
Organic Nitrogen Supplementation Increases Vegetative and Reproductive Biomass in a Versatile White Rot Fungus


{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Strains, Culture Maintenance, Growth Rate and Fruiting Setup
2.2. Assessment of Mycelial Growth and Reproductive Success
2.3. Statistical Analysis
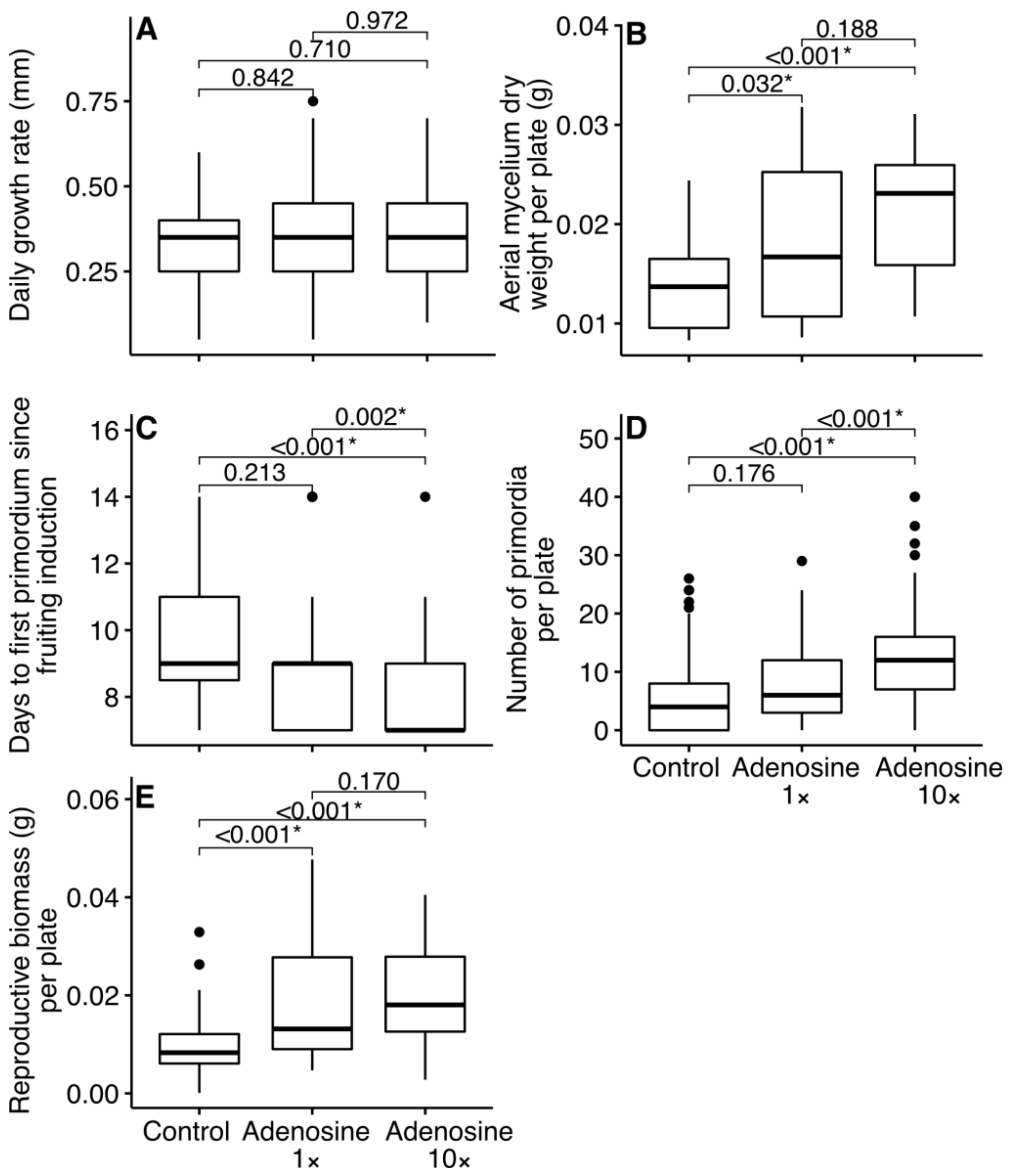
3. Results
4. Discussion
4.1. Effect of Adenosine Addition on Mycelium and Primordia Formation
4.2. Effect of Adenosine Addition on Reproductive Biomass in Fruit Bodies
4.3. Study Limitations and Future Perspective
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bar-On, Y.M.; Phillips, R.; Milo, R. The Biomass Distribution on Earth. Proc. Natl. Acad. Sci. USA 2018, 115, 6506–6511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, L.G.; Riley, R.; Tritt, A.; Adam, C.; Daum, C.; Floudas, D.; Sun, H.; Yadav, J.S.; Pangilinan, J.; Larsson, K.-H.; et al. Comparative Genomics of Early-Diverging Mushroom-Forming Fungi Provides Insights into the Origins of Lignocellulose Decay Capabilities. Mol. Biol. Evol. 2015, 33, 959–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, D.K.; Rühl, M.; Mishra, B.; Kleofas, V.; Hofrichter, M.; Herzog, R.; Pecyna, M.J.; Sharma, R.; Kellner, H.; Hennicke, F.; et al. The Genome Sequence of the Commercially Cultivated Mushroom Agrocybe aegerita Reveals a Conserved Repertoire of Fruiting-Related Genes and a Versatile Suite of Biopolymer-Degrading Enzymes. BMC Genom. 2018, 19, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jennings, D.H. The Physiology of Fungal Nutrition; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Cornwell, W.K.; Cornelissen, J.H.C.; Allison, S.D.; Bauhus, J.; Eggleton, P.; Preston, C.M.; Scarff, F.; Weedon, J.T.; Wirth, C.; Zanne, A.E. Plant Traits and Wood Fates across the Globe: Rotted, Burned, or Consumed? Glob. Chang. Biol. 2009, 15, 2431–2449. [Google Scholar] [CrossRef] [Green Version]
- Moore, D.; Robson, G.D.; Trinci, A.P. 21st Century Guidebook to Fungi; Cambridge University Press: Cambridge, UK, 2020. [Google Scholar]
- Allison, S.D.; Vitousek, P.M. Responses of Extracellular Enzymes to Simple and Complex Nutrient Inputs. Soil Biol. Biochem. 2005, 37, 937–944. [Google Scholar] [CrossRef]
- Noordeloos, M.E.; Kuyper, T.W.; Vellinga, E. Flora Agaricina Neerlandica; CRC Press: Boca Raton, FL, USA, 2005; Volume 6, pp. 204–221. [Google Scholar]
- Watkinson, S.; Davison, E.; Bramah, J. The Effect of Nitrogen Availability on Growth and Cellulolysis by Serpula lacrimans. New Phytol. 1981, 89, 295–305. [Google Scholar] [CrossRef]
- Domondon, D.L.; He, W.; De Kimpe, N.; Höfte, M.; Poppe, J. β-Adenosine, a Bioactive Compound in Grass Chaff Stimulating Mushroom Production. Phytochemistry 2004, 65, 181–187. [Google Scholar] [CrossRef]
