

Antifungal Activity of 6-Methylcoumarin against Valsa mali and Its Possible Mechanism of Action

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Pathogens and Cultures
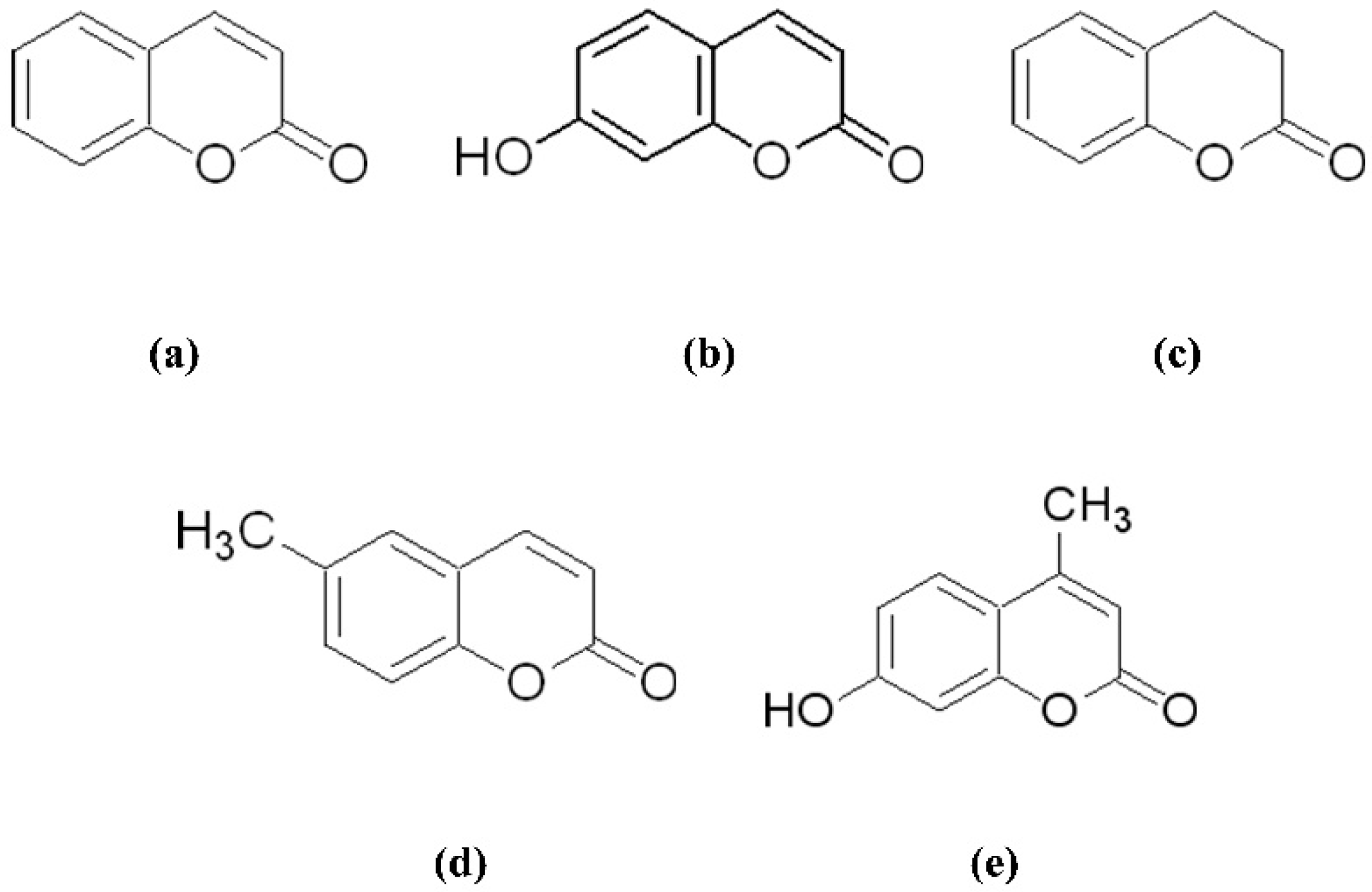
2.3. Antifungal Activity of Coumarins against Forest Pathogenic Fungi
2.4. Antifungal Activity of 6-MCM against V. mali
2.5. Effects of 6-MCM on the Cell Membrane of V. mali
2.6. The Effect of 6-MCM on V. mali Enzyme Activity
2.7. Scanning Electron Microscopy (SEM)
2.8. Transmission Electron Microscopy (TEM)
2.9. Control Effect
2.10. Statistical Analysis
3. Results
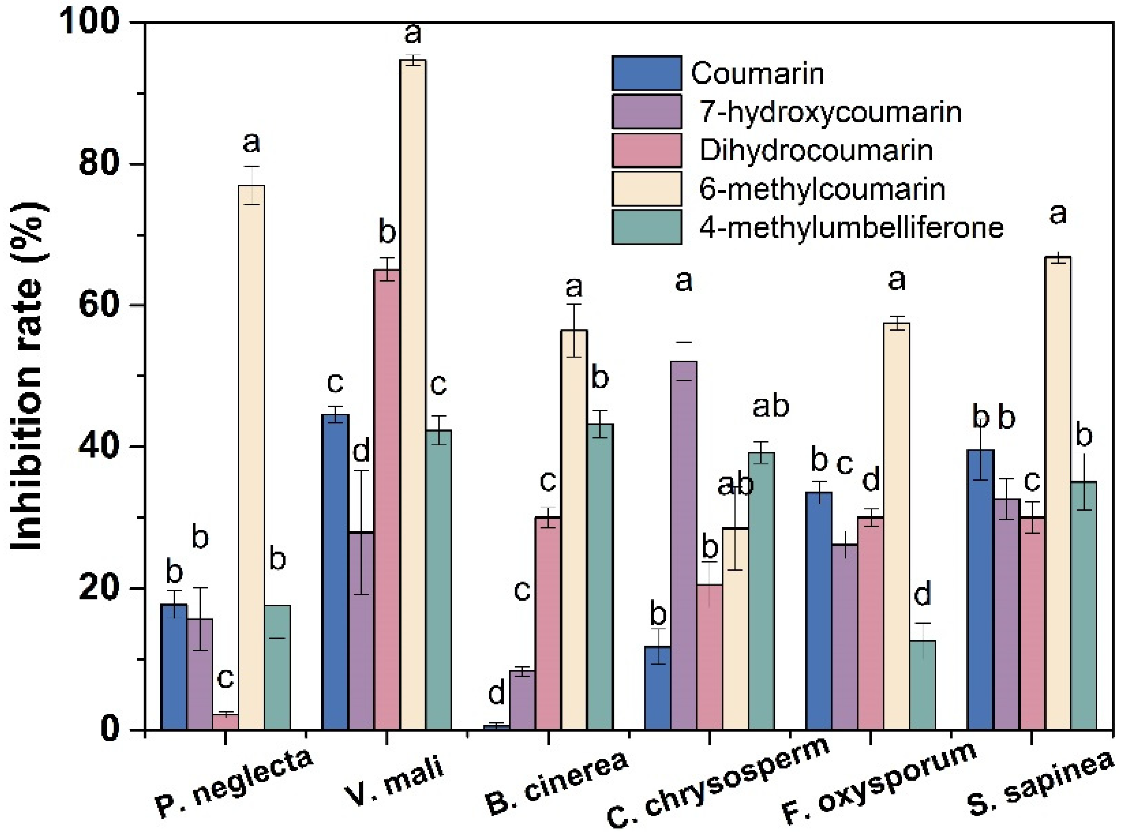
3.1. Antifungal Activity of Coumarin Compounds against Forest Pathogenic Fungi
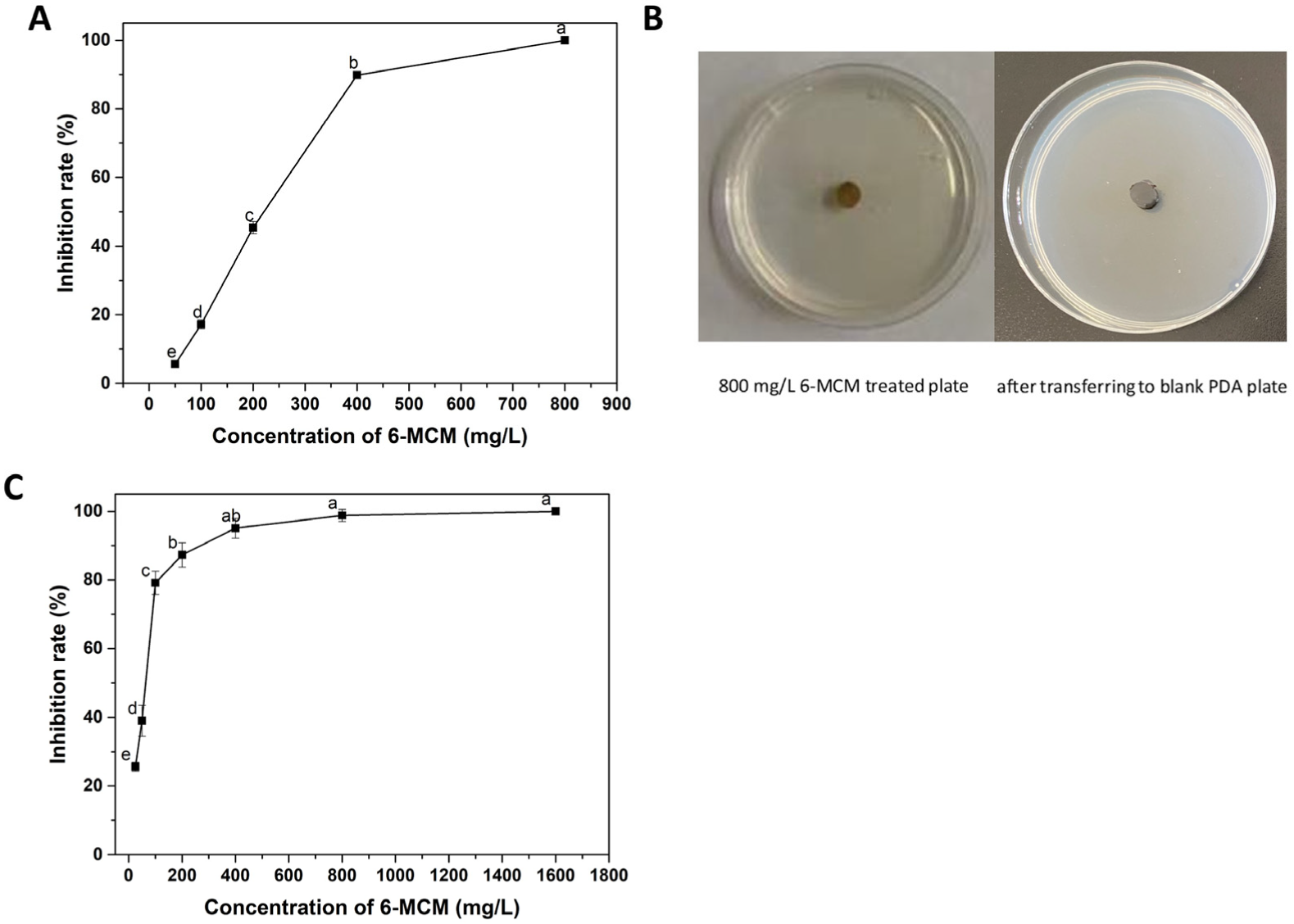
3.2. Virulence of 6-MCM against V. mali
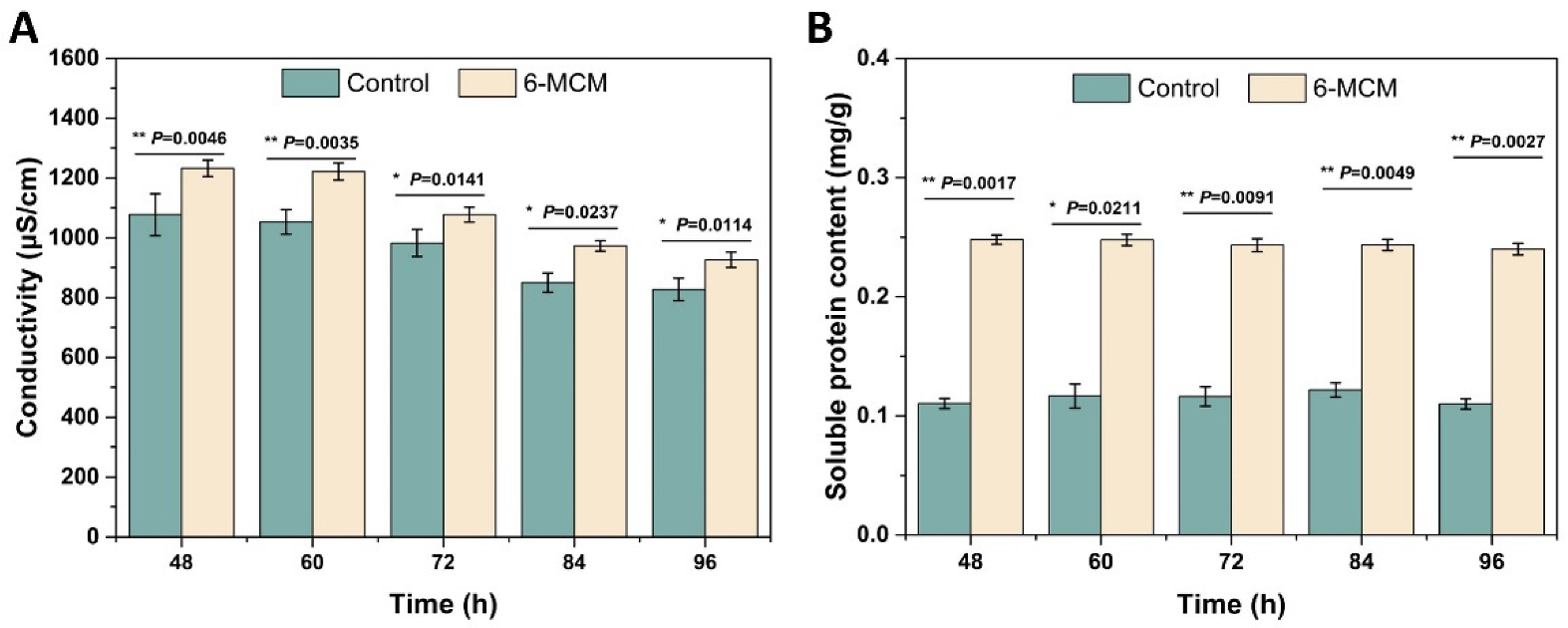
3.3. Effects of 6-MCM on the Cell Membrane of V. mali
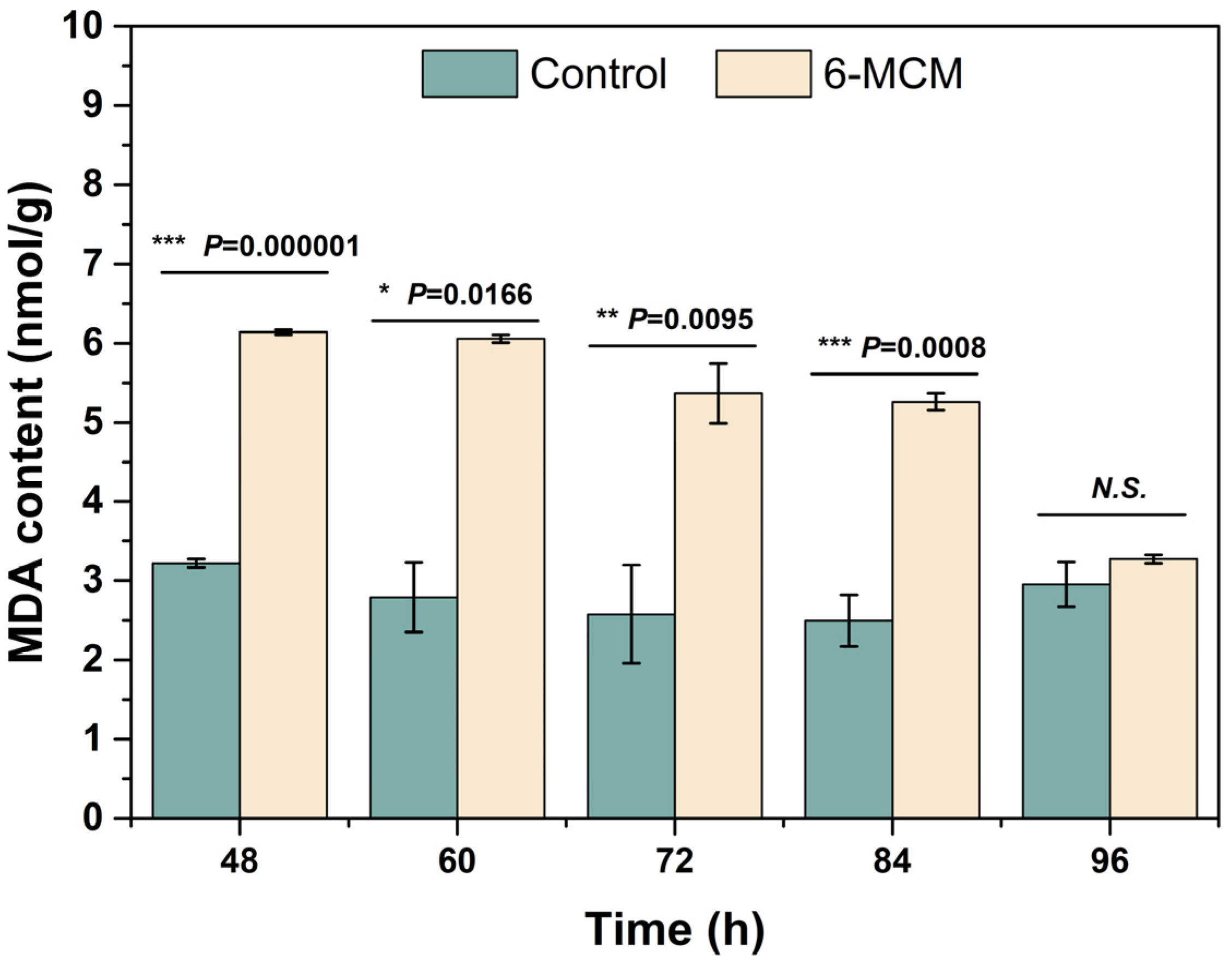
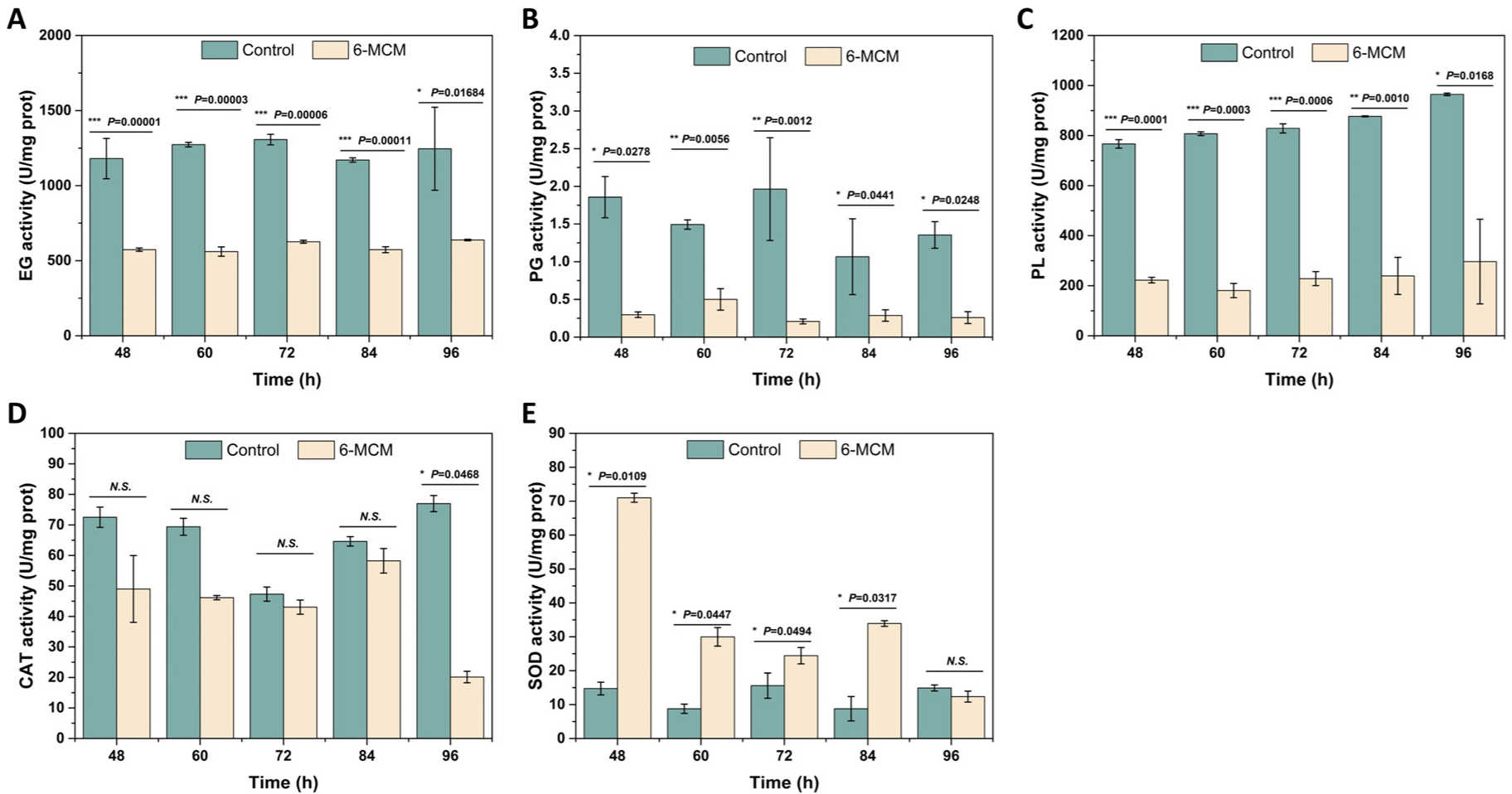
3.4. Effects of 6-MCM on Enzyme Activity of V. mali
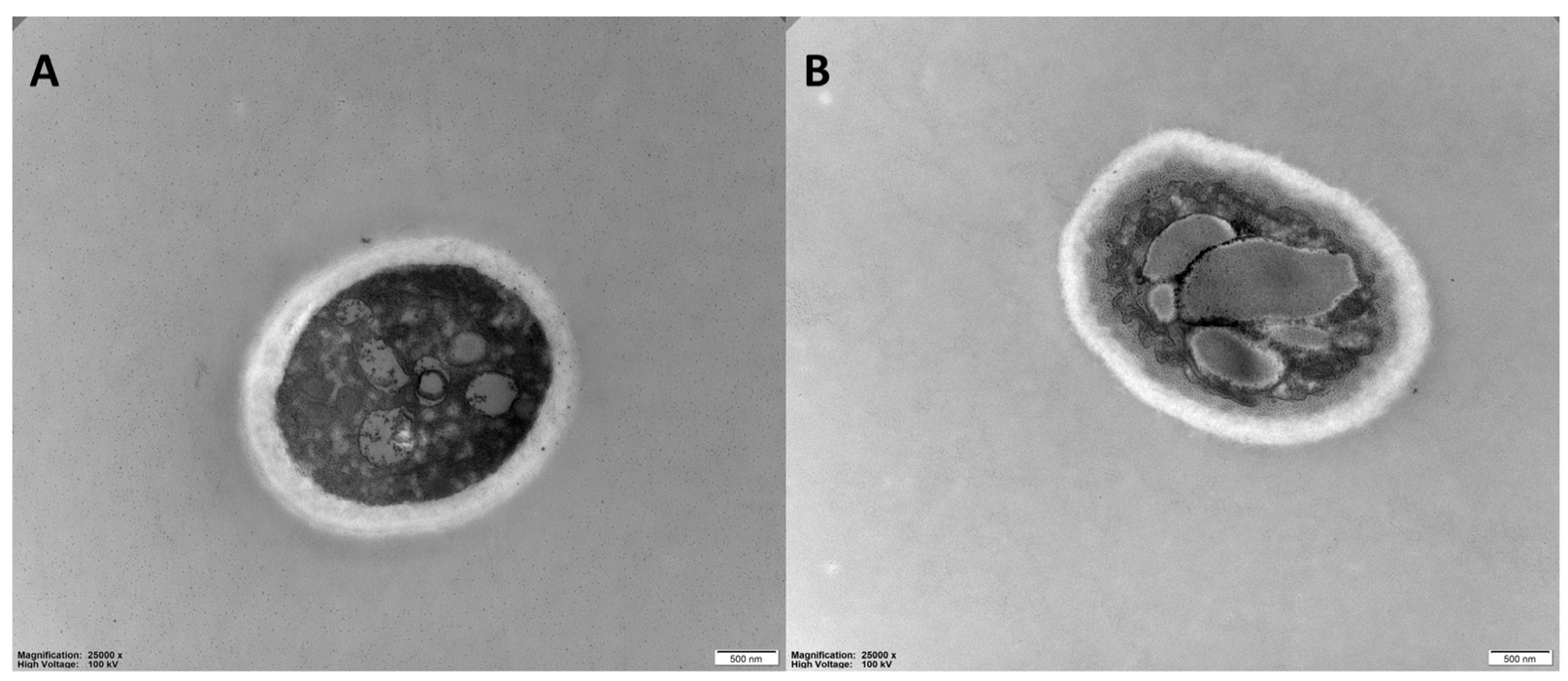
3.5. Effects of 6-MCM on Microscopic Morphology and Ultrastructure of V. mali Hyphae
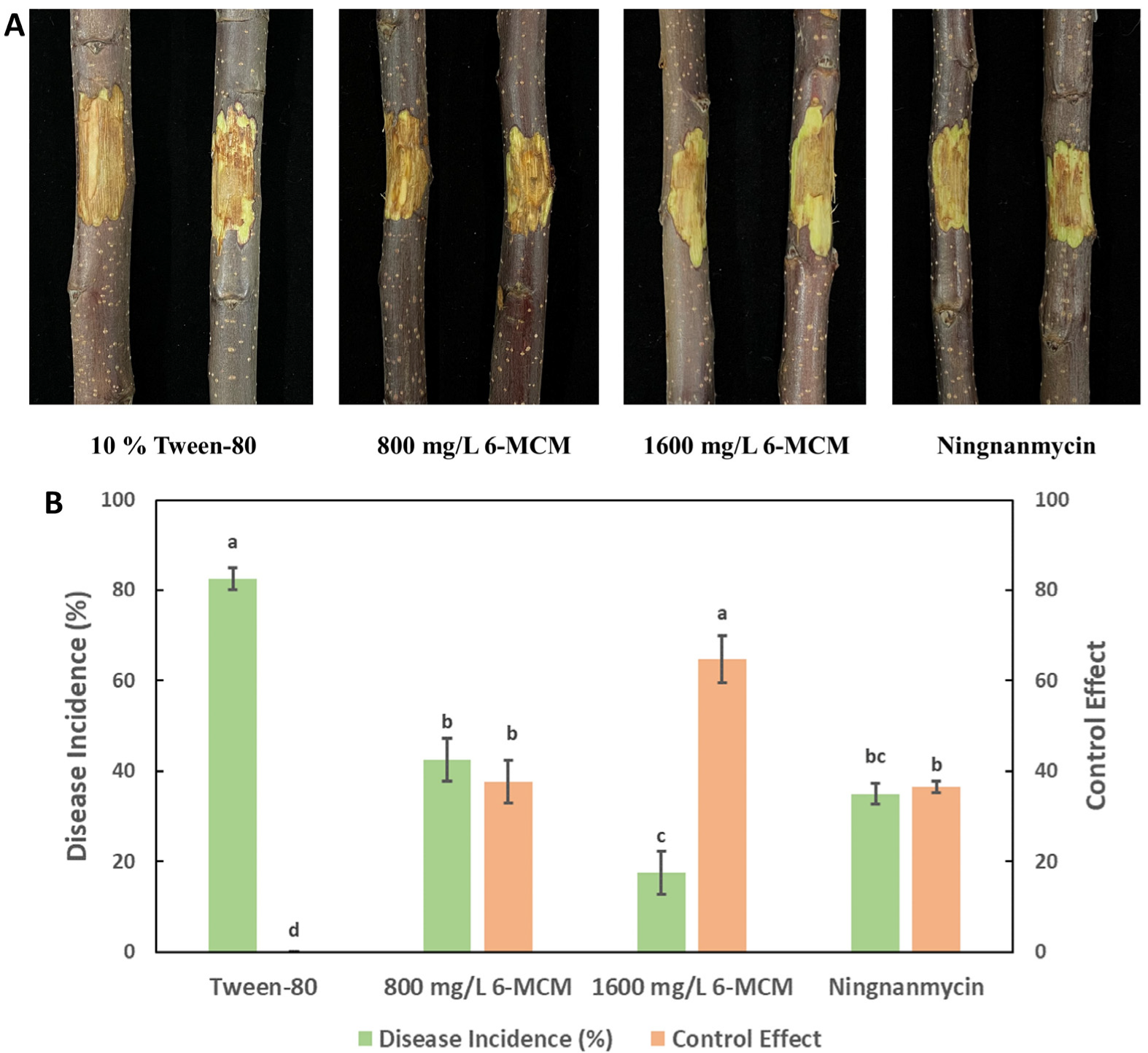
3.6. Control Effects
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, R.H.; Li, J.Y.; Zhang, F.R.; Zheng, D.A.; Chang, Y.L.; Xu, L.S.; Huang, L.L. Biocontrol activity of Bacillus velezensis D4 against apple Valsa canker. Biol. Control 2021, 163, 104760. [Google Scholar] [CrossRef]
- Bessho, H.; Tsuchiya, S.; Soejima, J. Screening methods of apple trees for resistance to Valsa canker. Euphytica 1994, 77, 18. [Google Scholar] [CrossRef]
