1 | ND | 4 | TR34/L98H | A | MAT1-2 |
| C91 | >16 | 1 | ND | ND | TR34/L98H | A | MAT1-1 |
| C92 | >16 | 1 | ND | ND | TR34/L98H | A | MAT1-2 |
| C93 | 16 | 2 | ND | ND | TR34/L98H | A | MAT1-1 |
| C94 | 16 | 2 | ND | 4 | TR34/L98H | A | MAT1-1 |
| C95 | ND | 0.25 | ND | ND | WT | B | MAT1-2 |
| C96 | >16 | 2 | ND | ND | TR34/L98H | A | MAT1-1 |
| C97 | 4 | 1 | 0.25 | ND | TR34/L98H | A | MAT1-1 |
| C99 | 1 | 2 | ND | ND | WT | B | MAT1-2 |
| C100 | 8 | 1 | ND | ND | WT | B | MAT1-2 |
| C101 | 16 | 1 | ND | ND | TR34/L98H | A | MAT1-2 |
| C102 | >16 | 0.125 | ND | ND | G54R | B | MAT1-1 |
| C103 | >16 | 0.25 | 0.5 | ND | G54E | A | MAT1-1 |
| C104 | >16 | 2 | ND | ND | TR34/L98H | A | MAT1-2 |
| C105 | >16 | 2 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| C106 | >16 | 0.125 | 4 | ND | G54W | B | MAT1-1 |
| C107 | >16 | 2 | 2 | 4 | TR34/L98H | A | MAT1-2 |
| C108 | >16 | 0.03 | 2 | ND | G54W | B | MAT1-1 |
| C109 | >16 | 0.25 | 0.125 | ND | WT | B | MAT1-2 |
| C110 | >16 | 2 | 1 | ND | WT | B | MAT1-2 |
| C111 | 8 | 2 | 0.5 | 1 | WT | B | MAT1-2 |
| C112 | >16 | 0.125 | 8 | ND | G54W | B | MAT1-2 |
| C113 | >16 | 0.125 | 16 | ND | G54W | B | MAT1-2 |
| C114 | >16 | 0.125 | 8 | ND | G54W | B | MAT1-2 |
| C115 | >16 | 2 | 2 | ND | TR34/L98H | A | MAT1-2 |
| C116 | 16 | 2 | 1 | ND | TR34/L98H | A | MAT1-1 |
| C117 | >16 | 0.125 | 16 | ND | G54W | B | MAT1-2 |
| C118 | >16 | 2 | 2 | 3 | WT | B | MAT1-2 |
| C119 | >16 | 2 | 2 | 3 | WT | B | MAT1-2 |
| C120 | >16 | 0.25 | 2 | ND | P216L | B | MAT1-1 |
| C121 | >16 | 2 | 1 | 1 | WT | B | MAT1-2 |
| C122 | >16 | 0.125 | 16 | ND | G54W | B | MAT1-1 |
| C123 | 16 | 1 | 0.25 | ND | L98H | A | MAT1-1 |
| C124 | >16 | 0.06 | 0.25 | ND | G54W | A | MAT1-1 |
| C125 | 16 | 2 | 0.5 | ND | TR34/L98H | A | MAT1-2 |
| C126 | >16 | 2 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| C127 | >16 | 0.125 | 0.5 | ND | P216L | B | MAT1-2 |
| C128 | 0.5 | 0.125 | 0.03 | ND | WT | B | MAT1-2 |
| C129 | 1 | 0.125 | 0.03 | ND | WT | B | MAT1-2 |
| C130 | 2 | 0.125 | 0.03 | ND | WT | B | MAT1-2 |
| C131 | >16 | 0.25 | 0.125 | ND | TR34/L98H | A | MAT1-1 |
| C132 | 16 | 0.25 | 0.125 | ND | TR34/L98H | A | MAT1-1 |
| C133 | >16 | 0.25 | 0.125 | ND | TR34/L98H | A | MAT1-1 |
| C134 | 0.5 | 0.125 | 0.03 | ND | WT | B | MAT1-2 |
| C135 | 1 | 0.125 | 0.03 | ND | WT | B | MAT1-2 |
| C136 | 1 | 0.125 | 0.03 | ND | WT | B | MAT1-2 |
| C137 | 16 | >16 | 4 | 4 | TR34/L98H/T289A/ I364V/G448S | A | MAT1-2 |
| C138 | 16 | >16 | 4 | 4 | TR34/L98H/T289A/ I364V/G448S | A | MAT1-2 |
| C139 | 16 | >16 | 4 | ND | TR34/L98H/T289A/ I364V/G448S | A | MAT1-2 |
| C140 | 16 | >16 | 4 | 4 | TR34/L98H/T289A/ I364V/G448S | A | MAT1-2 |
| C141 | >16 | 2 | 0.5 | 4 | TR34/L98H | A | MAT1-2 |
| C142 | >16 | 2 | 0.5 | 3 | TR34/L98H | A | MAT1-2 |
| C143 | 4 | 0.5 | 0.125 | 4 | TR34/L98H | A | MAT1-1 |
| C144 | 2 | 2 | 1 | ND | WT | A | MAT1-1 |
| C145 | >16 | 1 | >16 | ND | G54W | A | MAT1-1 |
| C146 | 2 | 1 | 0.125 | 3 | WT | B | MAT1-2 |
| C147 | 2 | 0.5 | 0.125 | ND | WT | B | MAT1-1 |
| C148 | 1 | 0.25 | 0.06 | ND | WT | B | MAT1-2 |
| C149 | 0.06 | 0.06 | 0.03 | ND | WT | B | MAT1-1 |
| C150 | 2 | 0.25 | 0.03 | ND | WT | B | MAT1-1 |
| C151 | 0.06 | 0.06 | 0.015 | 1 | WT | B | MAT1-2 |
| C152 | 0.06 | 0.06 | 0.015 | ND | WT | B | MAT1-2 |
| C153 | 16 | 0.5 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| C154 | 0.06 | 0.06 | 0.03 | 1 | WT | B | MAT1-1 |
| C155 | 0.06 | 0.06 | 0.015 | ND | WT | B | MAT1-1 |
| C156 | 0.06 | 0.06 | 0.03 | ND | WT | B | MAT1-1 |
| C157 | 0.06 | 0.06 | 0.03 | ND | WT | B | MAT1-2 |
| C158 | 0.06 | 0.06 | 0.03 | ND | WT | B | MAT1-1 |
| C159 | 0.06 | 0.125 | 0.03 | ND | WT | A | MAT1-1 |
| C160 | 0.06 | 0.06 | 0.03 | ND | WT | B | MAT1-1 |
| C161 | 0.06 | 0.06 | 0.03 | ND | WT | B | MAT1-1 |
| C162 | 0.06 | 0.06 | 0.03 | 4 | WT | B | MAT1-2 |
| C163 | 0.06 | 0.125 | 0.06 | ND | WT | B | MAT1-1 |
| C164 | 0.125 | 0.125 | 0.03 | ND | WT | B | MAT1-2 |
| C165 | 0.125 | 0.06 | 0.03 | 1 | WT | B | MAT1-2 |
| C166 | 0.06 | 0.06 | 0.03 | ND | WT | B | MAT1-2 |
| C167 | 0.06 | 0.06 | 0.03 | ND | WT | B | MAT1-1 |
| C168 | 0.06 | 0.06 | 0.03 | ND | WT | A | MAT1-1 |
| C169 | 0.06 | 0.06 | 0.03 | ND | WT | B | MAT1-2 |
| C170 | 0.125 | 0.06 | 0.03 | ND | WT | A | MAT1-1 |
| C171 | >16 | 0.06 | 0.25 | ND | G54R | B | MAT1-2 |
| C172 | 0.06 | 0.03 | 0.03 | ND | WT | B | MAT1-1 |
| C173 | 0.06 | 0.06 | 0.015 | 3 | WT | B | MAT1-2 |
