
In Vitro Activity of Novel Lipopeptides against Triazole-Resistant Aspergillus fumigatus

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolates
2.2. Antifungals
2.3. Peptide Synthesis
2.4. Preparation of Fungal Suspensions
2.5. MIC Determination
2.6. Time–Kill Assay
2.7. XTT Assay
2.8. Optical Microscopy
2.9. Checkerboard Assays
2.10. Statistical Analysis
3. Results
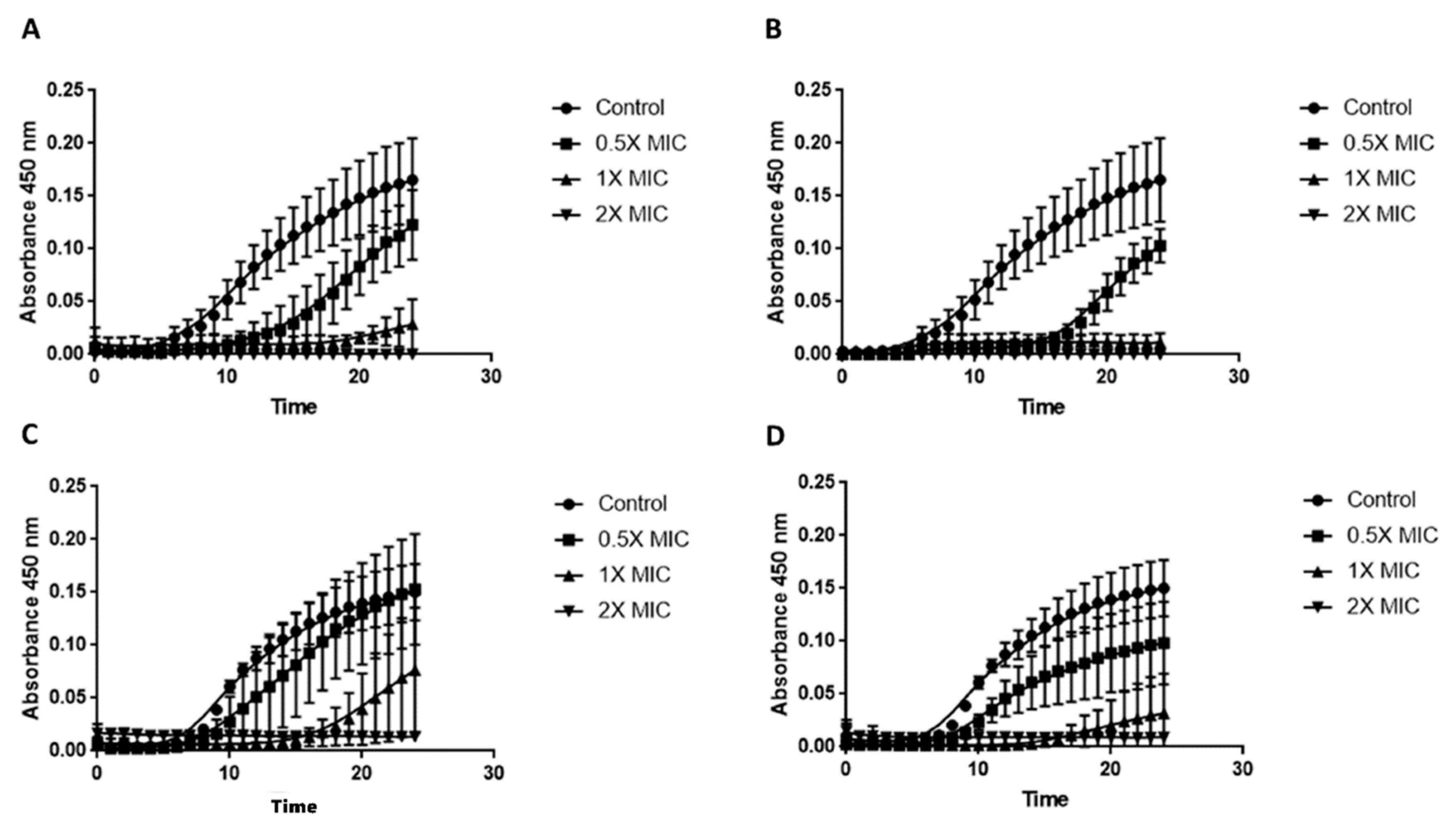
3.1. Time–Kill Assay
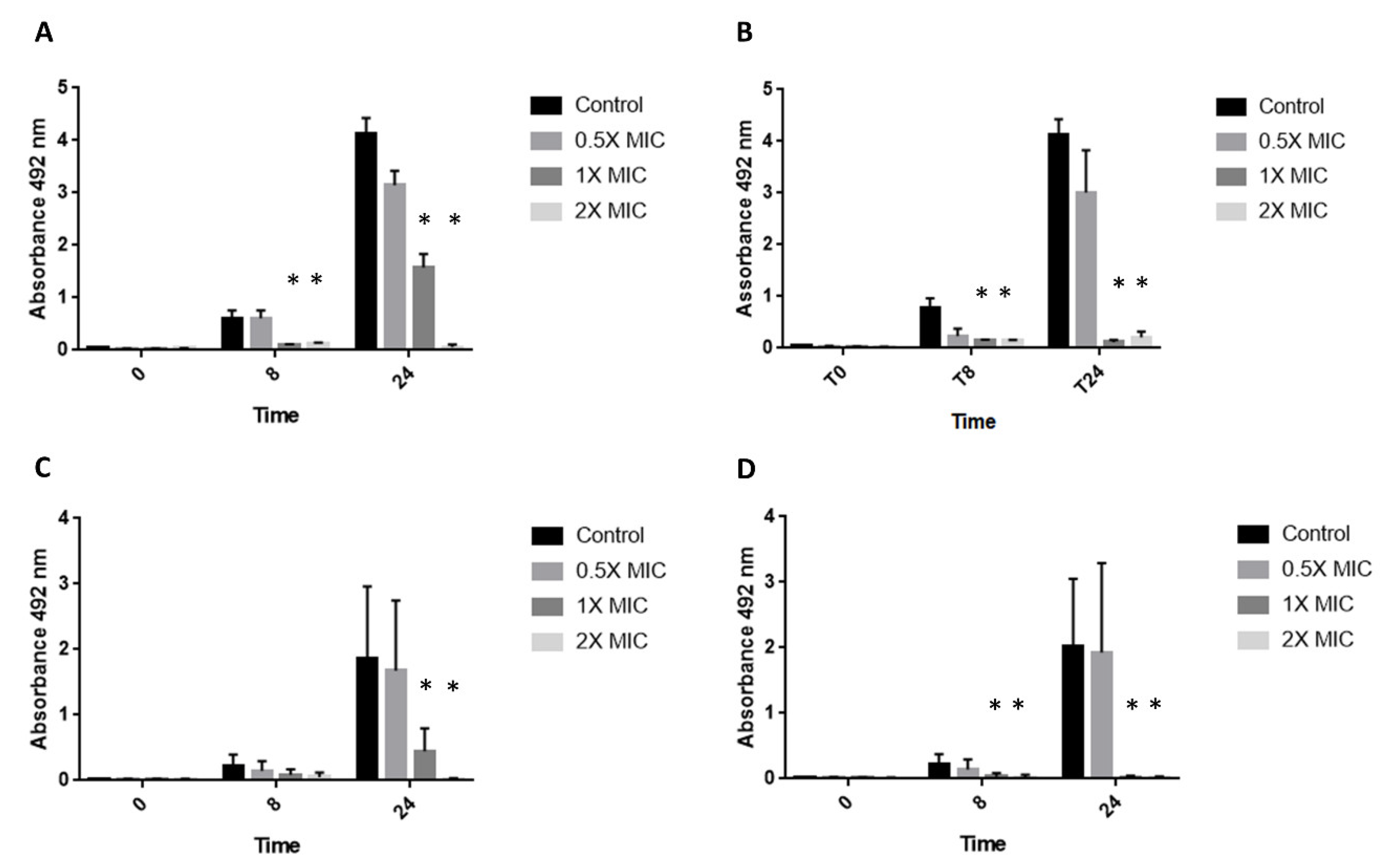
3.2. XTT Assay
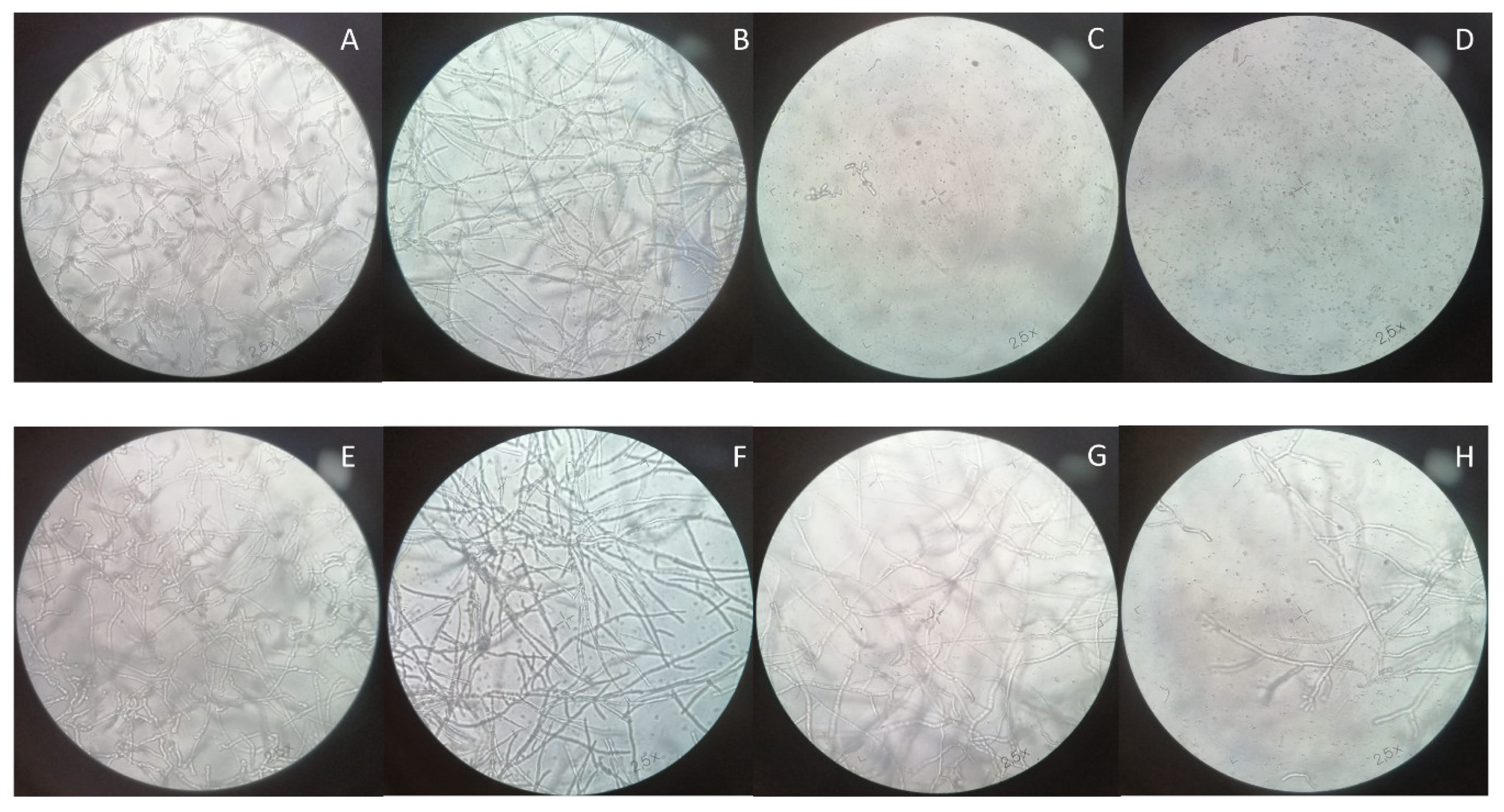
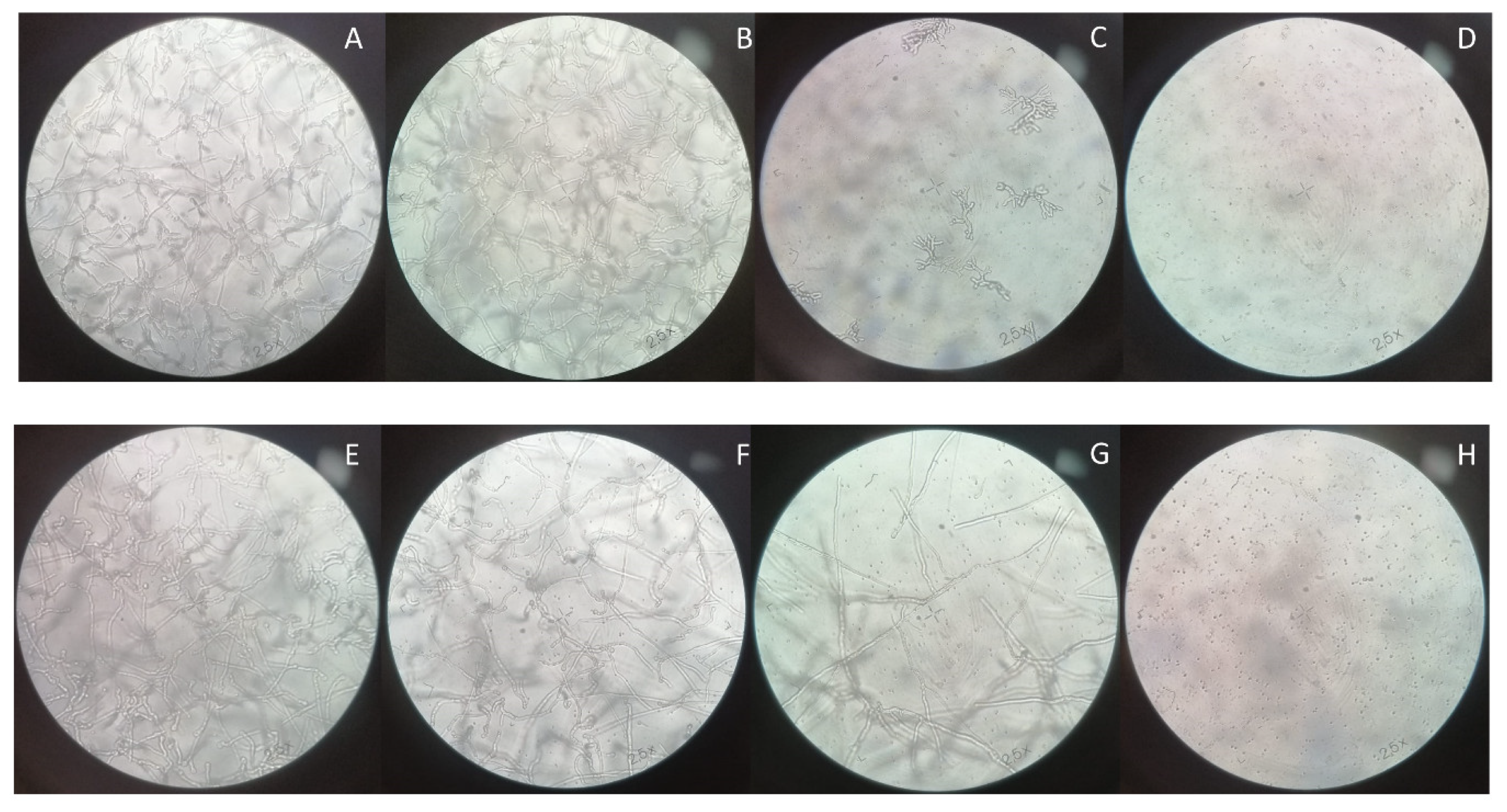
3.3. Optical Microscopy
3.4. Checkerboard Assays
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cadena, J.; Thompson, G.R.; Patterson, T.F. Aspergillosis: Epidemiology, Diagnosis, and Treatment. Infect. Dis. Clin. N. Am. 2021, 35, 415–434. [Google Scholar] [CrossRef] [PubMed]
