
Whole Genome Sequencing Shows Genetic Diversity, as Well as Clonal Complex and Gene Polymorphisms Associated with Fluconazole Non-Susceptible Isolates of Candida tropicalis

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolates and Susceptibility Testing
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Whole Genome Sequencing Analysis
2.3.1. MLST
2.3.2. Phylogenetic Analysis
2.3.3. Assessment of Antifungal Drug Resistance Markers
3. Results
3.1. Phenotypic Susceptibility to Antifungal Agents
3.2. Multilocus Sequence Typing (MLST)
3.3. Phylogenetic Analysis
3.4. Gene Mutations and Antifungal Resistance
3.4.1. Azole Resistance
3.4.2. Echinocandin Resistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ko, J.H.; Jung, D.S.; Lee, J.Y.; Kim, H.A.; Ryu, S.Y.; Jung, S.I.; Joo, E.J.; Cheon, S.; Kim, Y.S.; Kim, S.W.; et al. Poor prognosis of candida tropicalis among non-albicans candidemia: A retrospective multicenter cohort study, korea. Diagn. Microbiol. Infect. Dis. 2019, 95, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.L.; Huang, Y.T.; Hsieh, M.H.; Hii, I.M.; Lee, Y.L.; Ho, M.W.; Liu, C.E.; Chen, Y.H.; Wang, F.D. Clinical characteristics of candida tropicalis fungaemia with reduced triazole susceptibility in taiwan: A multicentre study. Int. J. Antimicrob. Agents 2019, 53, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Ruiz, M.; Puig-Asensio, M.; Guinea, J.; Almirante, B.; Padilla, B.; Almela, M.; Díaz-Martín, A.; Rodríguez-Baño, J.; Cuenca-Estrella, M.; Aguado, J.M. Candida tropicalis bloodstream infection: Incidence, risk factors and outcome in a population-based surveillance. J. Infect. 2015, 71, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Chapman, B.; Slavin, M.; Marriott, D.; Halliday, C.; Kidd, S.; Arthur, I.; Bak, N.; Heath, C.H.; Kennedy, K.; Morrissey, C.O.; et al. Changing epidemiology of candidaemia in australia. J. Antimicrob. Chemother. 2017, 72, 1103–1108. [Google Scholar] [CrossRef] [Green Version]
- Keighley, C.; Chen, S.C.; Marriott, D.; Pope, A.; Chapman, B.; Kennedy, K.; Bak, N.; Underwood, N.; Wilson, H.L.; McDonald, K.; et al. Candidaemia and a risk predictive model for overall mortality: A prospective multicentre study. BMC Infect. Dis. 2019, 19, 445. [Google Scholar] [CrossRef]
- Keighley, C.L.; Pope, A.; Marriott, D.J.E.; Chapman, B.; Bak, N.; Daveson, K.; Hajkowicz, K.; Halliday, C.; Kennedy, K.; Kidd, S.; et al. Risk factors for candidaemia: A prospective multi-centre case-control study. Mycoses 2020, 64, 257–263. [Google Scholar] [CrossRef]