| C191 | 4 | 2 | 0.5 | ND | TR34/L98H | A | MAT1-2 |
| C192 | 2 | 2 | 0.5 | ND | TR34/L98H | A | MAT1-1 |
| C193 | 0.5 | 2 | 0.5 | ND | TR34/L98H | A | MAT1-2 |
| C194 | ND | ND | ND | ND | TR34/L98H | A | MAT1-1 |
| C195 | ND | ND | ND | ND | TR46/Y121F/T289A | A | MAT1-2 |
| C196 | 32 | 2 | 0.5 | ND | TR34/L98H | A | MAT1-2 |
| C197 | ND | ND | ND | ND | TR46/Y121F/T289A | A | MAT1-2 |
| C198 | ND | ND | ND | ND | WT | B | MAT1-2 |
| C199 | 0.75 | 0.125 | 0.25 | ND | WT | A | MAT1-2 |
| C200 | 0.03 | 0.12 | 0.25 | ND | WT | B | MAT1-2 |
| C307 | ND | ND | ND | 2 | TR34/L98H | A | MAT1-1 |
| C323 | >4 | 0.25 | 0.06 | ND | TR34/L98H | A | MAT1-2 |
| C360 | 0.06 | 0.3 | 0.015 | 1 | WT | B | MAT1-2 |
| C372 | ND | ND | ND | ND | G54W | B | MAT1-2 |
| C376 | <0.015 | <0.015 | <0.015 | 1 | WT | B | MAT1-2 |
| C402 | ND | ND | ND | ND | WT | B | MAT1-1 |
| C403 | 0.25 | 0.25 | 0.063 | ND | WT | B | MAT1-2 |
| C404 | 0.25 | 0.5 | 0.063 | ND | WT | B | MAT1-2 |
| C405 | 0.25 | 0.25 | 0.063 | ND | WT | B | MAT1-2 |
| C406 | 0.5 | 0.5 | 0.125 | ND | WT | B | MAT1-2 |
| C407 | 0.25 | 0.5 | 0.063 | ND | WT | B | MAT1-2 |
| C408 | 0.125 | 0.5 | 0.031 | ND | WT | B | MAT1-1 |
| C409 | 0.5 | 0.25 | 0.125 | ND | WT | B | MAT1-1 |
| C410 | 0.5 | 0.5 | 0.063 | ND | WT | B | MAT1-1 |
| C422 | ND | ND | ND | 1 | WT | B | MAT1-2 |
| C423 | ND | ND | ND | 1 | WT | B | MAT1-1 |
| C424 | ND | ND | ND | 1 | WT | B | MAT1-2 |
| C425 | ND | ND | ND | 1 | WT | A | MAT1-1 |
| C426 | ND | ND | ND | 1 | WT | B | MAT1-1 |
| C427 | ND | ND | ND | 1 | WT | A | MAT1-2 |
| C428 | ND | ND | ND | 1 | WT | B | MAT1-2 |
| C429 | ND | ND | ND | 1 | WT | A | MAT1-1 |
| C430 | ND | ND | ND | 1 | WT | A | MAT1-1 |
| C431 | ND | ND | ND | 1 | WT | B | MAT1-1 |
| C432 | ND | ND | ND | 1 | WT | B | MAT1-2 |
| C433 | ND | ND | ND | 3 | WT | A | MAT1-2 |
| C434 | ND | ND | ND | 1 | WT | A | MAT1-1 |
| C435 | ND | ND | ND | 1 | WT | B | MAT1-2 |
| C436 | ND | ND | ND | 1 | WT | B | MAT1-1 |
| C437 | ND | ND | ND | 1 | WT | B | MAT1-1 |
| C438* | >16 | 4 | 2 | 3 | TR34/L98H | A | MAT1-1 |
| C439 | ND | ND | ND | 1 | WT | B | MAT1-2 |
| C440 | ND | ND | ND | 1 | WT | B | MAT1-2 |
| C441* | 8 | 4 | 0.5 | 4 | TR34/L98H | A | MAT1-1 |
| C442 | ND | ND | ND | 3 | TR34/L98H | A | MAT1-1 |
| C443 | ND | ND | ND | 1 | WT | A | MAT1-1 |
| C444 | >32 | 2 | 1 | 3 | TR34/L98H | A | MAT1-1 |
| C611 | ND | ND | ND | 1 | WT | B | MAT1-2 |
| C612 | ND | ND | ND | 1 | WT | B | MAT1-2 |
| CAPA-A* | 0.5 | 0.5 | 1 | ND | WT | A | MAT1-2 |
| CAPA-B* | 0.5 | 0.5 | 1 | ND | WT | B | MAT1-2 |
| CAPA-C* | 0.5 | 0.5 | 1 | ND | WT | B | MAT1-1 |
| CAPA-D* | 0.5 | 0.5 | 1 | ND | WT | B | MAT1-1 |
| E9 | >16 | 2 | 0.12 | 3 | TR34/L98H | A | MAT1-2 |
| E10 | >16 | 2 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| E11 | >16 | 2 | 0.12 | ND | TR34/L98H | A | MAT1-2 |
| E12 | >16 | 8 | <0.5 | 4 | TR34/L98H | B | MAT1-2 |
| E13 | >16 | 2 | 0.12 | 3 | TR34/L98H | A | MAT1-2 |
| E14 | >16 | 2 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| E15 | >16 | 4 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| E16 | >16 | 2 | 0.12 | ND | TR34/L98H | A | MAT1-2 |
| E17 | >16 | 8 | 0.5 | ND | TR34/L98H | A | MAT1-2 |
| E18 | >16 | 2 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| E19 | >16 | 4 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| E20 | >16 | 2 | 0.12 | ND | TR34/L98H | A | MAT1-2 |
| E21 | >16 | >8 | 0.5 | ND | TR34/L98H | A | MAT1-2 |
| E22 | >16 | 8 | <0.5 | ND | TR34/L98H | A | MAT1-2 |
| E23 | >16 | 2 | 0.12 | ND | TR34/L98H | A | MAT1-2 |
| E24 | >16 | 2 | 0.12 | ND | TR34/L98H | A | MAT1-2 |
| E25 | >16 | 2 | 0.12 | ND | TR34/L98H | A | MAT1-2 |
| E26 | >16 | 1 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| E27 | >16 | 2 | <0.25 | ND | TR34/L98H | A | MAT1-2 |
| E28 | >16 | 2 | 0.025 | 3 | TR34/L98H | A | MAT1-2 |
| E29 | >16 | 2 | 0.12 | 3 | TR34/L98H | A | MAT1-2 |
| E30 | >16 | 2 | 0.12 | ND | TR34/L98H | A | MAT1-2 |
| E31 | >16 | 2 | 0.12 | ND | TR34/L98H | A | MAT1-2 |
| E32 | >16 | 4 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| E33 | >16 | 0.25 | 1 | ND | G54R | A | MAT1-2 |
| E34 | >16 | 1 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| E35 | >16 | 4 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| E36 | >16 | 4 | 0.5 | ND | TR34/L98H | A | MAT1-2 |
| E37 | >16 | 2 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| E38 | >16 | 2 | 0.12 | ND | TR34/L98H | A | MAT1-2 |
| E39 | ND | ND | ND | 3 | TR34/L98H | A | MAT1-2 |