- Jenks, J.D.; Nam, H.H.; Hoenigl, M. Invasive aspergillosis in critically ill patients: Review of definitions and diagnostic approaches. Mycoses 2021, 64, 1002–1014. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, A.; Osman, M.; Dabboussi, F.; Rafei, R.; Mallat, H.; Papon, N.; Bouchara, J.P.; Hamze, M. Recent trends in the epidemiology, diagnosis, treatment, and mechanisms of resistance in clinical Aspergillus species: A general review with a special focus on the Middle Eastern and North African region. J. Infect. Public Health 2020, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; Tiseo, G.; Menichetti, F. Pulmonary Aspergillosis: An Evolving Challenge for Diagnosis and Treatment. Infect. Dis. Ther. 2020, 9, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Latge, J.; Chamilos, G. Aspergillus fumigatus and Aspergillosis in 2019. Clin. Microbiol. Rev. 2019, 33, e00140-18. [Google Scholar] [CrossRef] [PubMed]
- Prigitano, A.; Esposto, A.M.; Grancini, A.; Biffi, A.; Innocenti, P.; Cavanna, C.; Lallitto, F.; Mollaschi, E.M.G.; Bandettini, R.; Oltolini, C.; et al. Azole resistance in Aspergillus isolates by different types of patients and correlation with environment—An Italian prospective multicentre study (ARiA study). Mycoses 2021, 64, 528–536. [Google Scholar] [CrossRef]
