

Pseudocercospora fijiensis Conidial Germination Is Dominated by Pathogenicity Factors and Effectors
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material and Preparation
2.2. RNA Isolation and Transcriptome Sequencing
2.3. Sequence Cleaning and Transcriptome Assembly
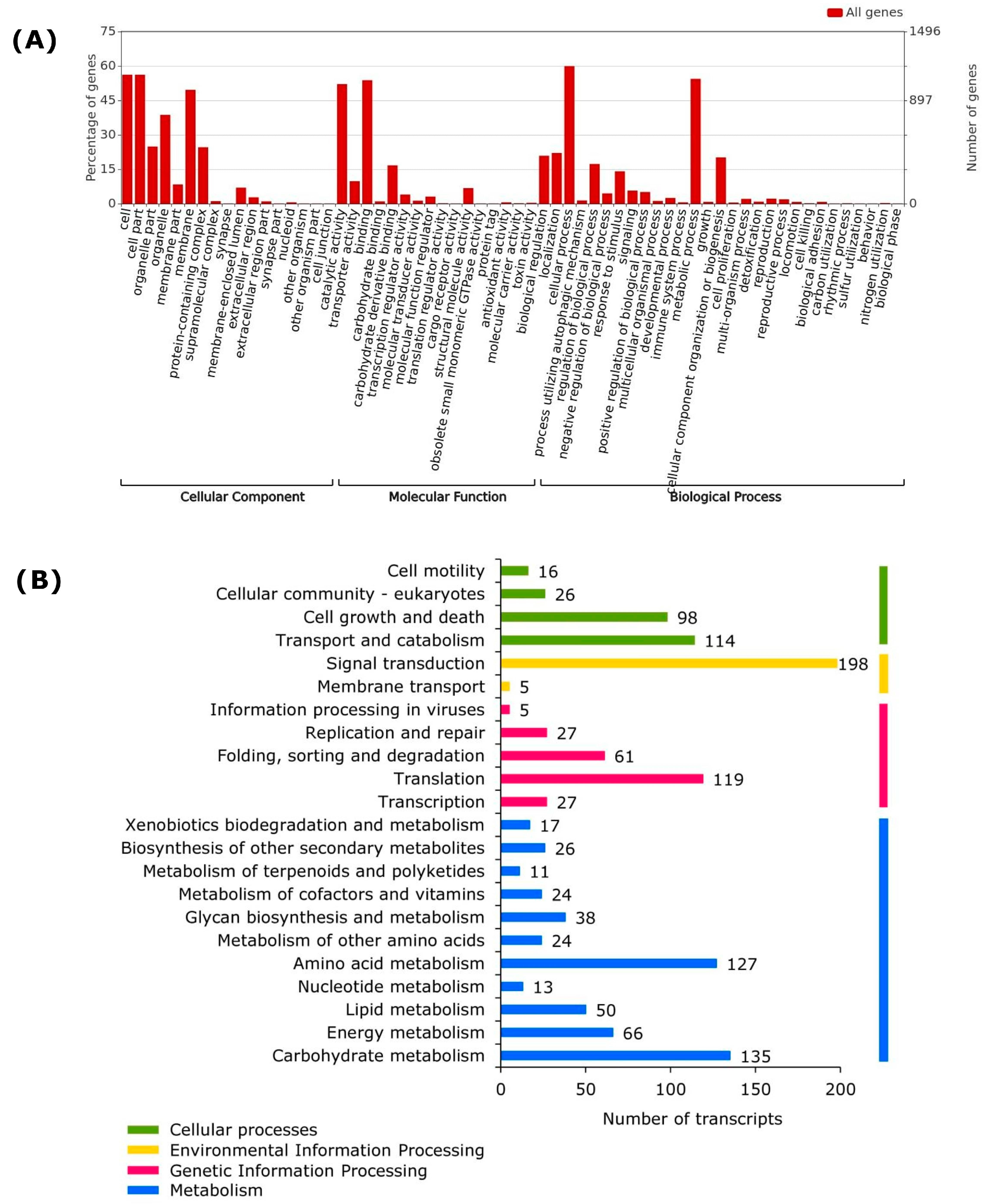
2.4. Gene Ontology Distribution and Functional Annotation
2.5. Identification of Transcripts Involved in Pathogen–Host Interaction
2.6. Prediction of Effector Proteins
2.7. Differential Expression of Genes (DEGs) of Top Conidial Transcripts
3. Results
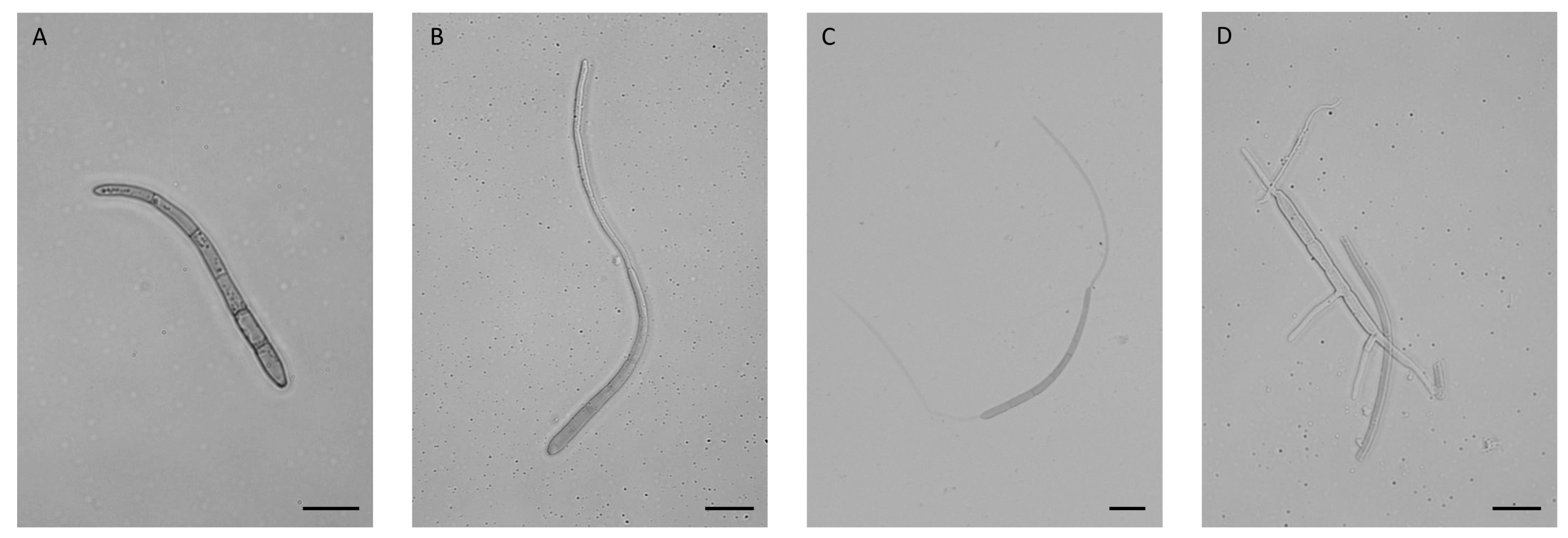
3.1. Conidia Germination
3.2. Sequence Quality Analysis, Assembly, and Functional Annotation
3.3. Transcriptional and Translational Activities
3.4. Most Active Pathways
3.5. Secretome
3.6. Other Important Processes Usually Found in Conidia
3.7. Pathogenicity-Associated Functions
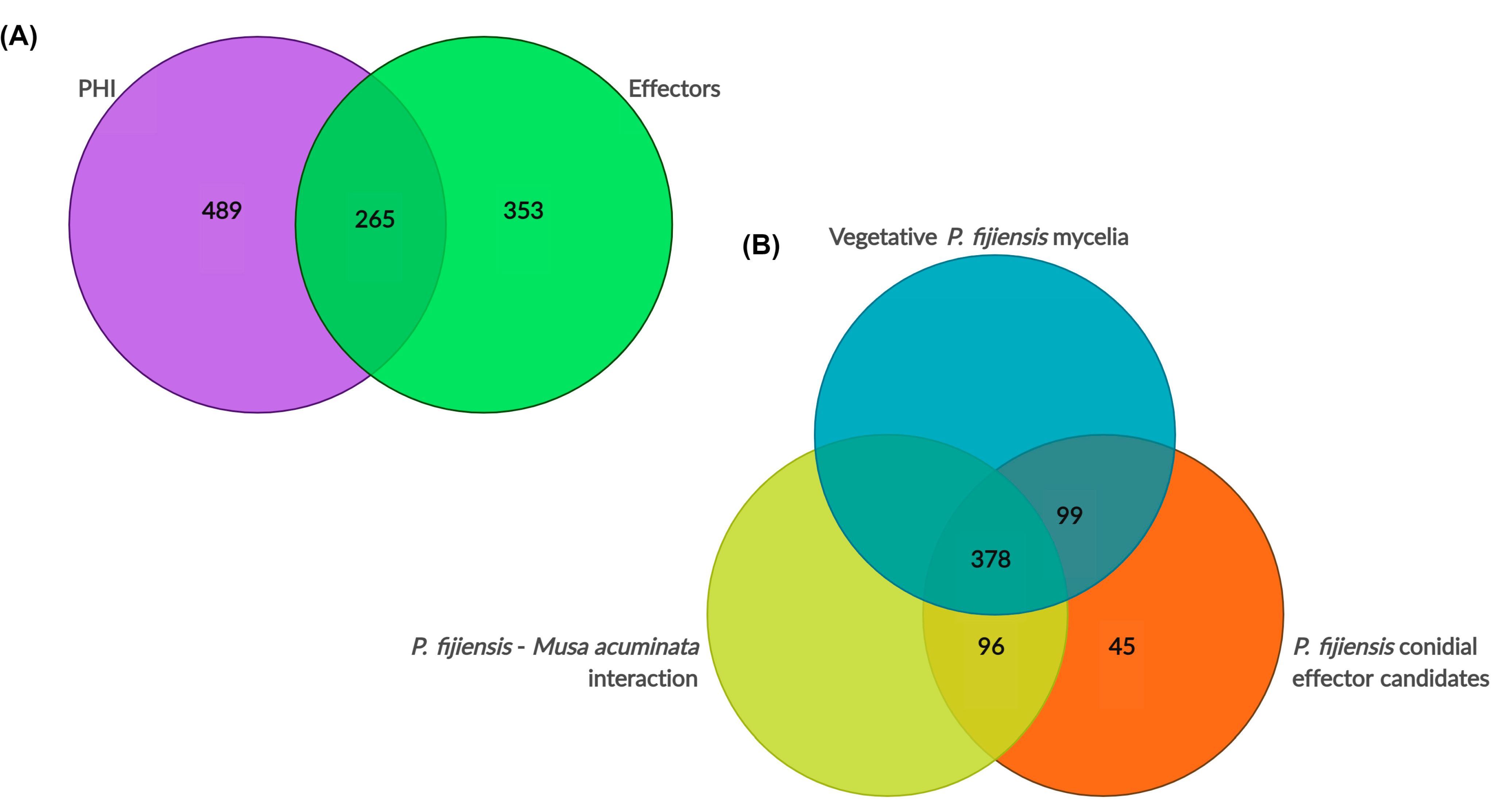
3.7.1. PHI Homologs
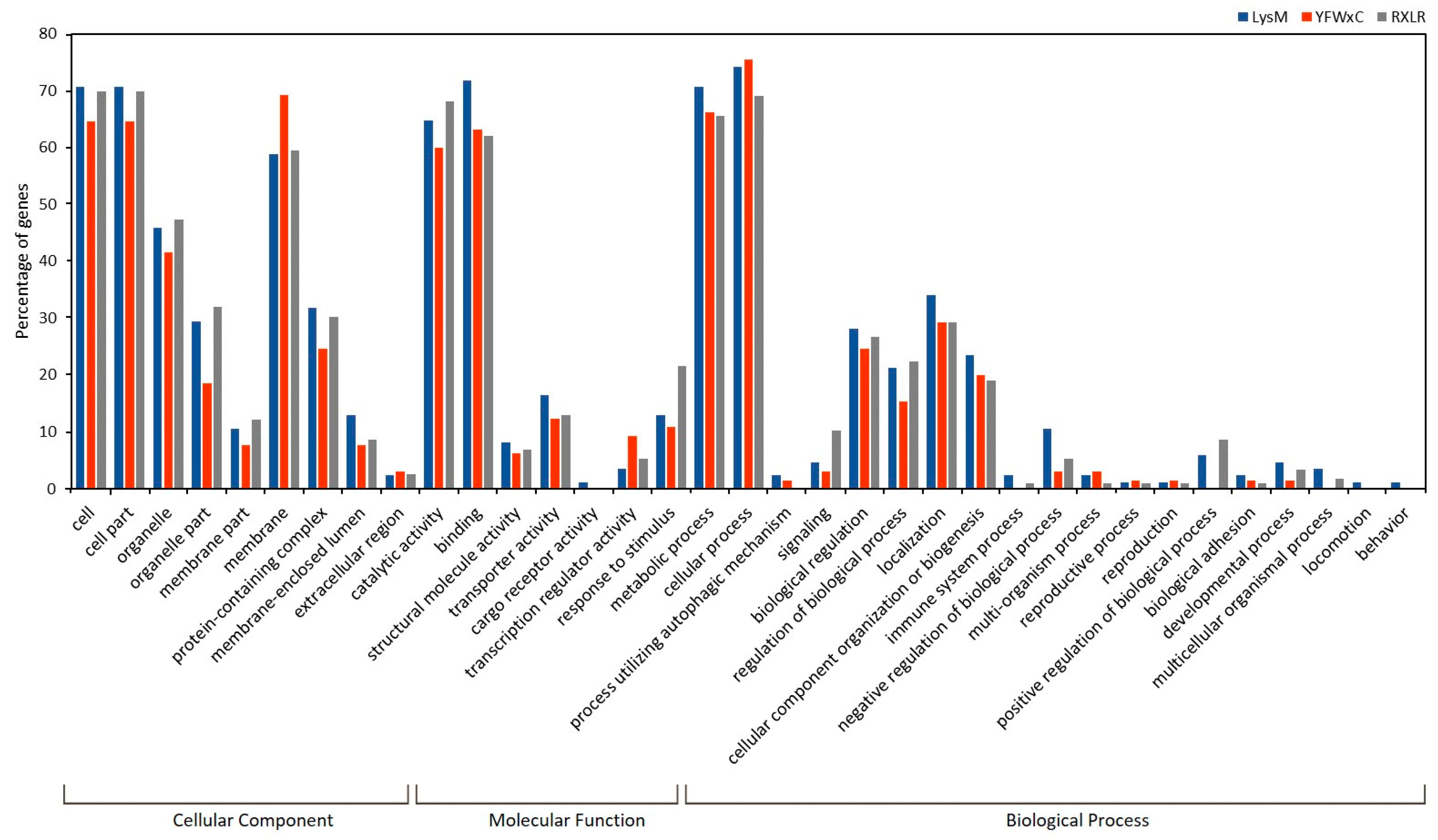
3.7.2. Effectorome Prediction
4. Discussion
4.1. High Transcriptional Expression Levels
4.2. Storage and Energy Metabolisms
4.3. Defense and Oxidative Stress
4.4. Pathogenicity
4.5. Conidial Processes Regulation by Gene Silencing
4.6. Transcriptome Representation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olmedo, M.; Ruger-Herreros, C.; Luque, E.M.; Corrochano, L.M. A Complex Photoreceptor System Mediates the Regulation by Light of the Conidiation Genes Con-10 and Con-6 in Neurospora crassa. Fungal Genet. Biol. 2010, 47, 352–363. [Google Scholar] [CrossRef]
- Lee, S.-J.; Kong, M.; Morse, D.; Hijri, M. Expression of Putative Circadian Clock Components in the Arbuscular Mycorrhizal Fungus Rhizoglomus irregulare. Mycorrhiza 2018, 28, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Kikuma, T.; Arioka, M.; Kitamoto, K. Autophagy during Conidiation and Conidial Germination in Filamentous Fungi. Autophagy 2007, 3, 128–129. [Google Scholar] [CrossRef] [PubMed]
- Blango, M.G.; Kniemeyer, O.; Brakhage, A.A. Conidial Surface Proteins at the Interface of Fungal Infections. PLoS Pathog. 2019, 15, e1007939. [Google Scholar] [CrossRef] [PubMed]
- Grünbacher, A.; Throm, T.; Seidel, C.; Gutt, B.; Röhrig, J.; Strunk, T.; Vincze, P.; Walheim, S.; Schimmel, T.; Wenzel, W.; et al. Six Hydrophobins Are Involved in Hydrophobin Rodlet Formation in Aspergillus nidulans and Contribute to Hydrophobicity of the Spore Surface. PLoS ONE 2014, 9, e94546. [Google Scholar] [CrossRef]
- Qu, J.; Zou, X.; Yu, J.; Zhou, Y. The Conidial Mucilage, Natural Film Coatings, Is Involved in Environmental Adaptability and Pathogenicity of Hirsutella satumaensis Aoki. Sci. Rep. 2017, 7, 1301. [Google Scholar] [CrossRef]
- Santi, L.; Beys Da Silva, W.O.; Berger, M.; Guimarães, J.A.; Schrank, A.; Vainstein, M.H. Conidial Surface Proteins of Metarhizium anisopliae: Source of Activities Related with Toxic Effects, Host Penetration and Pathogenesis. Toxicon 2010, 55, 874–880. [Google Scholar] [CrossRef]
- Teutschbein, J.; Albrecht, D.; Pötsch, M.; Guthke, R.; Aimanianda, V.; Clavaud, C.; Latgé, J.-P.; Brakhage, A.A.; Kniemeyer, O. Proteome Profiling and Functional Classification of Intracellular Proteins from Conidia of the Human-Pathogenic Mold Aspergillus Fumigatus. J. Proteome Res. 2010, 9, 3427–3442. [Google Scholar] [CrossRef]
- Kim, S.; Kim, C.-Y.; Park, S.-Y.; Kim, K.-T.; Jeon, J.; Chung, H.; Choi, G.; Kwon, S.; Choi, J.; Jeon, J.; et al. Two Nuclear Effectors of the Rice Blast Fungus Modulate Host Immunity via Transcriptional Reprogramming. Nat. Commun. 2020, 11, 5845. [Google Scholar] [CrossRef]
- Lazar, N.; Mesarich, C.H.; Petit-Houdenot, Y.; Talbi, N.; Li De La Sierra-Gallay, I.; Zélie, E.; Blondeau, K.; Gracy, J.; Ollivier, B.; Blaise, F.; et al. A New Family of Structurally Conserved Fungal Effectors Displays Epistatic Interactions with Plant Resistance Proteins. PLoS Pathog. 2022, 18, e1010664. [Google Scholar] [CrossRef]
- Rocafort, M.; Bowen, J.K.; Hassing, B.; Cox, M.P.; McGreal, B.; De La Rosa, S.; Plummer, K.M.; Bradshaw, R.E.; Mesarich, C.H. The Venturia inaequalis Effector Repertoire Is Dominated by Expanded Families with Predicted Structural Similarity, but Unrelated Sequence, to Avirulence Proteins from Other Plant-Pathogenic Fungi. BMC Biol. 2022, 20, 246. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, A.; Ghosh, S.; Sahoo, D.; Jha, G. Fungal Effectors, the Double Edge Sword of Phytopathogens. Curr. Genet. 2021, 67, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, C.; Si, J.; Han, Z.; Chen, D. Action Mechanisms of Effectors in Plant-Pathogen Interaction. Int. J. Mol. Sci. 2022, 23, 6758. [Google Scholar] [CrossRef] [PubMed]
- Ryder, L.S.; Talbot, N.J. Regulation of Appressorium Development in Pathogenic Fungi. Curr. Opin. Plant Biol. 2015, 26, 8–13. [Google Scholar] [CrossRef]