| E40 | ND | ND | ND | 4 | TR46/Y121F/T289A | A | MAT1-2 |
| E41 | ND | ND | ND | ND | TR34/L98H | A | MAT1-2 |
| E42 | ND | ND | ND | 4 | TR34/L98H | A | MAT1-2 |
| E43 | ND | ND | ND | 4 | TR34/L98H | A | MAT1-1 |
| E44 | ND | ND | ND | ND | TR46/Y121F/T289A | B | MAT1-1 |
| E45 | ND | ND | ND | 4 | TR34/L98H | A | MAT1-2 |
| E46 | ND | ND | ND | 4 | TR46/Y121F/T289A | A | MAT1-2 |
| E48 | ND | ND | ND | ND | TR46/Y121F/T289A | B | MAT1-1 |
| E49 | ND | ND | ND | ND | TR34/L98H | A | MAT1-2 |
| E50 | ND | ND | ND | ND | TR46/Y121F/T289A | A | MAT1-2 |
| E52 | ND | ND | ND | ND | WT | B | MAT1-1 |
| E53 | ND | ND | ND | ND | WT | B | MAT1-2 |
| E81 | 0.06 | 0.06 | 0.06 | ND | WT | A | MAT1-1 |
| E82 | 0.125 | 0.06 | 0.06 | 1 | WT | A | MAT1-2 |
| E83 | 0.06 | 0.06 | 0.03 | ND | WT | B | MAT1-2 |
| E84 | 0.125 | 0.06 | 0.06 | 1 | WT | A | MAT1-1 |
| E107 | 0.125 | 0.125 | 0.06 | ND | WT | A | MAT1-2 |
| E133 | 0.125 | 0.125 | 0.03 | ND | WT | B | MAT1-2 |
| E136 | 0.125 | 0.125 | 0.06 | ND | WT | B | MAT1-2 |
| E190 | >16 | 0.5 | 0.25 | ND | TR34/L98H | A | MAT1-1 |
| E191 | >16 | 0.5 | 0.25 | ND | TR34/L98H | A | MAT1-2 |
| E192 | 0.06 | 0.06 | 0.03 | ND | WT | B | MAT1-2 |
| E193 | 0.125 | 0.06 | 0.03 | ND | WT | B | MAT1-1 |
| E194 | 0.25 | 0.06 | 0.03 | ND | WT | B | MAT1-2 |
| E195 | 0.25 | 0.06 | 0.03 | ND | WT | B | MAT1-1 |
| E201 | 4 | 0.5 | ND | 4 | TR34/L98H | A | MAT1-1 |
| E202 | 16 | 2 | ND | ND | TR34 | A | MAT1-1 |
| E203 | 4 | 0.25 | ND | ND | TR34/L98H | A | MAT1-1 |
| E204 | 1 | 0.5 | ND | ND | WT | B | MAT1-2 |
| E205 | 4 | 0.5 | ND | ND | TR34/L98H | A | MAT1-2 |
| E206 | 1 | 0.5 | ND | ND | TR34/L98H | A | MAT1-1 |
| U1 | 0.03 | 0.25 | 0.015 | ND | WT | B | MAT1-2 |
| U2 | 0.03 | 0.12 | 0.015 | ND | WT | B | MAT1-2 |
| U3 | 0.06 | 0.25 | 0.03 | ND | WT | B | MAT1-1 |




References
- Brackin, A.; Hemmings, S.; Fisher, M.; Rhodes, J. Fungal Genomics in Respiratory Medicine: What, How and When? Mycopathologia 2021, 186, 589–608. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, A.; Aguado, J.; Arikan-Akdagli, S.; Denning, D.; Groll, A.; Lagrou, K.; Lass-Flörl, C.; Munoz, R.L.P.; Verweij, P.; Warris, A.; et al. Diagnosis and management of Aspergillus diseases: Executive summary of the 2017 ESCMID-ECMM-ERS guideline. Clin. Microbiol. Infect. 2018, 24, e1–e38. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.; Denning, D.; Gow, N.; Levitz, S.; Neta, M.; White, T. Hidden Killers: Human Fungal Infections. Sci. Transl. Med. 2012, 4, 165rv113. [Google Scholar] [CrossRef] [PubMed]
- Bongomin, F.; Gago, S.; Oladele, R.; Denning, D. Global and Multi-National Prevalence of Fungal Diseases—Estimate Precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- Pakzad, R.; Malekifar, P.; Shateri, Z.; Zandi, M.; Rezayat, S.; Soleymani, M.; Karimi, M.; Ahmadi, S.; Shahbahrami, R.; Pakzad, I.; et al. Worldwide prevalence of microbial agents’ coinfection among COVID-19 patients: A comprehensive updated systematic review and meta-analysis. J. Clin. Lab. Anal. 2022, 36, e24151. [Google Scholar] [CrossRef]
- Baddley, J.; Thompson, G.; Chen, S.; White, P.; Johnson, M.; Nguyen, M.; Schwartz, I.; Spec, A.; Ostrosky-Zeichner, L.; Jackson, B.; et al. Coronavirus Disease 2019-Associated Invasive Fungal Infection. Open Forum Infect. Dis. 2021, 16, ofab510. [Google Scholar] [CrossRef]
- Koehler, P.; Bassetti, M.; Chakrabarti, A.; Chen, S.; Colombo, A.; Hoenigl, M.; Klimko, N.; Lass-Florl, C.; Oladale, R.; Vinh, D.; et al. Defining and managing COVID-19-associated pulmonary aspergillosis: The 2020 ECMM/ISHAM consensus criteria for research and clinical guidance. Lancet Infect. Dis. 2021, 21, e149–e162. [Google Scholar] [CrossRef]
- Casalini, G.; Giacomelli, A.; Ridolfo, A.; Gervasoni, C. Invasive Fungal Infections Complicating COVID-19: A Narrative Review. J. Fungi 2021, 7, 921. [Google Scholar] [CrossRef]
- Feys, S.; Almyroudi, M.; Braspenning, R.; Lagrou, K.; Spriet, I.; Dimopoulos, G.; Wauters, J. A Visual and Comprehensive Review on COVID-19-Associated Pulmonary Aspergillosis (CAPA). J. Fungi 2021, 7, 1067. [Google Scholar] [CrossRef]
- Gangneux, J.-P.; Dannaoui, E.; Fekkar, A.; Luyt, C.-E.; Botterel, F.; Prost, N.D.; Tadié, J.-M.; Reizine, F.; Houzé, S.; Timsit, J.-F.; et al. Fungal infections in mechanically ventilated patients with COVID-19 during the first wave: The French multicentre MYCOVID study. Lancet Respir. Med. 2022, 10, 180–190. [Google Scholar] [CrossRef]
