
Symbiotic Fungi Alter the Acquisition of Phosphorus in Camellia oleifera through Regulating Root Architecture, Plant Phosphate Transporter Gene Expressions and Soil Phosphatase Activities


 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Symbiotic Fungal Inoculums
2.2. Plant Culture
2.3. Experimental Design
2.4. Determinations of Variables
2.5. Statistical Analysis
3. Results
3.1. Changes in Fungal Colonization
3.2. Growth Responses under Different Fungal Inoculations
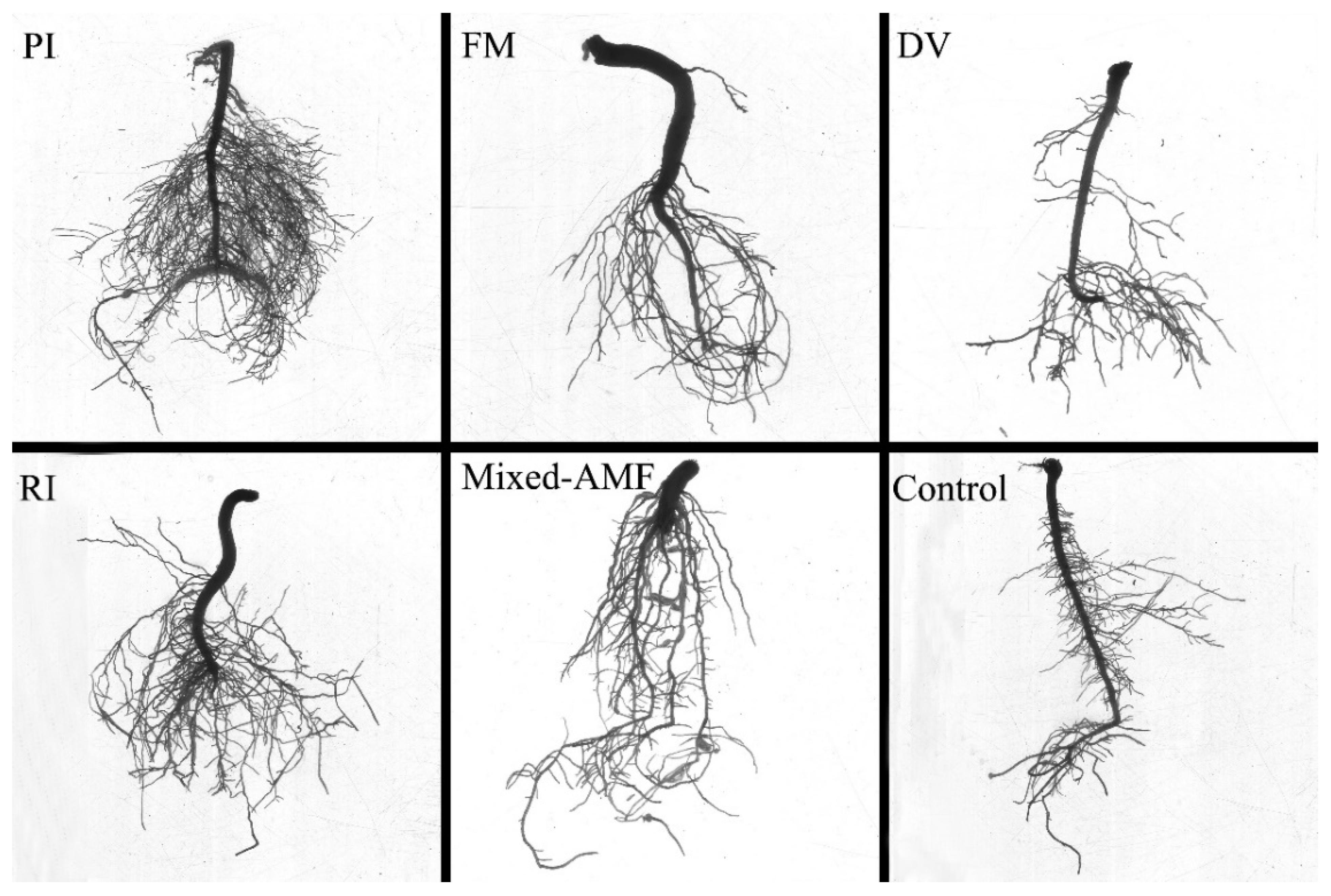
3.3. Root Architecture Responses under Different Fungal Inoculations
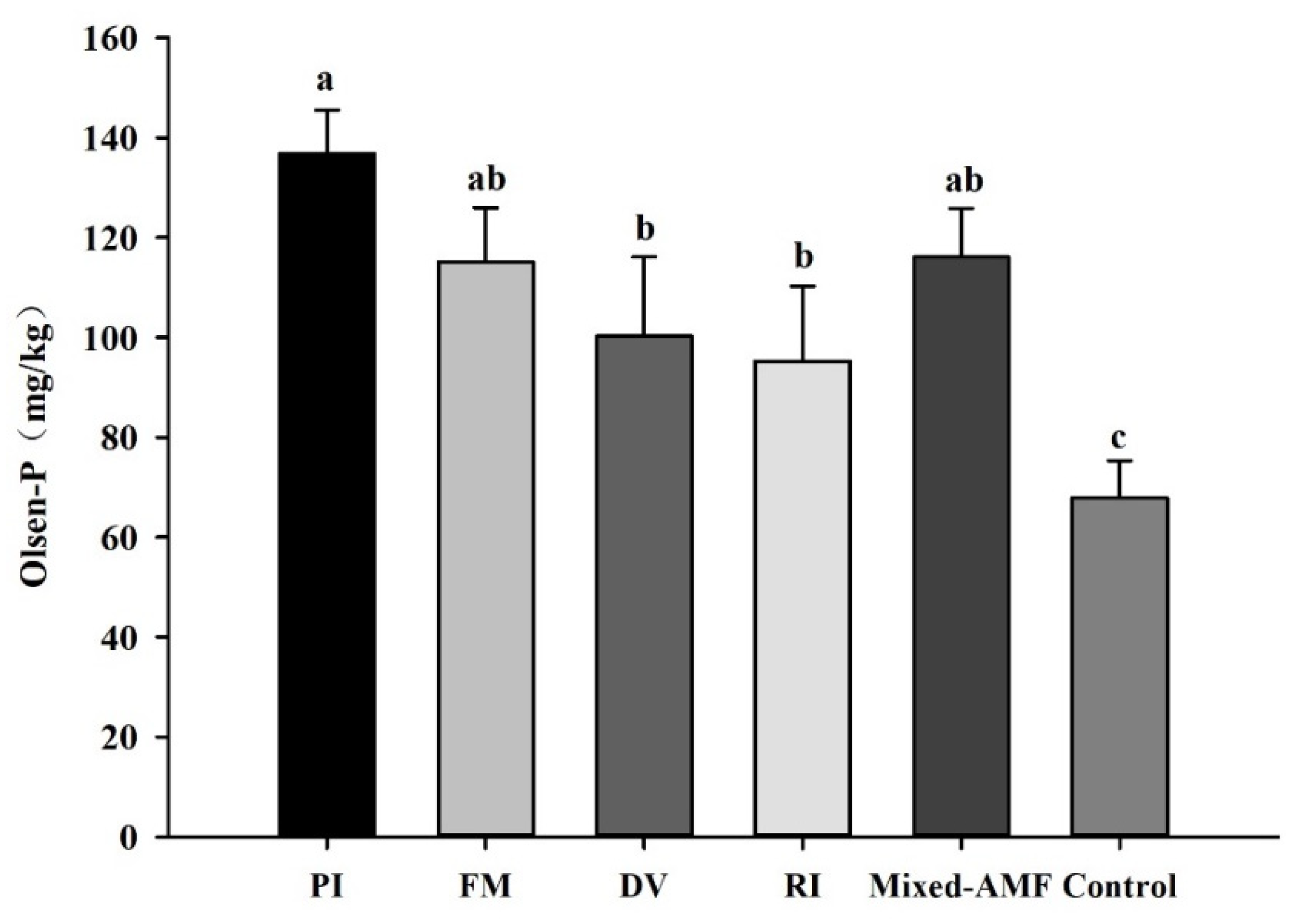
3.4. Soil Olsen-P Responses under Different Fungal Inoculations
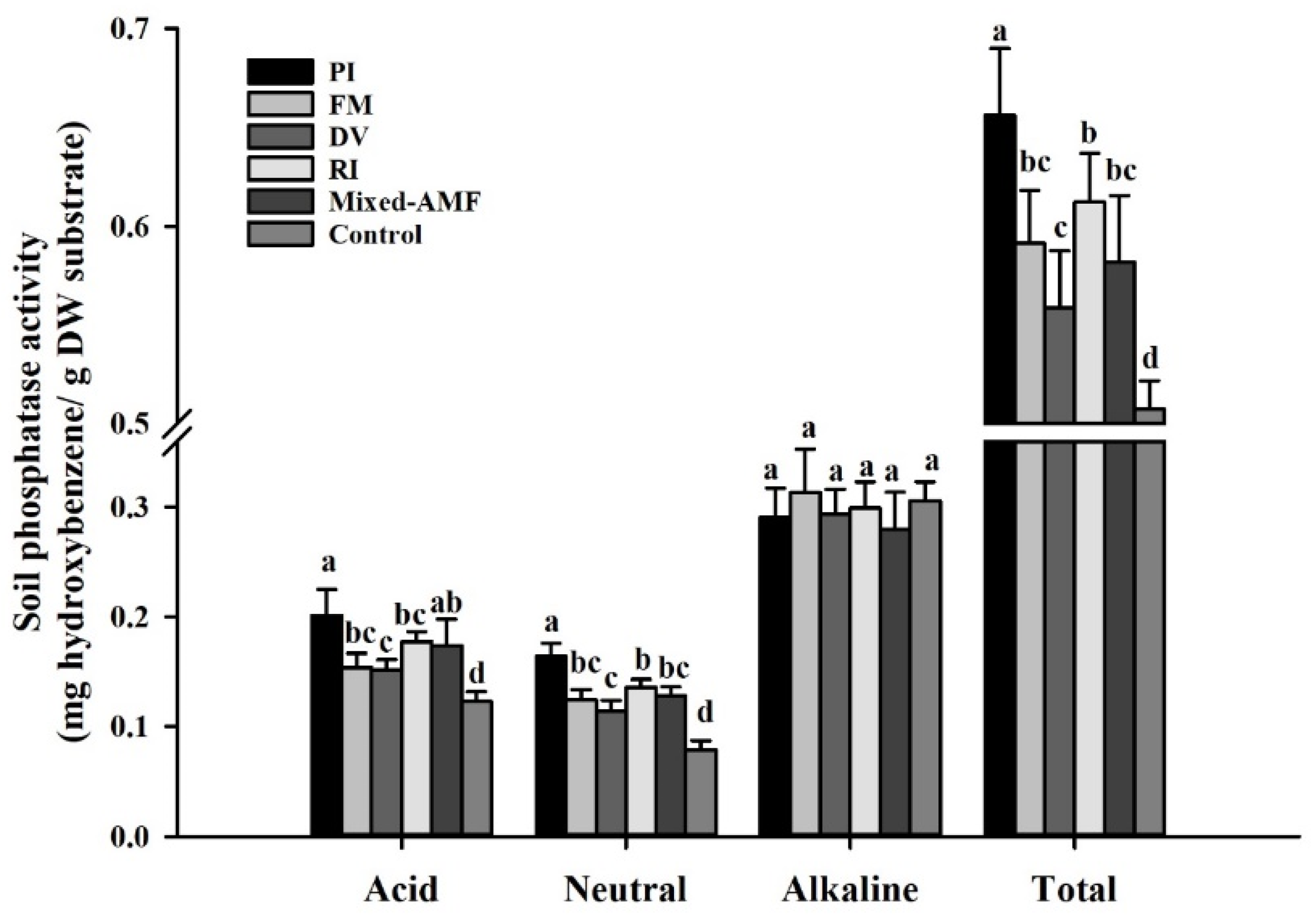
3.5. Soil Phosphatase Responses under Different Fungal Inoculations
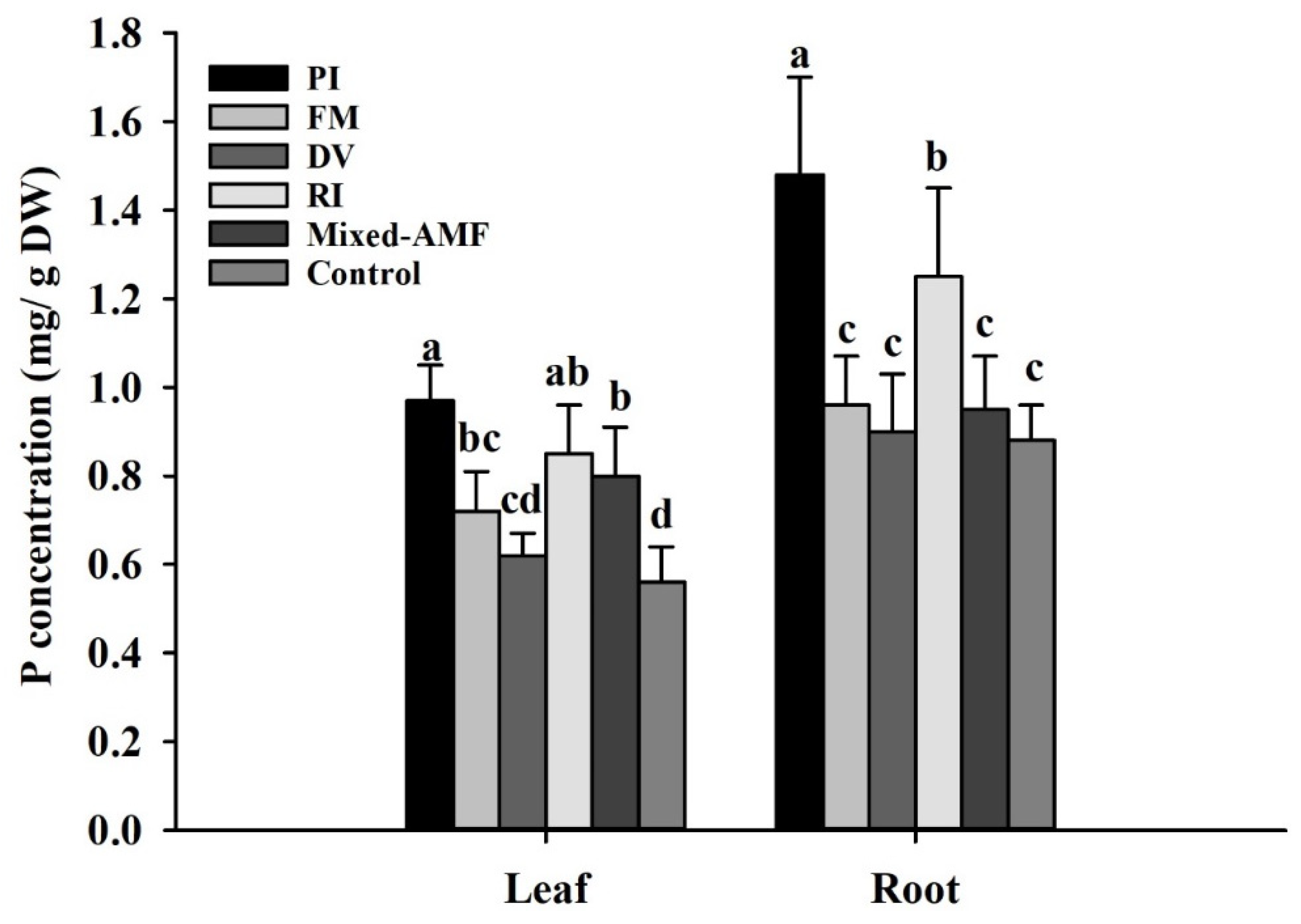
3.6. Plant P Responses under Different Fungal Inoculations
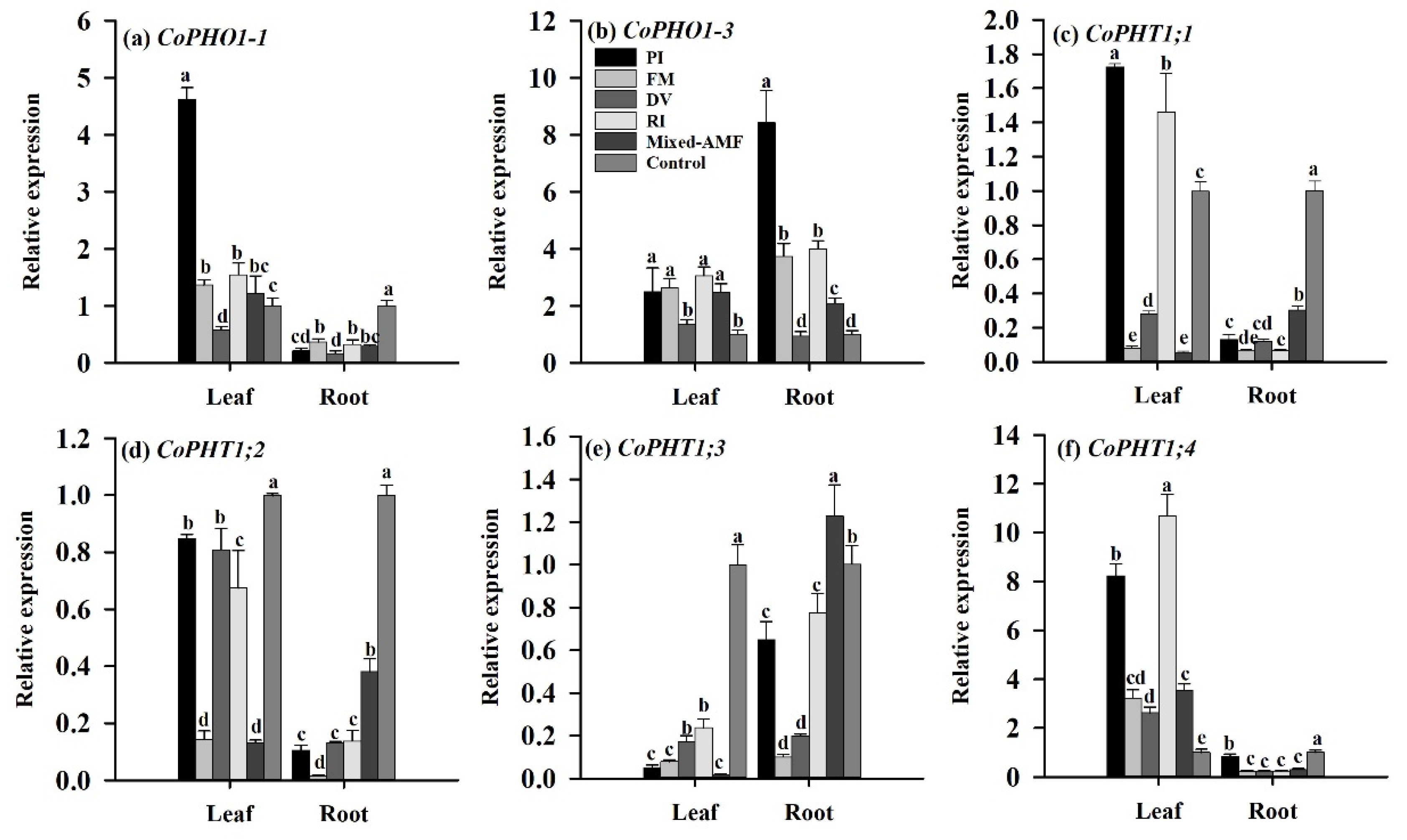
3.7. PT Gene Expression Responses under Different Fungal Inoculations
3.8. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, S.E.; Jakobsen, I.; Gronlund, M.; Smith, F.A. Roles of arbuscular mycorrhizas in plant phosphorus nutrition: Interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol. 2011, 156, 1050–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Lv, S.L.; Jiang, P.; Li, Y.X. Roles, regulation, and agricultural application of plant phosphate transporters. Front. Plant Sci. 2017, 8, 817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.X.; Hoffland, E.; Feng, G.; Kuyper, T.W. Arbuscular mycorrhizal symbiosis increases phosphorus uptake and productivity of mixtures of maize varieties compared to monocultures. J. Appl. Ecol. 2020, 57, 