
The Biosynthesis of 1-octene-3-ol by a Multifunctional Fatty Acid Dioxygenase and Hydroperoxide Lyase in Agaricus bisporus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Plasmids
2.2. Bioinformatics Analysis
2.3. Gene Cloning, Expression, and Purification
2.4. Dioxygenase Activity Assay
2.5. HPL Activity Assay
2.6. Enzymatic Synthesis of 1-octene-3-ol from Linoleic Acid
2.7. Headspace Solid-Phase Micro-Extraction Gas Chromatography–Mass Spectrometry (HS-SPME-GC–MS) Analysis
2.8. Construction of Antisense AbLOX Transformants
2.9. Quantitative RT–PCR
2.10. Statistical Analysis
3. Results
3.1. Homology Analysis of 1-octen-3-ol Synthesis-Related Genes in Agaricus bisporus
3.2. Enzyme Activity of Prokaryotic-Expression Proteins
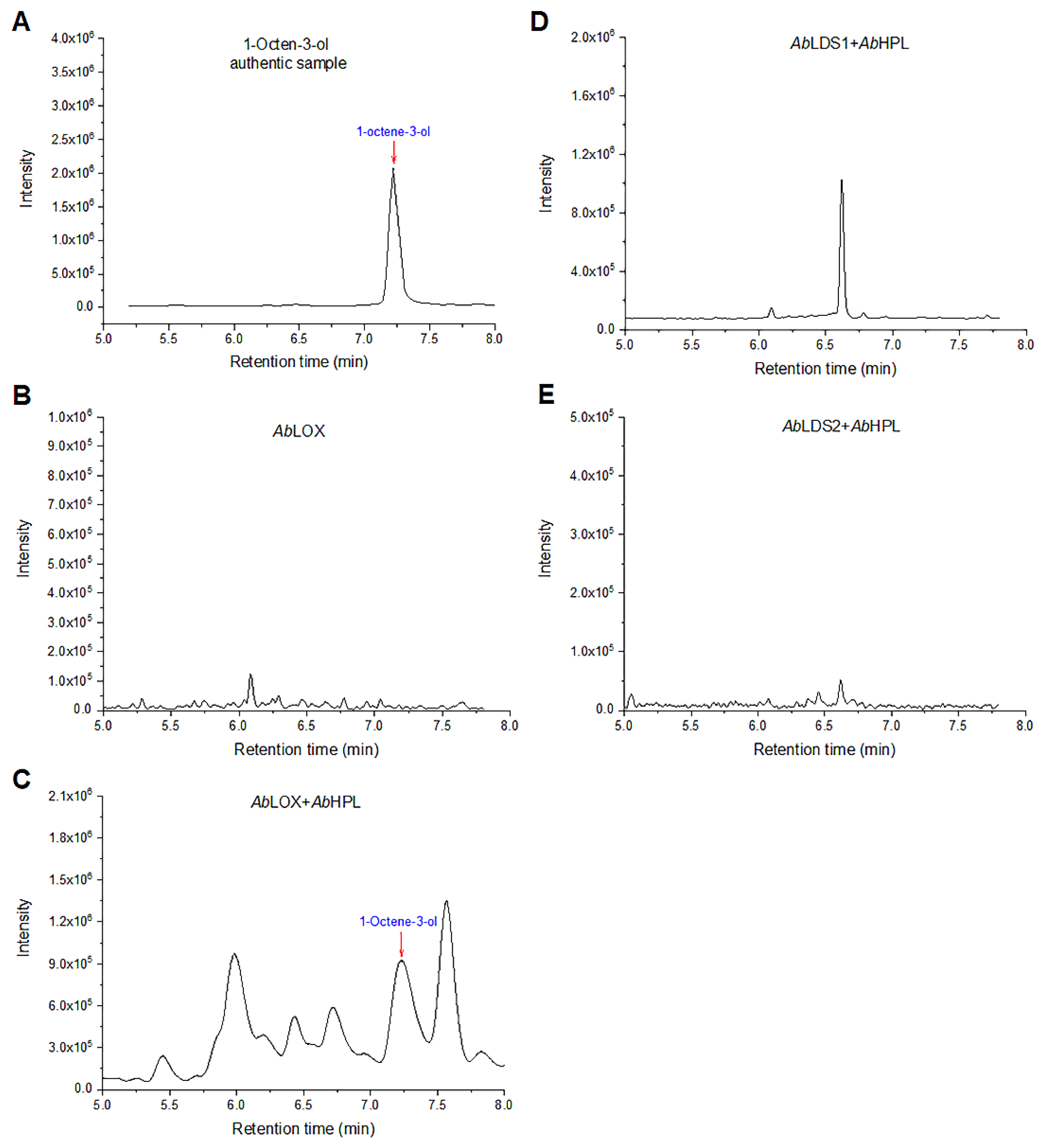
3.3. Analysis of the Volatile Products of Enzymatic Linoleic Acid Degradation by HS-SPME-GC–MS
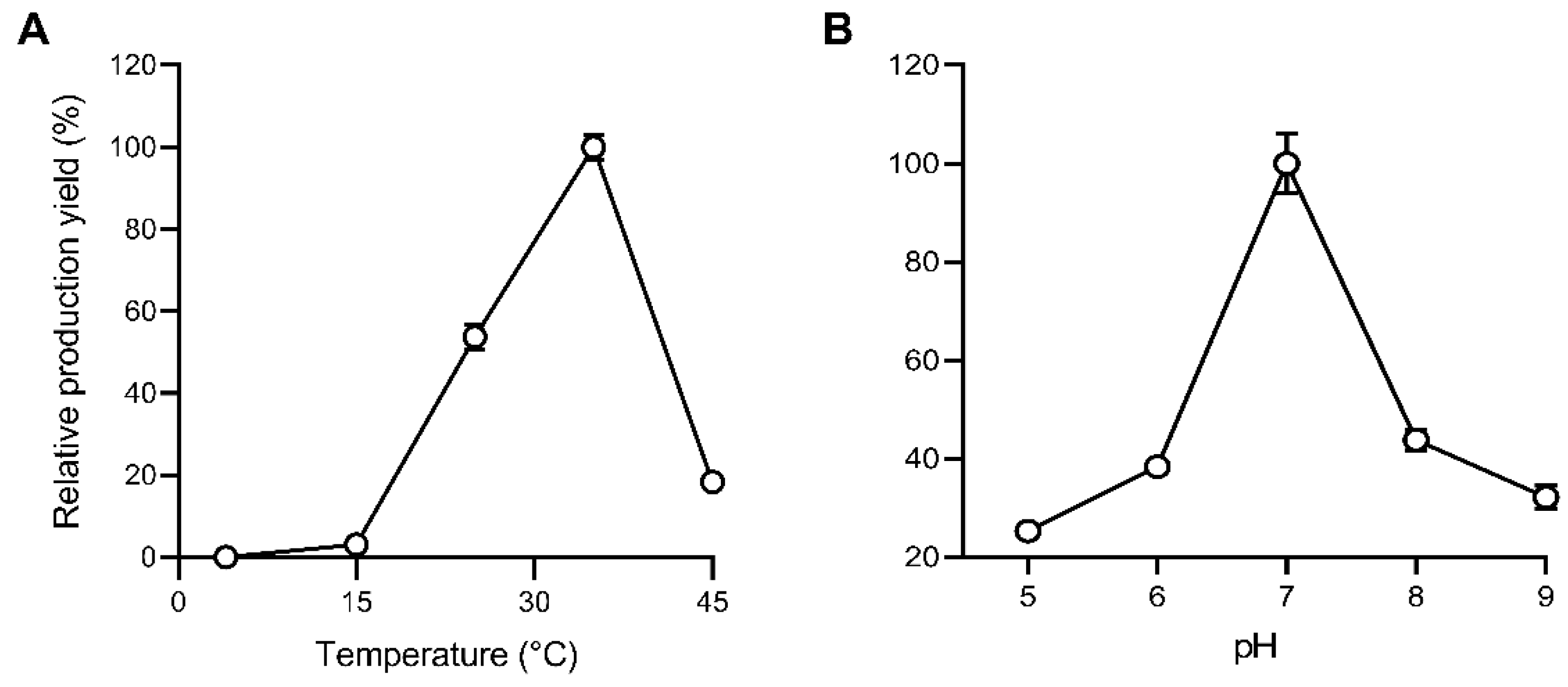
3.4. The Optimal Temperature and pH for the Enzymatic Synthesis of 1-octen-3-ol from Linoleic Acid
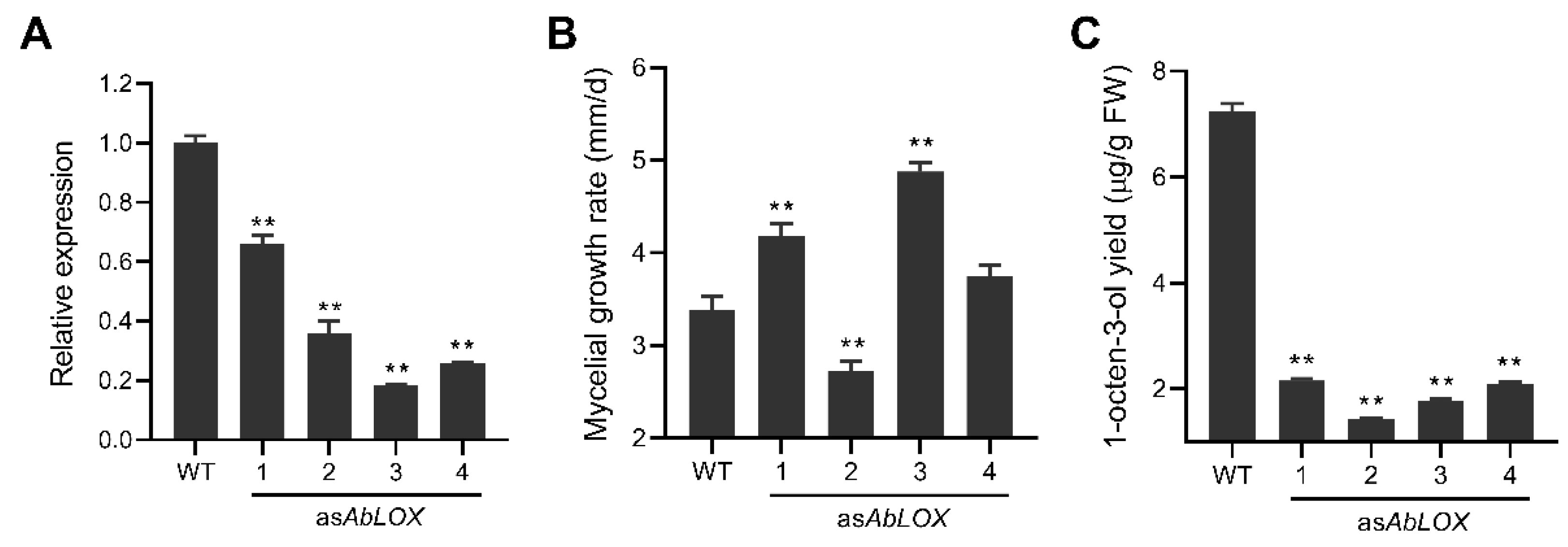
3.5. Effect of Reduced AbLOX Expression on Agaricus bisporus Mycelial Growth and the Production of 1-octen-3-ol
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murahashi, S. Über die riechstoffe des matsutake. Sci. Pap. Inst. Phys. Chem. Res. 1938, 34, 155–172. [Google Scholar]
- Maga, J.A. Mushroom flavor. J. Agric. Food Chem. 1981, 29, 1–4. [Google Scholar] [CrossRef]
- Wurzenberger, M.; Grosch, W. The formation of 1-octen-3-ol from the 10-hydroperoxide isomer of linoleic acid by a hydroperoxide lyase in mushrooms (Psalliota bispora). Biochim. Biophys. Acta BBA-Lipids Lipid Metab. 1984, 794, 25–30. [Google Scholar] [CrossRef]
- Dickschat, J.S. Fungal volatiles—A survey from edible mushrooms to moulds. Nat. Prod. Rep. 2017, 34, 310–328. [Google Scholar] [CrossRef]
- Hoffmann, G. 1-Octen-3-ol and its relation to other oxidative cleavage products from esters of linoleic acid. J. Am. Oil Chem. Soc. 1962, 39, 439–444. [Google Scholar] [CrossRef]
- Stark, W.; Forss, D.A. A compound responsible for mushroom flavour in dairy products. J. Dairy Res. 1964, 31, 253–259. [Google Scholar] [CrossRef]