- Tlalka, M.; Bebber, D.; Darrah, P.; Watkinson, S.; Fricker, M. Quantifying Dynamic Resource Allocation Illuminates Foraging Strategy in Phanerochaete velutina. Fungal Genet. Biol. 2008, 45, 1111–1121. [Google Scholar] [CrossRef]
- Weißhaupt, P.; Pritzkow, W.; Noll, M. Nitrogen Metabolism of Wood Decomposing Basidiomycetes and Their Interaction with Diazotrophs as Revealed by IRMS. Int. J. Mass Spectrom. 2011, 307, 225–231. [Google Scholar] [CrossRef]
- Philpott, T.; Prescott, C.; Chapman, W.; Grayston, S. Nitrogen Translocation and Accumulation by a Cord-Forming Fungus (Hypholoma fasciculare) into Simulated Woody Debris. For. Ecol. Manag. 2014, 315, 121–128. [Google Scholar] [CrossRef]
- Zhong, H.; Smith, C.; Robinson, B.; Kim, Y.-N.; Dickinson, N. Plant Litter Variability and Soil N Mobility. Soil Res. 2016, 55, 253–263. [Google Scholar] [CrossRef]
- Rinne, K.T.; Rajala, T.; Peltoniemi, K.; Chen, J.; Smolander, A.; Mäkipää, R. Accumulation Rates and Sources of External Nitrogen in Decaying Wood in a Norway Spruce Dominated Forest. Funct. Ecol. 2017, 31, 530–541. [Google Scholar] [CrossRef]
- Ganney, G. Observations on Cultural Aspects Affecting the Consistent Production of Good Quality Closed Hybrid Mushrooms. In Developments in Crop Science; Elsevier: Amsterdam, The Netherlands, 1987; Volume 10, pp. 295–300. [Google Scholar]
- Kües, U. Life History and Developmental Processes in the Basidiomycete Coprinus cinereus. Microbiol. Mol. Biol. Rev. 2000, 64, 316–353. [Google Scholar] [CrossRef] [Green Version]
- Stamets, P. Growing Gourmet and Medicinal Mushrooms; Ten Speed Press: Berkeley, CA, USA, 2011. [Google Scholar]
- Sakamoto, Y. Influences of Environmental Factors on Fruiting Body Induction, Development and Maturation in Mushroom-Forming Fungi. Fungal Biol. Rev. 2018, 32, 236–248. [Google Scholar] [CrossRef]
- Elliot, C. Reproduction in Fungi. In Genetical and Physiological Aspects; Chapman Hall: London, UK, 1994. [Google Scholar]
- Moore, D. Fungal Morphogenesis; Cambridge University Press: Cambridge, MA, USA, 2003. [Google Scholar]
- Kumar, P.; Eid, E.M.; Al-Huqail, A.A.; Širić, I.; Adelodun, B.; Abou Fayssal, S.; Valadez-Blanco, R.; Goala, M.; Ajibade, F.O.; Choi, K.S.; et al. Kinetic Studies on Delignification and Heavy Metals Uptake by Shiitake (Lentinula edodes) Mushroom Cultivated on Agro-Industrial Wastes. Horticulturae 2022, 8, 316. [Google Scholar] [CrossRef]
- Uhart, M.; Piscera, J.M.; Albertó, E. Utilization of New Naturally Occurring Strains and Supplementation to Improve the Biological Efficiency of the Edible Mushroom Agrocybe cylindracea. J. Ind. Microbiol. Biotechnol. 2008, 35, 595–602. [Google Scholar] [CrossRef]