2203–2211. [Google Scholar] [CrossRef]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, G.H.; Luo, X.Z.; Hou, M.H.; Zheng, M.H.; Zhang, L.L.; He, X.J.; Shen, W.J.; Wen, D.Z. Mycorrhizal fungi and phosphatase involvement in rhizosphere phosphorus transformations improves plant nutrition during subtropical forest succession. Soil Biol. Biochem. 2021, 153, 108099. [Google Scholar] [CrossRef]
- Jiang, F.Y.; Zhang, L.; Zhou, J.C.; George, T.S.; Feng, G. Arbuscular mycorrhizal fungi enhance mineralization of organic phosphorus (P) by carrying bacteria along their extraradical hyphae. New Phytol. 2020, 230, 304–315. [Google Scholar] [CrossRef]
- Yang, L.; Zou, Y.N.; Tian, Z.H.; Wu, Q.S.; Kuča, K. Effects of beneficial endophytic fungal inoculants on plant growth and nutrient absorption of trifoliate orange seedlings. Sci. Hortic. 2021, 277, 109815. [Google Scholar] [CrossRef]
- Chen, E.J.; Liao, H.X.; Chen, B.M.; Peng, S.L. Arbuscular mycorrhizal fungi are a double-edged sword in plant invasion controlled by phosphorus concentration. New Phytol. 2020, 226, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Standish, R.J.; Daws, M.I.; Moral, T.K.; Speijers, J.; Koch, J.M.; Hobbs, R.J.; Tibbett, M. Phosphorus supply affects seedling growth of mycorrhizal but not cluster-root forming jarrah-forest species. Plant Soil 2022, 472, 577–594. [Google Scholar] [CrossRef]
- Cheng, X.F.; Xie, M.M.; Li, Y.; Liu, B.Y.; Liu, C.Y.; Wu, Q.S.; Kuča, K. Effects of field inoculation with arbuscular mycorrhizal fungi and endophytic fungi on fruit quality and soil properties of Newhall navel orange. Appl. Soil Ecol. 2022, 170, 104308. [Google Scholar] [CrossRef]