- Meng, X.-L.; Qi, X.-H.; Han, Z.-Y.; Guo, Y.-B.; Wang, Y.-N.; Hu, T.-L.; Wang, L.-M.; Cao, K.-Q.; Wang, S.-T. Latent Infection of Valsa mali in the Seeds, Seedlings and Twigs of Crabapple and Apple Trees is a Potential Inoculum Source of Valsa Canker. Sci. Rep. 2019, 9, 7738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutton, T.B.; Aldwinckle, H.S.; Agnello, A.M.; Walgenbach, J.F. Compendium of Apple and Pear Diseases and Pests, 2nd ed.; The American Phytopathological Society: Saint Paul, MN, USA, 2014. [Google Scholar]
- Salazar, C.; Rand, J. Pesticide use, production risk and shocks. The case of rice producers in Vietnam. J. Environ. Manag. 2020, 253, 109705. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Ren, D.; Gao, C.; Li, J.; Du, B.; Wang, Z.; Qian, S. Recent advances for alkaloids as botanical pesticides for use in organic agriculture. Int. J. Pest Manag. 2021. [Google Scholar] [CrossRef]
- Souto, A.L.; Sylvestre, M.; Tolke, E.D.; Tavares, J.F.; Barbosa, J.M.; Cebrian-Torrejon, G. Plant-Derived Pesticides as an Alternative to Pest Management and Sustainable Agricultural Production: Prospects, Applications and Challenges. Molecules 2021, 26, 4835. [Google Scholar] [CrossRef]
- Song, P.P.; Zhao, J.; Liu, Z.L.; Duan, Y.B.; Hou, Y.P.; Zhao, C.Q.; Wu, M.; Wei, M.; Wang, N.H.; Lv, Y.; et al. Evaluation of antifungal activities and structure-activity relationships of coumarin derivatives. Pest Manag. Sci. 2017, 73, 94–101. [Google Scholar] [CrossRef]