- Hennicke, F.; Cheikh-Ali, Z.; Liebisch, T.; Maciá-Vicente, J.G.; Bode, H.B.; Piepenbring, M. Distinguishing Commercially Grown Ganoderma lucidum from Ganoderma lingzhi from Europe and East Asia on the Basis of Morphology, Molecular Phylogeny, and Triterpenic Acid Profiles. Phytochemistry 2016, 127, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Esser, K.; Semerdžieva, M.; Stahl, U. Genetische Untersuchungen an dem Basidiomyceten Agrocybe aegerita. Theor. Appl. Genet. 1974, 45, 77–85. [Google Scholar] [CrossRef]
- Labarere, J.; Noel, T. Mating Type Switching in the Tetrapolar Basidiomycete Agrocybe aegerita. Genetics 1992, 131, 307–319. [Google Scholar] [CrossRef]
- Herzog, R.; Solovyeva, I.; Rühl, M.; Thines, M.; Hennicke, F. Dikaryotic Fruiting Body Development in a Single Dikaryon of Agrocybe aegerita and the Spectrum of Monokaryotic Fruiting Types in Its Monokaryotic Progeny. Mycol. Prog. 2016, 15, 947–957. [Google Scholar] [CrossRef]
- Orban, A.; Weber, A.; Herzog, R.; Hennicke, F.; Rühl, M. Transcriptome of Different Fruiting Stages in the Cultivated Mushroom Cyclocybe aegerita Suggests a Complex Regulation of Fruiting and Reveals Enzymes Putatively Involved in Fungal Oxylipin Biosynthesis. BMC Genom. 2021, 22, 324. [Google Scholar] [CrossRef] [PubMed]
- Elders, H.; Hennicke, F. The Pacific Tree-Parasitic Fungus Cyclocybe parasitica Exhibits Monokaryotic Fruiting, Showing Phenotypes Known from Bracket Fungi and from Cyclocybe aegerita. J. Fungi 2021, 7, 394. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Tang, M.; Chen, H. Growth Promotion of Suillus Luteus by Adenosine in Vitro. Mycoscience 2010, 51, 139–143. [Google Scholar] [CrossRef]
- Ruytinx, J.; Miyauchi, S.; Hartmann-Wittulsky, S.; de Freitas Pereira, M.; Guinet, F.; Churin, J.-L.; Put, C.; Le Tacon, F.; Veneault-Fourrey, C.; Martin, F.; et al. A Transcriptomic Atlas of the Ectomycorrhizal Fungus Laccaria bicolor. Microorganisms 2021, 9, 2612. [Google Scholar] [CrossRef]
- Frings, R.A.; Maciá-Vicente, J.G.; Buße, S.; Čmoková, A.; Kellner, H.; Hofrichter, M.; Hennicke, F. Multilocus Phylogeny- and Fruiting Feature-Assisted Delimitation of European Cyclocybe aegerita from a New Asian Species Complex and Related Species. Mycol. Prog. 2020, 19, 1001–1016. [Google Scholar] [CrossRef]
- Roy, J.; Stobo, I.; Shotton, S.M.; Ganderton, P.; Gillies, C.M. The Nutritive Value of Non-Milk Proteins for the Preruminant Calf. The Effect of Replacement of Milk Protein by Soya-Bean Flour or Fish-Protein Concentrate. Br. J. Nutr. 1977, 38, 167–187. [Google Scholar] [CrossRef] [Green Version]
- Dawson, D.; Morrill, J.; Reddy, P.; Minocha, H.; Ramsey, H. Soy Protein Concentrate and Heated Soy Flours as Protein Sources in Milk Replacer for Preruminant Calves. J. Dairy Sci. 1988, 71, 1301–1309. [Google Scholar] [CrossRef]
- Stochmal, A.; Piacente, S.; Pizza, C.; De Riccardis, F.; Leitz, R.; Oleszek, W. Alfalfa (Medicago sativa L.) Flavonoids. 1. Apigenin and Luteolin Glycosides from Aerial Parts. J. Agric. Food Chem. 2001, 49, 753–758. [Google Scholar] [CrossRef]