- Bonfante, P. The future has roots in the past: The ideas and scientists that shaped mycorrhizal research. New Phytol. 2018, 220, 982–995. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.C.; Gao, W.Q.; Srivastava, A.K.; Zou, Y.N.; Kuča, K.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.S. Differential effects of exogenous glomalin-related soil proteins on plant growth of trifoliate orange through regulating auxin changes. Front. Plant Sci. 2021, 12, 745402. [Google Scholar] [CrossRef]
- Lǚ, L.H.; Zou, Y.N.; Wu, Q.S. Mycorrhizas mitigate soil replant disease of peach through regulating root exudates, soil microbial population, and soil aggregate stability. Commun. Soil Sci. Plant Anal. 2019, 50, 909–921. [Google Scholar] [CrossRef]
- Ding, Y.E.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Mycorrhizal fungi regulate daily rhythm of circadian clock in trifoliate orange under drought stress. Tree Physiol. 2022, 42, 616–628. [Google Scholar] [CrossRef]
- Wu, M.Y.; Wei, Q.; Xu, L.; Li, H.Z.; Oelmüller, R.; Zhang, W.Y. Piriformospora indica enhances phosphorus absorption by stimulating acid phosphatase activities and organic acid accumulation in Brassica napus. Plant Soil 2018, 432, 333–344. [Google Scholar] [CrossRef]
- He, J.D.; Chi, G.G.; Zou, Y.N.; Shu, B.; Wu, Q.S.; Srivastava, A.K.; Kuča, K. Contribution of glomalin-related soil proteins to soil organic carbon in trifoliate orange. Appl. Soil Ecol. 2020, 154, 103592. [Google Scholar] [CrossRef]
- Abdelrahman, M.; EI-Sayed, M.A.; Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Burritt, D.J.; Tran, L.S.P. Metabolomics and transcriptomics in legumes under phosphate deficiency in relation to nitrogen fixation by root nodules. Front. Plant Sci. 2018, 9, 922. [Google Scholar] [CrossRef] [Green Version]