- Bernhard, R.A.; Simone, M.J. The locus of aroma in the mushroom (Agaricus campestris L.). Food Res. 1958, 24, 165–166. [Google Scholar] [CrossRef]
- Wurzenberger, M.; Grosch, W. Determination of 1 octen-3-ol in mushrooms and in products containing mushrooms. Z Lebensm-Unters-Forsch 1983, 176, 16–19. [Google Scholar] [CrossRef]
- Kim, K.J.; Kim, Y.H. Optimization for effective bioproduction of natural (-)-1-octen-3-ol by lipoxygenase and hydroperoxide lyase from Agaricus bisporus. J. Korean Soc. Food Sci. Nutr. 2005, 34, 899–903. [Google Scholar]
- Zawirska-Wojtasiak, R. Optical purity of (R)-(−)-1-octen-3-ol in the aroma of various species of edible mushrooms. Food Chem. 2004, 86, 113–118. [Google Scholar] [CrossRef]
- Von Arx, M.; Schmidt-Büsser Dguerin, P.M. Plant volatiles enhance behavioral responses of grapevine moth males, Lobesia botrana to sex pheromone. J. Chem. Ecol. 2012, 38, 222–225. [Google Scholar] [CrossRef]
- Kües, U.; Khonsuntia, W.; Subba, S.; Dörnte, B. Volatiles in communication of Agaricomycetes. In Physiology and Genetics, 2nd ed.; Anke, T., Schuffler, A., Eds.; Springer: Cham, Switzerland, 2018; pp. 149–212. [Google Scholar]
- Holighaus, G.; Rohlfs, M. Volatile and non-volatile fungal oxylipins in fungus-invertebrate interactions. Fungal Ecol. 2019, 38, 28–36. [Google Scholar] [CrossRef]
- Gessler, N.N.; Filippovich, S.Y.; Bachurina, G.P.; Kharchenko, E.A.; Groza, N.V.; Belozerskaya, T.A. Oxylipins and oxylipin synthesis pathways in fungi. Appl. Biochem. Microbiol. 2017, 53, 628–639. [Google Scholar] [CrossRef]
- Mehmood, A.; Liu, G.; Wang, X.; Meng, G.; Wang, C.; Liu, Y. Fungal quorum-sensing molecules and inhibitors with potential antifungal activity, a review. Molecules 2019, 24, 1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noble, R.; Dobrovin-Pennington, A.; Hobbs, P.J.; Pederby, J.; Rodger, A. Volatile C8 compounds and pseudomonads influence primordium formation of Agaricus bisporus. Mycologia 2009, 101, 583–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eastwood, D.C.; Herman, B.; Noble, R.; Dobrovin-Pennington, A.; Sreenivasaprasad, S.; Burton, K.S. Environmental regulation of reproductive phase change in Agaricus bisporus by 1-octen-3-ol, temperature and CO2. Fungal Genet. Biol. 2013, 55, 54–66. [Google Scholar] [CrossRef]
