
Colletotrichum Species Associated with Anthracnose Disease of Watermelon (Citrullus lanatus) in China

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Isolates
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Phylogenetic Analyses
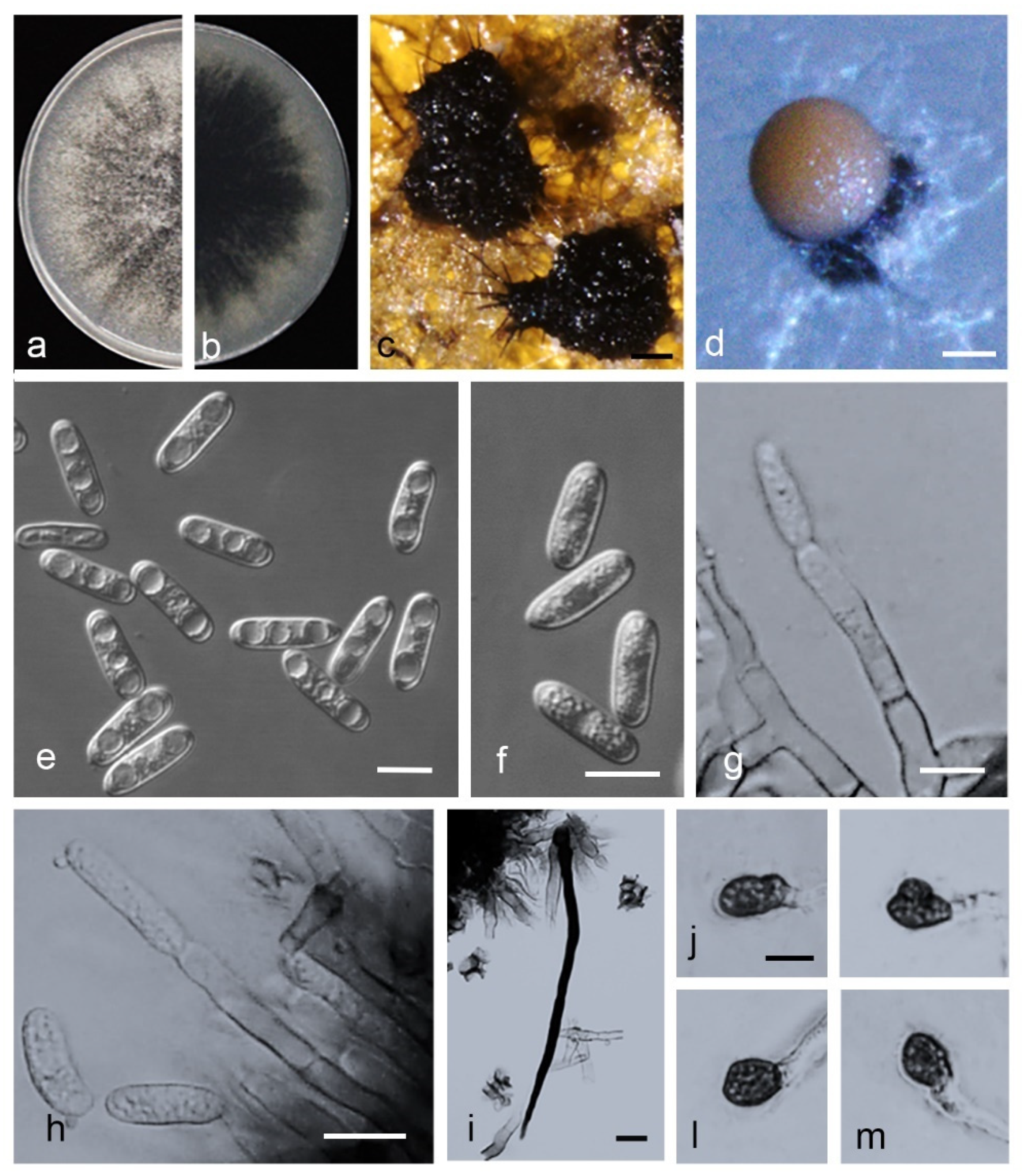
2.4. Morphological Analysis
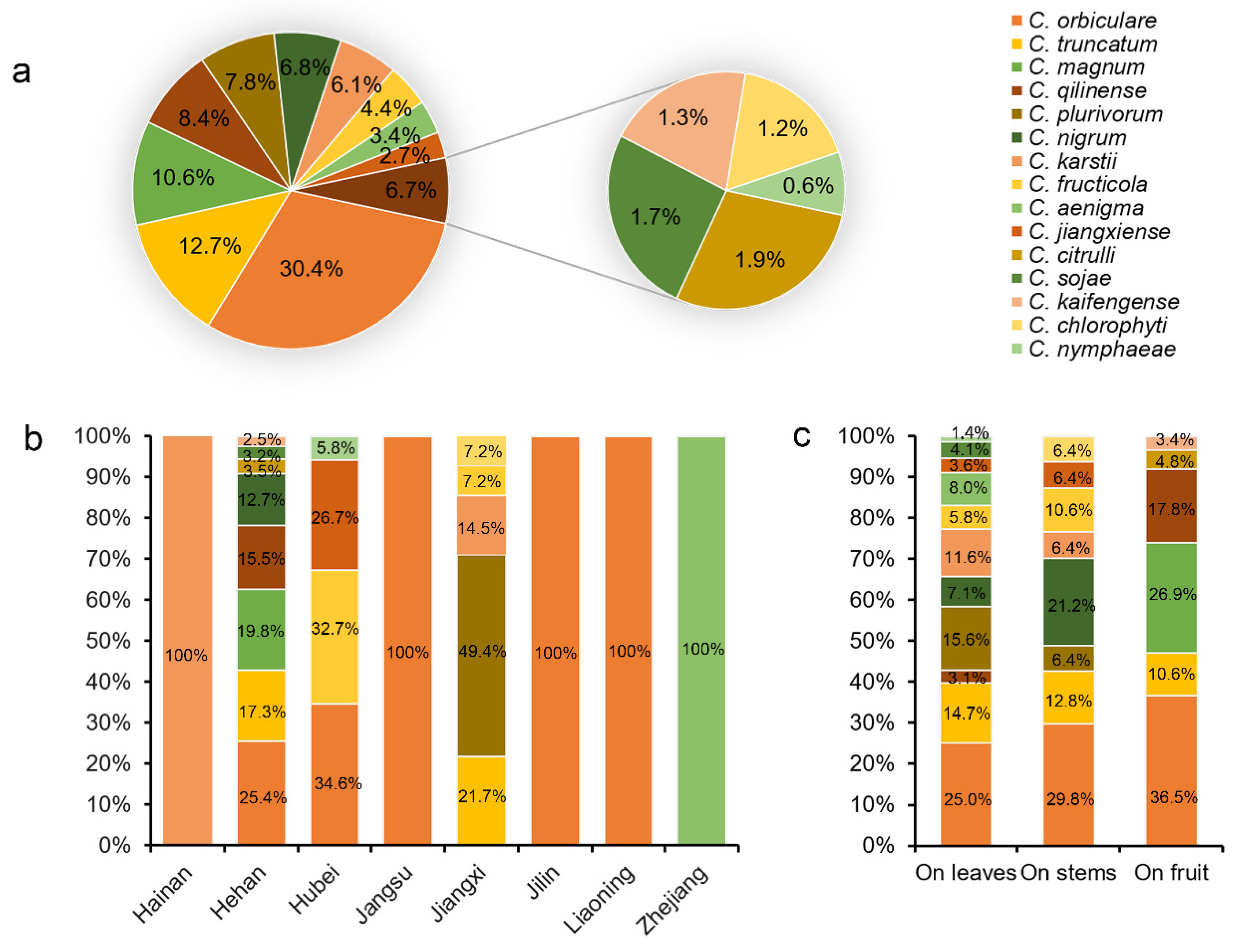
2.5. Prevalence
2.6. Pathogenicity Tests
3. Results
3.1. Collection of Watermelon Anthracnose Samples and Strain Isolation
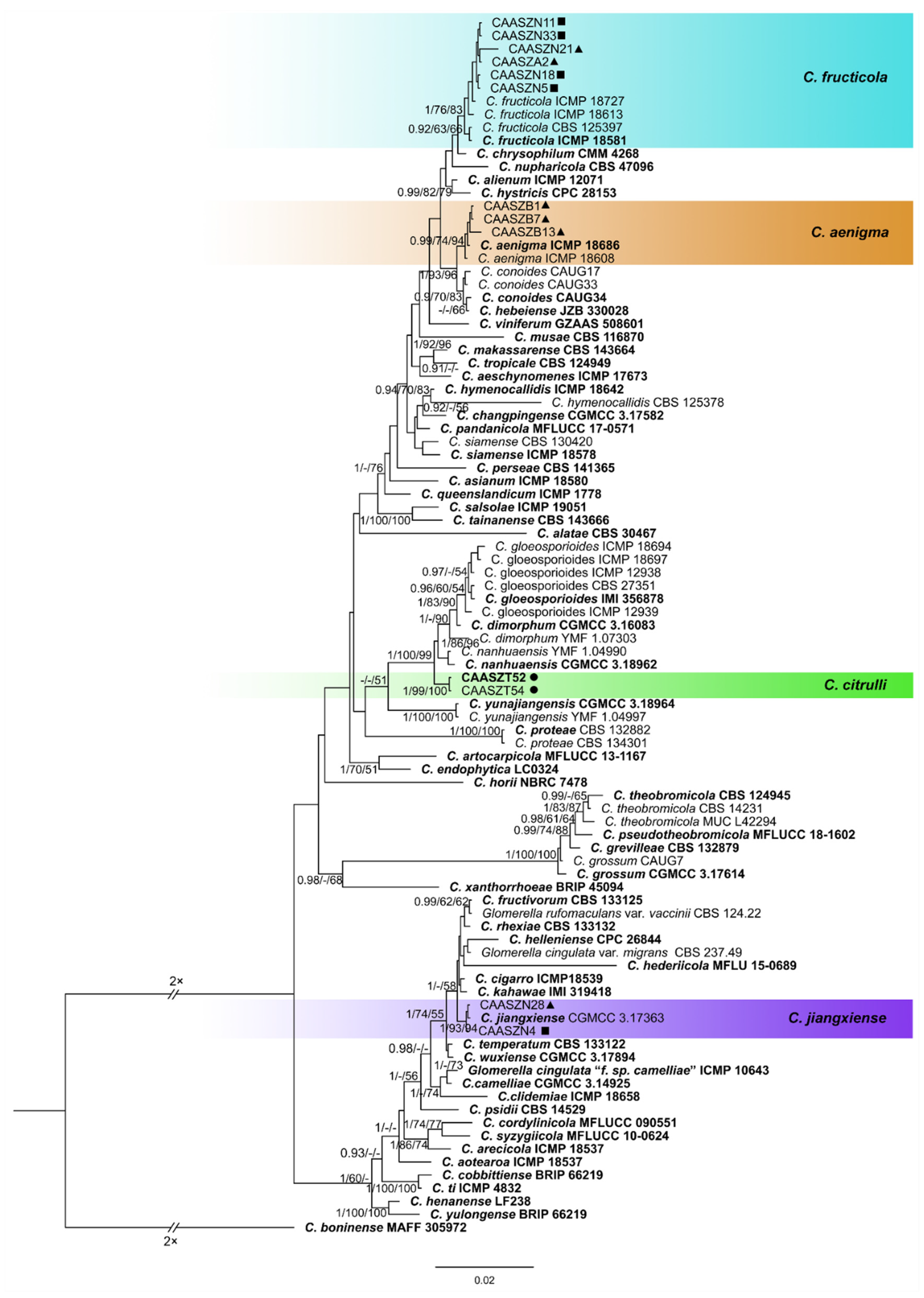
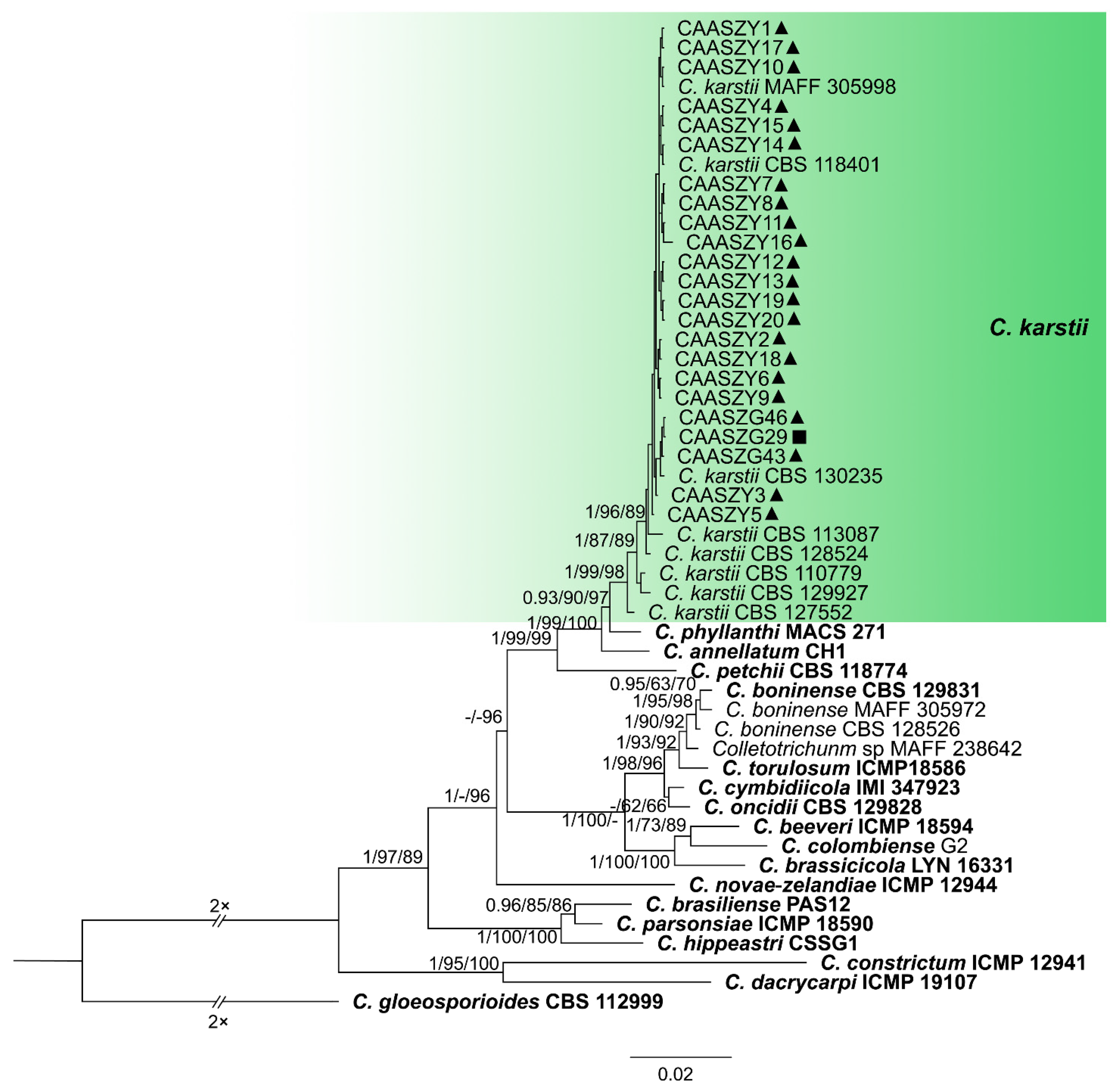
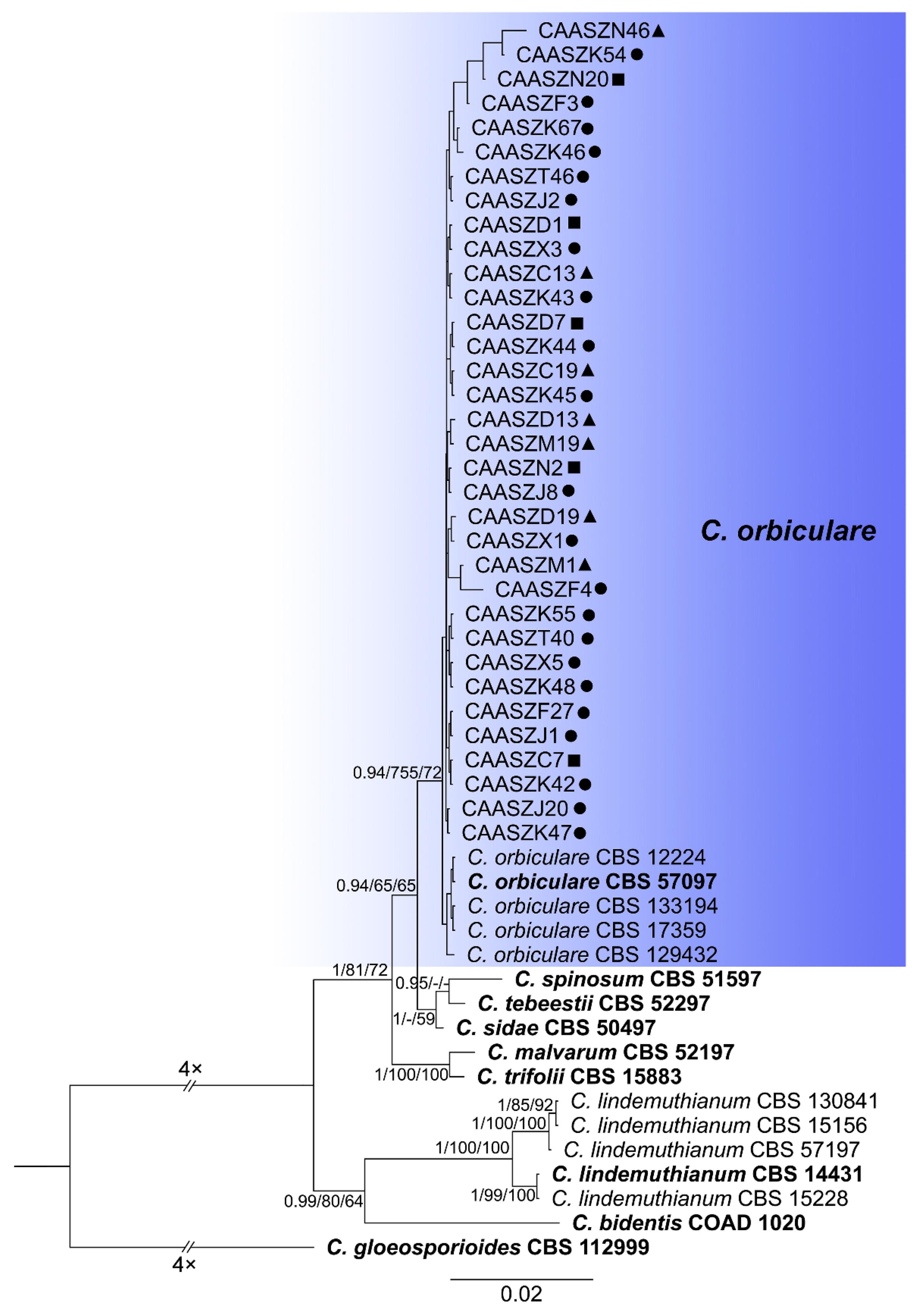
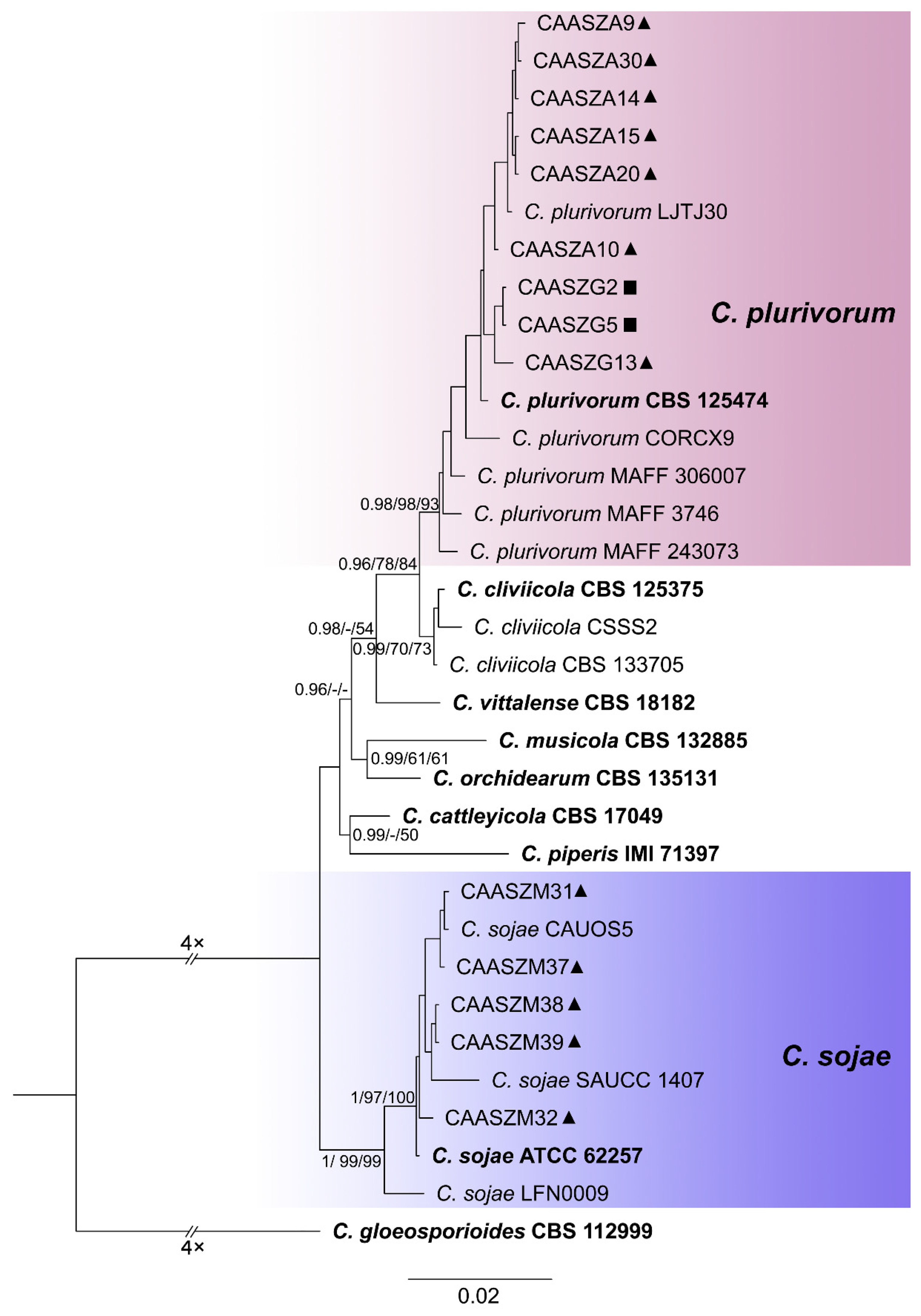
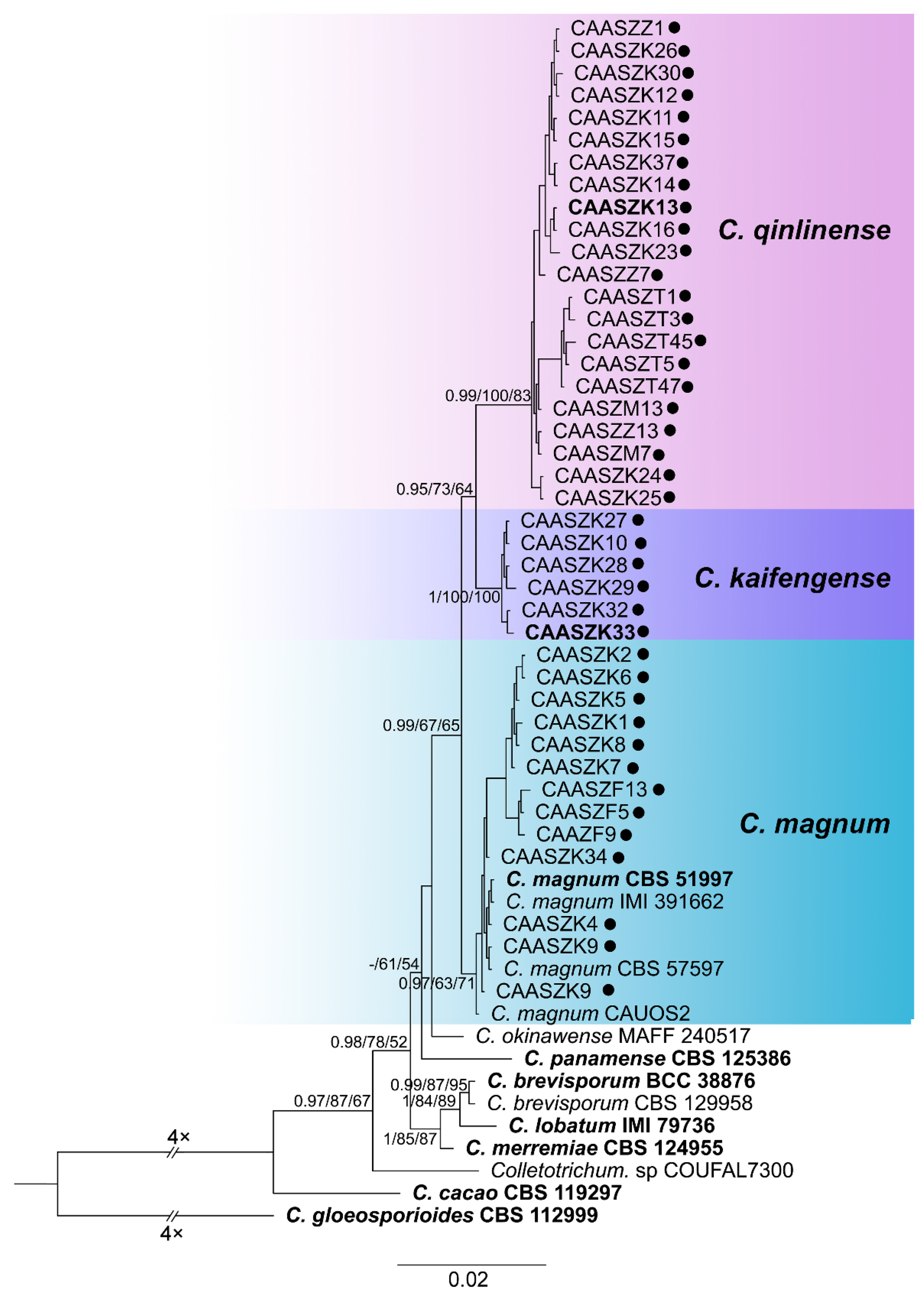
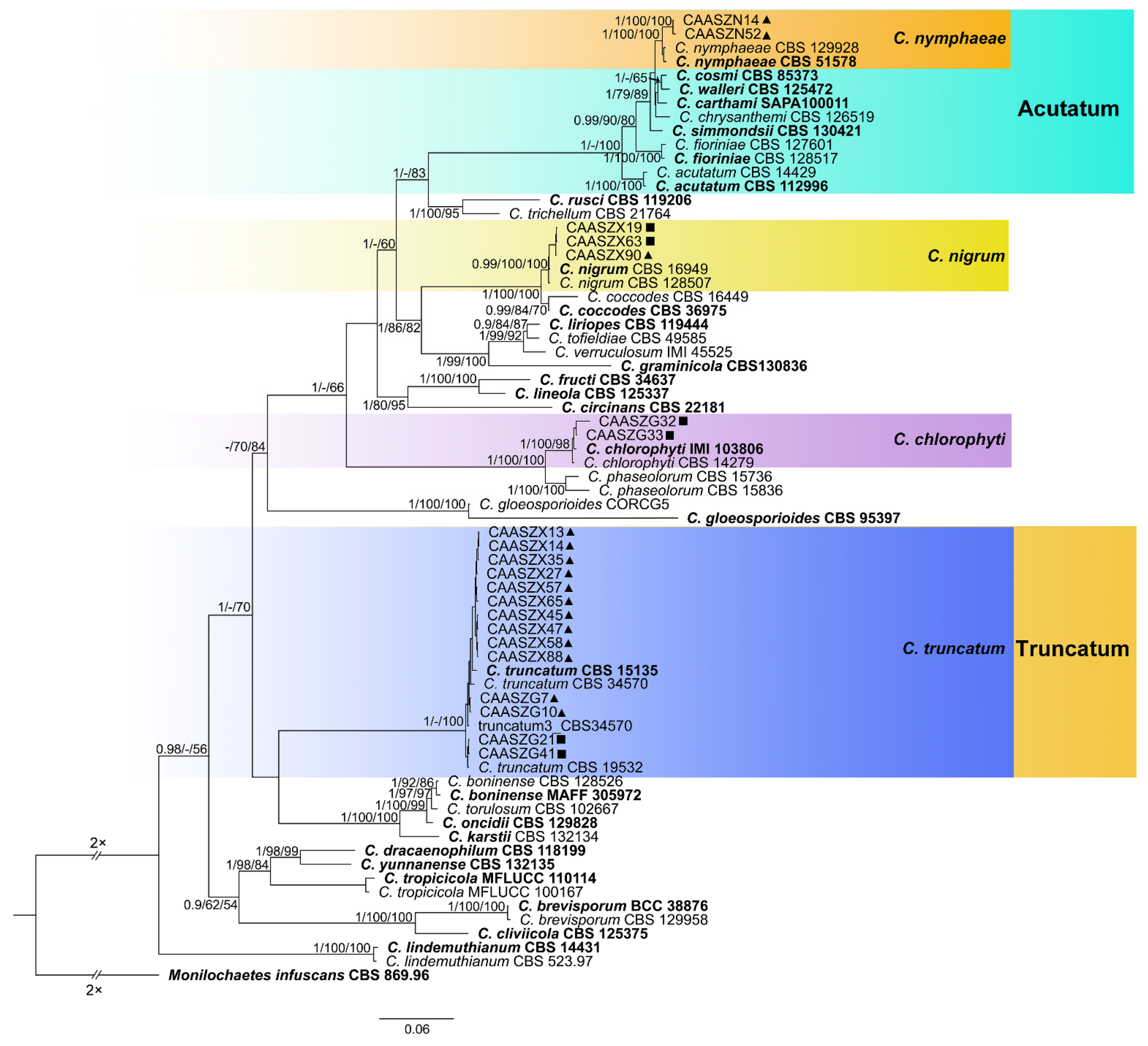
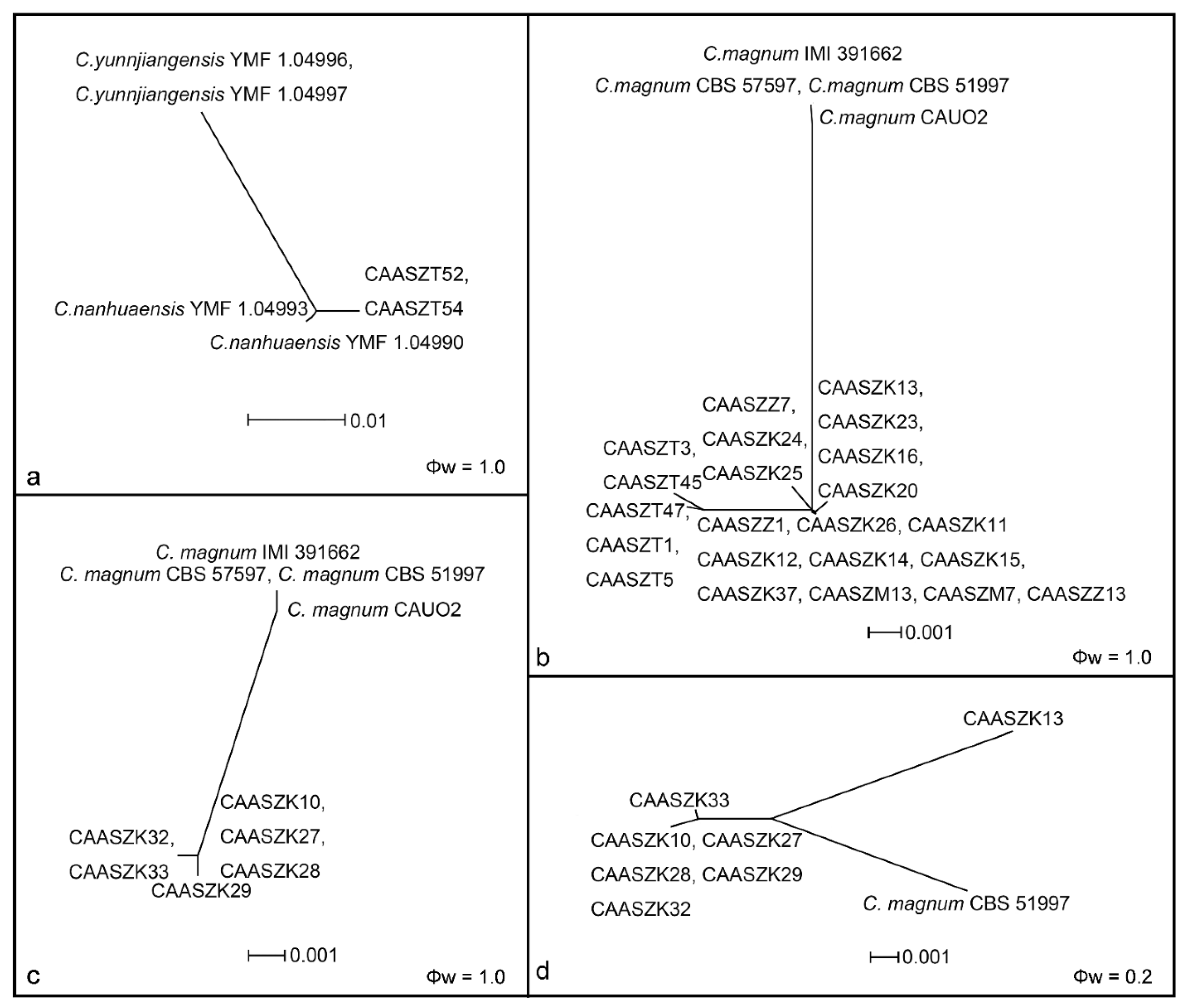
3.2. Multi-Locus Phylogenetic Analyses
3.3. Multi-Locus Phylogenetic Analyses
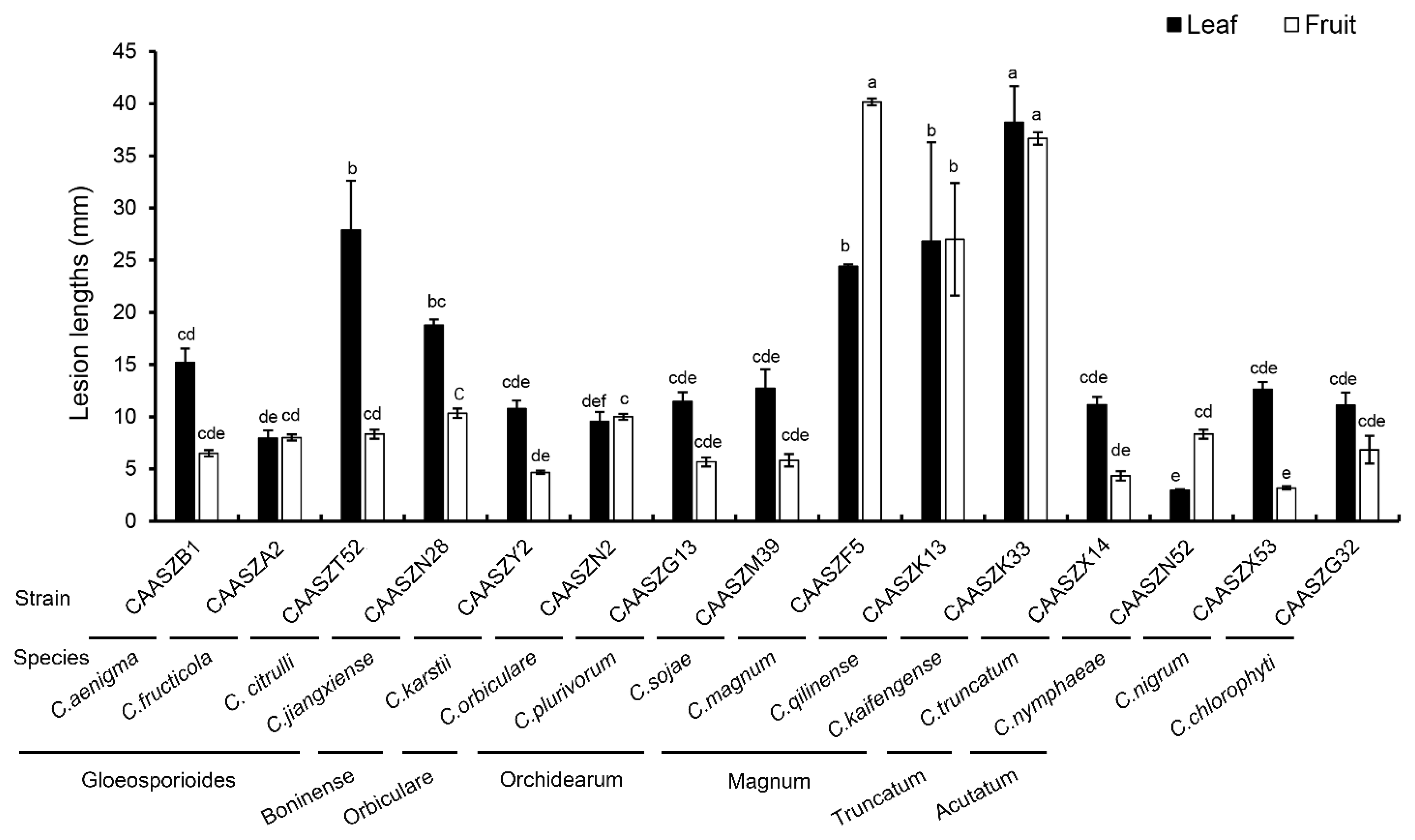
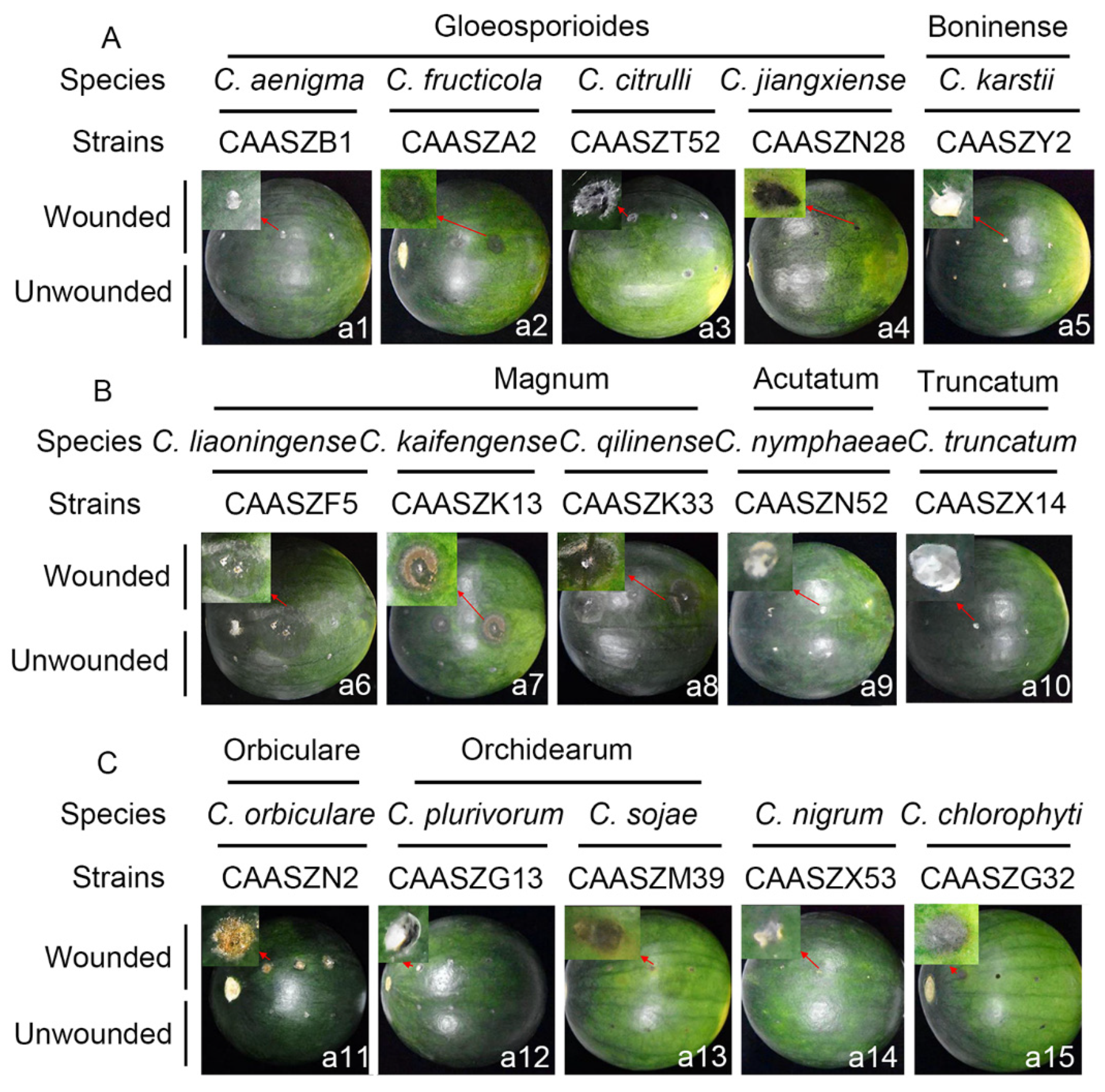
3.4. Pathogenicity Assay
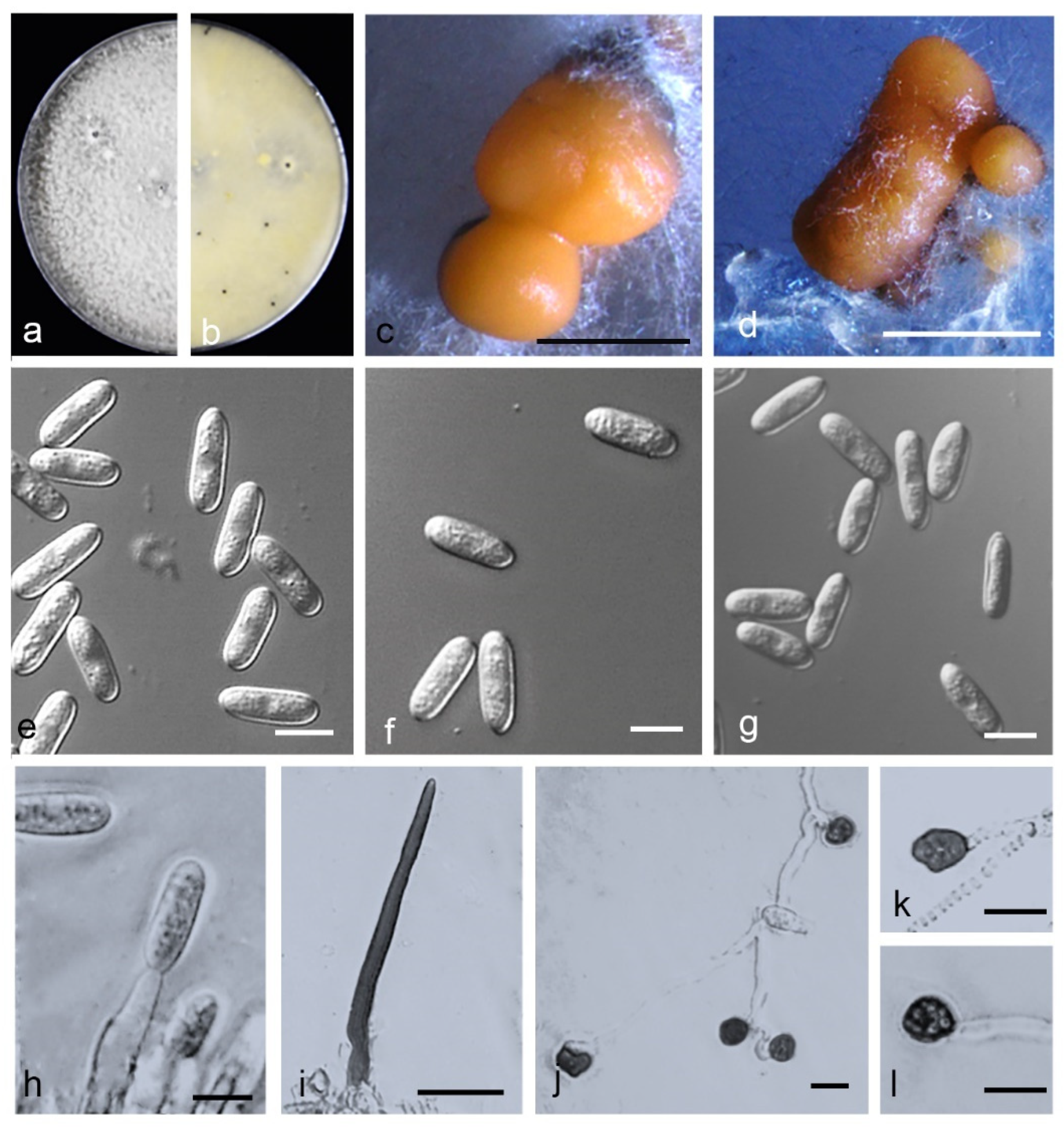
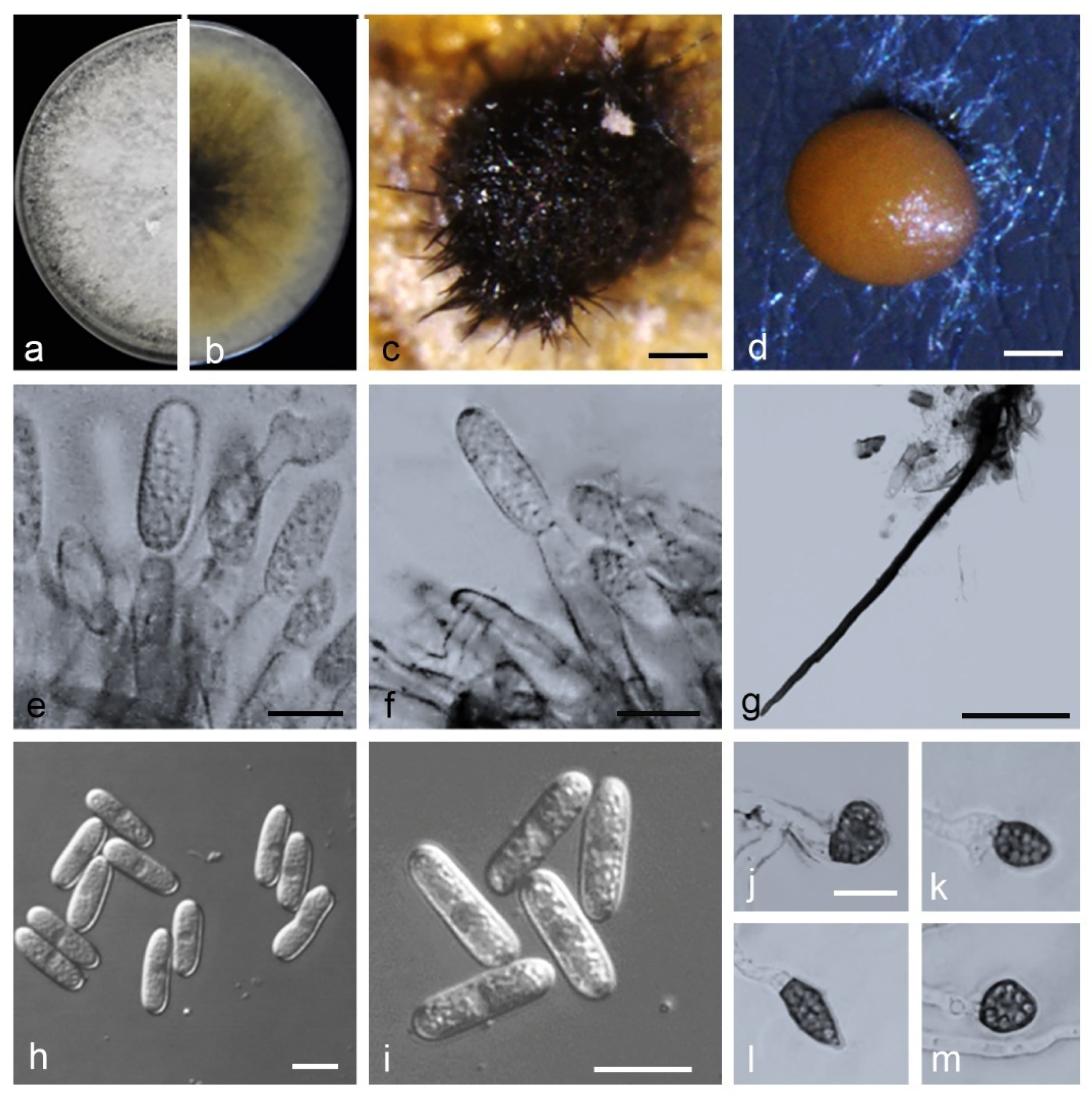
3.5. Taxonomy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum-current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Yu, Z.; Jiang, X.; Fang, L.; Qiao, M. Endophytic Colletotrichum Species from Aquatic Plants in Southwest China. J. Fungi 2022, 8, 87. [Google Scholar] [CrossRef]
- Denoyes-Rothan, B.; Guerin, G.; Delye, C.; Smith, B.; Minz, D.; Maymon, M.; Freeman, S. Genetic diversity and pathogenic variability among isolates of Colletotrichum species from strawberry. Phytopathology 2003, 93, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Prihastuti, H.; McKenzie, E.; Hyde, K.; Cai, L.; Mckenzie, H.; Hyde, E. Characterization of Colletotrichum species associated with coffee berries in northern Thailand. Fungal Divers. 2009, 39, 89–109. [Google Scholar]