- Prathap, V.; Kumar, A.; Maheshwari, C.; Tyagi, A. Phosphorus homeostasis: Acquisition, sensing, and long-distance signaling in plants. Mol. Biol. Rep. 2022. [Google Scholar] [CrossRef]
- Wege, S.; Khan, G.A.; Jung, J.Y.; Vogiatzaki, E.; Pradervand, S.; Aller, I.; Meyer, A.J.; Poirier, Y. The EXS domain of PHO1 participates in the response of shoots to phosphate deficiency via a root-to-shoot signal. Plant Physiol. 2015, 170, 385–400. [Google Scholar] [CrossRef] [Green Version]
- Sahodaran, N.K.; Arun, A.K.; Ray, J.G. Native arbuscular mycorrhizal fungal isolates (Funneliformis mosseae and Glomus microcarpum) improve plant height and nutritional status of banana plants. Exp. Agr. 2019, 55, 924–933. [Google Scholar] [CrossRef]
- Campos, P.M.S.; Borie, F.; Cornejo, P.; Meier, S.; López-Ráez, J.A.; López-Garcia, Á.; Seguel, A. Wheat root trait plasticity, nutrient acquisition and growth responses are dependent on specific arbuscular mycorrhizal fungus and plant genotype interactions. J. Plant. Physiol. 2021, 256, 153297. [Google Scholar] [CrossRef] [PubMed]
- Mohandas, S. Arbuscular mycorrhizal fungi benefit mango (Mangifera indica L.) plant growth in the field. Sci. Hortic. 2012, 143, 43–48. [Google Scholar] [CrossRef]
- Wu, F.; Li, J.R.; Chen, Y.L.; Zhang, L.P.; Zhang, Y.; Wang, S.; Shi, X.; Li, L.; Liang, J.S. Effects of phosphate solubilizing bacteria on the growth, photosynthesis, and nutrient uptake of Camellia oleifera Abel. Forests 2019, 10, 348. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.C.; Xiao, Z.Y.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.S. Mycorrhizal fungal diversity and its relationship with soil properties in Camellia oleifera. Agriculture 2021, 11, 470. [Google Scholar] [CrossRef]
