
A Putative TRAPα Protein of Microsporidia Nosema bombycis Exhibits Non-Canonical Alternative Polyadenylation in Transcripts

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Spores Purification
2.2. Genomic DNA and Total RNA Extraction
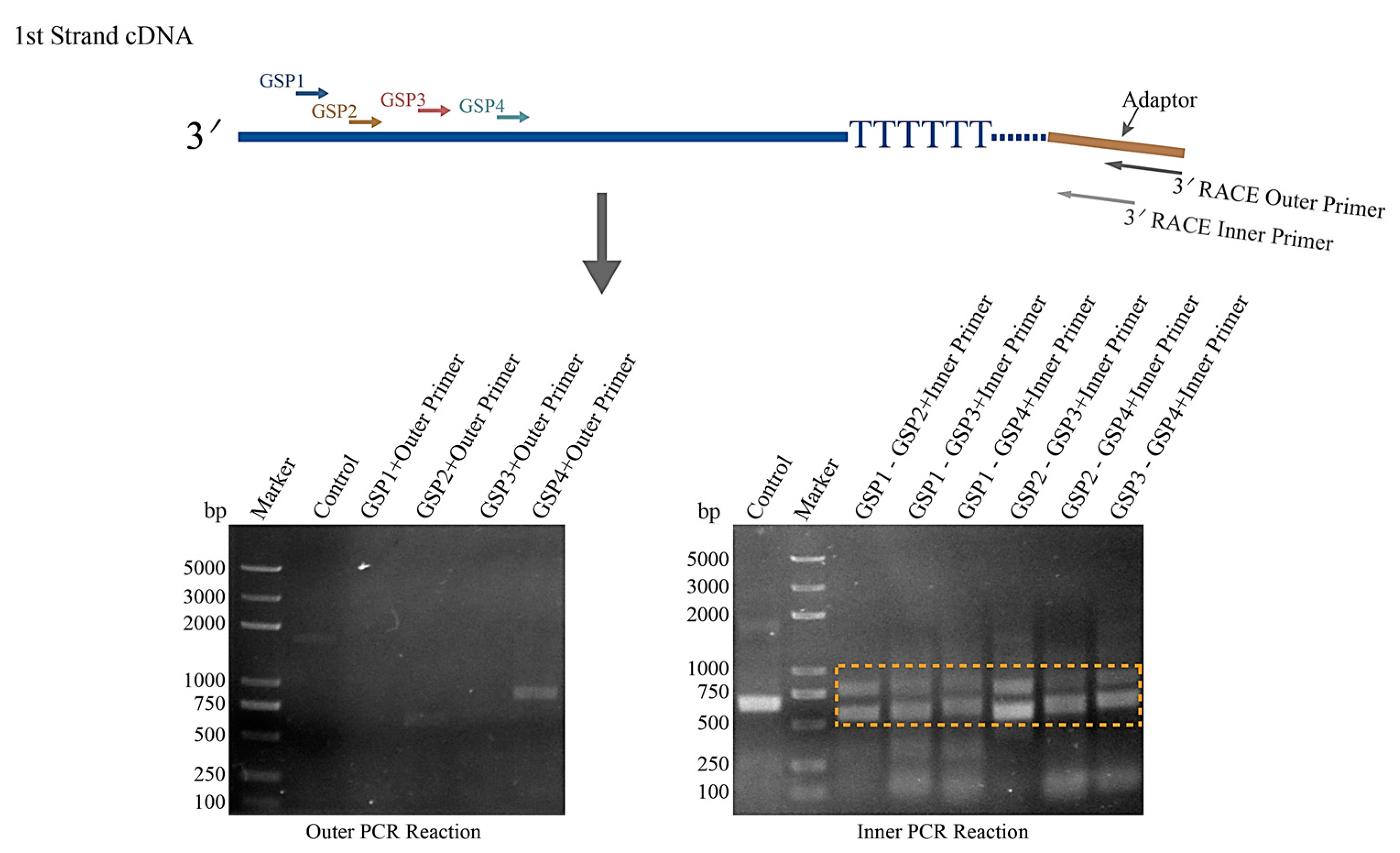
2.3. 3′ RACE Analysis
2.4. Pathogen Inoculation
2.5. The Specific HNbTRAPα-Antisera Preparation
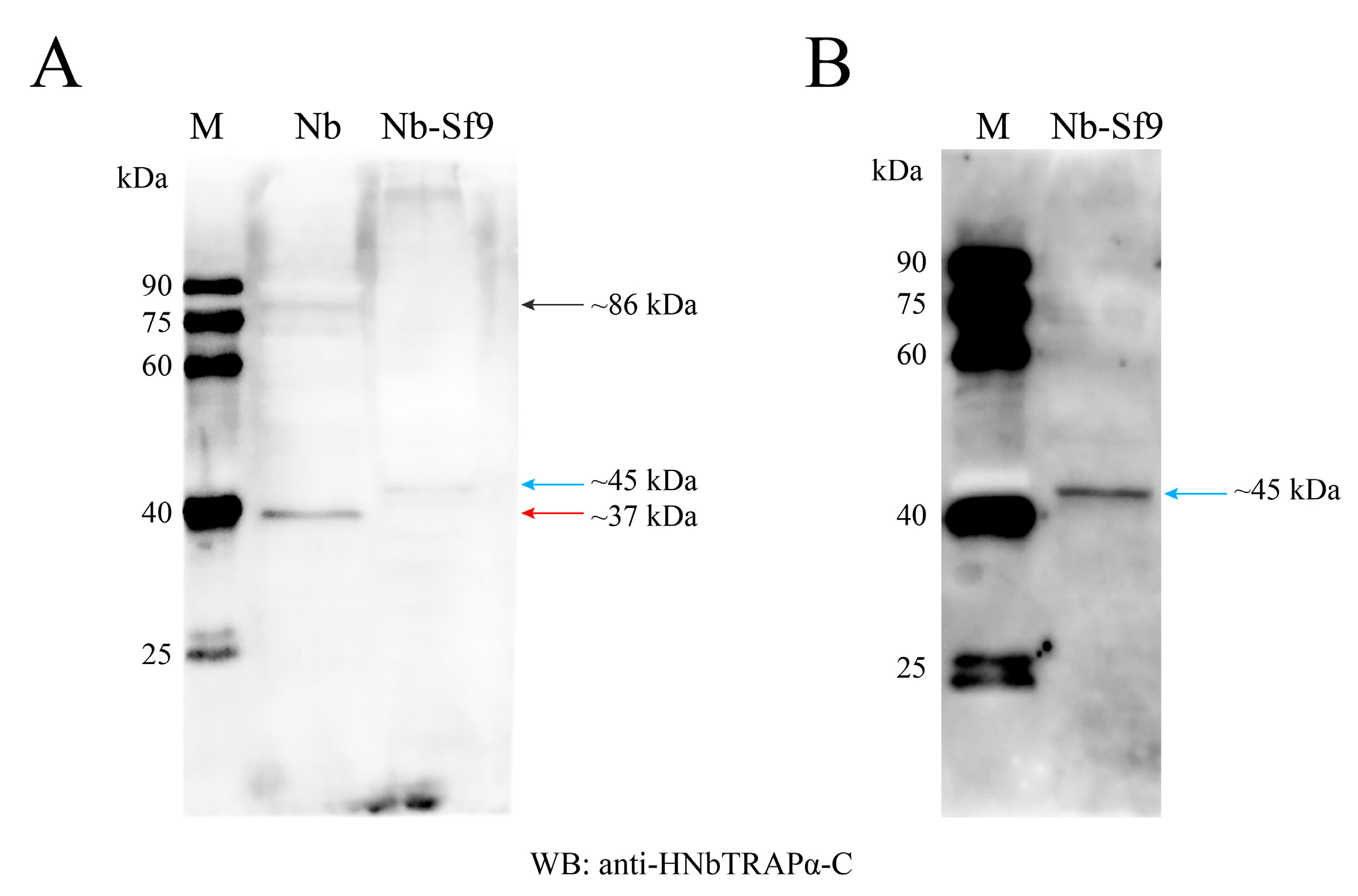
2.6. Immunoblottings
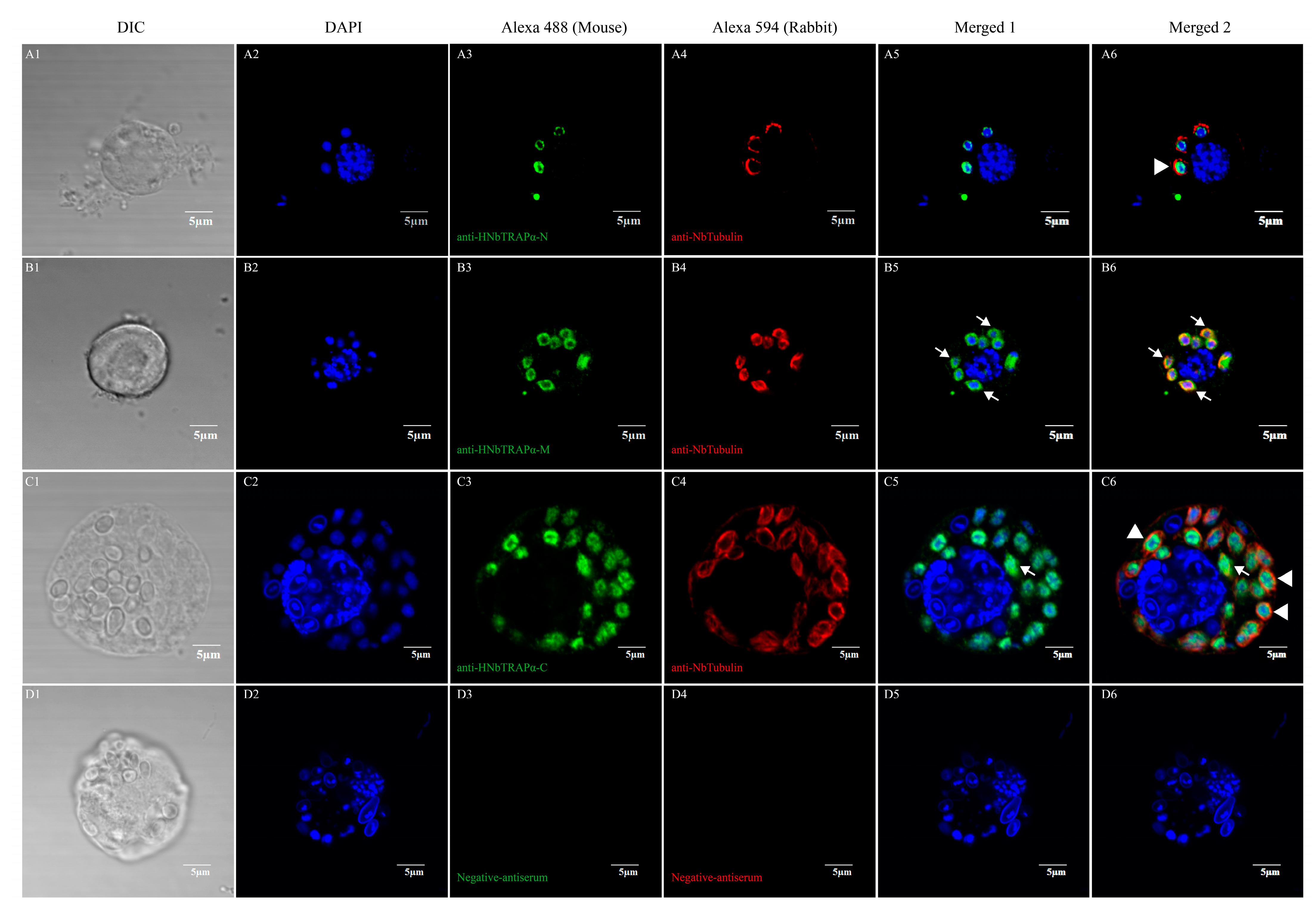
2.7. Immunofluorescence Analysis
3. Results
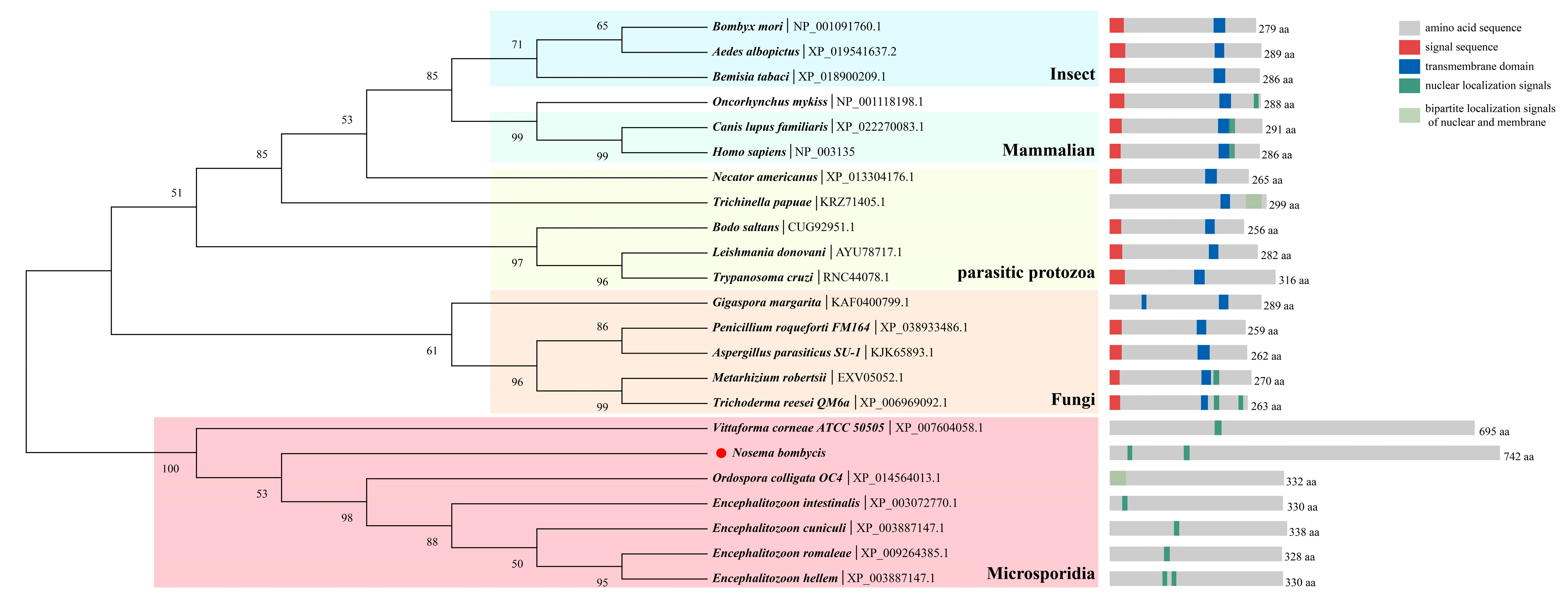
3.1. Sequence Analysis of HNbTRAPα
3.2. Two Transcriptional Isoform of HNbTRAPα
3.3. Three Protein Productions of HNbTRAPα Were Detected in N. bombycis
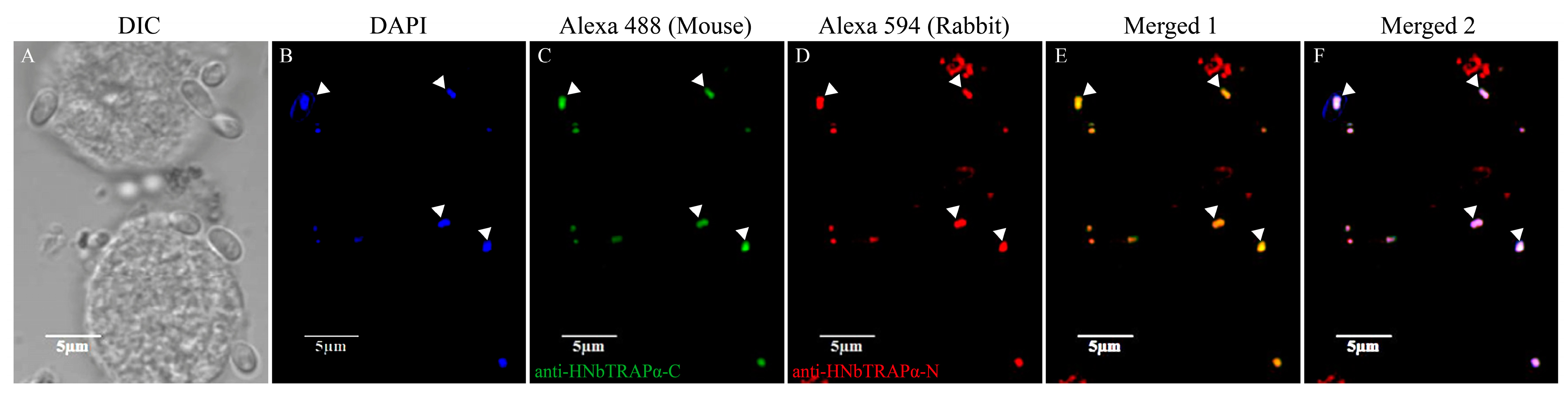
3.4. HNbTRAPα Localized at the Perinuclear Region or in the Nucleus during Different Development Stages
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cavalier-Smith, T. A 6-Klngdom Classification And A Unified Phylogeny. In Intracellular Space as Oligogenetic Ecosystem, Proceedings; Hainfried, E.A.S., Werner, S., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 1983; pp. 1027–1034. [Google Scholar]
- Han, B.; Weiss, L.M. Microsporidia: Obligate Intracellular Pathogens Within the Fungal Kingdom. Microbiol. Spectr. 2017, 5, FUNK-0018–2016. [Google Scholar] [CrossRef] [Green Version]
