
Cystathionine Gamma-Lyase Regulate Psilocybin Biosynthesis in Gymnopilus dilepis Mushroom via Amino Acid Metabolism Pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strain and Sample Collection
2.2. Psilocybin Extraction and Determination
2.3. L-Serine Extraction and Determination
2.4. Transcriptome Analysis
2.5. Quantitative Real-Time PCR (qRT-PCR) Analysis
2.6. Exogenous Addition Experiment
- PAG: 1 mM PAG was added to the culture medium, followed by incubation for 24 h;
- PAG + L-Cys: 1 mM PAG and 0.1% L-cysteine were added to the culture medium, followed by incubation for 24 h;
- Control group: the same amount of sterile water as the other groups was added, and the sample was incubated for 24 h.
2.7. Statistical Analysis
3. Results
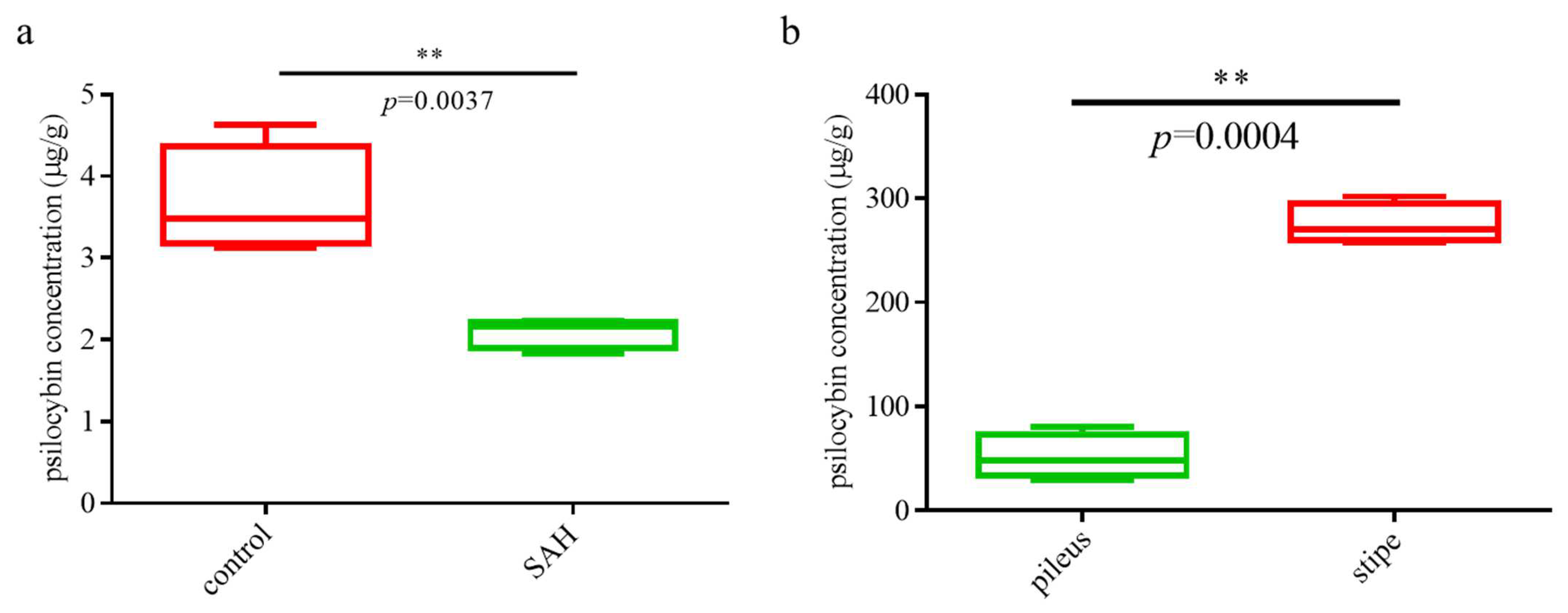
3.1. Comparative Analysis of Psilocybin in Samples from Different Treatments and Parts
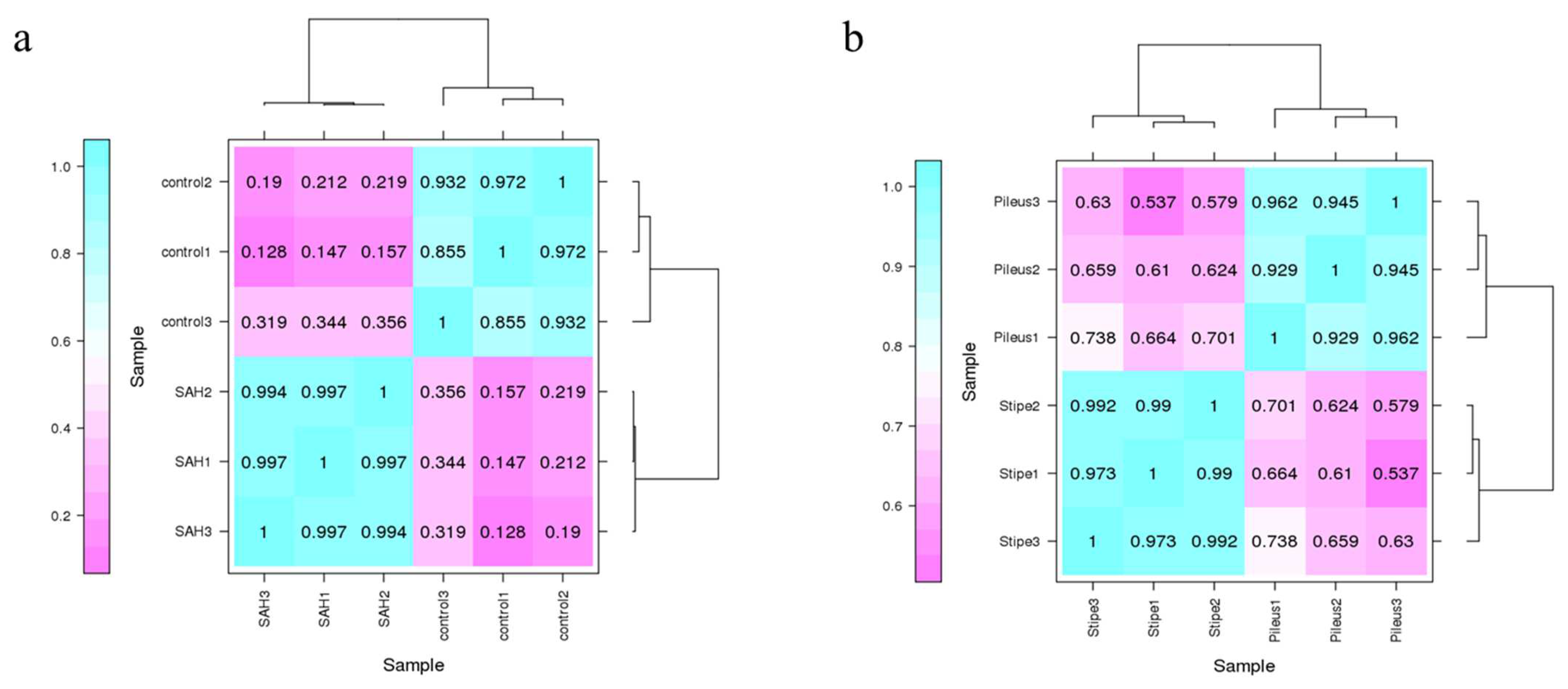
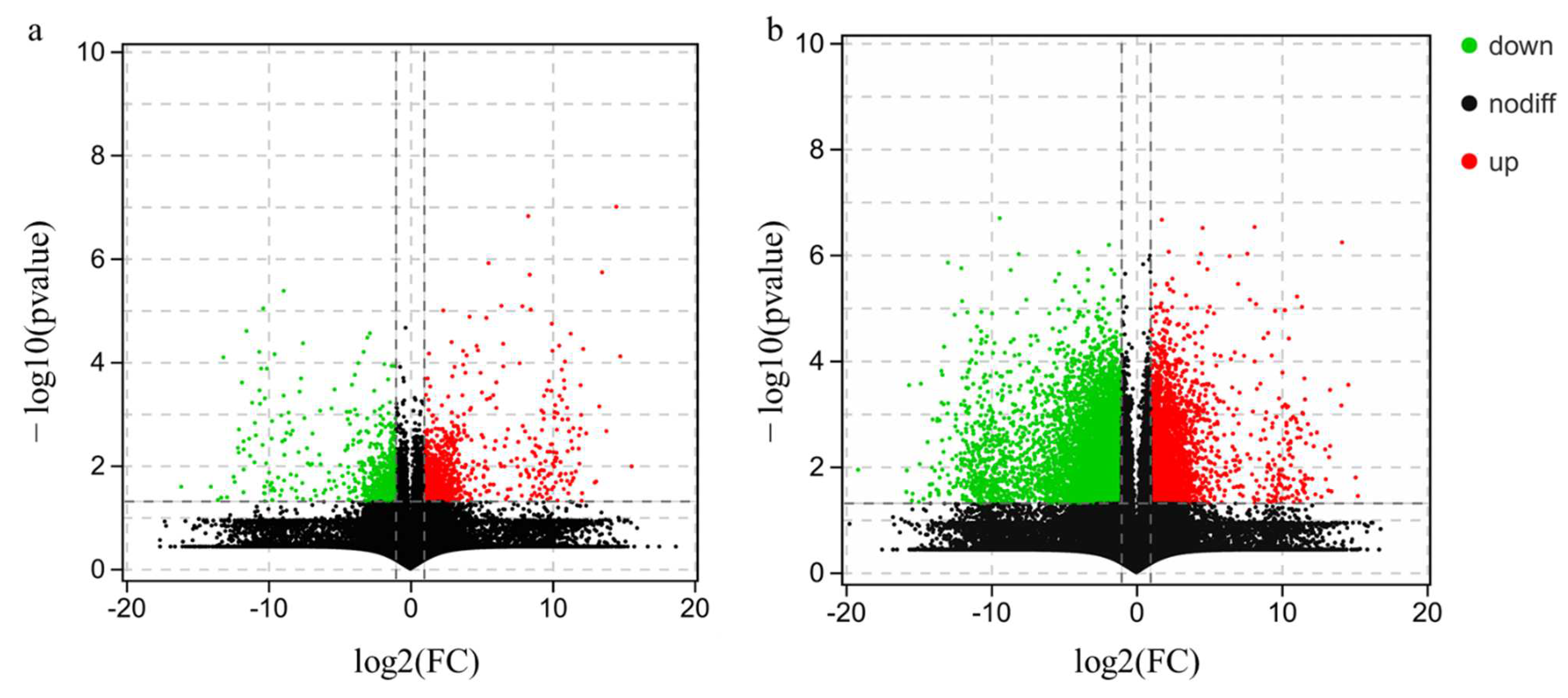
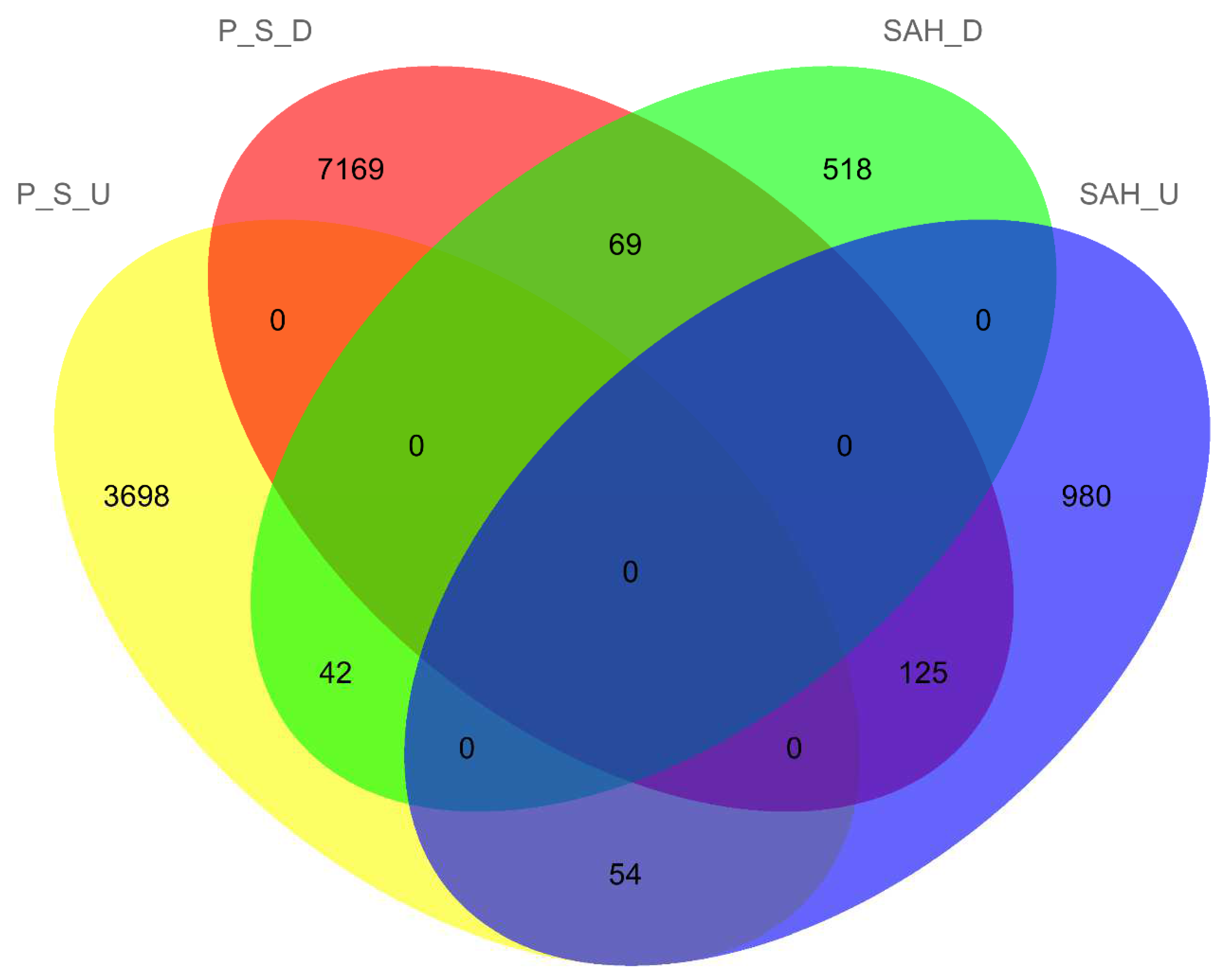
3.2. Sample Correlation Analysis and Differentially Expressed Genes (DEGs) Analysis
3.3. GO Enrichment Analysis for DEGs
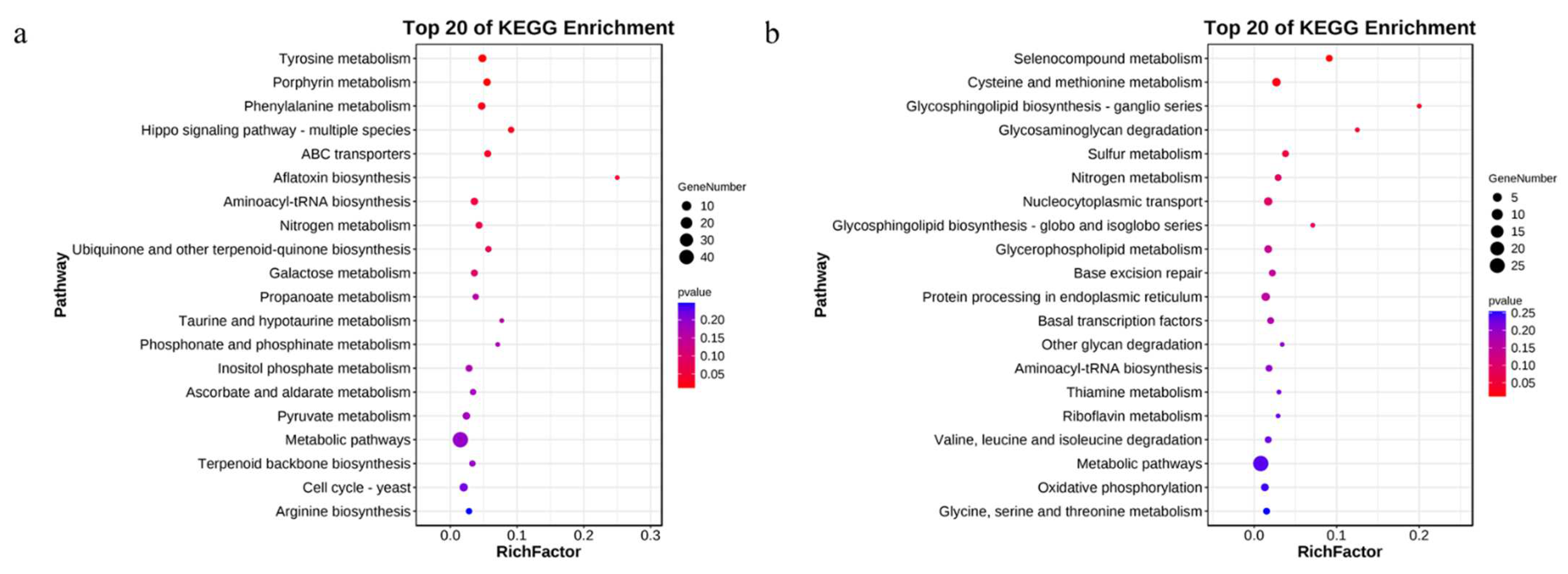
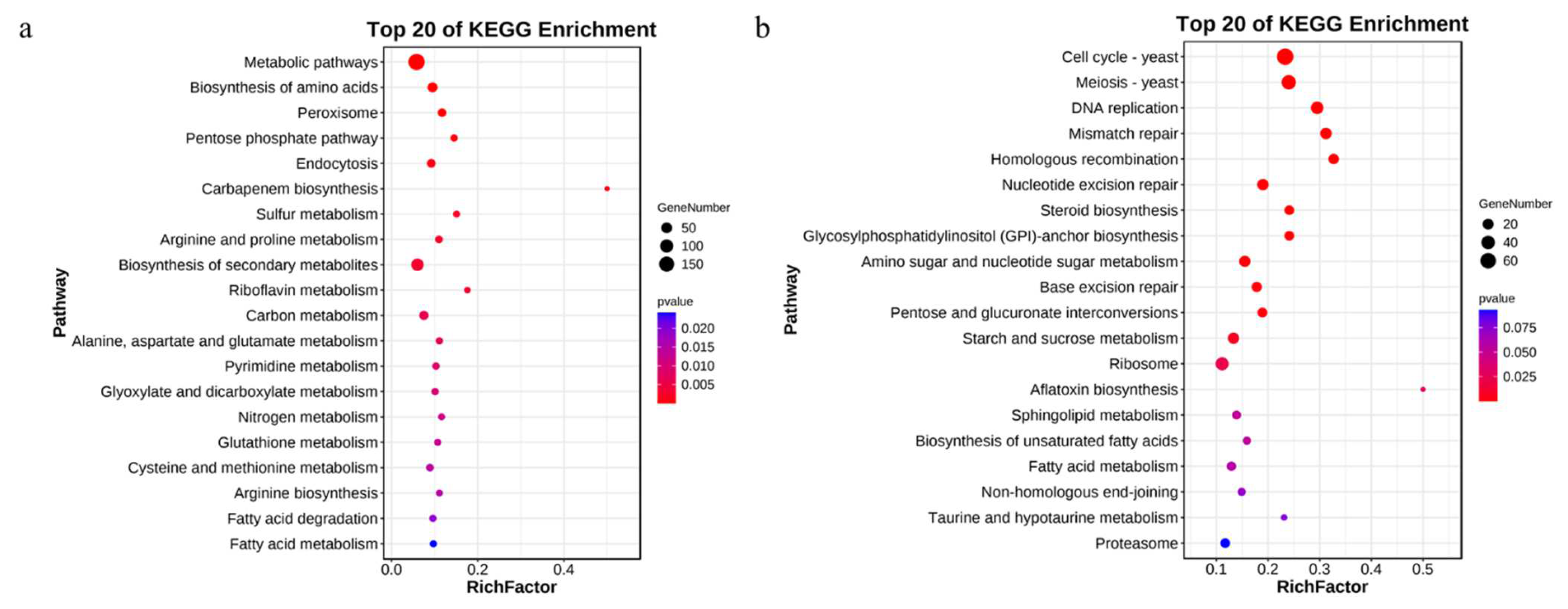
3.4. KEGG Enrichment Analysis of the DEGs
3.5. Cysteine and Methionine Metabolism Is Correlated with Psilocybin Synthesis
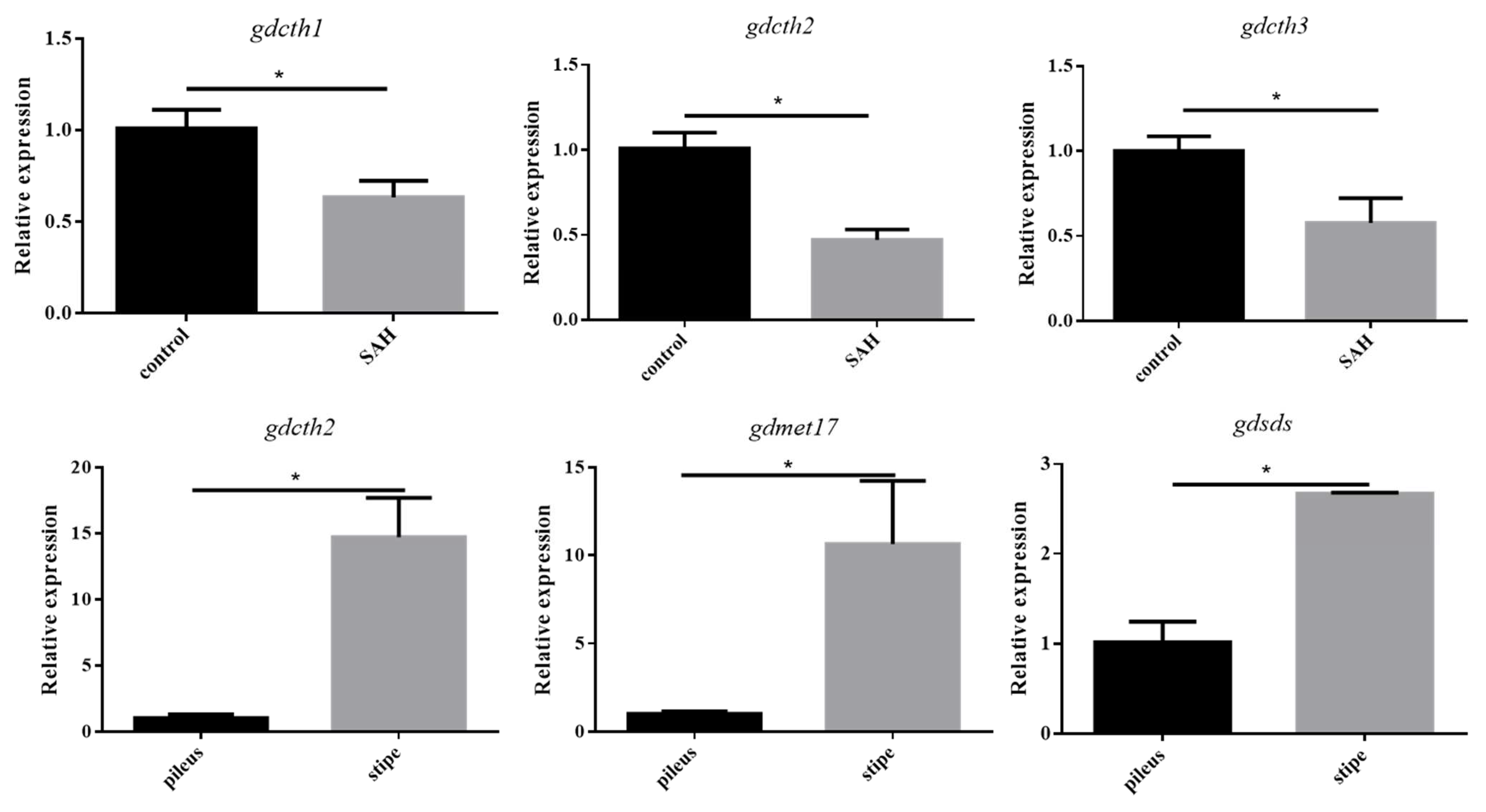
3.6. Validation of RNA-Seq Results Using qRT-PCR
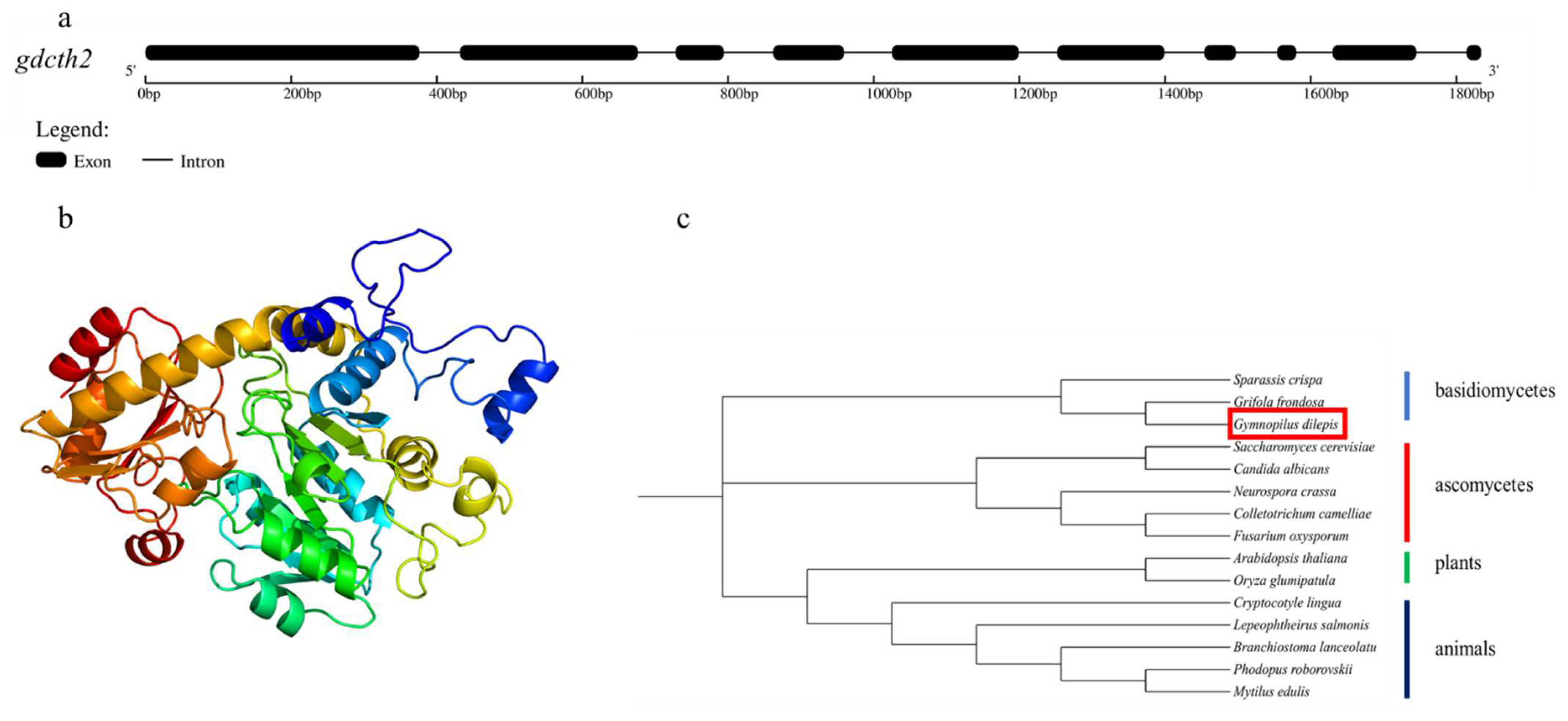
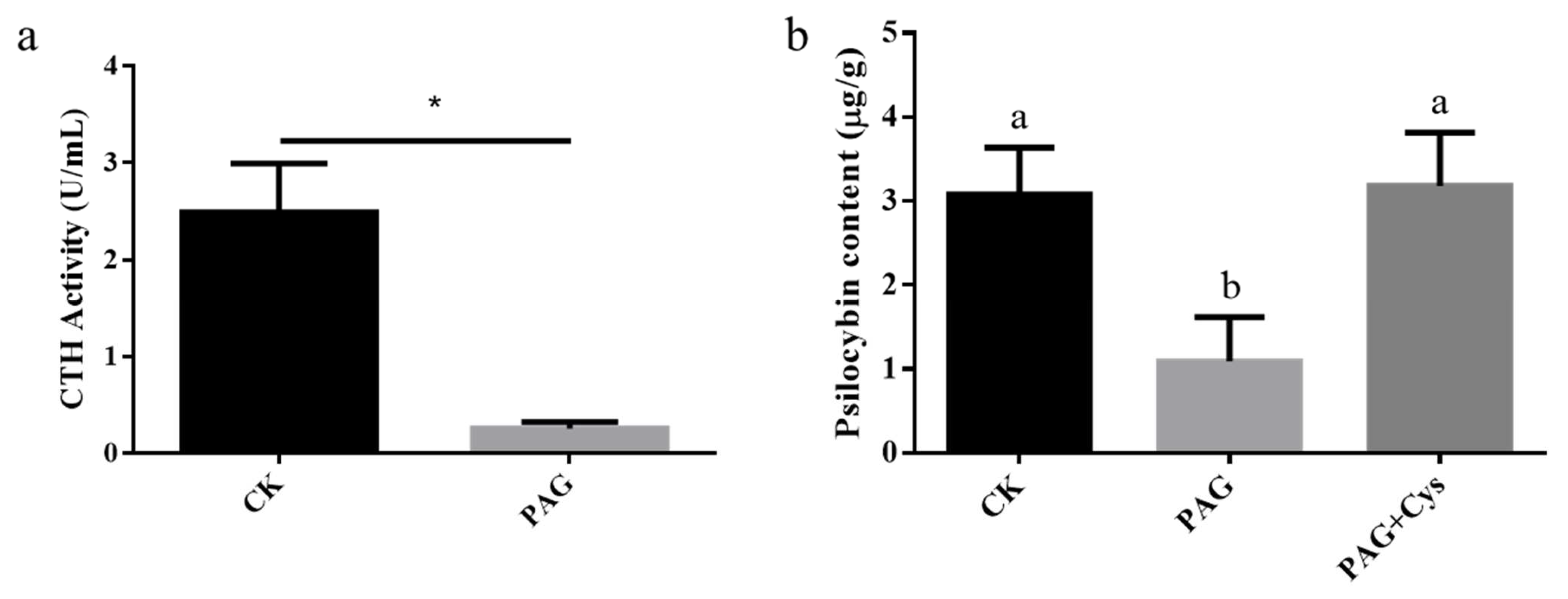
3.7. CTH Related to Biosynthesis of Psilocybin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andersson, C.; Kristinsson, J.; Gry, J. Occurrence and Use of Hallucinogenic Mushrooms Containing Psilocybin Alkaloids; Nordic Council of Ministers: Copenhagen, Denmark, 2009; pp. 75–90. [Google Scholar]
- Hofmann, A.; Heim, R.; Brack, A.; Kobel, H. Psilocybin, ein psychotroper Wirkstoff aus dem mexikanischen Rauschpilz Psilocybe mexicana Heim. Experientia 1958, 14, 107–109. [Google Scholar] [CrossRef] [PubMed]
