
Immunomodulatory Molecular Mechanisms of Luffa cylindrica for Downy Mildews Resistance Induced by Growth-Promoting Endophytic Fungi

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Endophytic Isolation, Characterization, and Growth-Promotion Plant Bioassays
- (1)
- Control;
- (2)
- ThM9;
- (3)
- TvA1;
- (4)
- TvA1 + ThM9.
2.2. Pathogen Isolation, Characterization, and Plant Pathogenicity Bioassay
2.3. In Vitro Screening of Fungal Endophytes for Anti-Fungal Enzymes and Metabolites
2.4. In Planta Screening for Endophytic Fungal Antagonism against DM
- (1)
- Control = nutrient solution + Czapek media;
- (2)
- Endophytic fungus 1 = ThM9;
- (3)
- Endophytic fungus 2 = TvA1;
- (4)
- Endophytic fungus 1 + 2 = TvA1+ThM9;
- (5)
- Control = nutrient solution + P. cubensis;
- (6)
- Endophytic fungus 1 + pathogen = ThM9 + P. cubensis;
- (7)
- Endophytic fungus 2 + pathogen = TvA1 + P. cubensis;
- (8)
- Endophytic fungus 1 + 2 + pathogen = TvA1 + ThM9 + P. cubensis.
- Grade 0: leaves (disease spots absent);
- Grade 1: Disease spots (<5%/total leaf area);
- Grade 2: Disease spots (6–10%/total leaf area);
- Grade 3: Disease spots (11–25%/total leaf area);
- Grade 4: Disease spots (26–50%/total leaf area);
- Grade 5: Disease spots (50–60%/total leaf area);