- Katinka, M.D.; Duprat, S.; Cornillot, E.; Metenier, G.; Thomarat, F.; Prensier, G.; Barbe, V.; Peyretaillade, E.; Brottier, P.; Wincker, P.; et al. Genome sequence and gene compaction of the eukaryote parasite Encephalitozoon cuniculi. Nature 2001, 414, 450–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, T.Y.; Pelin, A.; Bonen, L.; Ahrendt, S.; Sain, D.; Corradi, N.; Stajich, J.E. Shared signatures of parasitism and phylogenomics unite Cryptomycota and microsporidia. Curr. Biol. 2013, 23, 1548–1553. [Google Scholar] [CrossRef] [Green Version]
- James, T.Y.; Stajich, J.E.; Hittinger, C.T.; Rokas, A. Toward a Fully Resolved Fungal Tree of Life. Annu. Rev. Microbiol. 2020, 74, 291–313. [Google Scholar] [CrossRef] [PubMed]
- Corradi, N. Microsporidia: Eukaryotic Intracellular Parasites Shaped by Gene Loss and Horizontal Gene Transfers. Annu. Rev. Microbiol. 2015, 69, 167–183. [Google Scholar] [CrossRef]
- Barandun, J.; Hunziker, M.; Vossbrinck, C.R.; Klinge, S. Evolutionary compaction and adaptation visualized by the structure of the dormant microsporidian ribosome. Nat. Microbiol. 2019, 4, 1798–1804. [Google Scholar] [CrossRef]
- Pan, G.; Xu, J.; Li, T.; Xia, Q.; Liu, S.L.; Zhang, G.; Li, S.; Li, C.; Liu, H.; Yang, L.; et al. Comparative genomics of parasitic silkworm microsporidia reveal an association between genome expansion and host adaptation. BMC Genom. 2013, 14, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corradi, N.; Pombert, J.F.; Farinelli, L.; Didier, E.S.; Keeling, P.J. The complete sequence of the smallest known nuclear genome from the microsporidian Encephalitozoon intestinalis. Nat. Commun. 2010, 1, 77. [Google Scholar] [CrossRef] [Green Version]
- Jespersen, N.; Monrroy, L.; Barandun, J. Impact of Genome Reduction in Microsporidia. Exp Suppl. 2022, 114, 1–42. [Google Scholar] [CrossRef]