- Nichols, D.E. Psilocybin: From ancient magic to modern medicine. J. Antibiot. 2020, 73, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, R.R.; Richards, W.A.; Johnson, M.W.; McCann, U.D.; Jesse, R. Mystical-type experiences occasioned by psilocybin mediate the attribution of personal meaning and spiritual significance 14 months later. J. Psychopharmacol. 2008, 22, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Kometer, M.; Schmidt, A.; Bachmann, R.; Studerus, E.; Seifritz, E.; Vollenweider, F.X. Psilocybin biases facial recognition, goal-directed behavior, and mood state toward positive relative to negative emotions through different serotonergic subreceptors. Biol. Psychiatry 2012, 72, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Grob, C.S.; Danforth, A.L.; Chopra, G.S.; Hagerty, M.; McKay, C.R.; Halberstadt, A.L.; Greer, G.R. Pilot study of psilocybin treatment for anxiety in patients with advanced-stage cancer. Arch. Gen. Psychiatry 2011, 68, 71–78. [Google Scholar] [CrossRef]
- Carhart-Harris, R.L.; Bolstridge, M.; Rucker, J.; Day, C.M.; Erritzoe, D.; Kaelen, M.; Bloomfield, M.; Rickard, J.A.; Forbes, B.; Feilding, A.; et al. Psilocybin with psychological support for treatment-resistant depression: An open-label feasibility study. Lancet Psychiatry 2016, 3, 619–627. [Google Scholar] [CrossRef]
- Davis, A.K.; Barrett, F.S.; May, D.G.; Cosimano, M.P.; Sepeda, N.D.; Johnson, M.W.; Finan, P.H.; Griffiths, R.R. Effects of psilocybin-assisted therapy on major depressive disorder: A randomized clinical trial. JAMA Psychiatry 2021, 78, 481–489. [Google Scholar] [CrossRef]
- Rootman, J.M.; Kiraga, M.; Kryskow, P.; Harvey, K.; Stamets, P.; Santos-Brault, E.; Kuypers, K.P.C.; Walsh, Z. Psilocybin microdosers demonstrate greater observed improvements in mood and mental health at one month relative to non-microdosing controls. Sci. Rep. 2022, 12, 11091. [Google Scholar] [CrossRef]