- Ryder, L.S.; Cruz-Mireles, N.; Molinari, C.; Eisermann, I.; Eseola, A.B.; Talbot, N.J. The Appressorium at a Glance. J. Cell Sci. 2022, 135, jcs259857. [Google Scholar] [CrossRef]
- Yan, X.; Tang, B.; Ryder, L.S.; MacLean, D.; Were, V.M.; Eseola, A.B.; Cruz-Mireles, N.; Ma, W.; Foster, A.J.; Osés-Ruiz, M.; et al. The Transcriptional Landscape of Plant Infection by the Rice Blast Fungus Magnaporthe oryzae Reveals Distinct Families of Temporally Co-Regulated and Structurally Conserved Effectors. Plant Cell 2023, 35, 1360–1385. [Google Scholar] [CrossRef]
- Dodds, P.N.; Rafiqi, M.; Gan, P.H.P.; Hardham, A.R.; Jones, D.A.; Ellis, J.G. Effectors of Biotrophic Fungi and Oomycetes: Pathogenicity Factors and Triggers of Host Resistance. New Phytol. 2009, 183, 993–1000. [Google Scholar] [CrossRef]
- Chaudhari, P.; Ahmed, B.; Joly, D.L.; Germain, H. Effector Biology during Biotrophic Invasion of Plant Cells. Virulence 2014, 5, 703–709. [Google Scholar] [CrossRef]
- Figueroa, M.; Ortiz, D.; Henningsen, E.C. Tactics of Host Manipulation by Intracellular Effectors from Plant Pathogenic Fungi. Curr. Opin. Plant Biol. 2021, 62, 102054. [Google Scholar] [CrossRef]
- Chen, H.; He, S.; Zhang, S.; Runa, A.; Li, W.; Liu, S. The Necrotroph Botrytis cinerea BcSpd1 Plays a Key Role in Modulating Both Fungal Pathogenic Factors and Plant Disease Development. Front. Plant Sci. 2022, 13, 820767. [Google Scholar] [CrossRef]
- Kanyuka, K.; Igna, A.A.; Solomon, P.S.; Oliver, R.P. The Rise of Necrotrophic Effectors. New Phytol. 2022, 233, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Shao, D.; Smith, D.L.; Kabbage, M.; Roth, M.G. Effectors of Plant Necrotrophic Fungi. Front. Plant Sci. 2021, 12, 687713. [Google Scholar] [CrossRef] [PubMed]
- Vilanova, L.; Valero-Jiménez, C.A.; Van Kan, J.A.L. Deciphering the Monilinia fructicola Genome to Discover Effector Genes Possibly Involved in Virulence. Genes 2021, 12, 568. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jiang, N.; Liu, J.; Liu, W.; Wang, G.-L. The Role of Effectors and Host Immunity in Plant–Necrotrophic Fungal Interactions. Virulence 2014, 5, 722–732. [Google Scholar] [CrossRef]
- Luo, X.; Mao, H.; Wei, Y.; Cai, J.; Xie, C.; Sui, A.; Yang, X.; Dong, J. The Fungal-Specific Transcription Factor Vdpf Influences Conidia Production, Melanized Microsclerotia Formation and Pathogenicity in Verticillium dahliae: The Function of Vdpf in Verticillium Dahliae. Mol. Plant Pathol. 2016, 17, 1364–1381. [Google Scholar] [CrossRef]
- Hou, X.; An, B.; Wang, Q.; Guo, Y.; Luo, H.; He, C. SGE1 Is Involved in Conidiation and Pathogenicity of Fusarium Oxysporum f. Sp. Cubense. Can. J. Microbiol. 2018, 64, 349–357. [Google Scholar] [CrossRef]
- Gurdaswani, V.; Ghag, S.B.; Ganapathi, T.R. FocSge1 in Fusarium Oxysporum f. Sp. Cubense Race 1 Is Essential for Full Virulence. BMC Microbiol. 2020, 20, 255. [Google Scholar] [CrossRef]
- Ayukawa, Y.; Asai, S.; Gan, P.; Tsushima, A.; Ichihashi, Y.; Shibata, A.; Komatsu, K.; Houterman, P.M.; Rep, M.; Shirasu, K.; et al. A Pair of Effectors Encoded on a Conditionally Dispensable Chromosome of Fusarium oxysporum Suppress Host-Specific Immunity. Commun. Biol. 2021, 4, 707. [Google Scholar] [CrossRef]
- Mehta, N.; Patil, R.; Baghela, A. Differential Physiological Prerequisites and Gene Expression Profiles of Conidial Anastomosis Tube and Germ Tube Formation in Colletotrichum gloeosporioides. J. Fungi 2021, 7, 509. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, J.; Qi, Y.; Li, J.; Amin, R.; Yang, W.; Liu, D. Predicating the Effector Proteins Secreted by Puccinia triticina through Transcriptomic Analysis and Multiple Prediction Approaches. Front. Microbiol. 2020, 11, 538032. [Google Scholar] [CrossRef]
- Sánchez-Vallet, A.; Tian, H.; Rodriguez-Moreno, L.; Valkenburg, D.-J.; Saleem-Batcha, R.; Wawra, S.; Kombrink, A.; Verhage, L.; De Jonge, R.; Van Esse, H.P.; et al. A Secreted LysM Effector Protects Fungal Hyphae through Chitin-Dependent Homodimer Polymerization. PLoS Pathog. 2020, 16, e1008652. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; MacKenzie, C.I.; Rodriguez-Moreno, L.; Berg, G.C.M.; Chen, H.; Rudd, J.J.; Mesters, J.R.; Thomma, B.P.H.J. Three LysM Effectors of Zymoseptoria Tritici Collectively Disarm Chitin-triggered Plant Immunity. Mol. Plant Pathol. 2021, 22, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Mapuranga, J.; Zhang, N.; Zhang, L.; Chang, J.; Yang, W. Infection Strategies and Pathogenicity of Biotrophic Plant Fungal Pathogens. Front. Microbiol. 2022, 13, 799396. [Google Scholar] [CrossRef] [PubMed]
- Leiva-Mora, M.; Alvarado-Capó, Y.; Acosta-Suárez, M.; Cruz-Martín, M.; Roque-Morales, B. Enhanced Sporulation, Morphological and Pathogenic Characterization of Mycosphaerella fijiensis, Causal Agent of Musa Black Leaf Streak. Cent. Agric. 2008, 35, 33–39. [Google Scholar]