- Akiyoshi, D.E.; Morrison, H.G.; Lei, S.; Feng, X.; Zhang, Q.; Corradi, N.; Mayanja, H.; Tumwine, J.K.; Keeling, P.J.; Weiss, L.M.; et al. Genomic survey of the non-cultivatable opportunistic human pathogen, Enterocytozoon bieneusi. PLoS Pathog. 2009, 5, e1000261. [Google Scholar] [CrossRef] [Green Version]
- Desjardins, C.A.; Sanscrainte, N.D.; Goldberg, J.M.; Heiman, D.; Young, S.; Zeng, Q.; Madhani, H.D.; Becnel, J.J.; Cuomo, C.A. Contrasting host-pathogen interactions and genome evolution in two generalist and specialist microsporidian pathogens of mosquitoes. Nat. Commun. 2015, 6, 7121. [Google Scholar] [CrossRef] [Green Version]
- Keeling, P.J.; Corradi, N.; Morrison, H.G.; Haag, K.L.; Ebert, D.; Weiss, L.M.; Akiyoshi, D.E.; Tzipori, S. The reduced genome of the parasitic microsporidian Enterocytozoon bieneusi lacks genes for core carbon metabolism. Genome Biol. Evol. 2010, 2, 304–309. [Google Scholar] [CrossRef]
- Campbell, S.E.; Williams, T.A.; Yousuf, A.; Soanes, D.M.; Paszkiewicz, K.H.; Williams, B.A. The genome of Spraguea lophii and the basis of host-microsporidian interactions. PLoS Genet. 2013, 9, e1003676. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.C.; Gill, E.E.; Roy, S.W.; Fast, N.M. Constrained intron structures in a microsporidian. Mol. Biol. Evol. 2010, 27, 1979–1982. [Google Scholar] [CrossRef] [PubMed]