- Lee, N.Y.; Choi, D.H.; Kim, M.G.; Jeong, M.J.; Kwon, H.J.; Kim, D.H.; Kim, Y.G.; di Luccio, E.; Arioka, M.; Yoon, H.J.; et al. Biosynthesis of (R)-(-)-1-octen-3-ol in recombinant Saccharomyces cerevisiae with lipoxygenase-1 and hydroperoxide lyase genes from Tricholoma matsutake. J. Microbiol. Biotechnol. 2020, 30, 296–305. [Google Scholar] [CrossRef]
- Mosandl, A.; Heusinger, G.; Gessner, M. Analytical and sensory differentiation of 1-octen-3-ol enantiomers. J. Agric. Food Chem. 1986, 34, 119–122. [Google Scholar] [CrossRef]
- Yin, G.; Zhang, Y.; Fu, M.; Hua, S.S.T.; Huang, Q.; Pennerman, K.K.; Wu, G.; Jurick, W.M.; Lee, S.; Bu, L.; et al. Influence of R and S enantiomers of 1-octen-3-ol on gene expression of Penicillium chrysogenum. J. Ind. Microbiol. Biotechnol. 2019, 46, 977–991. [Google Scholar] [CrossRef]
- Morawicki, R.O.; Beelman, R.B.; Peterson, D.; Ziegler, G. Biosynthesis of 1-octen-3-ol and 10-oxo-trans-8-decenoic acid using a crude homogenate of Agaricus bisporus, Optimization of the reaction, kinetic factors. Process Biochem. 2005, 40, 131–137. [Google Scholar] [CrossRef]
- Morawicki, R.O.; Beelman, R.B. Study of the biosynthesis of 1-octen-3-ol using a crude homogenate of Agaricus bisporus in a bioreactor. J. Food Sci. 2008, 73, C135–C139. [Google Scholar] [CrossRef] [PubMed]
- Oliw, E.H.; Aragó, M.; Chen, Y.; Jernerén, F. A new class of fatty acid allene oxide formed by the DOX-P450 fusion proteins of human and plant pathogenic fungi, C. immitis and Z. tritici. J. Lipid Res. 2016, 57, 1518–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wadman, M.W. Dioxygenation of Polyunsaturated Fatty Acids in Fungi. Ph.D. Thesis, Utrecht University, Utrecht, The Netherlands, 2007. [Google Scholar]
- Shibata, Y.; Matsui, K.; Kajiwara, T.; Hatanaka, A. Fatty acid hydroperoxide lyase is a heme protein. Biochem. Biophys. Res. Commun. 1995, 207, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Oliw, E.H. Fatty acid dioxygenase-cytochrome P450 fusion enzymes of filamentous fungal pathogens. Fungal Genet. Biol. 2021, 157, 103623. [Google Scholar] [CrossRef]