- Grade 6: Disease spots (60–70%/total leaf area);
- Grade 7: Disease spots (>70%/total leaf area).
2.5. Characterization of Plants for Growth Promotion and Pathogen Resistance
2.6. Molecular Analysis for the Response of Disease-Resistance Genes
2.7. Statistical Analysis
3. Results
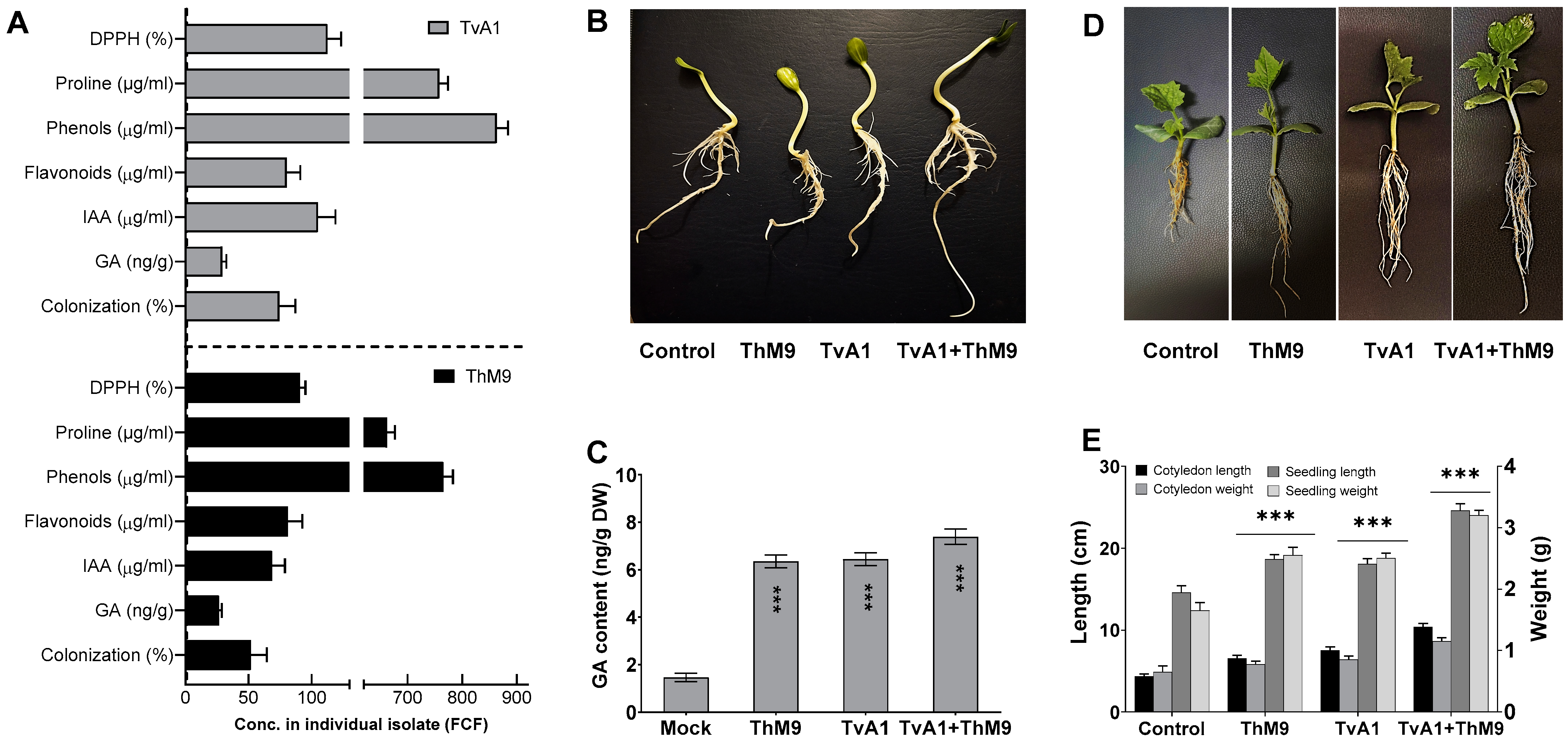
3.1. Isolation and Screening of Endophytic Fungal Isolates Based on Growth-Promoting Traits