- Chakrabarti, A.; Sood, P.; Rudramurthy, S.M.; Chen, S.; Kaur, H.; Capoor, M.; Chhina, D.; Rao, R.; Eshwara, V.K.; Xess, I.; et al. Incidence, characteristics and outcome of icu-acquired candidemia in india. Intensive Care Med. 2015, 41, 285–295. [Google Scholar] [CrossRef]
- Chai, L.Y.; Denning, D.W.; Warn, P. Candida tropicalis in human disease. Crit. Rev. Microbiol. 2010, 36, 282–298. [Google Scholar] [CrossRef]
- Tan, B.H.; Chakrabarti, A.; Li, R.Y.; Patel, A.K.; Watcharananan, S.P.; Liu, Z.; Chindamporn, A.; Tan, A.L.; Sun, P.L.; Wu, U.I.; et al. Incidence and species distribution of candidaemia in asia: A laboratory-based surveillance study. Clin. Microbiol. Infect. 2015, 21, 946–953. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, M.A.; Diekema, D.J.; Gibbs, D.L.; Newell, V.A.; Ellis, D.; Tullio, V.; Rodloff, A.; Fu, W.; Ling, T.A.; Global Antifungal Surveillance Group. Results from the artemis disk global antifungal surveillance study, 1997 to 2007: A 10.5-year analysis of susceptibilities of candida species to fluconazole and voriconazole as determined by clsi standardized disk diffusion. J. Clin. Microbiol. 2010, 48, 1366–1377. [Google Scholar] [CrossRef] [Green Version]
- Tulyaprawat, O.; Pharkjaksu, S.; Chongtrakool, P.; Ngamskulrungroj, P. An association of an eburst group with triazole resistance of candida tropicalis blood isolates. Front. Microbiol. 2020, 11, 934. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, X.; Wang, H.; Kudinha, T.; Mei, Y.-N.; Ni, F.; Pan, Y.-H.; Gao, L.-M.; Xu, H.; Kong, H.-S.; et al. Continual decline in azole susceptibility rates in candida tropicalis over a 9-year period in china. Front. Microbiol. 2021, 12, 1808. [Google Scholar]
- Yang, Y.L.; Lin, C.C.; Chang, T.P.; Lauderdale, T.L.; Chen, H.T.; Lee, C.F.; Hsieh, C.W.; Chen, P.C.; Lo, H.J. Comparison of human and soil candida tropicalis isolates with reduced susceptibility to fluconazole. PLoS ONE 2012, 7, e34609. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Slavin, M.; Nguyen, Q.; Marriott, D.; Playford, E.G.; Ellis, D.; Sorrell, T.; Australian Candidemia, S. Active surveillance for candidemia, australia. Emerg. Infect. Dis. 2006, 12, 1508–1516. [Google Scholar] [CrossRef]
- Oliveira, J.S.; Pereira, V.S.; Castelo-Branco, D.; Cordeiro, R.A.; Sidrim, J.J.C.; Brilhante, R.S.N.; Rocha, M.F.G. The yeast, the antifungal, and the wardrobe: A journey into antifungal resistance mechanisms of candida tropicalis. Can. J. Microbiol. 2020, 66, 377–388. [Google Scholar] [CrossRef]