- Burks, C.; Darby, A.; Gómez Londoño, L.; Momany, M.; Brewer, M.T. Azole-resistant Aspergillus fumigatus in the environment: Identifying key reservoirs and hotspot of fungal resistance. PLoS Pathog. 2021, 17, e1009711. [Google Scholar] [CrossRef] [PubMed]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial host defence peptides: Functions and clinical potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [CrossRef] [PubMed]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef]
- Simonetti, O.; Silvestri, C.; Arzeni, D.; Cirioni, O.; Kamysz, W.; Conte, I.; Staffolani, S.; Orsetti, E.; Morciano, A.; Castelli, P.; et al. In vitro activity of the protegrin IB-367 alone and in combination compared with conventional antifungal agents against dermatophytes. Mycoses 2014, 57, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, O.; Ganzetti, G.; Arzeni, D.; Campanati, A.; Marconi, B.; Silvestri, C.; Cirioni, O.; Gabrielli, E.; Lenci, I.; Kamysz, W.; et al. In vitro activity of Tachyplesin III alone and in combination with terbinafine against clinical isolates of dermatophytes. Peptides 2009, 30, 1794–1797. [Google Scholar] [CrossRef]
- Neubauer, D.; Jaśkiewicz, M.; Bauer, M.; Gołacki, K.; Kamysz, W. Ultrashort Cationic Lipopeptides–Effect of N-Terminal Amino Acid and Fatty Acid Type on Antimicrobial Activity and Hemolysis. Molecules 2020, 25, 257. [Google Scholar] [CrossRef]
- EUCAST Definitive Document E.DEF 9.4. 2022. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/AFST/Files/EUCAST_EDef_9.4_method_for_susceptibility_testing_of_moulds.pdf (accessed on 1 April 2022).