- Tressl, R.; Bahri, D.; Engel, K.H. Lipid oxidation in fruits and vegetables. ACS Symp. Ser. 1981, 170, 213–232. [Google Scholar]
- Wurzenberger, M.; Grosch, W. The enzymic oxidative breakdown of linoleic acid in mushrooms (Psalliota bispora). Z. Lebensm.-Unters.-Forsch. 1982, 175, 186–190. [Google Scholar] [CrossRef]
- Assaf, S.; Hadar, Y.; Dosoretz, C.G. 1-Octen-3-ol and 13-hydroperoxylinoleate are products of distinct pathways in the oxidative breakdown of linoleic acid by Pleurotus pulmonarius. Enzym. Microb. Technol. 1997, 21, 484–490. [Google Scholar] [CrossRef]
- Assaf, S.; Hadar, Y.; Dosoretz, C.G. Biosynthesis of 13-hydroperoxylinoleate, 10-oxo-8-decenoic acid, and 1-octen-3-ol from linoleic acid by a mycelial-pellet homogenate of Pleurotus pulmonarius. J. Agric. Food Chem. 1995, 43, 2173–2178. [Google Scholar] [CrossRef]
- Akakabe, Y.; Matsui, K.; Kajiwara, T. Stereochemical correlation between 10-hydroperoxyoctadecadienoic acid and 1-octen-3-ol in Lentinula edodes and Tricholoma matsutake mushrooms. Biosci. Biotechnol. Biochem. 2005, 69, 1539–1544. [Google Scholar] [CrossRef] [Green Version]
- Combet, E.; Henderson, J.; Eastwood, D.C.; Burton, K.S. Influence of sporophore development, damage, storage, and tissue specificity on the enzymic formation of volatiles in mushrooms (Agaricus bisporus). J. Agric. Food Chem. 2009, 57, 3709–3717. [Google Scholar] [CrossRef]
- Brodhun, F.; Schneider, S.; Göbel, C.; Hornung, E.; Feussner, I. PpoC from Aspergillus nidulans is a fusion protein with only one active haem. Biochem. J. 2010, 425, 553–565. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, R.; Watanabe, T.; Yano, S.; Mizutani, O.; Yamada, O.; Kasumi, T.; Ogihara, J. Aspergillus luchuensis fatty acid oxygenase ppoC is necessary for 1-octen-3-ol biosynthesis in rice koji. J. Biosci. Bioeng. 2020, 129, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, R.; Watanabe, T.; Hayashi, R.; Isogai, A.; Yamada, O.; Ogihara, J. Awamori fermentation test and 1-octen-3-ol productivity analysis using fatty acid oxygenase disruptants of Aspergillus luchuensis. J. Biosci. Bioeng. 2020, 130, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Stolterfoht, H.; Rinnofner, C.; Winkler, M.; Pichler, H. Recombinant lipoxygenases and hydroperoxide lyases for the synthesis of green leaf volatiles. J. Agric. Food Chem. 2019, 67, 13367–13392. [Google Scholar] [CrossRef]