- Wang, D.X.; Chen, G.C.; Jiang, Z.P. Effects of arbuscular mycorrhiza on the growth and absorption of nitrogen and phosphorus in Camellia oleifera seedlings. China For. Sci. Technol. 2010, 24, 23–26. [Google Scholar]
- Wu, F.; Li, Z.Y.; Lin, Y.L.; Zhang, L.P. Effects of Funneliformis mosseae on the utilization of organic phosphorus in Camellia oleifera Abel. Can. J. Microbiol. 2021, 65, 349–357. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Br. Mycol. Soc. Trans. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N.; He, X.H. Differences of hyphal and soil phosphatase activities in drought-stressed mycorrhizal trifoliate orange (Poncirus trifoliata) seedlings. Sci. Hortic. 2011, 129, 294–298. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zou, Y.N.; Zhang, F.; Srivastava, A.K.; Wu, Q.S.; Kuča, K. Arbuscular mycorrhizal fungi regulate polyamine homeostasis in roots of trifoliate orange for improved adaptation to soil moisture deficit stress. Front. Plant Sci. 2021, 11, 600792. [Google Scholar] [CrossRef]
- Cao, M.A.; Wang, P.; Hashem, A.; Wirth, S.; Abd_Allah, E.F.; Wu, Q.S. Field inoculation of arbuscular mycorrhizal fungi improves fruit quality and root physiological activity of citrus. Agriculture 2021, 11, 1297. [Google Scholar] [CrossRef]
- Standish, R.J.; Albornoz, F.E.; Morald, T.K.; Hobbs, R.J.; Tibbett, M. Mycorrhizal symbiosis and phosphorus supply determine interactions among plants with contrasting nutrient-acquisition strategies. J. Ecol. 2021, 109, 3892–3902. [Google Scholar] [CrossRef]