- Furnica, D.; Dittmer, S.; Sanders, M.I.; Steinmann, J.; Rath, P.M.; Kirchhoff, L. In Vitro and In Vivo Activity of Luliconazole (NND-502) against Planktonic Cells and Biofilms of Azole Resistant Aspergillus fumigatus. J. Fungi 2022, 8, 350. [Google Scholar] [CrossRef] [PubMed]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef]
- Ullmann, A.J.; Aguado, J.M.; Arikan-Akdagli, S.; Denning, D.W.; Groll, A.H.; Lagrou, K.; Lass-Flörl, C.; Lewis, R.E.; Munoz, P.; Verweij, P.E.; et al. Diagnosis and management of Aspergillus diseases: Executive summary of the 2017 ESCMID-ECMM-ERS guideline. Clin. Microbiol. Infect. 2018, 24 (Suppl. S1), e1–e38. [Google Scholar] [CrossRef] [PubMed]
- Verweij, P.E.; Lucas, J.A.; Arendrup, M.C.; Bowyer, P.; Brinkmann, A.J.F.; Denning, D.W.; Dyer, P.S.; Fisher, M.C.; Geenen, P.L.; Gisi, U.; et al. The one health problem of azole resistance in Aspergillus fumigatus: Current insights and future research agenda. Fungal Biol. Rev. 2020, 34, 202–214. [Google Scholar] [CrossRef]
- Meena, K.R.; Kanwar, S.S. Lipopeptides as the antifungal and antibacterial agents: Applications in food safety and therapeutics. Biomed. Res. Int. 2015, 2015, 473050. [Google Scholar] [CrossRef] [PubMed]
- Kamysz, E.; Simonetti, O.; Cirioni, O.; Arzeni, D.; Ganzetti, G.; Campanati, A.; Giacometti, A.; Gabrielli, E.; Silvestri, C.; Kamysz, W.; et al. In vitro activity of the lipopeptide PAL-Lys-Lys-NH2, alone and in combination with antifungal agents, against clinical isolates of Candida spp. Peptides 2011, 32, 99–103. [Google Scholar] [CrossRef]
- Simonetti, O.; Arzeni, D.; Ganzetti, G.; Silvestri, C.; Cirioni, O.; Gabrielli, E.; Castelletti, S.; Kamysz, W.; Kamysz, E.; Scalise, G.; et al. In vitro activity of the lipopeptide derivative (Pal-Lys-Lys-NH), alone and in combination with antifungal agents, against clinical isolates of dermatophytes. Br. J. Dermatol. 2009, 161, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Barchiesi, F.; Silvestri, C.; Arzeni, D.; Ganzetti, G.; Castelletti, S.; Simonetti, O.; Cirioni, O.; Kamysz, W.; Kamysz, E.; Spreghini, E.; et al. In vitro susceptibility of dermatophytes to conventional and alternative antifungal agents. Med. Mycol. 2009, 47, 321–326. [Google Scholar] [CrossRef]
- Simonetti, O.; Rizzetto, G.; Molinelli, E.; Cirioni, O.; Offidani, A. Review: A Safety Profile of Dalbavancin for On- and Off-Label Utilization. Ther. Clin. Risk Manag. 2021, 17, 223–232. [Google Scholar] [CrossRef]
- Huang, H.W. DAPTOMYCIN, its membrane-active mechanism vs. that of other antimicrobial peptide. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183395. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, O.; Lucarini, G.; Morroni, G.; Orlando, F.; Lazzarini, R.; Zizzi, A.; Brescini, L.; Provinciali, M.; Giacometti, A.; Offidani, A.; et al. New Evidence and Insights on Dalbavancin and Wound Healing in a Mouse Model of Skin Infection. Antimicrob. Agents Chemother. 2020, 64, e02062-19. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, O.; Lucarini, G.; Orlando, F.; Pierpaoli, E.; Ghiselli, R.; Provinciali, M.; Castelli, P.; Guerrieri, M.; Di Primio, R.; Offidani, A.; et al. Role of Daptomycin on Burn Wound Healing in an Animal Methicillin-Resistant Staphylococcus aureus Infection Model. Antimicrob. Agents Chemother. 2017, 61, e00606-17. [Google Scholar] [CrossRef]