- Prusty, J.S.; Kumar, A. Coumarins: Antifungal effectiveness and future therapeutic scope. Mol. Divers. 2020, 24, 1367–1383. [Google Scholar] [CrossRef]
- Navarro-Garcia, V.M.; Rojas, G.; Aviles, M.; Fuentes, M.; Zepeda, G. In vitro antifungal activity of coumarin extracted from Loeselia mexicana Brand. Mycoses 2011, 54, E569–E571. [Google Scholar] [CrossRef]
- Bhattarai, N.; Kumbhar, A.A.; Pokharel, Y.R.; Yadav, P.N. Anticancer Potential of Coumarin and its Derivatives. Mini-Rev. Med. Chem. 2021, 21, 2996–3029. [Google Scholar] [CrossRef]
- Cheke, R.S.; Patel, H.M.; Patil, V.M.; Ansari, I.A.; Ambhore, J.P.; Shinde, S.D.; Kadri, A.; Snoussi, M.; Adnan, M.; Kharkar, P.S.; et al. Molecular Insights into Coumarin Analogues as Antimicrobial Agents: Recent Developments in Drug Discovery. Antibiotics 2022, 11, 566. [Google Scholar] [CrossRef] [PubMed]
- Grover, J.; Jachak, S.M. Coumarins as privileged scaffold for anti-inflammatory drug development. RSC Adv. 2015, 5, 38892–38905. [Google Scholar] [CrossRef]
- Thuong, P.T.; Hung, T.M.; Ngoc, T.M.; Ha, D.T.; Min, B.S.; Kwack, S.J.; Kang, T.S.; Choi, J.S.; Bae, K. Antioxidant Activities of Coumarins from Korean Medicinal Plants and their Structure-Activity Relationships. Phytother. Res. 2010, 24, 101–106. [Google Scholar] [CrossRef] [PubMed]
- He, Y.H.; Shang, X.F.; Li, H.X.; Li, A.P.; Tang, C.; Zhang, B.Q.; Zhang, Z.J.; Wang, R.; Ma, Y.; Du, S.S.; et al. Antifungal Activity and Action Mechanism Study of Coumarins from Cnidium monnieri Fruit and Structurally Related Compounds. Chem. Biodivers. 2021, 18, e2100633. [Google Scholar] [CrossRef] [PubMed]