- Zuza-Alves, D.L.; Silva-Rocha, W.P.; Chaves, G.M. An update on candida tropicalis based on basic and clinical approaches. Front. Microbiol. 2017, 8, 1927. [Google Scholar] [CrossRef] [Green Version]
- Chew, K.L.; Cheng, J.W.S.; Jureen, R.; Lin, R.T.P.; Teo, J.W.P. Erg11 mutations are associated with high-level azole resistance in clinical candida tropicalis isolates, a singapore study. Mycoscience 2017, 58, 111–115. [Google Scholar] [CrossRef]
- Ford, C.B.; Funt, J.M.; Abbey, D.; Issi, L.; Guiducci, C.; Martinez, D.A.; Delorey, T.; Li, B.Y.; White, T.C.; Cuomo, C.; et al. The evolution of drug resistance in clinical isolates of candida albicans. Elife 2015, 4, e00662. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Cao, Z.; Wang, Q.; Wang, Y.; Wang, X.; Chen, H.; Wang, H. Mdr1 overexpression combined with erg11 mutations induce high-level fluconazole resistance in candida tropicalis clinical isolates. BMC Infect. Dis. 2018, 18, 162. [Google Scholar] [CrossRef] [Green Version]
- Morio, F.; Jensen, R.H.; Le Pape, P.; Arendrup, M.C. Molecular basis of antifungal drug resistance in yeasts. Int. J. Antimicrob. Agents 2017, 50, 599–606. [Google Scholar] [CrossRef]
- Flowers, S.A.; Colón, B.; Whaley, S.G.; Schuler, M.A.; Rogers, P.D. Contribution of clinically derived mutations in erg11 to azole resistance in candida albicans. Antimicrob. Agents Chemother. 2015, 59, 450–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.C.; Cardozo Bonfim Carbone, D.; Diniz, P.F.; Freitas Fernandes, F.; Fuzo, C.A.; Santos Pereira Cardoso Trindade, C.; Rodrigues Chang, M.; Silva, J.S. Modulation of erg genes expression in clinical isolates of candida tropicalis susceptible and resistant to fluconazole and itraconazole. Mycopathologia 2020, 185, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.J.; Won, E.J.; Shin, J.H.; Kim, S.H.; Lee, W.G.; Kim, M.N.; Lee, K.; Shin, M.G.; Suh, S.P.; Ryang, D.W.; et al. Resistance mechanisms and clinical features of fluconazole-nonsusceptible candida tropicalis isolates compared with fluconazole-less-susceptible isolates. Antimicrob. Agents Chemother. 2016, 60, 3653–3661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.; Xiao, M.; Zhang, D.; Huang, J.J.; Wang, H.; Hou, X.; Zhang, L.; Kong, F.; Chen, S.C.; Tong, Z.H.; et al. Molecular mechanisms of azole resistance in candida tropicalis isolates causing invasive candidiasis in china. Clin. Microbiol. Infect. 2019, 25, 885–891. [Google Scholar] [CrossRef]
- Todd, R.T.; Selmecki, A. Expandable and reversible copy number amplification drives rapid adaptation to antifungal drugs. eLife 2020, 9, e58349. [Google Scholar] [CrossRef]