- Phoulivong, S.; McKenzie, E.H.C.; Hyde, K.D. Cross infection of Colletotrichum species; a case study with tropical fruits. Curr. Res. Environ. Appl. Mycol. 2012, 2, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Chen, G.Q.; Hou, X.; Fu, Y.S.; Cai, L.; Hyde, K.D.; Li, H.Y. Colletotrichum species associated with cultivated citrus in China. Fungal Divers. 2013, 61, 61–74. [Google Scholar] [CrossRef]
- Lima, N.B.; Batista, M.V.D.A.; De Morais, M.A.; Barbosa, M.A.G.; Michereff, S.J.; Hyde, K.D.; Câmara, M.P.S. Five Colletotrichum species are responsible for mango anthracnose in northeastern Brazil. Fungal Divers. 2013, 61, 75–88. [Google Scholar] [CrossRef]
- Liu, F.; Damm, U.; Cai, L.; Crous, P.W. Species of the Colletotrichum gloeosporioides complex associated with anthracnose diseases of Proteaceae. Fungal Divers. 2013, 61, 89–105. [Google Scholar] [CrossRef]
- Schena, L.; Mosca, S.; Cacciola, S.O.; Faedda, R.; Sanzani, S.M.; Agosteo, G.E.; Sergeeva, V.; di San Lio, G.M. Species of the Colletotrichum gloeosporioides and C. boninense complexes associated with olive anthracnose. Plant Pathol. 2014, 63, 437–446. [Google Scholar] [CrossRef]
- Vieira, W.A.S.; Michereff, S.J.; de Morais, M.A.; Hyde, K.D.; Câmara, M.P.S. Endophytic species of Colletotrichum associated with mango in northeastern Brazil. Fungal Divers. 2014, 67, 181–202. [Google Scholar] [CrossRef]
- Yan, J.-Y.; Jayawardena, M.M.R.S.; Goonasekara, I.D.; Wang, Y.; Zhang, W.; Liu, M.; Huang, J.B.; Wang, Z.Y.; Shang, J.J.; Peng, Y.L.; et al. Diverse species of Colletotrichum associated with grapevine anthracnose in China. Fungal Divers. 2014, 71, 233–246. [Google Scholar] [CrossRef]
- La Hoz, C.J.P.-D.; Calderón, C.; Rincón, A.M.; Cárdenas, M.; Danies, G.; López-Kleine, L.; Restrepo, S.; Jiménez, P. Species from the Colletotrichum acutatum, Colletotrichum boninense and Colletotrichum gloeosporioidesspecies complexes associated with tree tomato and mango crops in Colombia. Plant Pathol. 2016, 65, 227–237. [Google Scholar] [CrossRef]
- Wang, Y.C.; Hao, X.Y.; Wang, L.; Bin, X.; Wang, X.C.; Yang, Y.J. Diverse Colletotrichum species cause anthracnose of tea plants (Camellia sinensis (L.) O. Kuntze) in China. Sci. Rep. 2016, 6, 35287. [Google Scholar] [CrossRef] [Green Version]