- Han, S.T.; Yang, L.; Wang, Y.; Ran, Y.A.; Li, S.L.; Ding, W. Preliminary Studies on the Antibacterial Mechanism of a New Plant-Derived Compound, 7-Methoxycoumarin, Against Ralstonia solanacearum. Front. Microbiol. 2021, 12, 697911. [Google Scholar] [CrossRef]
- Ali, E.M.; Alkuwayti, M.A.; Aldayel, M.F.; Abdallah, B.M. Coumarin derivative, 5′-hydroxy-auraptene, extracted from Lotus lalambensis, displays antifungal and anti-aflatoxigenic activities against Aspergillus flavus. J. King Saud Univ. Sci. 2021, 33, 101216. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Y.; He, X.B.; Xiao, Q.L.; Han, S.T.; Jia, Z.; Li, S.L.; Ding, W. Discovery of a novel plant-derived agent against Ralstonia solanacearum by targeting the bacterial division protein FtsZ. Pestic. Biochem. Physiol. 2021, 177, 104892. [Google Scholar] [CrossRef]
- Yang, J.; Lin, L.N.; Zhang, X.B.; Wu, Y.; Bi, B.; Zhang, G.C.; Zou, C.S. Sodium pheophorbide a has photoactivated fungicidal activity against Pestalotiopsis neglecta. Pestic. Biochem. Physiol. 2019, 158, 25–31. [Google Scholar]
- Yang, J.; Chen, Y.Z.; Tao, L.; Zhang, Y.D.; Wang, S.R.; Zhang, G.C.; Zhang, J. Inhibitory effects and mechanisms of vanillin on gray mold and black rot of cherry tomatoes. Pestic. Biochem. Physiol. 2021, 175, 104859. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ji, D.C.; Chen, T.; Ma, D.Y.; Liu, J.L.; Xu, Y.; Tian, S.P. Inhibitory effects of methyl thujate on mycelial growth of Botrytis cinerea and possible mechanisms. Postharvest Biol. Technol. 2018, 142, 46–54. [Google Scholar] [CrossRef]