- Senderskiy, I.V.; Timofeev, S.A.; Seliverstova, E.V.; Pavlova, O.A.; Dolgikh, V.V. Secretion of Antonospora (Paranosema) locustae proteins into infected cells suggests an active role of microsporidia in the control of host programs and metabolic processes. PLoS ONE 2014, 9, e93585. [Google Scholar] [CrossRef] [Green Version]
- Kurze, C.; Dosselli, R.; Grassl, J.; Le Conte, Y.; Kryger, P.; Baer, B.; Moritz, R.F. Differential proteomics reveals novel insights into Nosema-honey bee interactions. Insect. Biochem. Mol. Biol. 2016, 79, 42–49. [Google Scholar] [CrossRef]
- Luo, J.; He, Q.; Xu, J.Z.; Xu, C.; Han, Y.Z.; Gao, H.L.; Meng, X.Z.; Pan, G.Q.; Li, T.; Zhou, Z.Y. Microsporidia infection upregulates host energy metabolism but maintains ATP homeostasis. J. Invertebr. Pathol. 2021, 186, 107596. [Google Scholar] [CrossRef] [PubMed]
- Dean, P.; Hirt, R.P.; Embley, T.M. Microsporidia: Why Make Nucleotides if You can steal them. PLoS Pathog. 2016, 12, e1005870. [Google Scholar] [CrossRef] [Green Version]
- Nakjang, S.; Williams, T.A.; Heinz, E.; Watson, A.K.; Foster, P.G.; Sendra, K.M.; Heaps, S.E.; Hirt, R.P.; Martin Embley, T. Reduction and expansion in microsporidian genome evolution: New insights from comparative genomics. Genome Biol. Evol. 2013, 5, 2285–2303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, L.M.; Becnel, J.J. Microsporidia: Pathogens of Opportunity; Wiley Blackwell: Oxford, UK, 2014. [Google Scholar]
- Elkon, R.; Ugalde, A.P.; Agami, R. Alternative cleavage and polyadenylation: Extent, regulation and function. Nat. Rev. Genet. 2013, 14, 496–506. [Google Scholar] [CrossRef]
- Neve, J.; Patel, R.; Wang, Z.; Louey, A.; Furger, A.M. Cleavage and polyadenylation: Ending the message expands gene regulation. RNA Biol. 2017, 14, 865–890. [Google Scholar] [CrossRef] [Green Version]