3.2. Molecular Identification of Fungal Endophytes (M9 and A1) and Fungal Pathogen (DM1)
3.3. Compatibility Test for ThM9 and TvA1
3.4. Effect of ThM9 and TvA1 Endophytes on Growth Traits of Luffa under Laboratory Conditions
3.5. Effect of ThM9 and TvA1 Endophytes on Seed Germination and Seedling Vigor of Luffa
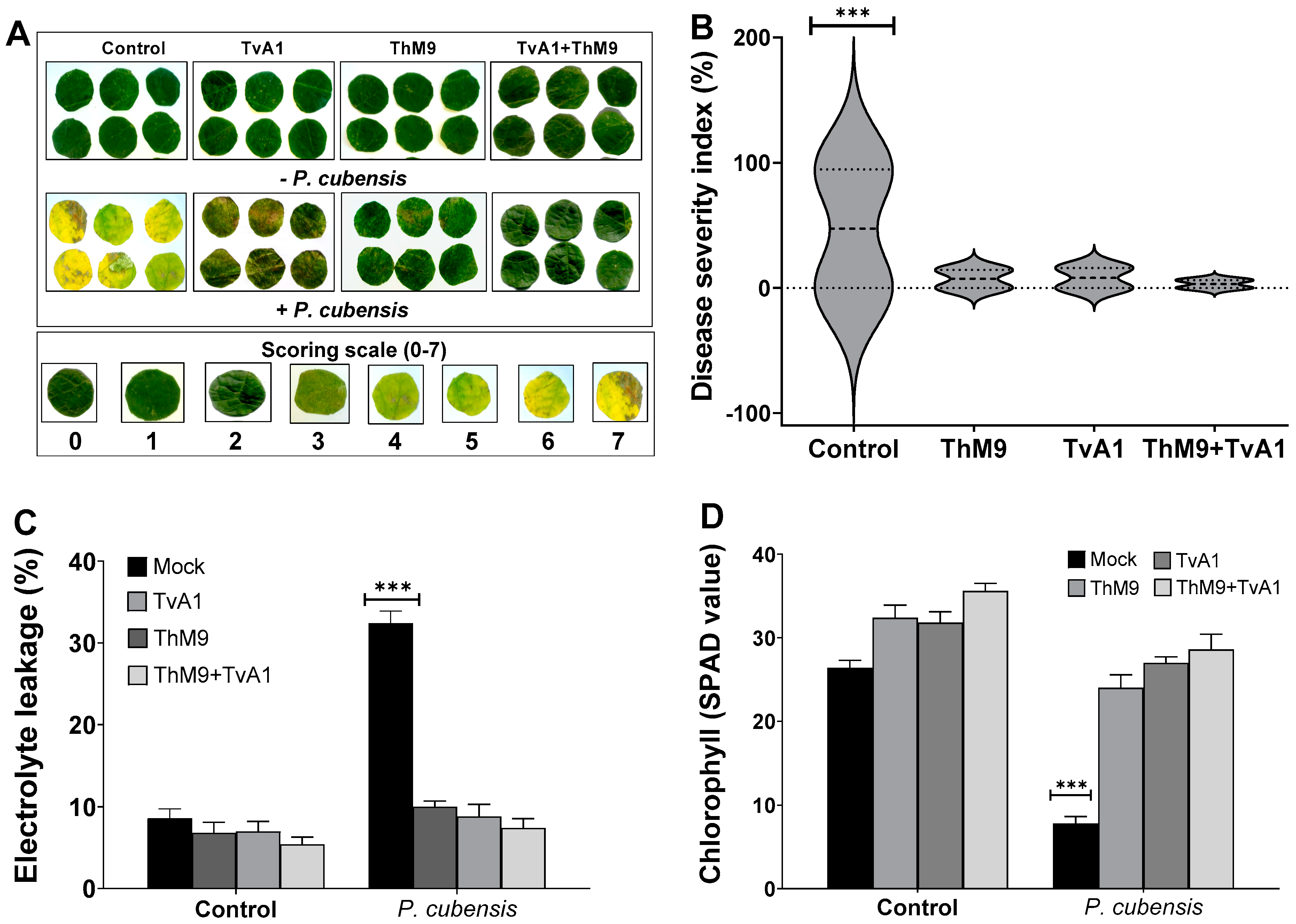
3.6. Screening of ThM9 and TvA1 Endophytes for Antagonistic Response against Luffa DM