- De Silva, D.D.; Ades, P.K.; Crous, P.W.; Taylor, P.W.J. Colletotrichum species associated with chili anthracnose in Australia. Plant Pathol. 2017, 66, 254–267. [Google Scholar] [CrossRef]
- Diao, Y.Z.; Zhang, C.; Liu, F.; Wang, W.Z.; Liu, L.; Cai, L.; Liu, X.L. Colletotrichum species causing anthracnose disease of chili in China. Persoonia 2017, 38, 20–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarnaccia, V.; Groenewald, J.Z.; Polizzi, G.; Crous, P.W. High species diversity in Colletotrichum associated with citrus diseases in Europe. Persoonia 2017, 39, 32–50. [Google Scholar] [CrossRef]
- Fu, M.; Crous, P.W.; Bai, Q.; Zhang, P.F.; Xiang, J.; Guo, Y.S.; Zhao, F.F.; Yang, M.M.; Hong, N.; Xu, W.X.; et al. Colletotrichum species associated with anthracnose of Pyrus spp. in China. Persoonia 2019, 42, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, J.Y.; Xu, X.; Cheng, J.; Zheng, L.; Huang, J.; Li, D.W. Identification and Characterization of Colletotrichum Species Associated with Anthracnose Disease of Camellia oleifera in China. Plant Dis. 2020, 104, 474–482. [Google Scholar] [CrossRef]
- Wang, W.; de Silva, D.D.; Moslemi, A.; Edwards, J.; Ades, P.K.; Crous, P.W.; Taylor, P.W.J. Colletotrichum Species Causing Anthracnose of Citrus in Australia. J. Fungi 2021, 7, 47. [Google Scholar] [CrossRef]
- Tan, Q.; Schnabel, G.; Chaisiri, C.; Yin, L.F.; Yin, W.X.; Luo, C.X. Colletotrichum Species Associated with Peaches in China. J. Fungi 2022, 8, 313. [Google Scholar] [CrossRef] [PubMed]
- Sutton, B.C. The Coelomycetes: Fungi Imperfecti with Pycnidia Acervuli and Stromata; Commonwelth Mycological Institute: Kew, UK, 1980. [Google Scholar]
- Von Arx, J.A. Die Arten der Gattung Colletotrichum Cda. Phytopathol. Z. 1957, 29, 413–468. [Google Scholar]
- Hyde, K.D.; Cai, L.; McKenzie, E.; Yang, Y.L.; Zhang, J.Z.; Prihastuti, H. Colletotrichum: A catalogue of confusion. Fungal Divers. 2009, 39, 1–17. [Google Scholar]