- Brash, A.R.; Niraula, N.P.; Boeglin, W.E.; Mashhadi, Z. An ancient relative of cyclooxygenase in cyanobacteria is a linoleate 10S-dioxygenase that works in tandem with a catalase-related protein with specific 10S-hydroperoxide lyase activity. J. Biol. Chem. 2014, 289, 13101–13111. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.M.; Zhu, Z.J.; Chen, J.J.; Yang, R.; Luo, Q.J.; Xu, J.L.; Shan, H.; Yan, X.J. A multifunctional lipoxygenase from Pyropia haitanensis—The cloned and functioned complex eukaryotic algae oxylipin pathway enzyme. Algal Res. 2015, 12, 316–327. [Google Scholar] [CrossRef]
- Zhu, Z.J.; Chen, H.M.; Chen, J.J.; Yang, R.; Yan, X.J. One-step bioconversion of fatty acids into C8–C9 volatile aroma compounds by a multifunctional lipoxygenase cloned from Pyropia haitanensis. J. Agric. Food Chem. 2018, 66, 1233–1241. [Google Scholar] [CrossRef]
- Senger, T.; Wichard, T.; Kunze, S.; Göbel, C.; Lerchl, J.; Pohnert, G.; Feussner, I. A multifunctional lipoxygenase with fatty acid hydroperoxide cleaving activity from the moss Physcomitrella patens. J. Biol. Chem. 2005, 280, 7588–7596. [Google Scholar] [CrossRef] [Green Version]
- Ntoruru, J.M.; Ohnishi, T.; Katsumata, F.; Koeduka, T.; Matsui, K. 1-Octen-3-ol is formed from its primeveroside after mechanical wounding of soybean leaves. Plant Mol. Biol. 2022, 109, 551–561. [Google Scholar] [CrossRef]
- Matsui, K.; Takemoto, H.; Koeduka, T.; Ohnishi, T. 1-Octen-3-ol is formed from its glycoside during processing of soybean [Glycine max (L.) Merr.] seeds. J. Agric. Food Chem. 2018, 66, 7409–7416. [Google Scholar] [CrossRef]