- Beekman, C.N.; Ene, I.V. Short-term evolution strategies for host adaptation and drug escape in human fungal pathogens. PLoS Pathog. 2020, 16, e1008519. [Google Scholar] [CrossRef]
- Berkow, E.L.; Lockhart, S.R. Fluconazole resistance in candida species: A current perspective. Infect. Drug Resist. 2017, 10, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Berman, J.; Krysan, D.J. Drug resistance and tolerance in fungi. Nat. Rev. Microbiol. 2020, 18, 319–331. [Google Scholar] [CrossRef]
- McTaggart, L.R.; Cabrera, A.; Cronin, K.; Kus, J.V. Antifungal susceptibility of clinical yeast isolates from a large canadian reference laboratory and application of whole genome sequence analysis to elucidate mechanisms of acquired resistance. Antimicrob. Agents Chemother. 2020, 64, e00402-20. [Google Scholar] [CrossRef]
- Magri, M.M.; Gomes-Gouvea, M.S.; de Freitas, V.L.; Motta, A.L.; Moretti, M.L.; Shikanai-Yasuda, M.A. Multilocus sequence typing of candida tropicalis shows the presence of different clonal clusters and fluconazole susceptibility profiles in sequential isolates from candidemia patients in sao paulo, brazil. J. Clin. Microbiol. 2013, 51, 268–277. [Google Scholar] [CrossRef] [Green Version]
- Corzo-Leon, D.E.; Alvarado-Matute, T.; Colombo, A.L.; Cornejo-Juarez, P.; Cortes, J.; Echevarria, J.I.; Guzman-Blanco, M.; Macias, A.E.; Nucci, M.; Ostrosky-Zeichner, L.; et al. Surveillance of candida spp bloodstream infections: Epidemiological trends and risk factors of death in two mexican tertiary care hospitals. PLoS ONE 2014, 9, e97325. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.Y.; Guo, H.; Wang, H.M.; Yi, G.H.; Zhou, L.M.; He, X.W.; Zhang, Y.; Xu, J. Multilocus sequence analyses reveal extensive diversity and multiple origins of fluconazole resistance in candida tropicalis from tropical china. Sci. Rep. 2017, 7, 42537. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, C.; Liu, J.Y.; Li, W.J.; Zhao, Y.; Xiang, M.J. Multilocus sequence typing of candida tropicalis shows clonal cluster enrichment in azole-resistant isolates from patients in shanghai, china. Infect. Genet. Evol. 2016, 44, 418–424. [Google Scholar] [CrossRef]
- Chen, P.Y.; Chuang, Y.C.; Wu, U.I.; Sun, H.Y.; Wang, J.T.; Sheng, W.H.; Lo, H.J.; Wang, H.Y.; Chen, Y.C.; Chang, S.C. Clonality of fluconazole-nonsusceptible candida tropicalis in bloodstream infections, taiwan, 2011-2017. Emerg. Infect. Dis. 2019, 25, 1660–1667. [Google Scholar] [CrossRef] [Green Version]