- Verweij, P.; Brüggemann, R.; Azoulay, E.; Bassetti, M.; Blot, S.; Buil, J.; Calandra, T.; Chiller, T.; Clancy, C.; Crnely, O.; et al. Taskforce report on the diagnosis and clinical management of COVID-19 associated pulmonary aspergillosis. Intensive Care Med. 2021, 47, 819–834. [Google Scholar] [CrossRef] [PubMed]
- Dellière, S.; Dudoignon, E.; Voicu, S.; Collet, M.; Fodil, S.; Plaud, B.; Chousterman, B.; Bretagne, S.; Azoulay, E.; Mebazaa, A.; et al. Combination of Mycological Criteria: A Better Surrogate to Identify COVID-19-Associated Pulmonary Aspergillosis Patients and Evaluate Prognosis? J. Clin. Microbiol. 2022, 60, e0216921. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.; Alastruey-Izquierdo, A.; Berman, J.; Bicanic, T.; Bignell, E.; Bowyer, P.; Bromley, M.; Bruggemann, R.; Garber, G.; Cornely, O.; et al. Tackling the emerging threat of antifungal resistance to human health. Nat. Rev. Microbiol. 2022, 20, 557–571. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. First Meeting of the WHO Antifungal Expert Group on Identifying Priority Fungal Pathogens: Meeting Report; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Duong, T.; Le, T.; Tran, K.; Nguyen, P.; Nguyen, H.; Nguyen, T.; Nguyen, H.; Nguyen, B.; Fisher, M.; Rhodes, J.; et al. Azole-resistant Aspergillus fumigatus is highly prevalent in the environment of Vietnam, with marked variability by land use type. Environ. Microbiol. 2021, 23, 7632–7642. [Google Scholar] [CrossRef] [PubMed]
- Shelton, J.; Collins, R.; Uzzelll, C.; Alghamdi, A.; Dyer, P.; Singer, A.; Fisher, M. Citizen Science Surveillance of Triazole-Resistant Aspergillus fumigatus in United Kingdom Residential Garden Soils. Appl. Environ. Microbiol. 2022, 88, e02061-02021. [Google Scholar] [CrossRef]
- Fischer, J.; Koningsbruggen-Reitschel, S.v.; Rietschel, E.; Vehreschild, M.; Wisplinghoff, H.; Krönke, M.; Hamprecht, A. Prevalence and molecular characterization of azole resistance in Aspergillus spp. isolates from German cystic fibrosis patients. J. Antimicrob. Chemother. 2014, 69, 1533–1536. [Google Scholar] [CrossRef]
- Risum, M.; Hare, R.; Gertsen, J.; Kristensen, L.; Rosenvinge, F.; Sulim, S.; Abou-Chakra, N.; Bangsborg, J.; Røder, B.; Marmolin, E.; et al. Azole resistance in Aspergillus fumigatus. The first 2-year’s Data from the Danish National Surveillance Study, 2018–2020. Mycoses 2022, 65, 419–428. [Google Scholar] [CrossRef]
- Shelton, J. Environmental Drivers of Azole-Resistance in Aspergillus fumigatus and Its Relevance to Human Health; Imperial College London: London, UK, 2021. [Google Scholar]
- Tsuchido, Y.; Tanaka, M.; Nakano, S.; Yamamoto, N.; Matsumura, Y.; Nagao, M. Prospective multicenter surveillance of clinically isolated Aspergillus species revealed azole-resistant Aspergillus fumigatus isolates with TR34/L98H mutation in the Kyoto and Shiga regions of Japan. Med. Mycol. 2019, 57, 997–1003. [Google Scholar] [CrossRef]
- Mead, M.; Castro, P.d.; Steenwyk, J.; Gangneux, J.; Hoenigl, M.; Prattes, J.; Rautemaa-Richardson, R.; Guegan, H.; Moore, C.; Lass-Flörl, C.; et al. COVID-19 Associated Pulmonary Aspergillosis isolates are genomically diverse but similar to each other in their responses to infection-relevant stresses. Microbiol. Spectr. 2023, 11, e05128-22. [Google Scholar] [CrossRef]
- Rasilla, T.d.l.; González-Jiménez, I.; Fernández-Arroyo, A.; Roldán, A.; Carretero-Ares, J.; García-Clemente, M.; Telenti-Asensio, M.; García-Prieto, E.; Martínez-Suarez, M.; Vázquez-Valdés, F.; et al. COVID-19 Associated Pulmonary Aspergillosis (CAPA): Hospital or Home Environment as a Source of Life-Threatening Aspergillus fumigatus Infection? J. Fungi 2022, 8, 316. [Google Scholar] [CrossRef]
- Kirchoff, L.; Braun, L.; Schmidt, D.; Dittmer, S.; Dedy, J.; Herbstreit, F.; Buer, J.; Rath, P.; Steinmann, J.; Verhasselt, H. COVID-19-associated pulmonary aspergillosis in ICU patients in a German reference centre: Phenotypic and molecular characterisation of Aspergillus fumigatus isolates. Mycoses 2022, 65, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Steenwyk, J.; Mead, M.; Castro, P.d.; Valero, C.; Damasio, A.; Santos, R.d.; Labella, A.; Li, Y.; Knowles, S.; Raja, H.; et al. Genomic and Phenotypic Analysis of COVID-19-Associated Pulmonary Aspergillosis Isolates of Aspergillus Fumigatus. Microbiol. Spectr. 2021, 9, e0001021. [Google Scholar] [CrossRef] [PubMed]