- Hyde, K.D.; Chomnunti, P.; Crous, P.W.; Groenewald, J.Z.; Damm, U.; Ko, T.W.K.; Shivas, R.G.; Summerell, B.A.; Tan, Y.P. A case for re-inventory of Australia’s plant pathogens. Persoonia 2010, 25, 50–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.Z.; Summerell, B.A.; Carnegie, A.J.; Burgess, T.I.; Crous, P.W. Introducing the Consolidated Species Concept to resolve species in the Teratosphaeriaceae. Persoonia 2014, 33, 1–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, C.C.; Tang, J.Y.M.; Lau, S.K.P.; Woo, P.C.Y. Taxonomy and evolution of Aspergillus, Penicillium and Talaromyces in the omics era-Past, present and future. Comput. Struct. Biotechnol. J. 2018, 16, 197–210. [Google Scholar] [CrossRef]
- Cai, L.; Hyde, K.; Taylor, P.; Weir, B.; Waller, J.; Abang, M.; Zhang, J.Z.; Yang, Y.L.; Phoulivong, S.; Liu, Z.Y. A polyphasic approach for studying Colletotrichum. Fungal Divers. 2009, 39, 183–204. [Google Scholar]
- Yang, Y.L.; Liu, Z.Y.; Cai, L.; Hyde, K.; Yu, Z.N.; McKenzie, E. Colletotrichum anthracnose of Amaryllidaceae. Fungal Divers. 2009, 39, 123–146. [Google Scholar]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Cai, L.; Crous, P.W.; Damm, U. The Colletotrichum gigasporum species complex. Persoonia 2014, 33, 83–97. [Google Scholar] [CrossRef] [Green Version]
- Damm, U.; Sato, T.; Alizadeh, A.; Groenewald, J.Z.; Crous, P.W. The Colletotrichum dracaenophilum, C. magnum and C. orchidearum species complexes. Stud. Mycol. 2019, 92, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Jiang, X.; Zheng, H.; Zhang, H.; Qiao, M. Fourteen New Species of Foliar Colletotrichum Associated with the Invasive Plant Ageratina adenophora and Surrounding Crops. J. Fungi 2022, 8, 185. [Google Scholar] [CrossRef] [PubMed]
- Damm, U.; Woudenberg, J.H.C.; Cannon, P.; Crous, P.W. Colletotrichum species with curved conidia from herbaceous hosts. Fungal Divers. 2009, 39, 45. [Google Scholar]