- Burdick, B.V.; Adinoff, B. A proposal to evaluate mechanistic efficacy of hallucinogens in addiction treatment. Am. J. Drug Alcohol AB 2013, 39, 291–297. [Google Scholar] [CrossRef]
- Johnson, M.W.; Garcia-Romeu, A.; Griffiths, R.R. Long-term follow-up of psilocybin-facilitated smoking cessation. Am. J. Drug Alcohol AB 2017, 43, 55–60. [Google Scholar] [CrossRef]
- Bogenschutz, M.P.; Forcehimes, A.A.; Pommy, J.A.; Wilcox, C.E.; Barbosa, P.C.; Strassman, R.J. Psilocybin-assisted treatment for alcohol dependence: A proof-of-concept study. J. Psychopharmacol. 2015, 29, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Studerus, E.; Kometer, M.; Hasler, F.; Vollenweider, F.X. Acute, subacute and long-term subjective effects of psilocybin in healthy humans: A pooled analysis of experimental studies. J. Psychopharmacol. 2011, 25, 1434–1452. [Google Scholar] [CrossRef] [PubMed]
- Barrett, F.S.; Krimmel, S.R.; Griffiths, R.R.; Seminowicz, D.A.; Mathur, B.N. Psilocybin acutely alters the functional connectivity of the claustrum with brain networks that support perception, memory, and attention. Neuroimage 2020, 218, 116980. [Google Scholar] [CrossRef] [PubMed]
- Barrett, F.S.; Doss, M.K.; Sepeda, N.D.; Pekar, J.J.; Griffiths, R.R. Emotions and brain function are altered up to one month after a single high dose of psilocybin. Sci. Rep. 2020, 10, 2214. [Google Scholar] [CrossRef] [PubMed]
- Gotvaldová, K.; Hájková, K.; Borovička, J.; Jurok, R.; Cihlářová, P.; Kuchař, M. Stability of psilocybin and its four analogs in the biomass of the psychotropic mushroom Psilocybe cubensis. Drug Test. Anal. 2021, 13, 439–446. [Google Scholar] [CrossRef]
- Demmler, R.; Fricke, J.; Dörner, S.; Gressler, M.; Hoffmeister, D. S-Adenosyl-l-Methionine salvage impacts psilocybin formation in “magic” mushrooms. ChemBioChem 2020, 21, 1364–1371. [Google Scholar] [CrossRef]