- Morais, S.; Toscano, C.; Simões, H.; Carpinterio, D.; Viegas, C.; Veríssimo, C.; Sabino, R. Comparison of Multi-locus Genotypes Detected in Aspergillus fumigatus Isolated from COVID Associated Pulmonary Aspergillosis (CAPA) and from Other Clinical and Environmental Sources. J. Fungi 2023, 9, 298. [Google Scholar] [CrossRef] [PubMed]
- Rogers, T.; Verweij, P.; Castanheira, M.; Dannaoui, E.; White, P.; Arendrup, M. Molecular mechanisms of acquired antifungal drug resistance in principal fungal pathogens and EUCAST guidance for their laboratory detection and clinical implications. J. Antimicrob. Chemother. 2022, 77, 2053–2753. [Google Scholar] [CrossRef] [PubMed]
- Barber, A.; Sae-Ong, T.; Kang, K.; Seelbinder, B.; Li, J.; Walther, G.; Panagiotou, g.; Kurzai, O. Aspergillus fumigatus pan-genome analysis identifies genetic variants associated with human infection. Nat. Microbiol. 2021, 6, 1526–1536. [Google Scholar] [CrossRef]
- Reis, T.d.; Silva, L.; Castro, A.d.; Carmo, R.d.; Marini, M.; Silveria, J.d.; Ferreira, B.; Rodrigues, F.; Lind, A.; Rokas, A.; et al. The Aspergillus fumigatus Mismatch Repair MSH2 Homolog Is Important for Virulence and Azole Resistance. Mol. Sphere 2019, 4, e00416–e00419. [Google Scholar] [CrossRef]
- Furukawa, T.; Rhijn, N.v.; Fraczek, M.; Gsaller, F.; Davis, E.; Carr, P.; Gago, S.; Fortune-Grant, R.; Rahman, S.; Gilsenan, J.; et al. The negative cofactor 2 complex is a key regulator of drug resistance in Aspergillus fumigatus. Nat. Commun. 2020, 11, 427. [Google Scholar] [CrossRef]
- Novak-Frazer, L.; Anees-Hill, S.; Hassan, D.; Masania, R.; Moore, C.; Richardson, M.; Denning, D.; Rautemaa-Richardson, R. Deciphering Aspergillus fumigatus cyp51A-mediated triazole resistance by pyrosequencing of respiratory specimens. J. Antimicrob. Chemother. 2020, 75, 3501–3509. [Google Scholar] [CrossRef]
- Takazono, T.; Ito, Y.; Tashiro, M.; Nakano, Y.; Hirayama, T.; Hosogaya, N.; Saijo, T.; Yamamoto, K.; Imamura, Y.; Miyazaki, T.; et al. Transition of triazole-resistant Aspergillus fumigatus isolates in a Japanese tertiary hospital and subsequent genetic analysis. J. Infect. Chemother. 2021, 27, 537–539. [Google Scholar] [CrossRef]
- Rhodes, J.; Abdolrasouli, A.; Dunne, K.; Sewell, T.; Zhang, Y.; Ballard, E.; Brackin, A.; Rhijn, N.v.; Chown, N.; Tsitsopoulou, A.; et al. Population genomics confirms acquisition of drug-resistant Aspergillus fumigatus infection by humans from the environment. Nat. Microbiol. 2022, 7, 663–674. [Google Scholar] [CrossRef]
- Armstrong-James, D.; Youngs, J.; Bicanic, T.; Abdolrasouli, A.; Denning, D.; Johnson, E.; Mehra, V.; Pagliuca, T.; Patel, B.; Rhodes, J.; et al. Confronting and mitigating the risk of COVID-19 Associated Pulmonary Aspergillosis (CAPA). Eur. Respir. J. 2020, 56, 2002554. [Google Scholar] [CrossRef]
- Donnelly, J.; Chen, S.; Kauffman, C.; Steinbach, W.; Baddley, J.; Verweij, P.; Clancy, C.; Wingard, J.; Lockhart, S.; Groll, A.; et al. Revision and Update of the Consensus Definitions of Invasive Fungal Disease from the European Organization for Research and Treatment of Cancer and the Mycoses Study Group Education and Research Consortium. Clin. Infect. Dis. 2020, 71, 1367–1376. [Google Scholar] [CrossRef] [PubMed]
- Koehler, P.; Cornely, O.; Böttiger, B.; Dusse, F.; Eichenauer, D.; Fuchs, F.; Hallek, M.; Jung, N.; Klein, F.; Persigehl, T.; et al. COVID-19 associated pulmonary aspergillosis. Mycoses 2020, 63, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.; Hassan, T.; Trzos-Grzybowska, M.; Thomas, J.; Quinn, A.; O’Sullivan, M.; Griffin, A.; Rogers, T.; Talento, A. Multi-triazole-resistant Aspergillus fumigatus and SARS-CoV-2 co-infection: A lethal combination. Med. Mycol. Case Rep. 2021, 31, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Argimón, S.; Abudahab, K.; Goater, R.; Fedosejev, A.; Bhai, J.; Glasner, C.; Feil, E.; Holden, M.; Yeats, C.; Grundmann, H.; et al. Microreact: Visualizing and sharing data for genomic epidemiology and phylogeography. Microbiol. Genom. 2016, 2, e000093. [Google Scholar] [CrossRef]
- Armstrong-James, D. (Department of Infectious Diseases, Imperial College London, London SW7 2AZ, UK); Scourfiled, A. (Department of Infectious Diseases, Imperial College London, London SW7 2AZ, UK); Abdolrasouli, A. (Department of Infectious Diseases, Imperial College London, London SW7 2AZ, UK). Azole drug susceptibility of clinical A. fumigatus isolates grown in minimal media using EUCAST method (C307, C323, C360, C372, and C376), 2018. Unpublished work.