- Gao, Y.; Xu, R.; Gu, S.; Chen, K.; Li, J.; He, X.; Shang, S.; Song, Z.; Song, J. Discovery of Natural Rosin Derivatives Containing Oxime Ester Moieties as Potential Antifungal Agents to Control Tomato Gray Mold Caused by Botrytis cinerea. J. Agric. Food Chem. 2022, 70, 5551–5560. [Google Scholar] [CrossRef] [PubMed]
- Percival, G.C.; Graham, S. The potential of resistance inducers and synthetic fungicide combinations for management of foliar diseases of nursery stock. Crop Prot. 2021, 145, 105636. [Google Scholar] [CrossRef]
- Nasab, N.H.; Azimian, F.; Kruger, H.G.; Kim, S.J. 3-Bromoacetylcoumarin, a Crucial Key for Facial Synthesis of Biological Active Compounds. Chemistryselect 2022, 7, 1734. [Google Scholar]
- Schiavi, D.; Francesconi, S.; Bischetti, G.; Giovanale, G.; Fortunati, E.; Balestra, G.M. Antibacterial activity of coumarin as an innovative organic control strategy for Xanthomonas euvesicatoria pv. euvesicatoria. J. Plant Dis. Prot. 2022, 129, 181–187. [Google Scholar] [CrossRef]
- Xu, L.; Yu, J.M.; Jin, L.; Pan, L. Design, Synthesis, and Antifungal Activity of 4-Amino Coumarin Based Derivatives. Molecules 2022, 27, 2738. [Google Scholar] [CrossRef]
- Wei, Y.; Peng, W.; Wang, D.; Hao, S.H.; Li, W.W.; Ding, F. Design, synthesis, antifungal activity, and 3D-QSAR of coumarin derivatives. J. Pestic. Sci. 2018, 43, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Ding, W.; Xu, Y.; Wu, D.; Li, S.; Chen, J.; Guo, B. New Insights into the Antibacterial Activity of Hydroxycoumarins against Ralstonia solanacearum. Molecules 2016, 21, 468. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, Y.G.; Cho, H.S.; Ryu, S.Y.; Cho, M.H.; Lee, J. Coumarins reduce biofilm formation and the virulence of Escherichia coli O157:H7. Phytomedicine 2014, 21, 1037–1042. [Google Scholar] [CrossRef]