- Wen, Z.H.; Li, H.B.; Shen, Q.; Tang, X.M.; Xiong, C.Y.; Li, H.G.; Pang, J.Y.; Ryan, M.H.; Lambers, H.; Shen, J.B. Tradeoffs among root morphology, exudation and mycorrhizal symbioses for phosphorus-acquisition strategies of 16 crop species. New Phytol. 2019, 223, 882–895. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, P.; Zou, Y.N.; Wu, Q.S. Effects of mycorrhizal fungi on root-hair growth and hormone levels of taproot and lateral roots in trifoliate orange under drought stress. Arch. Agron. Soil Sci. 2019, 65, 1316–1330. [Google Scholar] [CrossRef]
- Chenchouni, H.; Mekahlia, M.N.; Beddiar, A. Effect of inoculation with native and commercial arbuscular mycorrhizal fungi on growth and mycorrhizal colonization of olive (Olea europaea L.). Sci. Hortic. 2020, 261, 108969. [Google Scholar] [CrossRef]
- Liu, C.Y.; Wang, P.; Zhang, D.J.; Zou, Y.N.; Kuča, K.; Wu, Q.S. Mycorrhiza-induced change in root hair growth is associated with IAA accumulation and expression of EXPs in trifoliate orange under two P levels. Sci. Hortic. 2018, 234, 227–235. [Google Scholar] [CrossRef]
- Shao, Y.D.; Hu, X.C.; Wu, Q.S.; Yang, T.Y.; Srivastava, A.K.; Zhang, D.J.; Gao, X.B.; Kuča, K. Mycorrhizas promote P acquisition of tea plants through changes in root morphology and P transporter gene expression. S. Afr. J. Bot. 2021, 137, 455–462. [Google Scholar] [CrossRef]
- Feng, G.; Song, Y.C.; Li, X.L.; Christie, P. Contribution of arbuscular mycorrhizal fungi to utilization of organic sources of phosphorus by red clover in a calcareous soil. Appl. Soil Ecol. 2003, 22, 139–148. [Google Scholar] [CrossRef]
- Shu, B.; Xia, R.X.; Wang, P. Differential regulation of Pht1 phosphate transporters from trifoliate orange (Poncirus trifoliata L. Raf) seedlings. Sci. Hortic. 2012, 146, 115–123. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Names | Accessions | Sequences (5′→3′) |