- Chou, H.H.; Lo, H.J.; Chen, K.W.; Liao, M.H.; Li, S.Y. Multilocus sequence typing of candida tropicalis shows clonal cluster enriched in isolates with resistance or trailing growth of fluconazole. Diagn. Microbiol. Infect. Dis. 2007, 58, 427–433. [Google Scholar] [CrossRef]
- Wang, Q.Y.; Tang, D.L.; Tang, K.W.; Guo, J.; Huang, Y.; Li, C.R. Multilocus sequence typing reveals clonality of fluconazole-nonsusceptible candida tropicalis: A study from wuhan to the global. Front. Microbiol. 2020, 11, 554249. [Google Scholar] [CrossRef]
- Chew, K.L.; Octavia, S.; Lin, R.T.P.; Yan, G.Z.; Teo, J.W.P. Delay in effective therapy in anidulafungin-resistant candida tropicalis fungaemia: Potential for rapid prediction of antifungal resistance with whole-genome-sequencing. J. Glob. Antimicrob. Resist. 2019, 16, 105–107. [Google Scholar] [CrossRef]
- O’Brien, C.E.; Oliveira-Pacheco, J.; Ó Cinnéide, E.; Haase, M.A.B.; Hittinger, C.T.; Rogers, T.R.; Zaragoza, O.; Bond, U.; Butler, G. Population genomics of the pathogenic yeast candida tropicalis identifies hybrid isolates in environmental samples. PLoS Pathog. 2021, 17, e1009138. [Google Scholar] [CrossRef]
- Biswas, C.; Marcelino, V.R.; Van Hal, S.; Halliday, C.; Martinez, E.; Wang, Q.; Kidd, S.; Kennedy, K.; Marriott, D.; Morrissey, C.O.; et al. Whole genome sequencing of australian candida glabrata isolates reveals genetic diversity and novel sequence types. Front. Microbiol. 2018, 9, 2946. [Google Scholar] [CrossRef]
- Biswas, C.; Wang, Q.; van Hal, S.J.; Eyre, D.W.; Hudson, B.; Halliday, C.L.; Mazsewska, K.; Kizny Gordon, A.; Lee, A.; Irinyi, L.; et al. Genetic heterogeneity of australian candida auris isolates: Insights from a non-outbreak setting using whole-genome sequencing. Open Forum Infect. Dis. 2020, 7, ofaa158. [Google Scholar] [CrossRef]
- CLSI. Epidemiological cutoff values for antifungal susceptibility testing. In clsi Supplement m59, 3rd ed.; Clinical Laboratory Standards Institute: Wayne, PA, USA, 2020; Volume 40, pp. 1–34. [Google Scholar]
- CLSI. Performance standards for antifungal susceptibility testing of yeasts. 2nd ed. In Clsi Supplement m60, 3rd ed.; Clinical Laboratory Standards Institute: Wayne, PA, USA, 2020; Volume 40, pp. 1–34. [Google Scholar]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, snpeff: Snps in the genome of drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Tavanti, A.; Davidson, A.D.; Johnson, E.M.; Maiden, M.C.; Shaw, D.J.; Gow, N.A.; Odds, F.C. Multilocus sequence typing for differentiation of strains of candida tropicalis. J. Clin. Microbiol. 2005, 43, 5593–5600. [Google Scholar] [CrossRef] [Green Version]