- Kargbo, R.B.; Sherwood, A.; Walker, A.; Cozzi, N.V.; Dagger, R.E.; Sable, J.; O’Hern, K.; Kaylo, K.; Patterson, T.; Tarpley, G.; et al. Direct phosphorylation of psilocin enables optimized cGMP kilogram-scale manufacture of psilocybin. ACS Omega 2020, 5, 16959–16966. [Google Scholar] [CrossRef]
- Adams, A.M.; Kaplan, N.A.; Wei, Z.; Brinton, J.D.; Monnier, C.S.; Enacopol, A.L.; Ramelot, T.A.; Jones, J.A. In vivo production of psilocybin in E. coli. Metab. Eng. 2019, 56, 111–119. [Google Scholar] [CrossRef]
- Hoefgen, S.; Lin, J.; Fricke, J.; Stroe, M.C.; Mattern, D.J.; Kufs, J.E.; Hortschansky, P.; Brakhage, A.A.; Hoffmeister, D.; Valiante, V. Facile assembly and fluorescence-based screening method for heterologous expression of biosynthetic pathways in fungi. Metab. Eng. 2018, 48, 44–51. [Google Scholar] [CrossRef]
- Fricke, J.; Blei, F.; Hoffmeister, D. Enzymatic synthesis of psilocybin. Angew. Chem. Int. Ed. 2017, 56, 12352–12355. [Google Scholar] [CrossRef]
- Awan, A.R.; Winter, J.M.; Turner, D.; Shaw, W.M.; Suz, L.M.; Bradshaw, A.J.; Ellis, T.; Dentinger, B.T. Convergent evolution of psilocybin biosynthesis by psychedelic mushrooms. bioRxiv 2018, 374199. [Google Scholar] [CrossRef]
- Boyce, G.R.; Gluck-Thaler, E.; Slot, J.C.; Stajich, J.E.; Davis, W.J.; James, T.Y.; Cooley, J.R.; Panaccione, D.G.; Eilenberg, J.; Licht, H.H.; et al. Psychoactive plant-and mushroom-associated alkaloids from two behavior modifying cicada pathogens. Fungal Ecol. 2019, 41, 147–164. [Google Scholar] [CrossRef]
- Reynolds, H.T.; Vijayakumar, V.; Gluck-Thaler, E.; Korotkin, H.B.; Matheny, P.B.; Slot, J.C. Horizontal gene cluster transfer increased hallucinogenic mushroom diversity. Evol. lett. 2018, 2, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Blei, F.; Baldeweg, F.; Fricke, J.; Hoffmeister, D. Biocatalytic production of psilocybin and derivatives in tryptophan synthase—Enhanced reactions. Chem. Eur. J. 2018, 24, 10028–10031. [Google Scholar] [CrossRef] [PubMed]