- Yang, L.; Wu, L.T.; Yao, X.Y.; Zhao, S.Y.; Wang, J.; Li, S.L.; Ding, W. Hydroxycoumarins: New, effective plant-derived compounds reduce Ralstonia pseudosolanacearum populations and control tobacco bacterial wilt. Microbiol. Res. 2018, 215, 15–21. [Google Scholar] [CrossRef]
- Wang, B.; Li, P.R.; Xu, S.; Liu, L.Y.; Xu, Y.N.; Feng, X.; Zhao, X.Z.; Chen, Y. Inhibitory Effects of the Natural Product Esculetin on Phytophthora capsici and Its Possible Mechanism. Plant Dis. 2021, 105, 1814–1822. [Google Scholar] [CrossRef] [PubMed]
- Lionetti, V.; Metraux, J.P. Plant cell wall in pathogenesis, parasitism and symbiosis. Front. Plant Sci. 2014, 5, 612. [Google Scholar] [CrossRef] [PubMed]
- Lagaert, S.; Belien, T.; Volckaert, G. Plant cell walls: Protecting the barrier from degradation by microbial enzymes. Semin. Cell Dev. Biol. 2009, 20, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Kubicek, C.P.; Starr, T.L.; Glass, N.L. Plant Cell Wall-Degrading Enzymes and Their Secretion in Plant-Pathogenic Fungi. Annu. Rev. Phytopathol. 2014, 52, 427–451. [Google Scholar] [CrossRef]
- Huang, Y.; Yu, C.L.; Sun, C.C.; Saleem, M.; Li, P.L.; Li, B.H.; Wang, C.X. β-Glucosidase VmGlu2 Contributes to the Virulence of Valsa mali in Apple Tree. Front. Microbiol. 2021, 12, 695112. [Google Scholar] [CrossRef]