- Brackin, A.; Shelton, J.; Abdolrasouli, A.; Fisher, M.; Sewell, T. A low-cost tebuconazole-based screening test for azole-resistant aspergillus fumigatus. Curr. Protoc. Microbiol. 2020, 58, e112. [Google Scholar] [CrossRef]
- Buil, J.B.; Lee, H.A.L.v.d.; Rijs, A.J.M.M.; Zoll, J.; Hovestadt, J.A.M.F.; Melchers, W.J.G.; Verweij, P.E. Single-Center Evaluation of an Agar-Based Screening for Azole Resistance in Aspergillus fumigatus by Using VIPcheck. Antimicrob. Agents Chemother. 2017, 61, e01250-17. [Google Scholar] [CrossRef]
- Guinea, J. Updated EUCAST Clinical Breakpoints against Aspergillus, Implications for the Clinical Microbiology Laboratory. J. Fungi 2020, 6, 343. [Google Scholar] [CrossRef]
- Espinel-Ingroff, A.; Diekema, D.; Fothergill, A.; Johnson, E.; Pelaez, T.; Pfaller, M.; Rinaldi, M.; Canton, E.; Turnidge, J. Wild-Type MIC Distributions and Epidemiological Cutoff Values for the Triazoles and Six Aspergillus spp. for the CLSI Broth Microdilution Method (M38-A2 Document). J. Clin. Microbiol. 2010, 48, 3251–3257. [Google Scholar] [CrossRef]
- Korostin, D.; Kulemin, N.; Naumov, V.; Belova, V.; Kwon, D.; Gorbachev, A. Comparative analysis of novel MGISEQ-2000 sequencing platform vs. Illumina HiSeq 2500 for whole-genome sequencing. PLoS ONE 2020, 15, e0230301. [Google Scholar] [CrossRef]
- Nierman, W.; Pain, A.; Anderson, M.; Wortman, J.; Kim, H.; Arroyo, J.; Berriman, M.; Abe, K.; Archer, D.; Bermejo, C.; et al. Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus. Nature 2005, 438, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Heng, L.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Samson, R.; Visagie, C.; Houbraken, J.; Hong, S.; Hubka, V.; Klaasen, C.; Perrone, G.; Seifert, K.; Susca, A.; Tanney, J.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [PubMed]
- FungiDB. Afu4g10050 Calmodulin. 2015. Available online: https://fungidb.org/fungidb/app/record/gene/Afu4g10050 (accessed on 14 July 2022).
- Garnica, S.; Schön, M.; Abarenkov, K.; Riess, K.; Liimatainen, K.; Niskanen, T.; Dima, B.; Frøslev, T.; Jeppesen, T.; Peintner, U.; et al. Determining threshold values for barcoding fungi: Lessons from Cortinarius (Basidiomycota), a highly diverse and widespread ectomycorrhizal genus. FEMS Microbiol. Ecol. 2016, 92, fiw045. [Google Scholar] [CrossRef]
- Pyrzak, W.; Miller, K.; Miller, B. Mating Type Protein Mat1-2 from Asexual Aspergillus fumigatus Drives Sexual Reproduction in Fertile Aspergillus nidulans. Eukaryot. Cell 2008, 7, 1029–1040. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Jombart, T.; Collins, C. A tutorial for Discriminant Analysis of Principal Components (DAPC) Using Adegenet 2.0.0. Available online: https://adegenet.r-forge.r-project.org/files/tutorial-dapc.pdf (accessed on 14 July 2022).
- Garcia-Rubio, R.; Monzon, S.; Alcazar-Fuoli, L.; Cuesta, I.; Mellado, E. Genome-Wide Comparative Analysis of Aspergillus fumigatus Strains: The Reference Genome as a Matter of Concern. Genes 2018, 9, 363. [Google Scholar] [CrossRef]
- Paoletti, M.; Rydholm, C.; Schwier, E.; Anderson, M.; Szakacs, G.; Lutzoni, F.; Debeaupuis, J.; Latge, J.; Denning, D.; Dyer, P. Evidence for Sexuality in the Opportunistic Fungal Pathogen Aspergillus fumigatus. Curr. Biol. 2005, 15, 1242–1248. [Google Scholar] [CrossRef]
- Sewell, T.; Zhu, J.; Rhodes, J.; Hagen, G.; Meis, J.; Fisher, M.; Jombart, T. Nonrandom distribution of azole resistance across the global population of Aspergillus fumigatus. Mbio Am. Soc. Microbiol. 2019, 10, e00392–e00419. [Google Scholar] [CrossRef]
- Boyd, S.; Loh, K.; Lynch, J.; Alrashed, D.; Muzzammil, S.; Marsh, H.; Masoud, M.; Ihsan, S.; Matin-Loeches, I. Elevated Rates of Ventilator-Associated Pneumonia and COVID-19 Associated Pulmonary Aspergillosis in Critically Ill Patients with SARS-CoV2 Infection in the Second Wave: A Retrospective Chart Review. Antibiotics 2022, 11, 632. [Google Scholar] [CrossRef]
- Abdolrasouli, A.; Rhodes, J.; Beale, M.; Hagen, F.; Rogers, T.; Chowdhary, A.; Meis, J.; Armstrong-James, D.; Fisher, M. Genomic Context of Azole Resistance Mutations in Aspergillus fumigatus Determined Using Whole-Genome Sequencing. mBio 2015, 6, e00536-15. [Google Scholar] [CrossRef] [PubMed]
- Bruske, E.; Ottoe, T.; Frank, M. Whole genome sequencing and microsatellite analysis of the Plasmodium falciparum E5 NF54 strain show that the var, rifin and stevor gene families follow Mendelian inheritance. Malar. J. 2018, 17, 376. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rubio, R.; Escribano, P.; Gomez, A.; Guinea, J.; Mellado, E. Comparison of Two Highly Discriminatory Typing Methods to Analyze Aspergillus fumigatus Azole Resistance. Front. Microbiol. 2018, 9, 1626. [Google Scholar] [CrossRef] [PubMed]
- Meijer, E.; Dofferhoff, A.; Hoiting, O.; Buil, J.; Meis, J. Azole-Resistant COVID-19-Associated Pulmonary Aspergillosis in an Immunocompetent Host: A Case Report. J. Fungi 2020, 6, 79. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Jimenez, I.; Lucio, J.; Menéndez-Fraga, M.D.; Mellado, E.; Peláez, T. Hospital Environment as a Source of Azole-Resistant Aspergillus fumigatus Strains with TR34/L98H and G448S Cyp51A Mutations. J. Fungi 2021, 7, 22. [Google Scholar] [CrossRef]