- Torrens-Spence, M.P.; Liu, C.T.; Pluskal, T.; Chung, Y.K.; Weng, J.K. Monoamine biosynthesis via a noncanonical calcium-activatable aromatic amino acid decarboxylase in psilocybin mushroom. ACS Chem. Biol. 2018, 13, 3343–3353. [Google Scholar] [CrossRef]
- McKernan, K.; Kane, L.; Helbert, Y.; Zhang, L.; Houde, N.; McLaughlin, S. A whole genome atlas of 81 Psilocybe genomes as a resource for psilocybin production. F1000Research 2021, 10, 961. [Google Scholar] [CrossRef]
- Suwannarach, N.; Kumla, J.; Sri-Ngernyuang, K.; Lumyong, S. Gymnopilus dilepis, a new record in Thailand. Mycotaxon 2017, 132, 337–341. [Google Scholar] [CrossRef]
- Rees, B.J.; Marchant, A.; Zuccarello, G.C. A tale of two species-possible origins of red to purple-coloured Gymnopilus species in Europe. Australas. Mycol. 2004, 22, 57–72. [Google Scholar]
- Fricke, J.; Lenz, C.; Wick, J.; Blei, F.; Hoffmeister, D. Production options for psilocybin: Making of the magic. Chem. Eur. J. 2019, 25, 897–903. [Google Scholar] [CrossRef]
- Gambaro, V.; Roda, G.; Visconti, G.L.; Arnoldi, S.; Casagni, E.; Dell’Acqua, L.; Fare, F.; Paladino, E.; Rusconi, C.; Arioli, S.; et al. DNA-based taxonomic identification of basidiospores in hallucinogenic mushrooms cultivated in “grow-kits” seized by the police: LC-UV quali-quantitative determination of psilocybin and psilocin. J. Pharm. Biomed. 2016, 125, 427–432. [Google Scholar] [CrossRef]
- Guan, L.; Ju, B.; Zhao, M.; Zhu, H.; Chen, L.; Wang, R.; Gao, H.; Wang, Z. Influence of drying process on furostanoside and spirostanoside profiles of Paridis Rhizoma by combination of HPLC, UPLC and UPLC-QTOF-MS/MS analyses. J. Pharm. Biomed. 2021, 197, 113932. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.F.; Tycksen, E.D.; Sandell, L.J.; Brophy, R.H. Advantages of RNA-seq compared to RNA microarrays for transcriptome profiling of anterior cruciate ligament tears. J. Orthop. Res. 2018, 36, 484–497. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.J.; Tong, Z.J.; Liu, Y.Y.; Li, Y.N.; Zhao, C.; Mukhtar, I.; Tao, Y.X.; Chen, B.Z.; Deng, Y.J.; Xie, B.G. Comparative Transcriptomics of Flammulina filiformis suggests a high CO2 concentration inhibits early pileus expansion by decreasing cell division control pathways. Int. J. Mol. Sci. 2019, 20, 5923. [Google Scholar] [CrossRef]