- Gerke, J.; Schneider, D.; Poehlein, A.; Cornish, V.W.; Braus, G.H. Draft Genome Sequence of Saccharomyces cerevisiae LW2591Y, a Laboratory Strain for In Vivo Multigene Assemblies. Microbiol. Resour. Announc. 2021, 10, e01418-20. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-Length Transcriptome Assembly from RNA-Seq Data without a Reference Genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for Clustering the next-Generation Sequencing Data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Carreón-Anguiano, K.G.; Islas-Flores, I.; Vega-Arreguín, J.; Sáenz-Carbonell, L.; Canto-Canché, B. EffHunter: A Tool for Prediction of Effector Protein Candidates in Fungal Proteomic Databases. Biomolecules 2020, 10, 712. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating Signal Peptides from Transmembrane Regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L.L. Predicting Transmembrane Protein Topology with a Hidden Markov Model: Application to Complete genomes11Edited by F. Cohen. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef] [PubMed]
- The Gene Ontology Consortium; Carbon, S.; Douglass, E.; Good, B.M.; Unni, D.R.; Harris, N.L.; Mungall, C.J.; Basu, S.; Chisholm, R.L.; Dodson, R.J.; et al. The Gene Ontology Resource: Enriching a GOld Mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Ye, J.; Fang, L.; Zheng, H.; Zhang, Y.; Chen, J.; Zhang, Z.; Wang, J.; Li, S.; Li, R.; Bolund, L.; et al. WEGO: A Web Tool for Plotting GO Annotations. Nucleic Acids Res. 2006, 34, W293–W297. [Google Scholar] [CrossRef]
- Aramaki, T.; Blanc-Mathieu, R.; Endo, H.; Ohkubo, K.; Kanehisa, M.; Goto, S.; Ogata, H. KofamKOALA: KEGG Ortholog Assignment Based on Profile HMM and Adaptive Score Threshold. Bioinformatics 2020, 36, 2251–2252. [Google Scholar] [CrossRef]
- Urban, M.; Cuzick, A.; Seager, J.; Wood, V.; Rutherford, K.; Venkatesh, S.Y.; Sahu, J.; Iyer, S.V.; Khamari, L.; De Silva, N.; et al. PHI-Base in 2022: A Multi-Species Phenotype Database for Pathogen–Host Interactions. Nucleic Acids Res. 2022, 50, D837–D847. [Google Scholar] [CrossRef]
- Mount, D.W. Using the Basic Local Alignment Search Tool (BLAST). Cold Spring Harb. Protoc. 2007, 2007, pdb-top17. [Google Scholar] [CrossRef]
- Carreón-Anguiano, K.G.; Todd, J.N.A.; Chi-Manzanero, B.H.; Couoh-Dzul, O.J.; Islas-Flores, I.; Canto-Canché, B. WideEffHunter: An Algorithm to Predict Canonical and Non-Canonical Effectors in Fungi and Oomycetes. Int. J. Mol. Sci. 2022, 23, 13567. [Google Scholar] [CrossRef]
- Noar, R.D.; Daub, M.E. Transcriptome Sequencing of Mycosphaerella fijiensis during Association with Musa acuminata Reveals Candidate Pathogenicity Genes. BMC Genom. 2016, 17, 690. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, M.-C.; Konaté, M.M.; Chen, L.; Das, B.; Karlovich, C.; Williams, P.M.; Evrard, Y.A.; Doroshow, J.H.; McShane, L.M. TPM, FPKM, or Normalized Counts? A Comparative Study of Quantification Measures for the Analysis of RNA-Seq Data from the NCI Patient-Derived Models Repository. J. Transl. Med. 2021, 19, 269. [Google Scholar] [CrossRef] [PubMed]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-Enabled Heat Mapping for All. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.; Vogt, A.; Uslu, T.; Scalzitti, N.; Chennen, K.; Poch, O.; Thompson, J.D. CeGAL: Redefining a Widespread Fungal-Specific Transcription Factor Family Using an In Silico Error-Tracking Approach. J. Fungi 2023, 9, 424. [Google Scholar] [CrossRef] [PubMed]
- Knutson, B.A.; Hahn, S. TFIIB-Related Factors in RNA Polymerase I Transcription. Biochim. Biophys. Acta (BBA)—Gene Regul. Mech. 2013, 1829, 265–273. [Google Scholar] [CrossRef]
- Chen, H.-T.; Hahn, S. Mapping the Location of TFIIB within the RNA Polymerase II Transcription Preinitiation Complex. Cell 2004, 119, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Colbert, T.; Hahn, S. A Yeast TFIIB-Related Factor Involved in RNA Polymerase III Transcription. Genes Dev. 1992, 6, 1940–1949. [Google Scholar] [CrossRef]
- John, E.; Singh, K.B.; Oliver, R.P.; Tan, K. Transcription Factor Control of Virulence in Phytopathogenic Fungi. Mol. Plant Pathol. 2021, 22, 858–881. [Google Scholar] [CrossRef]
- Beranek, A.; Rechberger, G.; Knauer, H.; Wolinski, H.; Kohlwein, S.D.; Leber, R. Identification of a Cardiolipin-Specific Phospholipase Encoded by the Gene CLD1 (YGR110W) in Yeast. J. Biol. Chem. 2009, 284, 11572–11578. [Google Scholar] [CrossRef]
- Rafiei, V.; Vélëz, H.; Tzelepis, G. The Role of Glycoside Hydrolases in Phytopathogenic Fungi and Oomycetes Virulence. Int. J. Mol. Sci. 2021, 22, 9359. [Google Scholar] [CrossRef]
- Fan, X.; Zhang, P.; Batool, W.; Liu, C.; Hu, Y.; Wei, Y.; He, Z.; Zhang, S.-H. Contribution of the Tyrosinase (MoTyr) to Melanin Synthesis, Conidiogenesis, Appressorium Development, and Pathogenicity in Magnaporthe oryzae. J. Fungi 2023, 9, 311. [Google Scholar] [CrossRef]