- Abyzov, A.; Urban, A.E.; Snyder, M.; Gerstein, M. Cnvnator: An approach to discover, genotype, and characterize typical and atypical cnvs from family and population genome sequencing. Genome Res. 2011, 21, 974–984. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Dong, D.; Yu, B.; Cai, G.; Wang, X.; Ji, Y.; Peng, Y. Mechanisms of azole resistance in 52 clinical isolates of candida tropicalis in china. J. Antimicrob. Chemother. 2013, 68, 778–785. [Google Scholar] [CrossRef]
- Tan, J.; Zhang, J.; Chen, W.; Sun, Y.; Wan, Z.; Li, R.; Liu, W. The a395t mutation in erg11 gene confers fluconazole resistance in candida tropicalis causing candidemia. Mycopathologia 2015, 179, 213–218. [Google Scholar] [CrossRef]
- Lo, H.J.; Tsai, S.H.; Chu, W.L.; Chen, Y.Z.; Zhou, Z.L.; Chen, H.F.; Lee, C.F.; Yang, Y.L. Fruits as the vehicle of drug resistant pathogenic yeasts. J. Infect. 2017, 75, 254–262. [Google Scholar] [CrossRef]
- Arastehfar, A.; Polat, S.H.; Daneshnia, F.; Hafez, A.; Salehi, M.; Polat, F.; Yasar, M.; Arslan, N.; Hosbul, T.; Unal, N.; et al. Recent increase in the prevalence of fluconazole-non-susceptible candida tropicalis blood isolates in turkey: Clinical implication of azole-non-susceptible and fluconazole tolerant phenotypes and genotyping. Front. Microbiol. 2020, 11, 587278. [Google Scholar] [CrossRef]
- Castanheira, M.; Deshpande, L.M.; Messer, S.A.; Rhomberg, P.R.; Pfaller, M.A. Analysis of global antifungal surveillance results reveals predominance of erg11 y132f alteration among azole-resistant candida parapsilosis and candida tropicalis and country-specific isolate dissemination. Int. J. Antimicrob. Agents 2020, 55, 105799. [Google Scholar] [CrossRef]
- Sitterle, E.; Coste, A.T.; Obadia, T.; Maufrais, C.; Chauvel, M.; Sertour, N.; Sanglard, D.; Puel, A.; D’Enfert, C.; Bougnoux, M.E. Large-scale genome mining allows identification of neutral polymorphisms and novel resistance mutations in genes involved in candida albicans resistance to azoles and echinocandins. J. Antimicrob. Chemother. 2020, 75, 835–848. [Google Scholar] [CrossRef]
- Gonçalves, S.S.; Souza, A.C.R.; Chowdhary, A.; Meis, J.F.; Colombo, A.L. Epidemiology and molecular mechanisms of antifungal resistance in candida and aspergillus. Mycoses 2016, 59, 198–219. [Google Scholar] [CrossRef]
- Selmecki, A.; Forche, A.; Berman, J. Aneuploidy and isochromosome formation in drug-resistant candida albicans. Science 2006, 313, 367–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guin, K.; Chen, Y.; Mishra, R.; Muzaki, S.R.B.; Thimmappa, B.C.; O’Brien, C.E.; Butler, G.; Sanyal, A.; Sanyal, K. Spatial inter-centromeric interactions facilitated the emergence of evolutionary new centromeres. Elife 2020, 9, e58556. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