- Wang, X.; Ding, Y.; Gao, X.; Liu, H.; Zhao, K.; Gao, Y.; Qiu, L. Promotion of the growth and plant biomass degrading enzymes production in solid-state cultures of Lentinula edodes expressing Vitreoscilla hemoglobin gene. J. Biotechnol. 2019, 302, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Morin, E.; Kohler, A.; Baker, A.R.; Foulongne-Oriol, M.; Lombard, V.; Nagye, L.G.; Ohm, R.A.; Patyshakuliyeva, A.; Brun, A.; Aerts, A.L.; et al. Genome sequence of the button mushroom Agaricus bisporus reveals mechanisms governing adaptation to a humic-rich ecological niche. Proc. Natl. Acad. Sci. USA 2012, 109, 17501–17506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Zeng, X.; Yang, Y.L.; Xing, Y.M.; Zhang, Q.; Li, J.M.; Ma, K.; Liu, H.W.; Guo, S.X. Genomic and transcriptomic analyses reveal differential regulation of diverse terpenoid and polyketides secondary metabolites in Hericium erinaceus. Sci. Rep. 2017, 7, 10151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Zhang, J.; Gao, X.; Chen, J.; Zheng, Y.; Gao, Y.; Qiu, L. The molecular mechanism for the ethylene regulation of postharvest button mushrooms maturation and senescence. Postharvest Biol. Technol. 2019, 156, 110930. [Google Scholar] [CrossRef]
- Williams, D.C.; Lim, M.H.; Chen, A.O.; Pangborn, R.M.; Whitaker, J.R. Blanching of vegetables for freezing—Which indicator enzyme to choose. Food Technol. 1986, 40, 130–140. [Google Scholar]
- Satoh, T.; Matsuda, Y.; Takashio, M.; Satoh, K.; Beppu, T.; Arima, K. Isolation of lipoxygenase-like enzyme from Fusarium oxysporum. Agric. Biol. Chem. 1976, 40, 953–961. [Google Scholar] [CrossRef]
- Husson, F.; Bompas, D.; Kermasha, S. Biogeneration of 1-octen-3-ol by lipoxygenase and hydroperoxide lyase activities of Agaricus bisporus. Process Biochem. 2001, 37, 177–182. [Google Scholar] [CrossRef]
- Chen, X.; Stone, M.; Schlagnhaufer, C.; Romaine, C.P. A fruiting body tissue method for efficient Agrobacterium-mediated transformation of Agaricus bisporus. Appl. Environ. Microbiol. 2000, 66, 4510–4513. [Google Scholar] [CrossRef] [Green Version]
- Plagemann, I.; Zelena, K.; Arendt, P.; Ringel, P.D.; Krings, U.; Berger, R.G. LOXPsa1, the first recombinant lipoxygenase from a basidiomycete fungus. J. Mol. Catal. B Enzym. 2013, 87, 99–104. [Google Scholar] [CrossRef]
- Krahe, N.K.; Berger, R.G.; Kahlert, L.; Ersoy, F. Co-oxidative transformation of piperine to piperonal and 3, 4-methylenedioxycinnamaldehyde by a lipoxygenase from Pleurotus sapidus. ChemBioChem 2021, 22, 2857–2861. [Google Scholar] [CrossRef]