- Shivas, R.; Tan, Y.P. A taxonomic re-assessment of Colletotrichum acutatum, introducing C. fioriniae comb. et stat. nov. and C. simmondsii sp. nov. Fungal Divers. 2009, 39, 111–122. [Google Scholar]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.; Crous, P.W. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.; Johnston, P.R.; Weir, B.S.; Tan, Y.P.; Shivas, R.G.; Crous, P.W. The Colletotrichum boninense species complex. Stud. Mycol. 2012, 73, 1–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damm, U.; Cannon, P.F.; Liu, F.; Barreto, R.W.; Guatimosim, E.; Crous, P.W. The Colletotrichum orbiculare species complex: Important pathogens of field crops and weeds. Fungal Divers. 2013, 61, 29–59. [Google Scholar] [CrossRef]
- Damm, U.; O’Connell, R.J.; Groenewald, J.Z.; Crous, P.W. The Colletotrichum destructivum species complex-hemibiotrophic pathogens of forage and field crops. Stud. Mycol. 2014, 79, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Marin-Felix, Y.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Marincowitz, S.; Barnes, I.; Bensch, K.; Braun, U.; Camporesi, E.; Damm, U.; et al. Genera of phytopathogenic fungi: GOPHY 1. Stud. Mycol. 2017, 86, 99–216. [Google Scholar] [CrossRef] [Green Version]
- Jayawardena, R.S.; Hyde, K.D.; Chen, Y.J.; Papp, V.; Palla, B.; Papp, D.; Bhunjun, C.S.; Hurdeal, V.G.; Senwanna, C.; Manawasinghe, I.S.; et al. One stop shop IV: Taxonomic update with molecular phylogeny for important phytopathogenic genera: 76–100 (2020). Fungal Divers. 2020, 103, 87–218. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Phukhamsakda, C.; Jayawardena, R.S.; Jeewon, R.; Promputtha, I.; Hyde, K.D. Investigating species boundaries in Colletotrichum. Fungal Divers. 2021, 107, 107–127. [Google Scholar] [CrossRef]
- Liu, F.; Ma, Z.Y.; Hou, L.W.; Diao, Y.Z.; Wu, W.P.; Damm, U.; Song, S.; Cai, L. Updating species diversity of Colletotrichum, with a phylogenomic overview. Stud. Mycol. 2022, 101, 1–56. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, J.; Sun, H.; Salse, J.; Lucas, W.J.; Zhang, H.; Zheng, Y.; Mao, L.; Ren, Y.; Wang, Z.; et al. The draft genome of watermelon (Citrullus lanatus) and resequencing of 20 diverse accessions. Nat. Genet. 2013, 45, 51–58. [Google Scholar] [CrossRef] [Green Version]