- Dons, J.; De Vries, O.; Wessels, J. Characterization of the Genome of the Basidiomycete Schizophyllum commune. Biochim. Biophys. Acta (BBA) 1979, 563, 100–112. [Google Scholar] [CrossRef]
- Herzog, R.; Solovyeva, I.; Bölker, M.; Lugones, L.G.; Hennicke, F. Exploring Molecular Tools for Transformation and Gene Expression in the Cultivated Edible Mushroom Agrocybe aegerita. Mol. Genet. Genom. 2019, 294, 663–677. [Google Scholar] [CrossRef]
- Bässler, C.; Heilmann-Clausen, J.; Karasch, P.; Brandl, R.; Halbwachs, H. Ectomycorrhizal Fungi Have Larger Fruit Bodies than Saprotrophic Fungi. Fungal Ecol. 2014, 17, 205–212. [Google Scholar] [CrossRef]
- Krah, F.-S.; Bässler, C.; Heibl, C.; Soghigian, J.; Schaefer, H.; Hibbett, D.S. Evolutionary Dynamics of Host Specialization in Wood-Decay Fungi. BMC Evol. Biol. 2018, 18, 119–132. [Google Scholar] [CrossRef] [Green Version]
- Ackerman, D.; Millet, D.B.; Chen, X. Global Estimates of Inorganic Nitrogen Deposition across Four Decades. Glob. Biogeochem. Cycles 2019, 33, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Clemmensen, K.E.; Bahr, A.; Ovaskainen, O.; Dahlberg, A.; Ekblad, A.; Wallander, H.; Stenlid, J.; Finlay, R.D.; Wardle, D.A.; Lindahl, B.D. Roots and Associated Fungi Drive Long-Term Carbon Sequestration in Boreal Forest. Science 2013, 340, 1615–1618. [Google Scholar] [CrossRef]
- Liu, S.; García-Palacios, P.; Tedersoo, L.; Guirado, E.; van der Heijden, M.G.; Wagg, C.; Chen, D.; Wang, Q.; Wang, J.; Singh, B.K.; et al. Phylotype Diversity within Soil Fungal Functional Groups Drives Ecosystem Stability. Nat. Ecol. Evol. 2022, 6, 900–909. [Google Scholar] [CrossRef]
- Boddy, L.; Jones, T.H. Chapter 9 Interactions between Basidiomycota and Invertebrates. In British Mycological Society Symposia Series: Ecology of Saprotrophic Basidiomycetes; Boddy, L., Frankland, J.C., van West, P., Eds.; Academic Press: Cambridge, UK, 2008; Volume 28, pp. 155–179. ISBN 0275-0287. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hennicke, F.; Fleckenstein, L.; Bässler, C.; Krah, F.-S. Organic Nitrogen Supplementation Increases Vegetative and Reproductive Biomass in a Versatile White Rot Fungus. J. Fungi 2023, 9, 7. https://doi.org/10.3390/jof9010007
Hennicke F, Fleckenstein L, Bässler C, Krah F-S. Organic Nitrogen Supplementation Increases Vegetative and Reproductive Biomass in a Versatile White Rot Fungus. Journal of Fungi. 2023; 9(1):7. https://doi.org/10.3390/jof9010007
Chicago/Turabian StyleHennicke, Florian, Lena Fleckenstein, Claus Bässler, and Franz-Sebastian Krah. 2023. "Organic Nitrogen Supplementation Increases Vegetative and Reproductive Biomass in a Versatile White Rot Fungus" Journal of Fungi 9, no. 1: 7. https://doi.org/10.3390/jof9010007