- Yu, C.L.; Li, T.; Shi, X.P.; Saleem, M.; Li, B.H.; Liang, W.X.; Wang, C.X. Deletion of Endo-β-1,4-Xylanase VmXyl1 Impacts the Virulence of Valsa mali in Apple Tree. Front. Plant Sci. 2018, 9, 663. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.Y.; Liu, H.Q.; Li, Z.P.; Ke, X.W.; Dou, D.L.; Gao, X.N.; Song, N.; Dai, Q.Q.; Wu, Y.X.; Xu, J.R.; et al. Genome sequence of Valsa canker pathogens uncovers a potential adaptation of colonization of woody bark. New Phytol. 2015, 208, 1202–1216. [Google Scholar] [CrossRef]
- Ke, X.W.; Yin, Z.Y.; Song, N.; Dai, Q.Q.; Voegele, R.T.; Liu, Y.Y.; Wang, H.Y.; Gao, X.N.; Kang, Z.S.; Huang, L.L. Transcriptome profiling to identify genes involved in pathogenicity of Valsa mali on apple tree. Fungal Genet. Biol. 2014, 68, 31–38. [Google Scholar] [CrossRef]
- Wu, Y.X.; Xu, L.S.; Yin, Z.Y.; Dai, Q.Q.; Gao, X.N.; Feng, H.; Voegele, R.T.; Huang, L.L. Two members of the velvet family, VmVeA and VmVelB, affect conidiation, virulence and pectinase expression in Valsa mali. Mol. Plant Pathol. 2018, 19, 1639–1651. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.X.; Xu, L.S.; Yin, Z.Y.; Feng, H.; Huang, L.L. Transcription factor VmSeb1 is required for the growth, development, and virulence in Valsa mali. Microb. Pathog. 2018, 123, 132–138. [Google Scholar] [CrossRef]
- Xu, M.; Gao, X.N.; Chen, J.L.; Yin, Z.Y.; Feng, H.; Huang, L.L. The feruloyl esterase genes are required for full pathogenicity of the apple tree canker pathogen Valsa mali. Mol. Plant Pathol. 2018, 19, 1353–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Z.Y.; Ke, X.W.; Huang, D.X.; Gao, X.N.; Voegele, R.T.; Kang, Z.S.; Huang, L.L. Validation of reference genes for gene expression analysis in Valsa mali var. mali using real-time quantitative PCR. World J. Microbiol. Biotechnol. 