- FAO. Food and Agricultural Organization of the United Nations (FAOSTAT) Website. 2020. Available online: https://www.fao.org/faostat/zh/#data/QCL (accessed on 7 January 2022).
- Prusky, D. Pathogen quiescence in postharvest diseases. Annul. Rev. Phytopathol. 1996, 34, 413–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farr, D.F.; Aime, M.C.; Rossman, A.Y.; Palm, M.E. Species of Colletotrichum on agavaceae. Mycol. Res. 2006, 110, 1395–1408. [Google Scholar] [CrossRef] [PubMed]
- Du, M.Z.; Schardl, C.L.; Nuckles, E.M.; Vaillancourt, L.J. Using mating-type gene sequences for improved phylogenetic resolution of Colletotrichum species complexes. Mycologia 2005, 97, 641–658. [Google Scholar] [CrossRef]
- Zivkovic, S.T.; Stosic, S.S.; Stevanovic, M.L.; Gasic, K.M.; Aleksic, G.A.; Vucurovic, I.B.; Ristic, D.T. Colletotrichum orbiculare on watermelon: Identification and in vitro inhibition by antagonistic fungi. Zb. Matice Srp. Za Prir. Nauk. 2017, 331–343. [Google Scholar] [CrossRef]
- UASD (U.S. Department of Agriculture). U.S. National Fungus Collections Statistics. 2022. Available online: https://www.fao.org/faostat/zh/#data/QCL (accessed on 20 April 2022).
- Goh, K.S.; Balasubramaniam, J.; Sani, S.F.; Alam, M.W.; Ismail, N.A.; Gleason, M.L.; Rosli, H. First Report of Colletotrichum scovillei Causing Anthracnose on Watermelon (Citrullus lanatus) in Malaysia. Plant Dis. 2022; Advance online publication. [Google Scholar] [CrossRef]
- Da Silva, V.N.; Guzzo, S.D.; Lucon, C.M.M.; Harakava, R. Promoção de crescimento e indução de resistencia a antracnose por Trichoderma spp. em pepineiro. Pesqui. Agropecu. Bras. 2011, 46, 1609–1618. [Google Scholar] [CrossRef] [Green Version]