- Beltrán-García, M.J.; Prado, F.M.; Oliveira, M.S.; Ortiz-Mendoza, D.; Scalfo, A.C.; Pessoa, A.; Medeiros, M.H.G.; White, J.F.; Di Mascio, P. Singlet Molecular Oxygen Generation by Light-Activated DHN-Melanin of the Fungal Pathogen Mycosphaerella fijiensis in Black Sigatoka Disease of Bananas. PLoS ONE 2014, 9, e91616. [Google Scholar] [CrossRef] [PubMed]
- Kantún-Moreno, N.; Vázquez-Euán, R.; Tzec-Simá, M.; Peraza-Echeverría, L.; Grijalva-Arango, R.; Rodríguez-García, C.; James, A.C.; Ramírez-Prado, J.; Islas-Flores, I.; Canto-Canché, B. Genome-Wide in Silico Identification of GPI Proteins in Mycosphaerella fijiensis and Transcriptional Analysis of Two GPI-Anchored β-1,3-Glucanosyltransferases. Mycologia 2013, 105, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Sützl, L.; Foley, G.; Gillam, E.M.J.; Bodén, M.; Haltrich, D. The GMC Superfamily of Oxidoreductases Revisited: Analysis and Evolution of Fungal GMC Oxidoreductases. Biotechnol. Biofuels 2019, 12, 118. [Google Scholar] [CrossRef] [PubMed]
- Donzelli, B.G.G.; Churchill, A.C.L. A Dose-Response approach differentiating virulence of Mycosphaerella fijiensis strains on banana leaves uses either spores or mycelia as inocula. Acta Hortic. 2009, 828, 153–160. [Google Scholar] [CrossRef]
- Yang, Z.; Huang, J.; Geng, J.; Nair, U.; Klionsky, D.J. Atg22 Recycles Amino Acids to Link the Degradative and Recycling Functions of Autophagy. Mol. Biol. Cell 2006, 17, 5094–5104. [Google Scholar] [CrossRef]
- Duan, Z.; Chen, Y.; Huang, W.; Shang, Y.; Chen, P.; Wang, C. Linkage of Autophagy to Fungal Development, Lipid Storage and Virulence in Metarhizium robertsii. Autophagy 2013, 9, 538–549. [Google Scholar] [CrossRef]
- Nguyen, L.N.; Bormann, J.; Le, G.T.T.; Stärkel, C.; Olsson, S.; Nosanchuk, J.D.; Giese, H.; Schäfer, W. Autophagy-Related Lipase FgATG15 of Fusarium Graminearum Is Important for Lipid Turnover and Plant Infection. Fungal Genet. Biol. 2011, 48, 217–224. [Google Scholar] [CrossRef]
- Cingolani, F.; Czaja, M.J. Regulation and Functions of Autophagic Lipolysis. Trends Endocrinol. Metab. 2016, 27, 696–705. [Google Scholar] [CrossRef]
- Wang, Z.-Y.; Soanes, D.M.; Kershaw, M.J.; Talbot, N.J. Functional Analysis of Lipid Metabolism in Magnaporthe Grisea Reveals a Requirement for Peroxisomal Fatty Acid β-Oxidation during Appressorium-Mediated Plant Infection. Mol. Plant-Microbe Interact. 2007, 20, 475–491. [Google Scholar] [CrossRef]
- Campos, A.N.D.R.; Costa, M.D.; Tótola, M.R.; Borges, A.C. Total Lipid and Fatty Acid Accumulation during Basidiospore Formation in the Ectomycorrhizal Fungus pisolithus sp. Rev. Bras. Ciênc. Solo 2008, 32, 1531–1540. [Google Scholar] [CrossRef]
- Burgos-Canul, Y.Y.; Canto-Canché, B.; Berezovski, M.V.; Mironov, G.; Loyola-Vargas, V.M.; Barba De Rosa, A.P.; Tzec-Simá, M.; Brito-Argáez, L.; Carrillo-Pech, M.; Grijalva-Arango, R.; et al. The Cell Wall Proteome from Two Strains of Pseudocercospora Fijiensis with Differences in Virulence. World J. Microbiol. Biotechnol. 2019, 35, 105. [Google Scholar] [CrossRef] [PubMed]
- Vylkova, S. Environmental pH Modulation by Pathogenic Fungi as a Strategy to Conquer the Host. PLoS Pathog. 2017, 13, e1006149. [Google Scholar] [CrossRef] [PubMed]
- Churchill, A.C.L. Mycosphaerella fijiensis, the Black Leaf Streak Pathogen of Banana: Progress towards Understanding Pathogen Biology and Detection, Disease Development, and the Challenges of Control: M. fijiensis, BLS Pathogen of Banana. Mol. Plant Pathol. 2011, 12, 307–328. [Google Scholar] [CrossRef] [PubMed]
- Nfor, T.D.; Ajong, F.D.; Nuincho, L.I. Evaluation of Varietal Response to Black Sigatoka Caused by Mycosphaerella fijiensis Morelet in Banana Nursery. Int. Res. J. Plant Sci. 2011, 2, 299–304. [Google Scholar]
- Arango Isaza, R.E.; Diaz-Trujillo, C.; Dhillon, B.; Aerts, A.; Carlier, J.; Crane, C.F.; De Jong, T.V.; De Vries, I.; Dietrich, R.; Farmer, A.D.; et al. Combating a Global Threat to a Clonal Crop: Banana Black Sigatoka Pathogen Pseudocercospora fijiensis (Synonym Mycosphaerella fijiensis) Genomes Reveal Clues for Disease Control. PLoS Genet. 2016, 12, e1005876. [Google Scholar] [CrossRef] [PubMed]
- Luna-Moreno, D.; Sánchez-Álvarez, A.; Islas-Flores, I.; Canto-Canche, B.; Carrillo-Pech, M.; Villarreal-Chiu, J.; Rodríguez-Delgado, M. Early Detection of the Fungal Banana Black Sigatoka Pathogen Pseudocercospora fijiensis by an SPR Immunosensor Method. Sensors 2019, 19, 465. [Google Scholar] [CrossRef]
- Stover, R.H. Sigatoka Leaf Spots of Bananas And. Plant Dis. 1980, 64, 750. [Google Scholar] [CrossRef]
- Wu, M.-Y.; Mead, M.E.; Lee, M.-K.; Neuhaus, G.F.; Adpressa, D.A.; Martien, J.I.; Son, Y.-E.; Moon, H.; Amador-Noguez, D.; Han, K.-H.; et al. Transcriptomic, Protein-DNA Interaction, and Metabolomic Studies of VosA, VelB, and WetA in Aspergillus nidulans Asexual Spores. mBio 2021, 12, e03128-20. [Google Scholar] [CrossRef]