- Lee, D.S.; Nioche, P.; Hamberg, M.; Raman, C.S. Structural insights into the evolutionary paths of oxylipin biosynthetic enzymes. Nature 2008, 455, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Teder, T.; Lõhelaid, H.; Boeglin, W.E.; Calcutt, W.M.; Brash, A.R.; Samel, N. A catalase-related hemoprotein in coral is specialized for synthesis of short-chain aldehydes, Discovery of P450-type hydroperoxide lyase activity in a catalase. J. Biol. Chem. 2015, 290, 19823–19832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorina, S.S.; Toporkova, Y.Y.; Mukhtarova, L.S.; Grechkin, A.N. The CYP443C1 (CYP74 clan) cytochrome of sea anemone Nematostella vectensi—The first metazoan enzyme possessing hydroperoxide lyase/epoxyalcohol synthase activity. Dokl. Biochem. Biophys. 2019, 486, 192–196. [Google Scholar] [CrossRef]
- Skaggs, B.A.; Alexander, J.F.; Pierson, C.A.; Schweitzer, K.S.; Chun, K.T.; Koegel, C.; Barbuch, R.; Bard, M. Cloning and characterization of the Saccharomyces cerevisiae C-22 sterol desaturase gene, encoding a second cytochrome P-450 involved in ergosterol biosynthesis. Gene 1996, 169, 105–109. [Google Scholar] [CrossRef]
- Cruz, C.; Noël-Suberville, C.; Montury, M. Fatty acid content and some flavor compound release in two strains of Agaricus bisporus, according to three stages of development. J. Agric. Food Chem. 1997, 45, 64–67. [Google Scholar] [CrossRef]
- Feng, T.; Yang, M.; Ma, B.; Zhao, Y.; Zhuang, H.; Zhang, J.; Chen, D. Volatile profiles of two genotype Agaricus bisporus species at different growth stages. Food Res. Int. 2021, 140, 109761. [Google Scholar] [CrossRef] [PubMed]
- Mau, J.L.; Hwang, S.J. Volatile flavor compounds of mushroom mycelium. Food Sci. Agric. Chem. 1999, 1, 148–153. [Google Scholar]
- Chaisaena, W. Light Effects on Fruiting Body Development of Wildtype in Comparison to Light-Insensitive Mutant Strains of the Basidiomycete Coprinopsis cinerea, Grazing of Mites (Tyrophagus putrescentiae) on the Strains and Production of Volatile Organic Compounds during Fruiting Body Development. Ph.D. Thesis, University of Goettingen, Goettingen, Germany, 2009. [Google Scholar]