2013, 29, 1563–1571. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Stringlis, I.A.; van Bentum, S.; de Jonge, R.; Snoek, B.L.; Pieterse, C.M.J.; Bakker, P.; Berendsen, R.L. Transcriptome Signatures in Pseudomonas simiae WCS417 Shed Light on Role of Root-Secreted Coumarins in Arabidopsis-Mutualist Communication. Microorganisms 2021, 9, 575. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virulence Regression Equation | χ2 | R2 | EC50 (mg/L) | 95% Confidence Interval | EC90 (mg/L) | 95% Confidence Interval | |
|---|---|---|---|---|---|---|---|
| Mycelial growth inhibition | Y = 3.406X − 7.726 | 8.995 | 0.968 | 185.49 | 136.849–253.643 | 441.16 | 309.576–878.967 |
| Spore germination inhibition | Y = 2.059X − 3.578 | 6.812 | 0.979 | 54.62 | 46.288–189.456 | 228.892 | 189.456–289.685 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-Z.; Wang, S.-R.; Li, T.; Zhang, G.-C.; Yang, J. Antifungal Activity of 6-Methylcoumarin against Valsa mali and Its Possible Mechanism of Action. J. Fungi 2023, 9, 5. https://doi.org/10.3390/jof9010005
Chen Y-Z, Wang S-R, Li T, Zhang G-C, Yang J. Antifungal Activity of 6-Methylcoumarin against Valsa mali and Its Possible Mechanism of Action. Journal of Fungi. 2023; 9(1):5. https://doi.org/10.3390/jof9010005
Chicago/Turabian StyleChen, Yun-Ze, Shu-Ren Wang, Tao Li, Guo-Cai Zhang, and Jing Yang. 2023. "Antifungal Activity of 6-Methylcoumarin against Valsa mali and Its Possible Mechanism of Action" Journal of Fungi 9, no. 1: 5. https://doi.org/10.3390/jof9010005