- Tellez-Robledo, B.; Manzano, C.; Saez, A.; Navarro-Neila, S.; Silva-Navas, J.; de Lorenzo, L.; Gonzalez-Garcia, M.P.; Toribio, R.; Hunt, A.G.; Baigorri, R.; et al. The polyadenylation factor FIP1 is important for plant development and root responses to abiotic stresses. Plant J. 2019, 99, 1203–1219. [Google Scholar] [CrossRef]
- Ogorodnikov, A.; Levin, M.; Tattikota, S.; Tokalov, S.; Hoque, M.; Scherzinger, D.; Marini, F.; Poetsch, A.; Binder, H.; Macher-Göppinger, S.; et al. Transcriptome 3’end organization by PCF11 links alternative polyadenylation to formation and neuronal differentiation of neuroblastoma. Nat. Commun. 2018, 9, 5331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shell, S.A.; Hesse, C.; Morris, S.M., Jr.; Milcarek, C. Elevated levels of the 64-kDa cleavage stimulatory factor (CstF-64) in lipopolysaccharide-stimulated macrophages influence gene expression and induce alternative poly(A) site selection. J. Biol. Chem. 2005, 280, 39950–39961. [Google Scholar] [CrossRef] [Green Version]
- Pfeffer, S.; Dudek, J.; Schaffer, M.; Ng, B.G.; Albert, S.; Plitzko, J.M.; Baumeister, W.; Zimmermann, R.; Freeze, H.H.; Engel, B.D.; et al. Dissecting the molecular organization of the translocon-associated protein complex. Nat. Commun. 2017, 8, 14516. [Google Scholar] [CrossRef] [PubMed]
- Wiedmann, M.; Kurzchalia, T.V.; Hartmann, E.; Rapoport, T.A. A signal sequence receptor in the endoplasmic reticulum membrane. Nature 1987, 328, 830–833. [Google Scholar] [CrossRef]