- Escribano, P.; Rodríguez-Sánchez, B.; Díaz-García, J.; Martín-Gómez, M.; Ibáñez-Martínez, E.; Rodríguez-Mayo, M.; Pelâez, T.; Pedrosa, E.G.-G.d.l.; Tejero-García, R.; Marimón, J.; et al. Azole resistance survey on clinical Aspergillus fumigatus isolates in Spain. Clin. Microbiol. Infect. 2021, 27, 1170.E1171–1170.E1177. [Google Scholar] [CrossRef]
- Grootveld, R.v.; Paasen, J.v.; Boer, M.d.; Claas, E.; Kuijper, E.; Beek, M.v.d.; Group, L.-C.-R. Systematic screening for COVID-19-associated invasive aspergillosis in ICU patients by culture and PCR on tracheal aspirate. Myoces 2021, 64, 641–650. [Google Scholar] [CrossRef]
- Abdolrasouli, A.; Petrou, M.; Park, H.; Rhodes, J.; Rawson, T.; Moore, L.; Donaldson, H.; Holmes, A.; Fisher, M.; Armstrong-James, D. Surveillance for Azole-Resistant Aspergillus fumigatus in a Centralized Diagnostic Mycology Service, London, United Kingdom, 1998–2017. Front. Microbiol. 2018, 9, 2234. [Google Scholar] [CrossRef]
- Lestrade, P.; Bentvelsen, R.; Schauwvlieghe, A.; Schalekamp, S.; Velden, W.v.d.; Kuiper, E.; Paassen, J.v.; Hoven, B.v.d.; Lee, H.v.d.; Melchers, W.; et al. Voriconazole Resistance and Mortality in Invasive Aspergillosis: A Multicenter Retrospective Cohort Study. Clin. Infect. Dis. 2019, 68, 1463–1471. [Google Scholar] [CrossRef]
- Linden, J.v.d.; Snelders, E.; Kampinga, G.A.; Rijnders, B.; Mattsson, E.; Debets-Ossenkopp, Y.; Kuijper, E.; Tiel, F.; Melchers, W.; Verweij, P. Clinical Implications of Azole Resistance in Aspergillus fumigatus, the Netherlands, 2007–2009. Emerg. Infect. Dis. 2011, 17, 1846–1854. [Google Scholar] [CrossRef]
- Burks, C.; Darby, A.; Londoño, L.; Momany, M.; Brewer, M. Azole-resistant Aspergillus fumigatus in the environment: Identifying key reservoirs and hotspots of antifungal resistance. PLoS Pathog. 2021, 17, e1009711. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, B.; Illescas, L.; Posadas, A.; Campos, P. Azole resistance among clinical isolates of Aspergillus fumigatus in Lima-Peru. Med. Mycol. 2020, 58, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Sewell, T. Elevated Prevalence of Azole-Resistant Aspergillus fumigatus in Urban versus Rural Environments in the United Kingdom. Antimicrob. Agents Chemother. 2019, 63, e00548-19. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Sharma, C.; Boom, M.v.d.; Yntema, J.; Hagen, F.; Verweij, P.; Meis, J. Multi-azole-resistant Aspergillus fumigatus in the environment in Tanzania. J. Antimicrob. Chemother. 2014, 69, 2979–2983. [Google Scholar] [CrossRef]
- Chowdhary, A.; Kathuria, S.; Randhawa, H.; Gaur, S.; Klaassen, C.; Meis, J. Isolation of multiple-triazole-resistant Aspergillus fumigatus strains carrying the TR34/L98H mutations in the cyp51A gene in India. J. Antimicrob. Chemother. 2012, 67, 362–366. [Google Scholar] [CrossRef]
- Chang, H.; Ashu, E.; Sharma, C.; Kathuria, S.; Chowdhary, A.; Xu, J. Diversity and origins of Indian multi-triazole resistant strains of Aspergillus fumigatus. Mycoses 2016, 59, 450–466. [Google Scholar] [CrossRef]
- Talbot, J.; Subedi, S.; Halliday, C.; Hibbs, D.; Lopez-Ruiz, F.; Harper, L.; Park, R.; Cuddy, W.; Biswas, C.; Cooley, L.; et al. Surveillance for azole resistance in clinical and environmental isolates of Aspergillus fumigatus in Australia and cyp51A homology modelling of azole-resistant isolates. J. Antimicrob. Chemother. 2018, 73, 2347–2351. [Google Scholar] [CrossRef]
- Wu, C.; Wang, H.; Lee, J.; Lo, H.; Dai, C.; Chou, P.; Ko, W.; Chen, Y. Azole-resistant Aspergillus fumigatus isolates carrying TR34/L98H mutations in Taiwan. Mycoses 2015, 58, 544–549. [Google Scholar] [CrossRef]
- Vermeulen, E.; Maertens, J.; Schoemans, H.; Lagrou, K. Azole-resistant Aspergillus fumigatus due to TR46/Y121F/T289A mutation emerging in Belgium, July 2012. Euro Surveill. 2012, 17, 20326. [Google Scholar] [CrossRef]
- Buil, J.; Hare, R.; Zwaan, B.; Arendrup, M.; Melchers, W.; Verweij, P. The fading boundaries between patient and environmental routes of triazole resistance selection in Aspergillus fumigatus. PLoS Pathog. 2019, 15, e1007858. [Google Scholar] [CrossRef]
- Majima, H.; Arai, T.; Kusuya, Y.; Takahashi, H.; Watanabe, A.; Miyazaki, Y.; Kamei, K. Genetic differences between Japan and other countries in cyp51A polymorphisms of Aspergillus fumigatus. Mycoses 2021, 64, 1354–1365. [Google Scholar] [CrossRef] [PubMed]
- Zea, S.; Toyotome, T. Azole-resistant Aspergillus fumigatus as an emerging worldwide pathogen. Microbiol. Immunol. 2021, 66, 135–144. [Google Scholar] [CrossRef]
- Albarrg, A.; Anderson, M.; Howard, S.; Robson, G.; Warn, P.; Sanglard, D.; Denning, D. Interrogation of Related Clinical Pan-Azole-Resistant Aspergillus fumigatus Strains: G138C, Y431C, and G434C Single Nucleotide Polymorphisms in cyp51A, Upregulation of cyp51A, and Integration and Activation of Transposon Atf1 in the cyp51A Promoter. Antimicrob. Agents Chemother. 2011, 55, 5113–5121. [Google Scholar] [CrossRef]
- Howard, S.; Pasqualotto, A.; Anderson, M.; Leatherbarrow, H.; Albarrag, A.; Harrison, E.; Gregson, L.; Bowyer, P.; Denning, D. Major variations in Aspergillus fumigatus arising within aspergillomas in chronic pulmonary aspergillosis. Mycoses 2012, 56, 434–441. [Google Scholar] [CrossRef]