- Li, C.; Jia, S.; Rajput, S.; Qi, D.; Wang, S. Transcriptional Stages of Conidia Germination and Associated Genes in Aspergillus flavus: An Essential Role for Redox Genes. Toxins 2022, 14, 560. [Google Scholar] [CrossRef]
- Garron, M.-L.; Henrissat, B. The Continuing Expansion of CAZymes and Their Families. Curr. Opin. Chem. Biol. 2019, 53, 82–87. [Google Scholar] [CrossRef]
- Beltrán-García, M.J.; Manzo-Sanchez, G.; Guzmán-González, S.; Arias-Castro, C.; Rodríguez-Mendiola, M.; Avila-Miranda, M.; Ogura, T. Oxidative Stress Response of Mycosphaerella fijiensis, the Causal Agent of Black Leaf Streak Disease in Banana Plants, to Hydrogen Peroxide and Paraquat. Can. J. Microbiol. 2009, 55, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.-M.; Oakley, B.R.; Keller, N.P.; Wang, C.C.C. Unraveling Polyketide Synthesis in Members of the Genus aspergillus. Appl. Microbiol. Biotechnol. 2010, 86, 1719–1736. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Liao, Z.; Liu, T.; Hussain, K.; Chen, J.; Fang, Q.; Wang, J. Vital Roles of Pks11, a Highly Reducing Polyketide Synthase, in Fungal Conidiation, Antioxidant Activity, Conidial Cell Wall Integrity, and UV Tolerance of Beauveria bassiana. J. Invertebr. Pathol. 2021, 181, 107588. [Google Scholar] [CrossRef]
- Jones, D.A.B.; Rozano, L.; Debler, J.W.; Mancera, R.L.; Moolhuijzen, P.M.; Hane, J.K. An Automated and Combinative Method for the Predictive Ranking of Candidate Effector Proteins of Fungal Plant Pathogens. Sci. Rep. 2021, 11, 19731. [Google Scholar] [CrossRef] [PubMed]
- Sperschneider, J.; Dodds, P.N.; Gardiner, D.M.; Singh, K.B.; Taylor, J.M. Improved Prediction of Fungal Effector Proteins from Secretomes with EffectorP 2.0: Prediction of Fungal Effectors with EffectorP 2.0. Mol. Plant Pathol. 2018, 19, 2094–2110. [Google Scholar] [CrossRef]
- Chang, T.-C.; Salvucci, A.; Crous, P.W.; Stergiopoulos, I. Comparative Genomics of the Sigatoka Disease Complex on Banana Suggests a Link between Parallel Evolutionary Changes in Pseudocercospora fijiensis and Pseudocercospora eumusae and Increased Virulence on the Banana Host. PLoS Genet. 2016, 12, e1005904. [Google Scholar] [CrossRef]
- Li, J.; Liu, Q.; Li, J.; Lin, L.; Li, X.; Zhang, Y.; Tian, C. RCO-3 and COL-26 Form an External-to-Internal Module That Regulates the Dual-Affinity Glucose Transport System in Neurospora crassa. Biotechnol. Biofuels 2021, 14, 33. [Google Scholar] [CrossRef]
- Kawabe, M.; Okabe Onokubo, A.; Arimoto, Y.; Yoshida, T.; Azegami, K.; Teraoka, T.; Arie, T. GMC Oxidoreductase, a Highly Expressed Protein in a Potent Biocontrol Agent Fusarium Oxysporum Cong:1-2, Is Dispensable for Biocontrol Activity. J. Gen. Appl. Microbiol. 2011, 57, 207–217. [Google Scholar] [CrossRef]
- Etxebeste, O.; Herrero-García, E.; Cortese, M.S.; Garzia, A.; Oiartzabal-Arano, E.; De Los Ríos, V.; Ugalde, U.; Espeso, E.A. GmcA Is a Putative Glucose-Methanol-Choline Oxidoreductase Required for the Induction of Asexual Development in Aspergillus nidulans. PLoS ONE 2012, 7, e40292. [Google Scholar] [CrossRef]
- De Oliveira, B.V.; Teixeira, G.S.; Reis, O.; Barau, J.G.; Teixeira, P.J.P.L.; Do Rio, M.C.S.; Domingues, R.R.; Meinhardt, L.W.; Paes Leme, A.F.; Rincones, J.; et al. A Potential Role for an Extracellular Methanol Oxidase Secreted by Moniliophthora perniciosa in Witches’ Broom Disease in Cacao. Fungal Genet. Biol. 2012, 49, 922–932. [Google Scholar] [CrossRef]
- Fan, H.; Dong, H.; Xu, C.; Liu, J.; Hu, B.; Ye, J.; Mai, G.; Li, H. Pectin Methylesterases Contribute the Pathogenic Differences between Races 1 and 4 of Fusarium oxysporum f. sp. cubense. Sci. Rep. 2017, 7, 13140. [Google Scholar] [CrossRef] [PubMed]
- Coculo, D.; Del Corpo, D.; Martínez, M.O.; Vera, P.; Piro, G.; De Caroli, M.; Lionetti, V. Arabidopsis Subtilases Promote Defense-Related Pectin Methylesterase Activity and Robust Immune Responses to Botrytis Infection. Plant Physiol. Biochem. 2023, 201, 107865. [Google Scholar] [CrossRef] [PubMed]
- Tjallinks, G.; Boverio, A.; Maric, I.; Rozeboom, H.; Arentshorst, M.; Visser, J.; Ram, A.F.J.; Mattevi, A.; Fraaije, M.W. Structure Elucidation and Characterization of Patulin Synthase, Insights into the Formation of a Fungal Mycotoxin. FEBS J. 2023. [Google Scholar] [CrossRef]
- Albataineh, M.T.; Kadosh, D. Regulatory Roles of Phosphorylation in Model and Pathogenic Fungi. Med. Myco. 2016, 54, 333–352. [Google Scholar] [CrossRef]
- Batool, W.; Liu, C.; Fan, X.; Zhang, P.; Hu, Y.; Wei, Y.; Zhang, S.-H. AGC/AKT Protein Kinase SCH9 Is Critical to Pathogenic Development and Overwintering Survival in Magnaporthe oryzae. J. Fungi 2022, 8, 810. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; St. Leger, R.J.; Fang, W. Pyruvate Accumulation Is the First Line of Cell Defense against Heat Stress in a Fungus. mBio 2017, 8, e01284-17. [Google Scholar] [CrossRef]