- Görlich, D.; Prehn, S.; Hartmann, E.; Herz, J.; Otto, A.; Kraft, R.; Wiedmann, M.; Knespel, S.; Dobberstein, B.; Rapoport, T.A. The signal sequence receptor has a second subunit and is part of a translocation complex in the endoplasmic. J. Cell Biol. 1990, 111, 2283–2294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, E.; Görlich, D.; Kostka, S.; Otto, A.; Kraft, R.; Knespel, S.; Bürger, E.; Rapoport, T.A.; Prehn, S. A tetrameric complex of membrane proteins in the endoplasmic reticulum. Eur. J. Biochem. 1993, 214, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Fons, R.D.; Bogert, B.A.; Hegde, R.S. Substrate-specific function of the translocon-associated protein complex during translocation across the ER membrane. J. Cell Biol. 2003, 160, 529–539. [Google Scholar] [CrossRef] [Green Version]
- Prehn, S.; Herz, J.; Hartmann, E.; Frank, R.; Roemisch, K.; Dobberstein, B.; Rapoport, T.A. Structure and biosynthesis of the signal-sequence receptor. Eur. J. Biochem 1990, 188, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, E.; Prehn, S. The N-terminal region of the α-subunit of the TRAP complex has a conserved cluster of negative charges. FEBS Lett. 1994, 349, 324–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirama, T.; Miller, C.W.; Koeffler, H.P. Translocon-associated protein α transcripts are induced by granulocyte-macrophage colony-stimulating factor and exhibit complex alternative polyadenylation. FEBS Lett. 1999, 455, 223–227. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.K.; Mangalam, A.K.; Dwivedi, S.; Naik, S. Primer premier: Program for design of degenerate primers from a protein sequence. Biotechniques 1998, 24, 318–319. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Guo, W.; Dang, X.; Huang, Y.; Liu, F.; Meng, X.; An, Y.; Long, M.; Bao, J.; Zhou, Z.; et al. Easy labeling of proliferative phase and sporogonic phase of microsporidia Nosema bombycis in host cells. PLoS ONE 2017, 12, e0179618. [Google Scholar] [CrossRef] [Green Version]
- Williams, B.A.P.; Williams, T.A.; Trew, J. Comparative Genomics of Microsporidia. Exp. Suppl. 2022, 114, 43–69. [Google Scholar] [CrossRef]
- Cuomo, C.A.; Desjardins, C.A.; Bakowski, M.A.; Goldberg, J.; Ma, A.T.; Becnel, J.J.; Didier, E.S.; Fan, L.; Heiman, D.I.; Levin, J.Z.; et al. Microsporidian genome analysis reveals evolutionary strategies for obligate intracellular growth. Genome Res. 2012, 22, 2478–2488. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.; Manley, J.L. Alternative polyadenylation of mRNA precursors. Nat. Rev. Mol. Cell Biol. 2017, 18, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Burgess, R.J.; Zhang, Z. Histone chaperones in nucleosome assembly and human disease. Nat. Struct. Mol. Biol. 2013, 20, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Loyola, A.; Almouzni, G. Histone chaperones, a supporting role in the limelight. Biochim. Biophys. Acta 2004, 1677, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Eitoku, M.; Sato, L.; Senda, T.; Horikoshi, M. Histone chaperones: 30 years from isolation to elucidation of the mechanisms of nucleosome assembly and disassembly. Cell. Mol. Life Sci. 2008, 65, 414–444. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence (5′-3′) | Amplicon Region or Initiation Site (bp) |