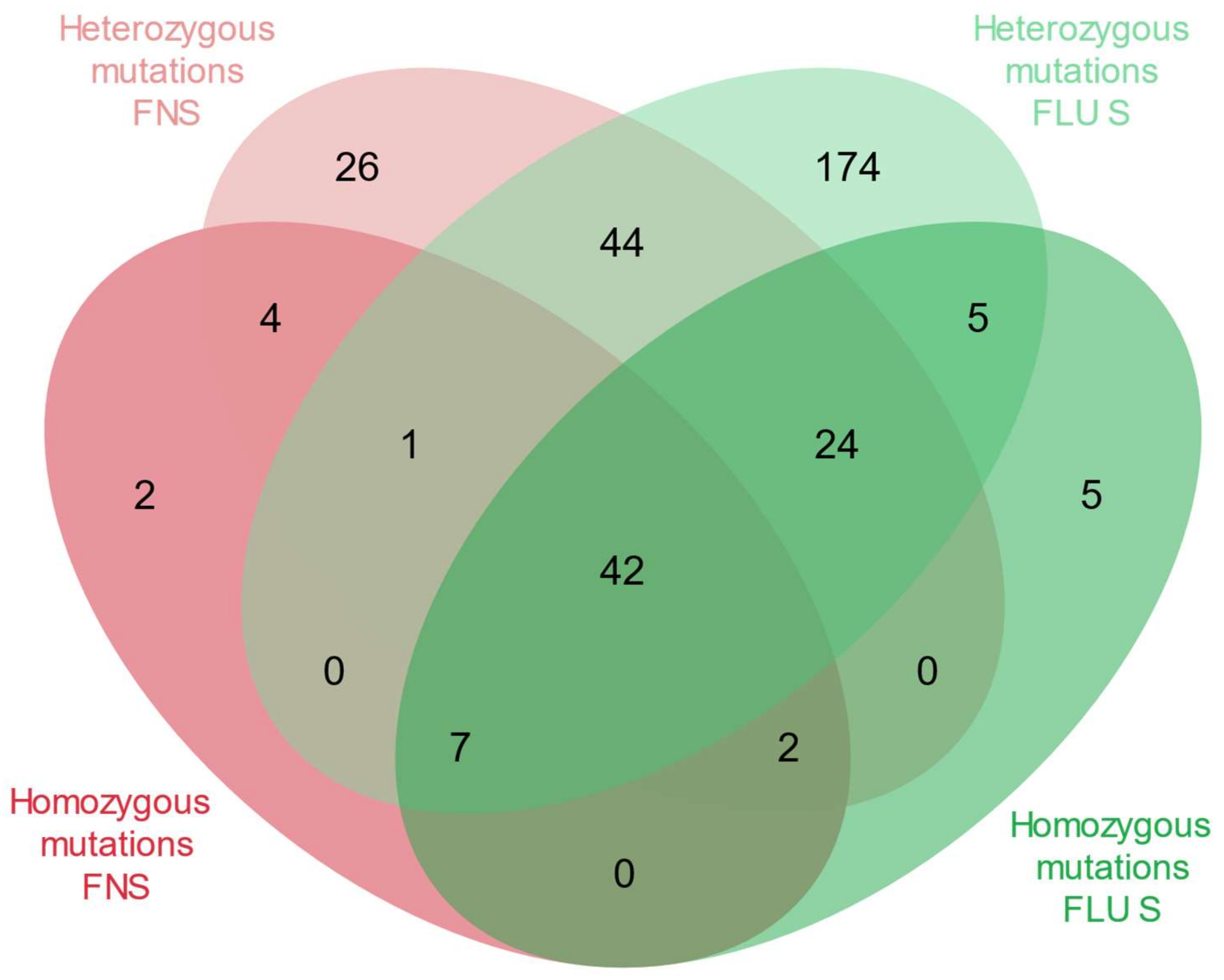{kind=link}
| MICs (mg/L) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| FLU | VOR | POS | ITR | AMP | 5FC | CAS | MCF | ANF | |
| MIC range | 0.5–>256 | 0.015–>8 | 0.03–2 | 0.06–>16 | <0.12–1 | <0.06–1 | 0.008–4 | <0.008–4 | <0.015–0.5 |
| MIC90 | 32 | 2 | 0.5 | 0.5 | 1 | 0.12 | 0.25 | 0.03 | 0.12 |
| GM MIC | 2.36 | 0.18 | 0.19 | 0.28 | 0.78 | 0.08 | 0.06 | 0.02 | 0.06 |
| S or WT n (%) | 66 (79.5) | 52 (63.9) | 34 (41.0) | 75 (90.4) | 83 (100) | 83 (100) | 80 (96.4) | 82 (98.8) | 82 (98.8) |
| I or SDD n (%) | 6 (7.2) | 16 (19.3) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 2 (2.4) | 0 (0) | 1 (1.2) |
| R or non-WT n (%) | 11 (13.3) | 14 (16.9) | 49 (59.0) | 8 (9.6) | 0 (0) | 0 (0) | 1 (1.2) | 1 (1.2) | 0 (0) |
| Isolates | MIC (mg/L) | Amino Acid Substitutions a | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| FLU | VOR | POS | ITR | MCF | ANF | CAS | AMP | 5FC | ||
| Isolates non-susceptible to fluconazole (isolate ID no.) | ||||||||||
| 19-008-0006 | >256 | >8 | 2 | >16 | 0.12 | 0.25 | 0.5 | 1 | <0.06 | S154F, Y132F |
| 19-008-0016 | >256 | 8 | 1 | 4 | 0.03 | 0.03 | 0.03 | 1 | <0.06 | S154F, Y132F |
| 19-008-0007 | 256 | >8 | 1 | 1 | 0.03 | 0.12 | 0.06 | 1 | <0.06 | S154F, Y132F |
| 19-008-0008 | 256 | >8 | 1 | 1 | 0.03 | 0.06 | 0.06 | 1 | <0.06 | S154F, Y132F |
| 19-008-0017 | 256 | 8 | 1 | 1 | 0.03 | 0.03 | 0.03 | 1 | 0.12 | S154F, Y132F |
| 19-5326-037-0003 | 256 | >8 | 0.5 | 1 | 0.03 | 0.06 | 0.25 | 1 | <0.06 | S154F, Y132F |
| 19-008-0055 | 128 | 4 | 0.12 | 0.12 | 0.008 | 0.03 | 0.03 | 1 | 0.5 | S154F, Y132F |
| 19-008-0045 | 64 | 2 | 0.12 | 0.25 | 0.015 | 0.06 | 0.03 | 1 | 0.25 | Y132F |
| 19-008-0015 | 32 | 2 | 0.5 | 0.5 | 0.03 | 0.03 | 0.015 | 1 | 0.06 | S154F, Y132F |
| 19-008-0044 | 32 | 2 | 0.5 | 0.5 | 0.03 | 0.06 | 0.06 | 1 | 0.12 | |
| SRR11235418 # | 32 | 2 | 1 | 0.5 | 0.03 | 0.06 | 0.12 | 1 | 0.12 | |
| 19-008-0005 | 8 | 0.5 | 1 | 1 | 0.12 | 0.12 | 0.25 | 1 | 0.06 | Y257H |
| 19-008-0042 | 4 | 0.5 | 0.25 | 0.25 | 0.015 | 0.03 | 0.03 | 1 | 0.06 | |
| 19-008-0062 | 4 | 0.5 | 0.25 | 0.25 | 0.03 | 0.25 | 0.25 | 1 | 0.12 | |
| 19-008-0043 | 4 | 0.25 | 0.25 | 0.25 | 0.015 | 0.03 | 0.03 | 1 | <0.06 | |
| MB-21-22330 | 4 | 0.5 | 0.25 | 0.25 | 0.015 | <0.015 | 0.03 | 1 | <0.06 | |
| 19-008-0021 | 4 | 0.12 | 0.25 | 0.25 | 0.03 | 0.06 | 0.03 | 0.5 | <0.06 | |
| 19-008-0063 | 4 | 0.12 | 0.06 | 0.12 | 0.03 | 0.06 | 0.03 | 0.25 | <0.06 | |