- Wu, C.; Zhang, X.; Fang, W. Increasing Pyruvate Concentration Enhances Conidial Thermotolerance in the Entomopathogenic Fungus Metarhizium robertsii. Front. Microbiol. 2019, 10, 519. [Google Scholar] [CrossRef]
- Gao, T.; He, D.; Liu, X.; Ji, F.; Xu, J.; Shi, J. The Pyruvate Dehydrogenase Kinase 2 (PDK2) Is Associated with Conidiation, Mycelial Growth, and Pathogenicity in Fusarium graminearum. Food Prod. Process. Nutr. 2020, 2, 11. [Google Scholar] [CrossRef]
- Dong, Z.; Yang, S.; Lee, B.H. Bioinformatic Mapping of a More Precise Aspergillus niger Degradome. Sci. Rep. 2021, 11, 693. [Google Scholar] [CrossRef]
- Sato, T.K.; Kawano, S.; Endo, T. Role of the Membrane Potential in Mitochondrial Protein Unfolding and Import. Sci. Rep. 2019, 9, 7637. [Google Scholar] [CrossRef]
- Medina, R.; Franco, M.E.E.; Bartel, L.C.; Martinez Alcántara, V.; Saparrat, M.C.N.; Balatti, P.A. Fungal Mitogenomes: Relevant Features to Planning Plant Disease Management. Front. Microbiol. 2020, 11, 978. [Google Scholar] [CrossRef] [PubMed]
- Tsugama, D.; Liu, S.; Takano, T. A bZIP Protein, VIP1, Is a Regulator of Osmosensory Signaling in Arabidopsis. Plant Physiol. 2012, 159, 144–155. [Google Scholar] [CrossRef] [PubMed]
- El Sarraf, N.; Gurel, F.; Tufan, F.; McGuffin, L.J. Characterisation of HvVIP1 and Expression Profile Analysis of Stress Response Regulators in Barley under Agrobacterium and Fusarium Infections. PLoS ONE 2019, 14, e0218120. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Gupta, A.; Yadav, S. Thioredoxins as Molecular Players in Plants, Pests, and Pathogens. In Plant-Pest Interactions: From Molecular Mechanisms to Chemical Ecology; Singh, I.K., Singh, A., Eds.; Springer: Singapore, 2021; pp. 107–125. ISBN 9789811524660. [Google Scholar]
- Filippovich, S.Y.; Bachurina, G.P. Nitric Oxide in Fungal Metabolism (Review). Appl. Biochem. Microbiol. 2021, 57, 694–705. [Google Scholar] [CrossRef]
- Tian, L.; Zhuang, J.; Li, J.-J.; Zhu, H.; Klosterman, S.J.; Dai, X.-F.; Chen, J.-Y.; Subbarao, K.V.; Zhang, D.-D. Thioredoxin VdTrx1, an Unconventional Secreted Protein, Is a Virulence Factor in Verticillium dahliae. Front. Microbiol. 2023, 14, 1130468. [Google Scholar] [CrossRef]
- Gao, M.; Glenn, A.E.; Blacutt, A.A.; Gold, S.E. Fungal Lactamases: Their Occurrence and Function. Front. Microbiol. 2017, 8, 1775. [Google Scholar] [CrossRef]
- Griffiths, S.; Mesarich, C.H.; Saccomanno, B.; Vaisberg, A.; De Wit, P.J.G.M.; Cox, R.; Collemare, J. Elucidation of Cladofulvin Biosynthesis Reveals a Cytochrome P450 Monooxygenase Required for Anthraquinone Dimerization. Proc. Natl. Acad. Sci. USA 2016, 113, 6851–6856. [Google Scholar] [CrossRef]
- Noar, R.D.; Thomas, E.; Daub, M.E. A Novel Polyketide Synthase Gene Cluster in the Plant Pathogenic Fungus Pseudocercospora fijiensis. PLoS ONE 2019, 14, e0212229. [Google Scholar] [CrossRef]
- Glenn, A.E.; Davis, C.B.; Gao, M.; Gold, S.E.; Mitchell, T.R.; Proctor, R.H.; Stewart, J.E.; Snook, M.E. Two Horizontally Transferred Xenobiotic Resistance Gene Clusters Associated with Detoxification of Benzoxazolinones by Fusarium Species. PLoS ONE 2016, 11, e0147486. [Google Scholar] [CrossRef]
- Yu, C.; Qi, J.; Han, H.; Wang, P.; Liu, C. Progress in Pathogenesis Research of Ustilago maydis, and the Metabolites Involved along with Their Biosynthesis. Mol. Plant Pathol. 2023, 24, 495–509. [Google Scholar] [CrossRef]
- Zapater, M.-F.; Abadie, C.; Pignolet, L.; Carlier, J.; Mourichon, X. Diagnosis of Mycosphaerella spp., Responsible for Mycosphaerella Leaf Spot Diseases of Bananas and Plantains, through Morphotaxonomic Observations. Fruits 2008, 63, 389–393. [Google Scholar] [CrossRef]
- He, Q.; McLellan, H.; Boevink, P.C.; Birch, P.R.J. All Roads Lead to Susceptibility: The Many Modes of Action of Fungal and Oomycete Intracellular Effectors. Plant Commun. 2020, 1, 100050. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, D.; Böhlenius, H.; Pedersen, C.; Zhang, Z.; Emmersen, J.; Thordal-Christensen, H. Powdery Mildew Fungal Effector Candidates Share N-Terminal Y/F/WxC-Motif. BMC Genom. 2010, 11, 317. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Shang, X.; Bi, W.; Yu, X.; Liu, D.; Kang, Z.; Wang, X.; Wang, X. Genome-Wide Identification of Effector Candidates with Conserved Motifs from the Wheat Leaf Rust Fungus Puccinia triticina. Front. Microbiol. 2020, 11, 1188. [Google Scholar] [CrossRef] [PubMed]
- Chitty, J.; Fraser, J. Purine Acquisition and Synthesis by Human Fungal Pathogens. Microorganisms 2017, 5, 33. [Google Scholar] [CrossRef]
- Sharma, V.; Chitranshi, N.; Agarwal, A.K. Significance and Biological Importance of Pyrimidine in the Microbial World. Int. J. Med. Chem. 2014, 2014, 202784. [Google Scholar] [CrossRef]