- Stajich, J.E.; Wilke, S.K.; Ahrén, D.; Au, C.H.; Birren, B.W.; Borodovsky, M.; Burns, C.; Canbäck, B.; Casselton, L.A.; Cheng, C.K.; et al. Insights into evolution of multicellular fungi from the assembled chromosomes of the mushroom Coprinopsis cinerea (Coprinus cinereus). Proc. Natl. Acad. Sci. USA 2010, 107, 11889–11894. [Google Scholar] [CrossRef] [Green Version]
- Agger, S.; Lopez-Gallego, F.; Schmidt-Dannert, C. Diversity of sesquiterpene synthases in the basidiomycete Coprinus cinereus. Mol. Microbiol. 2009, 72, 1181–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orban, A.; Hennicke, F.; Rühl, M. Volatilomes of Cyclocybe aegerita during different stages of monokaryotic and dikaryotic fruiting. Biol. Chem. 2020, 401, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Xin, G.; Hou, Z.; Zhao, X.; Xu, H.; Bao, X.; Xia, R.; Li, Y.; Li, L. Biosynthetic mechanism of key volatile biomarkers of harvested Lentinula edodes triggered by spore release. J. Agric. Food Chem. 2021, 69, 9350–9361. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Enzyme | Specific Activity (U/mg of Protein) |
|---|---|
| AbLOX | 55.9 ± 2.4 |
| AbLDS1 | 51.7 ± 1.5 |
| AbLDS2 | 67.5 ± 3.0 |
| AbHPL | 8.1 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, T.; Chen, Y.; Liu, H.; Gao, Y.; Guo, J.; Li, Y.; Qi, Y.; Qiu, L. The Biosynthesis of 1-octene-3-ol by a Multifunctional Fatty Acid Dioxygenase and Hydroperoxide Lyase in Agaricus bisporus. J. Fungi 2022, 8, 827. https://doi.org/10.3390/jof8080827
Su T, Chen Y, Liu H, Gao Y, Guo J, Li Y, Qi Y, Qiu L. The Biosynthesis of 1-octene-3-ol by a Multifunctional Fatty Acid Dioxygenase and Hydroperoxide Lyase in Agaricus bisporus. Journal of Fungi. 2022; 8(8):827. https://doi.org/10.3390/jof8080827
Chicago/Turabian StyleSu, Tongfu, Yuannan Chen, Haohao Liu, Yuqian Gao, Jiawen Guo, Yanan Li, Yuancheng Qi, and Liyou Qiu. 2022. "The Biosynthesis of 1-octene-3-ol by a Multifunctional Fatty Acid Dioxygenase and Hydroperoxide Lyase in Agaricus bisporus" Journal of Fungi 8, no. 8: 827. https://doi.org/10.3390/jof8080827