|---|---|---|
| HNbTRAPα-N_F | CGGGATCCATGACAAAAGCCCTTCATAA | 1–696 |
| HNbTRAPα-N_R | GCGTCGACAGGATTTCGAGCCATTAGATG | |
| HNbTRAPα-M_F | CGCGGATCCATGCCTTACAATTCATTC | 697–981 |
| HNbTRAPα-M_R | GTCGTCGACGTTTATTTTTGGCTGTAA | |
| HNbTRAPα-C_F | CGGGATCCATGGAGACTATTGAGAGC | 1704–2226 |
| HNbTRAPα-C_R | GTCGTCGACATTATACAAGCGTATTAT | |
| HNbTRAPα-GSP1 | GGAGAAATGAAGCAGACACC | 229 |
| HNbTRAPα-GSP2 | CAGATAGAAGCAGCAGTAGG | 376 |
| HNbTRAPα-GSP3 | GGATGATGAAGAAGAGGAGG | 405 |
| HNbTRAPα-GSP4 | AGAGAGTGCGTAATAGGGAG | 443 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Yu, Y.; Sun, Q.; Yu, Y.; Chen, J.; Li, T.; Meng, X.; Pan, G.; Zhou, Z. A Putative TRAPα Protein of Microsporidia Nosema bombycis Exhibits Non-Canonical Alternative Polyadenylation in Transcripts. J. Fungi 2023, 9, 407. https://doi.org/10.3390/jof9040407
Wu Y, Yu Y, Sun Q, Yu Y, Chen J, Li T, Meng X, Pan G, Zhou Z. A Putative TRAPα Protein of Microsporidia Nosema bombycis Exhibits Non-Canonical Alternative Polyadenylation in Transcripts. Journal of Fungi. 2023; 9(4):407. https://doi.org/10.3390/jof9040407
Chicago/Turabian StyleWu, Yujiao, Ying Yu, Quan Sun, Yixiang Yu, Jie Chen, Tian Li, Xianzhi Meng, Guoqing Pan, and Zeyang Zhou. 2023. "A Putative TRAPα Protein of Microsporidia Nosema bombycis Exhibits Non-Canonical Alternative Polyadenylation in Transcripts" Journal of Fungi 9, no. 4: 407. https://doi.org/10.3390/jof9040407