- Yan, J.; Chekanova, J.; Liu, Y.; Gan, B.; Long, Y.; Han, X.; Tong, Z.; Miao, J.; Lian, L.; Xie, B.; et al. Reactive Oxygen Species Distribution Involved in Stipe Gradient Elongation in the Mushroom Flammulina filiformis. Cells 2022, 11, 1896. [Google Scholar] [CrossRef] [PubMed]
- Saar-Reismaa, P.; Vaher, M.; Kaljurand, M.; Kulp, M.; Mazina-Šinkar, J. Simultaneous determination of γ-hydroxybutyric acid, ibotenic acid and psilocybin in saliva samples by capillary electrophoresis coupled with a contactless conductivity detector. Anal. Methods 2017, 9, 3128–3133. [Google Scholar] [CrossRef]
- Zhao, D.S.; Ma, X.L.; Li, X.X.; Zhang, H.B.; Mian, Q.H.; Meng, L.; Chen, J. Determination of 18 kinds of free amino acids in edible and medicinal Allium using pre-column derivatization HPLC. Chin. J. Pharm. Anal. 2013, 33, 963–968. [Google Scholar]
- Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.C.; Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007, 35, W182–W185. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
- Tao, Y.X.; Van Peer, A.F.; Huang, Q.H.; Shao, Y.; Zhang, L.; Xie, B.; Jiang, Y.J.; Zhu, J.; Xie, B.G. Identification of novel and robust internal control genes from Volvariella volvacea that are suitable for RT-qPCR in filamentous fungi. Sci. Rep. 2016, 6, 29236. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chrzanowska, M.; Grajewska, A.; Rozwadowska, M.D. Asymmetric synthesis of isoquinoline alkaloids: 2004–2015. Chem. Rev. 2016, 116, 12369–12465. [Google Scholar] [CrossRef] [PubMed]
- Fricke, J.; Kargbo, R.; Regestein, L.; Lenz, C.; Peschel, G.; Rosenbaum, M.A.; Sherwood, A.; Hoffmeister, D. Scalable hybrid synthetic/biocatalytic route to psilocybin. Chem. Eur. J. 2020, 26, 8281–8285. [Google Scholar] [CrossRef] [PubMed]