- Ballard, E.; Melchers, W.; Zoll, J.; Brown, A.; Verweij, P.; Warris, A. In-host microevolution of Aspergillus fumigatus: A phenotypic and genotypic analysis. Fungal Genet. Biol. 2018, 113, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Suzuki, J.; Watanabe, A.; Narumoto, O.; Kawashima, M.; Fukami, T.; Tamura, A.; Nagai, H.; Matsui, H.; Kamei, K. High detection rate of azole-resistant Aspergillus fumigatus after treatment with azole antifungal drugs among patients with chronic pulmonary aspergillosis in a single hospital setting with low azole resistance. Med. Mycol. 2020, 59, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Shalhoub, S.; Luong, M.; Howard, S.; Richardson, S.; Singer, L.; Chaparro, C.; Keshavjee, S.; Akinlolu, Y.; Rotstein, C.; Mazzulli, T.; et al. Rate of cyp51A mutation in Aspergillus fumigatus among lung transplant recipients with targeted prophylaxis. J. Antimicrob. Chemother. 2015, 70, 1064–1067. [Google Scholar] [CrossRef]
- Camps, S.; Dutilh, B.; Arendrup, M.; Rijs, A.; Snelders, E.; Huynen, M.; Verweij, P.; Melchers, W. Discovery of a hapE Mutation That Causes Azole Resistance in Aspergillus fumigatus through Whole Genome Sequencing and Sexual Crossing. PLoS ONE 2012, 7, e50034. [Google Scholar] [CrossRef]
- Ballard, E.; Weber, J.; Melchers, W.; Tammireddy, S.; Whitfield, P.; Brakhage, A.; Brown, A.; Verweij, P.; Warris, A. Recreation of in-host acquired single nucleotide polymorphisms by CRISPR-Cas9 reveals an uncharacterised gene playing a role in Aspergillus fumigatus azole resistance via a non-cyp51A mediated resistance mechanism. Fungal Genet. Biol. 2019, 130, 98–106. [Google Scholar] [CrossRef]
- Rybak, J. Challenging the Paradigm of Clinical Triazole Resistance in Aspergillus fumigatus; The University of Tennessee Health Science Center: Memphis, TN, USA, 2019. [Google Scholar] [CrossRef]






| Isolate ID | Country | Sample Type | No. of Aligned Reads (Millions) | Mean Depth of Coverage (x) | Percentage of Reference Genome Covered (%) |
|---|---|---|---|---|---|
| CAPA-A | Germany | CAPA—Possible | 35.8 | 146.7 | 98.4 |
| CAPA-B | Germany | CAPA—Probable | 40.1 | 166.7 | 98.5 |
| CAPA-C | Germany | CAPA—Probable | 37.5 | 157.4 | 98.1 |
| CAPA-D | Germany | CAPA—Possible | 34.2 | 147.5 | 97.3 |
| C403 | Netherlands | CAPA—Possible | 37.2 | 165.0 | 98.6 |
| C408 | Netherlands | CAPA—Possible | 28.5 | 127.7 | 98.3 |
| C422 | UK | CAPA—Probable | 11.8 | 45.5 | 96.2 |
| C423 | UK | CAPA—Probable | 15.6 | 56.2 | 96.5 |
| C424 | UK | CAPA—Probable | 11.2 | 45.1 | 98.6 |
| C425 | UK | CAPA—Probable | 11.0 | 43.0 | 97.2 |
| C434 | Ireland | CAPA—Probable | 10.6 | 41.9 | 96.6 |
| C435 | Ireland | CAPA—Probable | 10.3 | 41.4 | 97.7 |
| C436 | Ireland | CAPA—Probable | 9.8 | 39.3 | 98.2 |
| C437 | Ireland | CAPA—Probable | 9.6 | 38.9 | 97.4 |
| C438 | Ireland | CAPA—Probable | 10.5 | 42.6 | 97.7 |
| C439 | Ireland | CAPA—Probable | 11.4 | 45.1 | 98.3 |
| C440 | Ireland | CAPA—Probable | 7.8 | 27.6 | 97.9 |
| C441 | Ireland | CAPA—Probable | 9.2 | 36.3 | 97.9 |
| C443 | Ireland | CAPA—Probable | 9.7 | 39.2 | 97.5 |
| C444 | Ireland | CAPA—Probable | 11.0 | 43.2 | 97.9 |
| C611 | UK | CAPA—Probable | 11.0 | 43.7 | 97.6 |
| C612 | UK | CAPA—Possible | 11.0 | 44.0 | 97.5 |
| Isolate ID | VIPcheckTM Score | Tebucheck Score | Resistance Marker | ||
|---|---|---|---|---|---|
| ITR | VOR | POS | |||
| C154 | 0 | 0 | 0 | 1 | WT |
| C403 * | ND | ND | ND | ND | WT |
| C408 * | ND | ND | ND | ND | WT |
| C422 | ND | ND | ND | 1 | WT |
| C423 | ND | ND | ND | 1 | WT |
| C424 | ND | ND | ND | 1 | WT |
| C425 | ND | ND | ND | 1 | WT |
| C434 | 0 | 0 | 0 | 1 | WT |
| C435 | ND | ND | ND | 1 | WT |
| C436 | 0 | 0 | 0 | 1 | WT |
| C437 | 0 | 0 | 0 | 1 | WT |
| C438 | 1 | 1 | 1 | 3 | TR34/L98H |
| C439 | ND | ND | ND | 1 | WT |
| C440 | 0 | 0 | 0 | 1 | WT |
| C441 | 1 | 1 | 0 | 4 | TR34/L98H |
| C444 | ND | ND | ND | 3 | TR34/L98H |
| C611 | 0 | 0 | 0 | 1 | WT |
| C612 | ND | ND | ND | 1 | WT |
| CAPA-A * | ND | ND | ND | ND | WT |
| CAPA-B * | ND | ND | ND | ND | WT |
| CAPA-C * | ND | ND | ND | ND | WT |
| CAPA-D * | ND | ND | ND | ND | WT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simmons, B.C.; Rhodes, J.; Rogers, T.R.; Verweij, P.E.; Abdolrasouli, A.; Schelenz, S.; Hemmings, S.J.; Talento, A.F.; Griffin, A.; Mansfield, M.; et al. Genomic Epidemiology Identifies Azole Resistance Due to TR34/L98H in European Aspergillus fumigatus Causing COVID-19-Associated Pulmonary Aspergillosis. J. Fungi 2023, 9, 1104. https://doi.org/10.3390/jof9111104
Simmons BC, Rhodes J, Rogers TR, Verweij PE, Abdolrasouli A, Schelenz S, Hemmings SJ, Talento AF, Griffin A, Mansfield M, et al. Genomic Epidemiology Identifies Azole Resistance Due to TR34/L98H in European Aspergillus fumigatus Causing COVID-19-Associated Pulmonary Aspergillosis. Journal of Fungi. 2023; 9(11):1104. https://doi.org/10.3390/jof9111104
Chicago/Turabian StyleSimmons, Benjamin C., Johanna Rhodes, Thomas R. Rogers, Paul E. Verweij, Alireza Abdolrasouli, Silke Schelenz, Samuel J. Hemmings, Alida Fe Talento, Auveen Griffin, Mary Mansfield, and et al. 2023. "Genomic Epidemiology Identifies Azole Resistance Due to TR34/L98H in European Aspergillus fumigatus Causing COVID-19-Associated Pulmonary Aspergillosis" Journal of Fungi 9, no. 11: 1104. https://doi.org/10.3390/jof9111104