|---|---|---|
| CoPHT1;1 | JX403969 | F: GTTCTTGGCGGAGTCAATTTC |
| R: CATCCTCATCTTCCTCGTTCTC | ||
| CoPHT1;2 | JX412956 | F: TCCCTTTGCTTCTTCCGATTT |
| R: CGAGTCCTCTTGTTGGCATATT | ||
| CoPHT1;3 | KF989483 | F: GAGTCAGAGCAGCAGAAAGTAG |
| R: TGTAGTCCCAAGCAAGTGAAG | ||
| CoPHT1;4 | KF989484 | F: CCGTTACACCGCCCTTATC |
| R: CTGGGTTCTTCAGCCATCTT | ||
| CoPHO1-1 | KU161157 | F: AGCAGCCCTTGAAGTCATTAG |
| R: GAACTTGCCCGCATTGTTTAG | ||
| CoPHO1-3 | KU161156 | F: GAGCTTTCAGTGGCCTAACA |
| R: GTCGCCTCACCGAGTTTATC | ||
| EF-1α | KC337050 | F: AGACTGTGGCTGTTGGTGTT |
| R: ATCCAAACCCGCACAGTTCA |
| Treatments | Fungal Colonization Rate (%) | Height (cm) | Stem Diameter (mm) | Leaf Number (#/Plant) | Plant (Shoot + Root) Biomass (g/plant) |
|---|---|---|---|---|---|
| PI | 75 ± 7 a | 19.50 ± 3.70 a | 3.64 ± 0.40 a | 11.0 ± 1.1 a | 5.23 ± 0.55 a |
| FM | 54 ± 5 c | 11.92 ± 0.95 b | 2.50 ± 0.68 bc | 6.5 ± 1.0 b | 2.71 ± 0.38 bc |
| DV | 27 ± 4 d | 9.50 ± 1.03 c | 1.94 ± 0.46 cd | 5.0 ± 0.9 c | 2.19 ± 0.29 d |
| RI | 67 ± 7 ab | 14.07 ± 1.79 b | 2.64 ± 0.43 b | 7.7 ± 1.6 b | 3.18 ± 0.39 b |
| Mixed-AMF | 60 ± 8 b | 12.75 ± 1.17 b | 2.63 ± 0.65 b | 6.7 ± 1.2 b | 2.83 ± 0.40 b |
| Control | 0 ± 0 e | 9.60 ± 0.74 c | 1.85 ± 0.15 d | 4.8 ± 0.7 c | 2.30 ± 0.23 cd |
| Treatments | Length (cm) | Surface Area (cm2) | Average Diameter (mm) | Volume (cm3) |
|---|---|---|---|---|
| PI | 138.76 ± 14.80 a | 13.76 ± 1.35 a | 0.62 ± 0.05 b | 0.96 ± 0.12 a |
| FM | 65.50 ± 8.33 bc | 10.52 ± 0.65 b | 0.72 ± 0.11 b | 0.40 ± 0.04 c |
| DV | 41.38 ± 4.78 d | 7.51 ± 1.13 c | 0.95 ± 0.10 a | 0.23 ± 0.03 d |
| RI | 130.93 ± 17.80 a | 13.11 ± 1.05 a | 0.64 ± 0.07 b | 0.63 ± 0.07 b |