- Aruanno, M.; Glampedakis, E.; Lamoth, F. Echinocandins for the Treatment of Invasive Aspergillosis: From Laboratory to Bedside. Antimicrob. Agents Chemother. 2019, 63, e00399-19. [Google Scholar] [CrossRef] [PubMed]
- Makovitzki, A.; Avrahami, D.; Shai, Y. Ultrashort antibacterial and antifungal lipopeptides. Proc. Natl. Acad. Sci. USA 2006, 103, 15997–16002. [Google Scholar] [CrossRef] [PubMed]
- Ballarad, E.; Yucel, R.; Melchers, W.J.G.; Brown, A.J.P.; Verweij, P.E.; Warris, A. Antifungal Activity of Antimicrobial Peptides and Proteins against Aspergillus fumigatus. J. Fungi 2020, 6, 65. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, T.R.; Rhomberg, P.R.; Sader, H.S.; Jones, R.N. Antimicrobial activity of omiganan pentahydrochloride against contemporary fungal pathogens responsible for catheter-associated infections. Antimicrob. Agents Chemother. 2008, 52, 1187–1189. [Google Scholar] [CrossRef] [PubMed]
- D’auria, F.D.; Casciaro, B.; De Angelis, M.; Marcocci, M.E.; Palamara, A.T.; Nencioni, L.; Mangoni, M.L. Antifungal Activity of the Frog Skin Peptide Temporin G and Its Effect on Candida albicans Virulence Factors. Int. J. Mol. Sci. 2022, 23, 6345. [Google Scholar] [CrossRef] [PubMed]
- Dell’Olmo, E.; Gaglione, R.; Cesaro, A.; Cafaro, V.; Teertstra, W.R.; de Cock, H.; Notomista, E.; Haagsman, H.P.; Veldhuizen, E.J.A.; Arciello, A. Host defence peptides identified in human apolipoprotein B as promising antifungal agents. Appl. Microbiol. Biotechnol. 2021, 105, 1953–1964. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, M.; Zhu, X.; Peng, Y.; Fu, T.; Hu, C.H.; Cai, J.; Liao, G. Development of Lipo-γ-AA Peptides as Potent Antifungal Agents. J. Med. Chem. 2022, 65, 8029–8039. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Cantero, A.; López-Fernández, L.; Guarro, J.; Capilla, J. Azole resistance mechanisms in Aspergillus: Update and recent advances. Int. J. Antimicrob. Agents 2020, 55, 105807. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | MIC (µg/mL) | ||
|---|---|---|---|
| C14-NleRR-NH2 | C14-WRR-NH2 | Isavuconazole | |
| A. fumigatus SSI-4524 | 8 | 8 | 8 |
| A. fumigatus SSI-5586 | 16 | 16 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fioriti, S.; Cirioni, O.; Simonetti, O.; Franca, L.; Candelaresi, B.; Pallotta, F.; Neubauer, D.; Kamysz, E.; Kamysz, W.; Canovari, B.; et al. In Vitro Activity of Novel Lipopeptides against Triazole-Resistant Aspergillus fumigatus. J. Fungi 2022, 8, 872. https://doi.org/10.3390/jof8080872
Fioriti S, Cirioni O, Simonetti O, Franca L, Candelaresi B, Pallotta F, Neubauer D, Kamysz E, Kamysz W, Canovari B, et al. In Vitro Activity of Novel Lipopeptides against Triazole-Resistant Aspergillus fumigatus. Journal of Fungi. 2022; 8(8):872. https://doi.org/10.3390/jof8080872
Chicago/Turabian StyleFioriti, Simona, Oscar Cirioni, Oriana Simonetti, Lucia Franca, Bianca Candelaresi, Francesco Pallotta, Damian Neubauer, Elzbieta Kamysz, Wojciech Kamysz, Benedetta Canovari, and et al. 2022. "In Vitro Activity of Novel Lipopeptides against Triazole-Resistant Aspergillus fumigatus" Journal of Fungi 8, no. 8: 872. https://doi.org/10.3390/jof8080872