- Konkel, Z.; Scott, K.; Slot, J.C. Draft genome sequence of the termite-associated “cuckoo fungus,” Athelia (Fibularhizoctonia) sp. TMB Strain TB5. Microbiol. Resour. Ann. 2021, 10, e01230-20. [Google Scholar] [CrossRef] [PubMed]
- Back, K.; Tan, D.X.; Reiter, R.J. Melatonin biosynthesis in plants: Multiple pathways catalyze tryptophan to melatonin in the cytoplasm or chloroplasts. J. Pineal. Res. 2016, 61, 426–437. [Google Scholar] [CrossRef]
- Resende, D.I.; Boonpothong, P.; Sousa, E.; Kijjoa, A.; Pinto, M.M. Chemistry of the fumiquinazolines and structurally related alkaloids. Nat. Prod. Rep. 2019, 36, 7–34. [Google Scholar] [CrossRef]
- Schenck, C.A.; Maeda, H.A. Tyrosine biosynthesis, metabolism, and catabolism in plants. Phytochemistry 2018, 149, 82–102. [Google Scholar] [CrossRef]
- Xu, J.J.; Fang, X.; Li, C.Y.; Yang, L.; Chen, X.Y. General and specialized tyrosine metabolism pathways in plants. Abiotech 2020, 1, 97–105. [Google Scholar] [CrossRef]
- Pascual, M.B.; El-Azaz, J.; de la Torre, F.N.; Cañas, R.A.; Avila, C.; Cánovas, F.M. Biosynthesis and metabolic fate of phenylalanine in conifers. Front. Plant Sci. 2016, 7, 1030. [Google Scholar] [CrossRef]
- Yang, G.; Wei, Q.; Huang, H.; Xia, J. Amino acid transporters in plant cells: A brief review. Plants 2020, 9, 967. [Google Scholar] [CrossRef]
- Feng, G.; Qin, Z.; Yan, J.; Zhang, X.; Hu, Y. Arabidopsis ORGAN SIZE RELATED1 regulates organ growth and final organ size in orchestration with ARGOS and ARL. New Phytol. 2011, 191, 635–646. [Google Scholar] [CrossRef]
- Zheng, Y.; Pan, D. The Hippo signaling pathway in development and disease. Dev. Cell 2019, 50, 264–282. [Google Scholar] [CrossRef] [PubMed]
- Sidkey, B.A.J. Effect of plant growth regulators on secondary metabolites accumulation and antioxidant activity of Catharanthus roseus L. Int. J. Pharm. Sci. Res. 2020, 11, 241–245. [Google Scholar]