| Mixed-AMF | 74.44 ± 8.02 b | 11.62 ± 0.88 b | 0.68 ± 0.04 b | 0.34 ± 0.02 c |
| Control | 54.26 ± 6.15 cd | 8.44 ± 0.82 c | 0.89 ± 0.07 a | 0.25 ± 0.03 d |
| Fungal Colonization | Root Architecture | Soil Phosphatase | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Total Length | Surface Area | Average Diameter | Volume | Acid | Neutral | Alkaline | Total | ||
| Leaf P | 0.94 ** | 0.91 ** | 0.95 ** | −0.74 | 0.91 * | 0.97 ** | 0.95 ** | −0.42 | 0.95 ** |
| Root P | 0.73 | 0.95 ** | 0.85 * | −0.91 * | 0.99 ** | 0.85 * | 0.84 * | −0.21 | 0.87 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, M.-A.; Liu, R.-C.; Xiao, Z.-Y.; Hashem, A.; Abd_Allah, E.F.; Alsayed, M.F.; Harsonowati, W.; Wu, Q.-S. Symbiotic Fungi Alter the Acquisition of Phosphorus in Camellia oleifera through Regulating Root Architecture, Plant Phosphate Transporter Gene Expressions and Soil Phosphatase Activities. J. Fungi 2022, 8, 800. https://doi.org/10.3390/jof8080800
Cao M-A, Liu R-C, Xiao Z-Y, Hashem A, Abd_Allah EF, Alsayed MF, Harsonowati W, Wu Q-S. Symbiotic Fungi Alter the Acquisition of Phosphorus in Camellia oleifera through Regulating Root Architecture, Plant Phosphate Transporter Gene Expressions and Soil Phosphatase Activities. Journal of Fungi. 2022; 8(8):800. https://doi.org/10.3390/jof8080800
Chicago/Turabian StyleCao, Ming-Ao, Rui-Cheng Liu, Zhi-Yan Xiao, Abeer Hashem, Elsayed Fathi Abd_Allah, Mashail Fahad Alsayed, Wiwiek Harsonowati, and Qiang-Sheng Wu. 2022. "Symbiotic Fungi Alter the Acquisition of Phosphorus in Camellia oleifera through Regulating Root Architecture, Plant Phosphate Transporter Gene Expressions and Soil Phosphatase Activities" Journal of Fungi 8, no. 8: 800. https://doi.org/10.3390/jof8080800