- Fang, Z. Plant Disease Research Methods, 3rd ed.; China Agriculture Press: Beijing, China, 1998; pp. 122–145. [Google Scholar]
- Chomnunti, P.; Hongsanan, S.; Aguirre-Hudson, B.; Tian, Q.; Peršoh, D.; Dhami, M.K.; Alias, A.S.; Xu, J.; Liu, X.; Stadler, M.; et al. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.; Gelfand, D.; Sninsky, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols; Academic Press: San Diego, CA, USA, 1990; Volume 31, pp. 315–322. [Google Scholar]
- Guerber, J.C.; Liu, B.; Correll, J.C.; Johnston, P.R. Characterization of Diversity in Colletotrichum acutatum sensu lato by Sequence Analysis of Two Gene Introns, mtDNA and Intron RFLPs, and Mating Compatibility. Mycologia 2003, 95, 872–895. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z.; Risede, J.-M.; Simoneau, P.; Hywel-Jones, N. Calonectria species and their Cylindrocladium anamorphs: Species with sphaeropedunculate vesicles. Stud. Mycol. 2004, 50, 415–430. [Google Scholar]
- O’Donnell, K.; Cigelnik, E. Two Divergent Intragenomic rDNA ITS2 Types within a Monophyletic Lineage of the Fungus Fusarium Are Nonorthologous. Mol. Phyl. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Glass, L.N.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from Filamentous Ascomycetes. Am. Soc. Microbiol. 1995, 61, 1320–1330. [Google Scholar] [CrossRef] [Green Version]
- Woudenberg, J.H.; Aveskamp, M.M.; de Gruyter, J.; Spiers, A.G.; Crous, P.W. Multiple Didymella teleomorphs are linked to the Phoma clematidina morphotype. Persoonia 2009, 22, 56–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J. MrModeltest V2. Program Distributed by the Author. Bioinformatics 2004, 24, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swofford, L.D.; Bell, D.C. PAUP 4.0 b10. In Phylogenetic Analysis Using Parsimony (* and Other Methods); Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree v. 1.4.4. Institute of Evolutionary Biology, University of Edinburgh. 2018. Available online: https://tree.bio.ed.ac.uk/software/figtree/ (accessed on 1 March 2022).