- Bienaimé, C.; Melin, A.; Bensaddek, L.; Attoumbré, J.; Nava-Saucedo, E.; Baltora-Rosset, S. Effects of plant growth regulators on cell growth and alkaloids production by cell cultures of Lycopodiella inundata. Plant Cell Tissue Org. 2015, 123, 523–533. [Google Scholar] [CrossRef]
- Sharma1, M.; Singh, A.K.; Salgotra, R.K.; Sharma, M.; Sharma, B.K.; Gupta, S.K.; Sharma, V. Optimization of conditions for callus induction from BC2F1 population (ranbir basmati x PAU148) through anther culture. Biotechnol. J. Int. 2019, 23, 1–7. [Google Scholar] [CrossRef]
- Effendi, S.R.N.; Rofiqoh, I. Embryogenesis Callus Induction of Carica pubescens Using Divine Smoke Particulates Containing Amino Acids. J. Phys. Conf. Ser. 2019, 1241, 012004. [Google Scholar]
- Nurwahyuni, I.; Gusvani, V. The role of cysteine in improving somatic embryos of salak Sidempuan (Salacca sumatrana Becc.). J. Phys. Conf. Ser. 2018, 1116, 052019. [Google Scholar]
- Jeong, J.; Jung, Y.; Na, S.; Jeong, J.; Lee, E.; Kim, M.S.; Choi, S.; Shin, D.H.; Paek, E.; Lee, H.Y.; et al. Novel oxidative modifications in redox-active cysteine residues. Mol. Cell Proteom. 2011, 10, M110.000513. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, S.; Wei, C.; Lin, H.; Zhang, P.; Liu, Y.; Deng, Y.; Huang, Q.; Xie, B. Cystathionine Gamma-Lyase Regulate Psilocybin Biosynthesis in Gymnopilus dilepis Mushroom via Amino Acid Metabolism Pathways. J. Fungi 2022, 8, 870. https://doi.org/10.3390/jof8080870
Yao S, Wei C, Lin H, Zhang P, Liu Y, Deng Y, Huang Q, Xie B. Cystathionine Gamma-Lyase Regulate Psilocybin Biosynthesis in Gymnopilus dilepis Mushroom via Amino Acid Metabolism Pathways. Journal of Fungi. 2022; 8(8):870. https://doi.org/10.3390/jof8080870
Chicago/Turabian StyleYao, Sen, Chuanzheng Wei, Hui Lin, Peng Zhang, Yuanyuan Liu, Youjin Deng, Qianhui Huang, and Baogui Xie. 2022. "Cystathionine Gamma-Lyase Regulate Psilocybin Biosynthesis in Gymnopilus dilepis Mushroom via Amino Acid Metabolism Pathways" Journal of Fungi 8, no. 8: 870. https://doi.org/10.3390/jof8080870