- Sun, M.; Bian, Z.; Luan, Q.; Chen, Y.; Wang, W.; Dong, Y.; Chen, L.; Hao, C.; Xu, J.; Liu, H. Stage-specific Regulation of Purine Metabolism during Infectious Growth and Sexual Reproduction in Fusarium graminearum. New Phytol. 2021, 230, 757–773. [Google Scholar] [CrossRef]
- Nguyen, S.; Truong, J.Q.; Bruning, J.B. Targeting Unconventional Pathways in Pursuit of Novel Antifungals. Front. Mol. Biosci. 2021, 7, 621366. [Google Scholar] [CrossRef]
- Wu, W.; Lan, W.; Wu, C.; Fei, Q. Synthesis and Antifungal Activity of Pyrimidine Derivatives Containing an Amide Moiety. Front. Chem. 2021, 9, 695628. [Google Scholar] [CrossRef]
- Li, C.; Liu, Y.; Ren, X.; Tan, Y.; Jin, L.; Zhou, X. Design, Synthesis and Bioactivity of Novel Pyrimidine Sulfonate Esters Containing Thioether Moiety. Int. J. Mol. Sci. 2023, 24, 4691. [Google Scholar] [CrossRef]
- Pantazopoulou, A.; Diallinas, G. Fungal Nucleobase Transporters. FEMS Microbiol. Rev. 2007, 31, 657–675. [Google Scholar] [CrossRef] [PubMed]
- Qin, T.; Hao, W.; Sun, R.; Li, Y.; Wang, Y.; Wei, C.; Dong, T.; Wu, B.; Dong, N.; Wang, W.; et al. Verticillium dahliae VdTHI20, Involved in Pyrimidine Biosynthesis, Is Required for DNA Repair Functions and Pathogenicity. Int. J. Mol. Sci. 2020, 21, 1378. [Google Scholar] [CrossRef] [PubMed]
- Stallknecht, G.F. Fixation and Incorporation of CO2 into Ribonucleic Acid by Germinating Uredospores of Uromyces phaseoli. Phytopathology 1971, 61, 400. [Google Scholar] [CrossRef]
- Mahanty, B.; Mishra, R.; Joshi, R.K. Cross-Kingdom Small RNA Communication between Plants and Fungal Phytopathogens-Recent Updates and Prospects for Future Agriculture. RNA Biol. 2023, 20, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Wytinck, N.; Sullivan, D.S.; Biggar, K.T.; Crisostomo, L.; Pelka, P.; Belmonte, M.F.; Whyard, S. Clathrin Mediated Endocytosis Is Involved in the Uptake of Exogenous Double-Stranded RNA in the White Mold Phytopathogen Sclerotinia sclerotiorum. Sci. Rep. 2020, 10, 12773. [Google Scholar] [CrossRef]
- Bitencourt, T.A.; Pessoni, A.M.; Oliveira, B.T.M.; Alves, L.R.; Almeida, F. The RNA Content of Fungal Extracellular Vesicles: At the “Cutting-Edge” of Pathophysiology Regulation. Cells 2022, 11, 2184. [Google Scholar] [CrossRef]
- Bartholomew, H.P.; Lichtner, F.J.; Bradshaw, M.; Gaskins, V.L.; Fonseca, J.M.; Bennett, J.W.; Jurick, W.M. Comparative Penicillium spp. Transcriptomics: Conserved Pathways and Processes Revealed in Ungerminated Conidia and during Postharvest Apple Fruit Decay. Microorganisms 2022, 10, 2414. [Google Scholar] [CrossRef]
- Portal, O.; Izquierdo, Y.; De Vleesschauwer, D.; Sánchez-Rodríguez, A.; Mendoza-Rodríguez, M.; Acosta-Suárez, M.; Ocaña, B.; Jiménez, E.; Höfte, M. Analysis of Expressed Sequence Tags Derived from a Compatible Mycosphaerella fijiensis–Banana Interaction. Plant Cell Rep. 2011, 30, 913–928. [Google Scholar] [CrossRef]
- Rodriguez, H.A.; Rodriguez-Arango, E.; Morales, J.G.; Kema, G.; Arango, R.E. Defense Gene Expression Associated with Biotrophic Phase of Mycosphaerella fijiensis M. Morelet Infection in Banana. Plant Dis. 2016, 100, 1170–1175. [Google Scholar] [CrossRef]
- Rodriguez, H.A.; Hidalgo, W.F.; Sanchez, J.D.; Menezes, R.C.; Schneider, B.; Arango, R.E.; Morales, J.G. Differential Regulation of Jasmonic Acid Pathways in Resistant (Calcutta 4) and Susceptible (Williams) Banana Genotypes during the Interaction with Pseudocercospora fijiensis. Plant Pathol. 2020, 69, 872–882. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Read Statistics | # seqs |
|---|---|
| Raw reads | 23,429 |
| Clean reads | 22,940 |
| Total contigs | 2501 |
| Total mapped | 2424 |
| GC content | 53% |
| Sequence repetition (%) | 85.16% |
| JGI ID | NCBI Description | GO Term | RPKM |
|---|---|---|---|
| 210035 | K(+)/H(+) antiporter 1 | GO:0015297 GO:0016020 GO:1902600 | 555.70 |
| 158260 | Protein vip1 | GO:0003723 GO:0004408 GO:0004843 GO:0005743 GO:0006511 GO:0016579 GO:0046872 | 555.70 |
| 52736 | Putative glucose transporter rco-3 | GO:0005829 GO:0008643 GO:0009086 GO:0015144 GO:0015295 GO:0016020 GO:0035999 GO:0055085 GO:0071949 GO:0106312 GO:0106313 | 431.76 |
| 77759 | GMC oxidoreductase | GO:0016614 GO:0050660 | 417.66 |
| 42490 | Mono-oxygenase | GO:0004499 GO:0050660 GO:0050661 | 412.41 |
| 156715 | Serine/threonine-protein kinase | GO:0000781 GO:0000922 GO:0000939 GO:0004674 GO:0005524 GO:0005813 GO:0005814 GO:0005876 GO:0006468 GO:0007052 GO:0010971 GO:0032133 GO:0033316 GO:0034501 GO:0034503 GO:0035175 GO:0090148 GO:0090267 GO:0120110 GO:0140429 GO:0140602 GO:1902412 GO:1903380 GO:1904967 GO:1905824 GO:1990023 GO:1990385 | 395.61 |
| 216890 | Fluconazole resistance protein 1 | GO:0000981 GO:0006357 GO:0008270 | 388.58 |
| 51000 | Thioredoxin-like protein | GO:0003756 GO:0005773 GO:0005783 GO:0005886 | 386.86 |
| 212439 | Uncharacterized protein | GO:0016020 | 364.29 |