- Huson, D.H. SplitsTree: Analyzing and visualizing evolutionary data. Bioinformatics 1998, 14, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Kloepper, T.H. Computing recombination networks from binary sequences. Bioinformatics 2005, 21 (Suppl. S2), ii159–ii165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Nirenberg, H. Untersuchungen über die Morphologische und Biologische Differenzierung in der Fusarium-Sektion Liseola. Mitteilungen aus der Biologischen Bundesanstalt für Landund Forstwirtschaft (Berlin-Dahlem). 1976. No. 169. Available online: http://pascal-francis.inist.fr/vibad/index.php?action=getRecordDetail&idt=PASCAL7710018408 (accessed on 1 March 2022).
- Lin, Q.; Kanchana-udomkarn, C.; Jaunet, T.; Mongkolporn, O. Genetic analysis of resistance to pepper anthracnose caused by Colletotrichum capsici. Thail. J. Agric. Sci. 2004, 35, 259–264. [Google Scholar]
- Than, P.P.; Jeewon, R.; Hyde, K.D.; Pongsupasamit, S.; Mongkolporn, O.; Taylor, P.W.J. Characterization and pathogenicity of Colletotrichum species associated with anthracnose on chilli (Capsicum spp.) in Thailand. Plant Pathol. 2008, 57, 562–572. [Google Scholar] [CrossRef]
- Crouch, J.A.; Clarke, B.; Hillman, B. Unraveling Evolutionary Relationships Among the Divergent Lineages of Colletotrichum Causing Anthracnose Disease in Turfgrass and Corn. Phytopathology 2006, 96, 46–60. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Weir, B.S.; Damm, U.; Crous, P.W.; Wang, Y.; Liu, B.; Wang, M.; Zhang, M.; Cai, L. Unravelling Colletotrichum species associated with Camellia: Employing ApMat and GS loci to resolve species in the C. gloeosporioides complex. Persoonia 2015, 35, 63–86. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Chen, X.; Ren, G.; Wang, J.; Liu, L.; Qian, W.; Wang, J. First report of Colletotrichum chlorophyti causing peanut anthracnose in China. Plant Dis. 2020, 105, 226. [Google Scholar] [CrossRef]
- Tashiro, N.; Manabe, K.; Ide, Y. Emergence and frequency of highly benzimidazole-resistant Colletotrichum gloeosporioides, pathogen of Japanese pear anthracnose, after discontinued use of benzimidazole. J. Gen. Plant Pathol. 2012, 78, 221–226. [Google Scholar] [CrossRef]
- Peres, N.A.; Timmer, L.W.; Adaskaveg, J.E.; Correll, J.C. Lifestyles of Colletotrichum acutatum. Plant Dis. 2005, 89, 784–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkan, N.; Friedlander, G.; Ment, D.; Prusky, D.; Fluhr, R. Simultaneous transcriptome analysis of Colletotrichum gloeosporioides and tomato fruit pathosystem reveals novel fungal pathogenicity and fruit defense strategies. New Phytol. 2015, 205, 801–815. [Google Scholar] [CrossRef] [PubMed]
- De Silva, D.D.; Crous, P.W.; Ades, P.K.; Hyde, K.D.; Taylor, P.W.J. Life styles of Colletotrichum species and implications for plant biosecurity. Fungal Biol. Rev. 2017, 31, 155–168. [Google Scholar] [CrossRef]
- Jiang, J.; Zhai, H.; Li, H.; Wang, Z.; Chen, Y.; Hong, N.; Wang, G.; Chofong, G.N.; Xu, W. Identification and characterization of Colletotrichum fructicola causing black spots on young fruits related to bitter rot of pear (Pyrus bretschneideri Rehd.) in China. Crop Prot. 2014, 58, 41–48. [Google Scholar] [CrossRef]
- De Silva, D.D.; Groenewald, J.Z.; Crous, P.W.; Ades, P.K.; Nasruddin, A.; Mongkolporn, O.; Taylor, P.W.J. Identification, prevalence and pathogenicity of Colletotrichum species causing anthracnose of Capsicum annuum in Asia. IMA Fungus 2019, 10, 8. [Google Scholar] [CrossRef]
- Shivas, R.G.; Tan, Y.P.; Edwards, J.; Dinh, Q.; Maxwell, A.; Andjic, V.; Liberato, J.R.; Anderson, C.; Beasley, D.R.; Bransgrove, K.; et al. Colletotrichum species in Australia. Australas. Plant Pathol. 2016, 45, 447–464. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Host Tissue | Year | Latitude and Longitude | Number of Isolates |
|---|---|---|---|---|
| Jiyang, Hainan | leaf and stem | 2018 | 109.58° E, 18.28° N | 20 |
| Tongxu, Henan | fruit | 2019 | 114.47° E, 34.48° N | 8 |
| Weishi, Henan | fruit | 2019 | 114.29° E, 34.35° N | 16 |
| Xiangfu, Henan | fruit | 2019 | 114.35° E, 34.77° N | 6 |
| Jinming, Henan | fruit | 2019 | 114.31° E, 34.87° N | 19 |
| fruit | 2020 | 26 | ||
| Zhongmou, Henan | leaf and stem | 2019 | 113.98° E, 34.71° N | 25 |
| leaf and stem | 2020 | 9 | ||
| Xinzheng, Henan | fruit | 2019 | 113.74° E, 34.39° N | 8 |
| Fengcheng, Liaoning | leaf and stem | 2019 | 124.06° E, 40.45° N | 24 |
| Ningbo, Zhejiang | leaf and stem | 2019 | 121.54° E, 29.87° N | 18 |
| Taikang, Hennan | fruit | 2019 | 114.62° E, 34.11° N | 44 |
| Ganzhou, Jingxi | leaf and stem | 2020 | 115.78° E, 25.60° N | 48 |
| Xinxiang, Henan | leaf, stem and fruit | 2020 | 113.85° E, 35.30° N | 91 |
| Enshi, Hubei | leaf and stem | 2020 | 109.47° E, 30.29° N | 52 |
| Fugou, Henan | fruit | 2020 | 114.39° E, 34.05° N | 31 |
| Yancheng, Jiangsu | fruit | 2020 | 120.15° E, 33.34° N | 22 |
| Gaoan, Jiangxi | leaf and stem | 2020 | 115.37° E, 28.41° N | 35 |
| Tongyu, Jilin | leaf | 2020 | 123.08° E, 44.80° N | 24 |
| Total | 526 |
| Gene | ITS | gadph | chs-1 | his3 | act | tub2 | gs |
|---|---|---|---|---|---|---|---|
| Gloeosporioides clade | GTR + I | HKY + I | K80 + G | GTR + G | GTR + I | SYM + G | – |
| Boninense clade | SYM + I + G | HKY + G | K80 + G | GTR + I+G | GTR + G | K80 + I | – |
| Orbiculare clade | GTR | SYM | SYM + I | GTR + G | HKY + G | HKY | GTR + G |
| Orchidearum clade | GTR + I | HKY | GTR + G | HKY + G | HKY | HKY + I | – |
| Magnum clade | GTR + G | HKY + G | GTR + I | HKY + I | HKY + G | GTR + G | – |
| Acutatum clade and other taxa | GTR + I + G | HKY + I + G | GTR + I+G | – | HKY + I + G | HKY + I + G | – |
| Species | Strain | Infection Incidence % | |
|---|---|---|---|
| Leaf Bioassay | Fruit Bioassay | ||
| C.aenigma | CAASZB1 | 44.4 | 0.0 |
| C.fructicola | CAASZA2 | 0.0 | 0.0 |
| C. citrullus | CAASZT52 | 66.7 | 66.7 |
| C.jiangxiense | CAASZN28 | 44.4 | 33.3 |
| C.karstii | CAASZY2 | 44.4 | 100.0 |
| C.orbiculare | CAASZN2 | 44.4 | 100.0 |
| C.plurivorum | CAASZG13 | 55.6 | 66.7 |
| C.sojae | CAASZM39 | 33.3 | 100.0 |
| C.magnum | CAASZF5 | 66.7 | 100.0 |
| C.qilinense | CAASZK13 | 77.8 | 66.7 |
| C.kaifengense | CAASZK33 | 66.7 | 66.7 |
| C.nymphaeae | CAASZN52 | 0.0 | 66.7 |
| C.nigrum | CAASZX53 | 44.4 | 0.0 |
| C.chlorophyti | CAASZG32 | 0.0 | 33.3 |
| C.truncatum | CAASZX14 | 33.3 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Z.; Luo, C.-X.; Wu, H.-J.; Peng, B.; Kang, B.-S.; Liu, L.-M.; Zhang, M.; Gu, Q.-S. Colletotrichum Species Associated with Anthracnose Disease of Watermelon (Citrullus lanatus) in China. J. Fungi 2022, 8, 790. https://doi.org/10.3390/jof8080790
Guo Z, Luo C-X, Wu H-J, Peng B, Kang B-S, Liu L-M, Zhang M, Gu Q-S. Colletotrichum Species Associated with Anthracnose Disease of Watermelon (Citrullus lanatus) in China. Journal of Fungi. 2022; 8(8):790. https://doi.org/10.3390/jof8080790
Chicago/Turabian StyleGuo, Zhen, Chao-Xi Luo, Hui-Jie Wu, Bin Peng, Bao-Shan Kang, Li-Ming Liu, Meng Zhang, and Qin-Sheng Gu. 2022. "Colletotrichum Species Associated with Anthracnose Disease of Watermelon (Citrullus lanatus) in China" Journal of Fungi 8, no. 8: 790. https://doi.org/10.3390/jof8080790