| Isolates non-susceptible to echinocandins (isolate ID no.) | ||||||||||
| 19-008-0018 | 2 | 0.25 | 0.5 | 0.25 | 4 | 0.5 | 4 | 1 | <0.06 | S30P |
| SRR11235416 # | 1 | 0.06 | 0.12 | 0.25 | 2 | 1 | 8 | 1 | <0.06 | S30P |
| Description | Amino Acid Substitution | Homozygous (%) | Heterozygous (%) | |
|---|---|---|---|---|
| ERG11 | Sterol 14-demethylase | S154F | 39 | 6 |
| Y132F | 44 | 6 | ||
| Y257H | 6 | |||
| CDR1 | ABC multidrug transporter | P418A | 6 | |
| E697G | 11 | |||
| P706T | 6 | |||
| CDR11 | ABC multidrug transporter | K1467R | 11 | |
| D779Y | 6 | |||
| T332I | 6 | |||
| N240K | 39 | |||
| A168G | 11 | |||
| MDR1 | MFS multidrug efflux pump | V15F | 6 | |
| E350K | 11 | |||
| K523E | 6 | |||
| MMR1 | Regulator of MDR1 expression | G697D | 6 | |
| L854S | 6 | |||
| NDT80 | Activator of CDR1 | A89T | 6 | |
| SNQ2 | ABC multidrug transporter | G186D | 6 | |
| V291A | 6 | |||
| V629I | 6 | |||
| I765V | 6 | |||
| G791V | 6 | |||
| STB5 | Regulator of ABC transporter expression | L129F | 6 | 33 |
| T313N | 6 | 22 | ||
| TAC1 | Inducer of ABC transporter expression | F936V | 6 | |
| A446E | 33 | |||
| TPO3 | MFS multidrug efflux pump | T50I | 6 | |
| UPC2 | Regulator of ERG11 | L168P | 28 | |
| YOR | ABC multidrug transporter | T8I | 6 | |
| P44L | 6 | |||
| V575A | 6 | |||
| I1314T | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keighley, C.; Gall, M.; van Hal, S.J.; Halliday, C.L.; Chai, L.Y.A.; Chew, K.L.; Biswas, C.; Slavin, M.A.; Meyer, W.; Sintchenko, V.; et al. Whole Genome Sequencing Shows Genetic Diversity, as Well as Clonal Complex and Gene Polymorphisms Associated with Fluconazole Non-Susceptible Isolates of Candida tropicalis. J. Fungi 2022, 8, 896. https://doi.org/10.3390/jof8090896
Keighley C, Gall M, van Hal SJ, Halliday CL, Chai LYA, Chew KL, Biswas C, Slavin MA, Meyer W, Sintchenko V, et al. Whole Genome Sequencing Shows Genetic Diversity, as Well as Clonal Complex and Gene Polymorphisms Associated with Fluconazole Non-Susceptible Isolates of Candida tropicalis. Journal of Fungi. 2022; 8(9):896. https://doi.org/10.3390/jof8090896
Chicago/Turabian StyleKeighley, Caitlin, Mailie Gall, Sebastiaan J. van Hal, Catriona L. Halliday, Louis Yi Ann Chai, Kean Lee Chew, Chayanika Biswas, Monica A. Slavin, Wieland Meyer, Vitali Sintchenko, and et al. 2022. "Whole Genome Sequencing Shows Genetic Diversity, as Well as Clonal Complex and Gene Polymorphisms Associated with Fluconazole Non-Susceptible Isolates of Candida tropicalis" Journal of Fungi 8, no. 9: 896. https://doi.org/10.3390/jof8090896