| 212045 | FAD-linked oxidoreductase sorD | GO:0016020 GO:0016491 GO:0071949 GO:1901360 | 359.30 |
| Processes | Genes Annotated | RPKM |
|---|---|---|
| Adhesion, perception, and signal transduction | 59 | 16.35–395.61 |
| Conidial germination | 71 | 20.87–266.56 |
| Ergosterol biosynthesis | 10 | 82.63–224.76 |
| Glycosyl hydrolases (secreted) | 16 | 44.47–158.96 |
| Autophagy | 22 | 42.40–224.18 |
| Gene silencing | 9 | 91.84–166.85 |
| JGI ID | Homolog in PHI | PHI Description | GO Term | RPKM |
|---|---|---|---|---|
| 52736 | Magnaporthe oryzae (G4N740) | Unaffected pathogenicity | GO:0005829 GO:0008643 GO:0009086 GO:0015144 GO:0015295 GO:0016020 GO:0035999 GO:0055085 GO:0071949 GO:0106312 GO:0106313 | 431.76 |
| 77759 | Brucella abortus (Q2YIV2) | Reduced virulence | GO:0016614 GO:0050660 | 417.66 |
| 156715 | Fusarium graminearum (V6RFV4) | Lethal | GO:0000781 GO:0000922 GO:0000939 GO:0004674 GO:0005524 GO:0005813 GO:0005814 GO:0005876 GO:0006468 GO:0007052 GO:0010971 GO:0032133 GO:0033316 GO:0034501 GO:0034503 GO:0035175 GO:0090148 GO:0090267 GO:0120110 GO:0140429 GO:0140602 GO:1902412 GO:1903380 GO:1904967 GO:1905824 GO:1990023 GO:1990385 | 395.61 |
| 209751 | Magnaporthe oryzae (G4MXS1) | Unaffected pathogenicity | GO:0000050 GO:0000287 GO:0004053 GO:0004743 GO:0005524 GO:0005737 GO:0006091 GO:0006525 GO:0016301 GO:0016310 GO:0016491 GO:0016836 GO:0016866 GO:0030955 GO:0042866 GO:0046031 GO:0046034 GO:0061621 | 351.60 |
| 129014 | Parastagonospora nodorum (Q6Y392) | Unaffected pathogenicity | GO:0016020 GO:0035442 GO:0071916 | 346.95 |
| 87169 | Helicobacter pylori (O25656) | Reduced virulence | GO:0004222 GO:0006627 GO:0016020 GO:0017087 GO:0044237 GO:0046872 | 333.70 |
| 57565 | Botrytis cinerea (Q9UW03) | Unaffected pathogenicity | GO:0005524 GO:0015288 GO:0016887 GO:0034219 GO:0046930 GO:0140359 | 296.71 |
| 212004 | Fulvia fulva (B3VBK9) | Effector | - | 294.05 |
| 79446 | Xanthomonas campestris (Q4UWM4) | Reduced virulence | GO:0016020 GO:0031411 GO:0043227 | 282.64 |
| 135140 | Cryptococcus neoformans (Q874K8) | Reduced virulence | GO:0003735 GO:0004585 GO:0005247 GO:0005769 GO:0005773 GO:0005794 GO:0005840 GO:0005886 GO:0006412 GO:0015078 GO:0015297 GO:0016597 GO:0016787 GO:0019240 GO:0022853 GO:0042450 GO:1902476 GO:1990904 | 269.85 |
| JGI ID | NCBI Description | GO Term | RPKM |
|---|---|---|---|
| 42490 | Mono-oxygenase | GO:0004499 GO:0050660 GO:0050661 | 412.41 |
| 212439 | Uncharacterized protein | GO:0016020 | 364.29 |
| 88290 | Putative mitochondrial recombination protein | GO:0000002 GO:0000150 GO:0003697 GO:0003735 GO:0004553 GO:0005634 GO:0005762 GO:0005975 GO:0006261 GO:0006310 GO:0010557 GO:0016020 GO:0031328 GO:0045740 GO:0071897 | 349.72 |
| 202529 | Oligopeptide transporter | GO:0000329 GO:0035672 GO:0035673 | 313.75 |
| 181747 | Uncharacterized protein | - | 303.58 |
| 212004 | Extracellular protein 6 | - | 294.05 |
| 210624 | Kinase-like protein | GO:0004672 GO:0005524 GO:0006468 | 266.56 |
| 163251 | Ferric/cupric reductase transmembrane component 7 | GO:0000293 GO:0005886 GO:0006826 GO:0006879 GO:0009987 GO:0065008 | 266.56 |
| 52972 | ECP2 protein | GO:0003677 GO:0045892 GO:0046677 | 266.56 |
| 89258 | Putative altered inheritance of mitochondria protein | - | 263.34 |
| ID Gene | NCBI Description | GO Term | RPKM |
| Pfijiensis_857 | No hit | - | 216.41 |
| Pfijiensis_645 | No hit | - | 215.34 |
| Pfijiensis_2011 | No hit | - | 211.18 |
| Pfijiensis_1733 | No hit | - | 209.16 |
| Pfijiensis_887 | No hit | - | 190.89 |
| Pfijiensis_944 | No hit | - | 188.43 |
| Pfijiensis_1491 | No hit | - | 187.62 |
| Pfijiensis_1874 | No hit | - | 180.64 |
| Pfijiensis_424 | No hit | - | 174.16 |
| Pfijiensis_1096 | No hit | - | 173.47 |
| JGI ID | NCBI description | GO term | RPKM |
| Pfijiensis_2311 | No hit | - | 105.08 |
| Pfijiensis_287 | No hit | - | 64.47 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carreón-Anguiano, K.G.; Gómez-Tah, R.; Pech-Balan, E.; Ek-Hernández, G.E.; De los Santos-Briones, C.; Islas-Flores, I.; Canto-Canché, B. Pseudocercospora fijiensis Conidial Germination Is Dominated by Pathogenicity Factors and Effectors. J. Fungi 2023, 9, 970. https://doi.org/10.3390/jof9100970
Carreón-Anguiano KG, Gómez-Tah R, Pech-Balan E, Ek-Hernández GE, De los Santos-Briones C, Islas-Flores I, Canto-Canché B. Pseudocercospora fijiensis Conidial Germination Is Dominated by Pathogenicity Factors and Effectors. Journal of Fungi. 2023; 9(10):970. https://doi.org/10.3390/jof9100970
Chicago/Turabian StyleCarreón-Anguiano, Karla Gisel, Rufino Gómez-Tah, Efren Pech-Balan, Gemaly Elisama Ek-Hernández, César De los Santos-Briones, Ignacio Islas-Flores, and Blondy Canto-Canché. 2023. "Pseudocercospora fijiensis Conidial Germination Is Dominated by Pathogenicity Factors and Effectors" Journal of Fungi 9, no. 10: 970. https://doi.org/10.3390/jof9100970