3.7. The Predominance of Plant-Disease-Resistance Traits of Endophytes
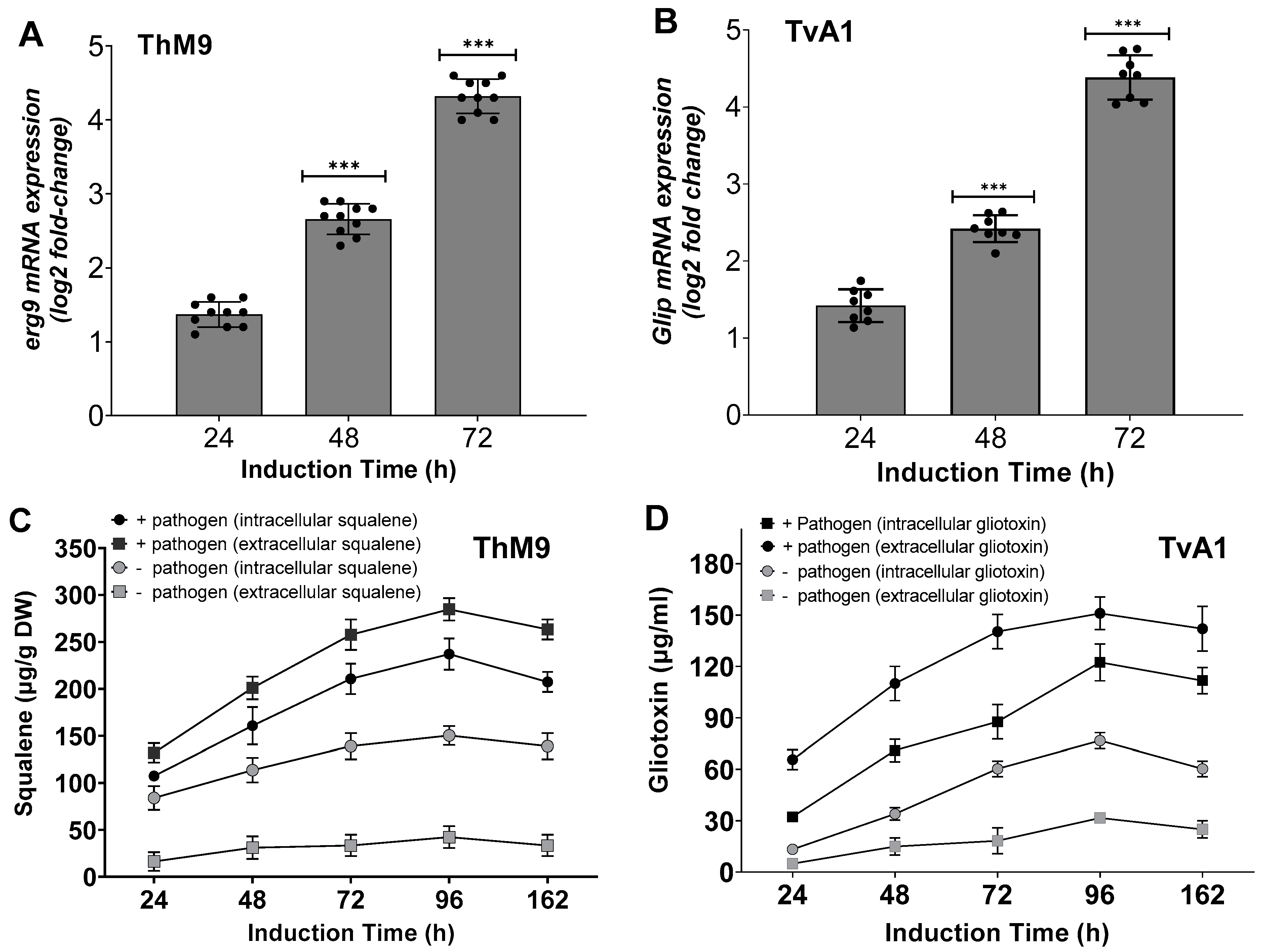
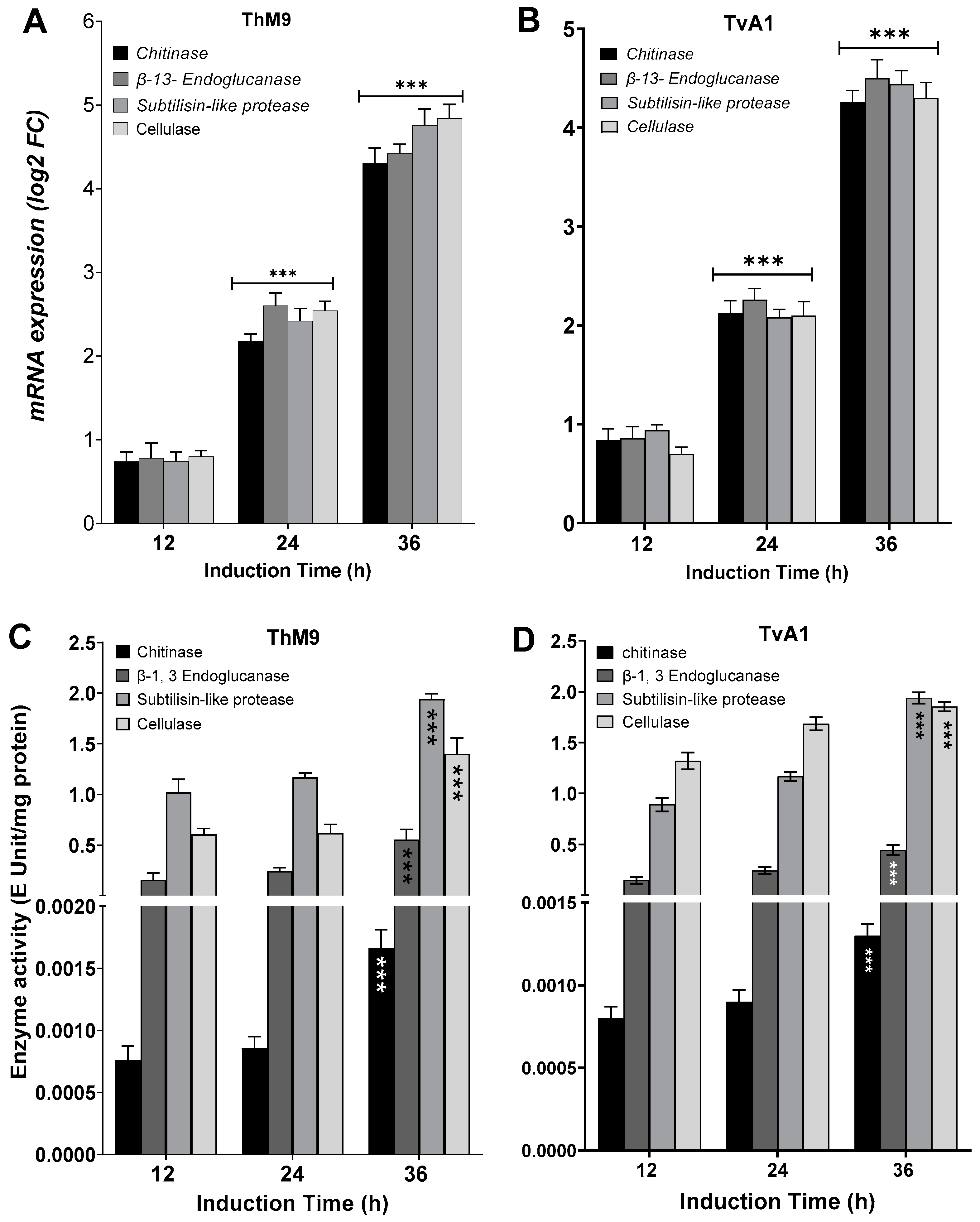
3.8. Induction of Defense-Related Hydrolytic Enzymes of ThM9 and TvA1
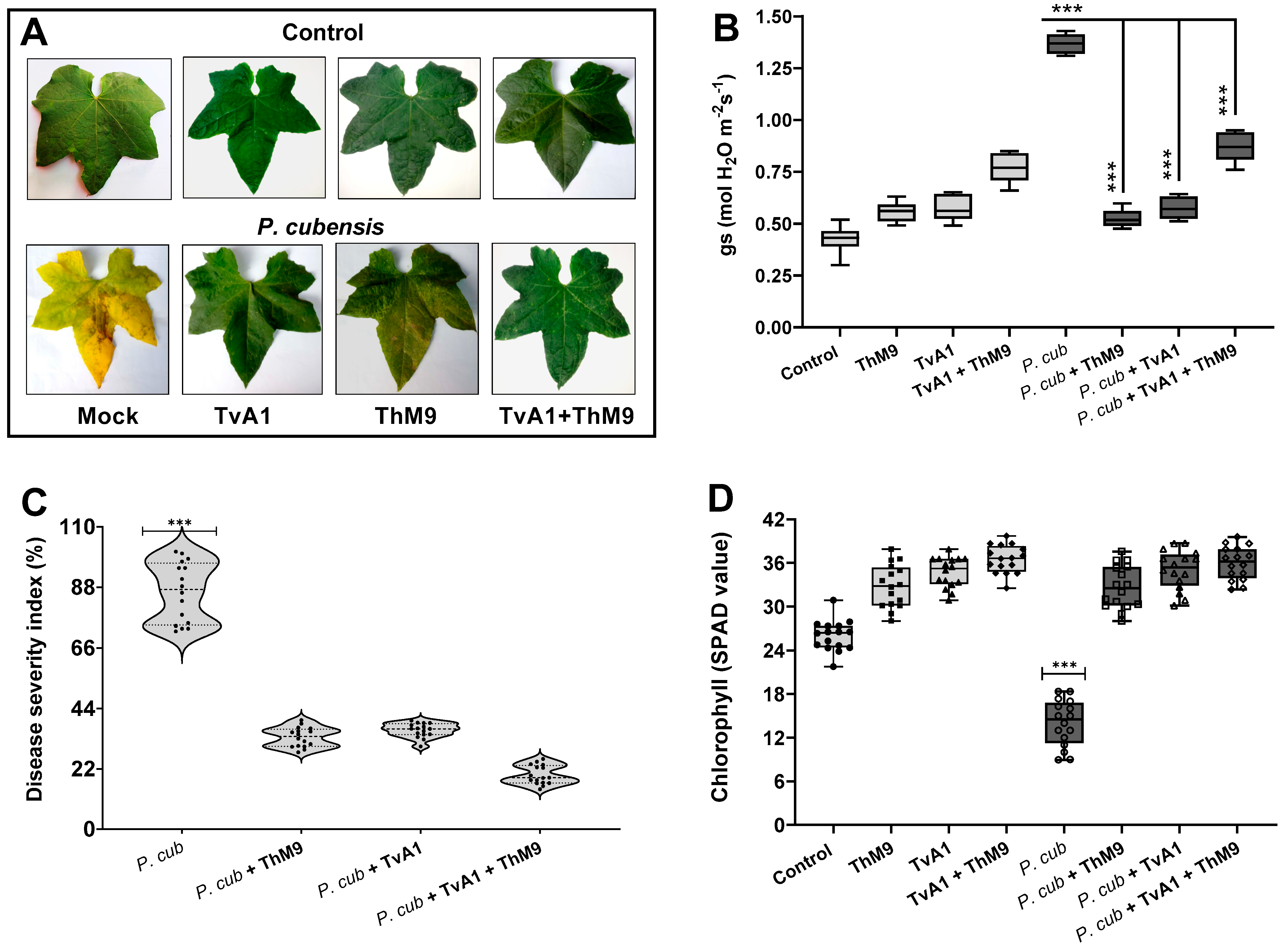
3.9. Effect of ThM9 and TvA1 Isolates on DM Disease Severity in Luffa
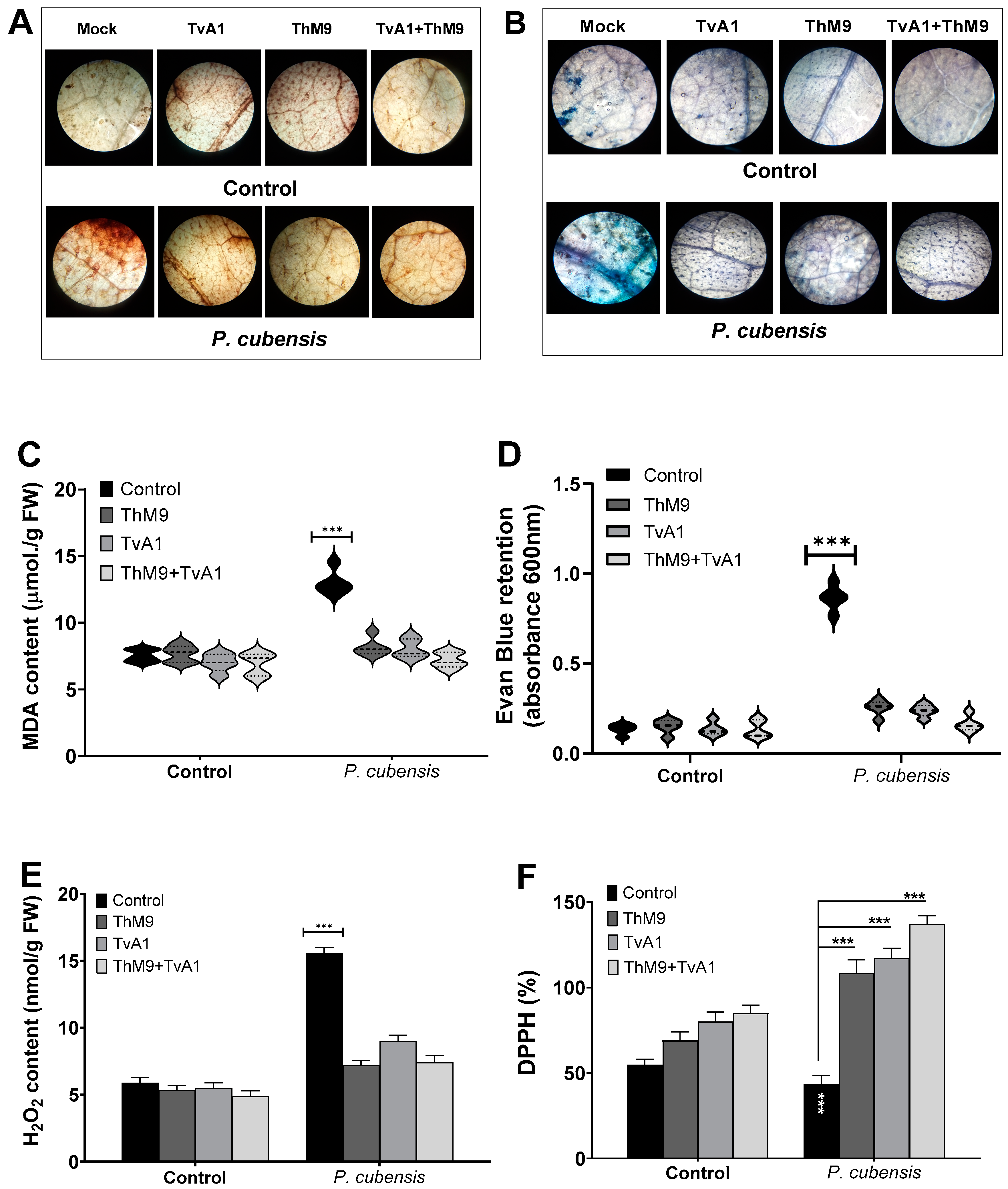
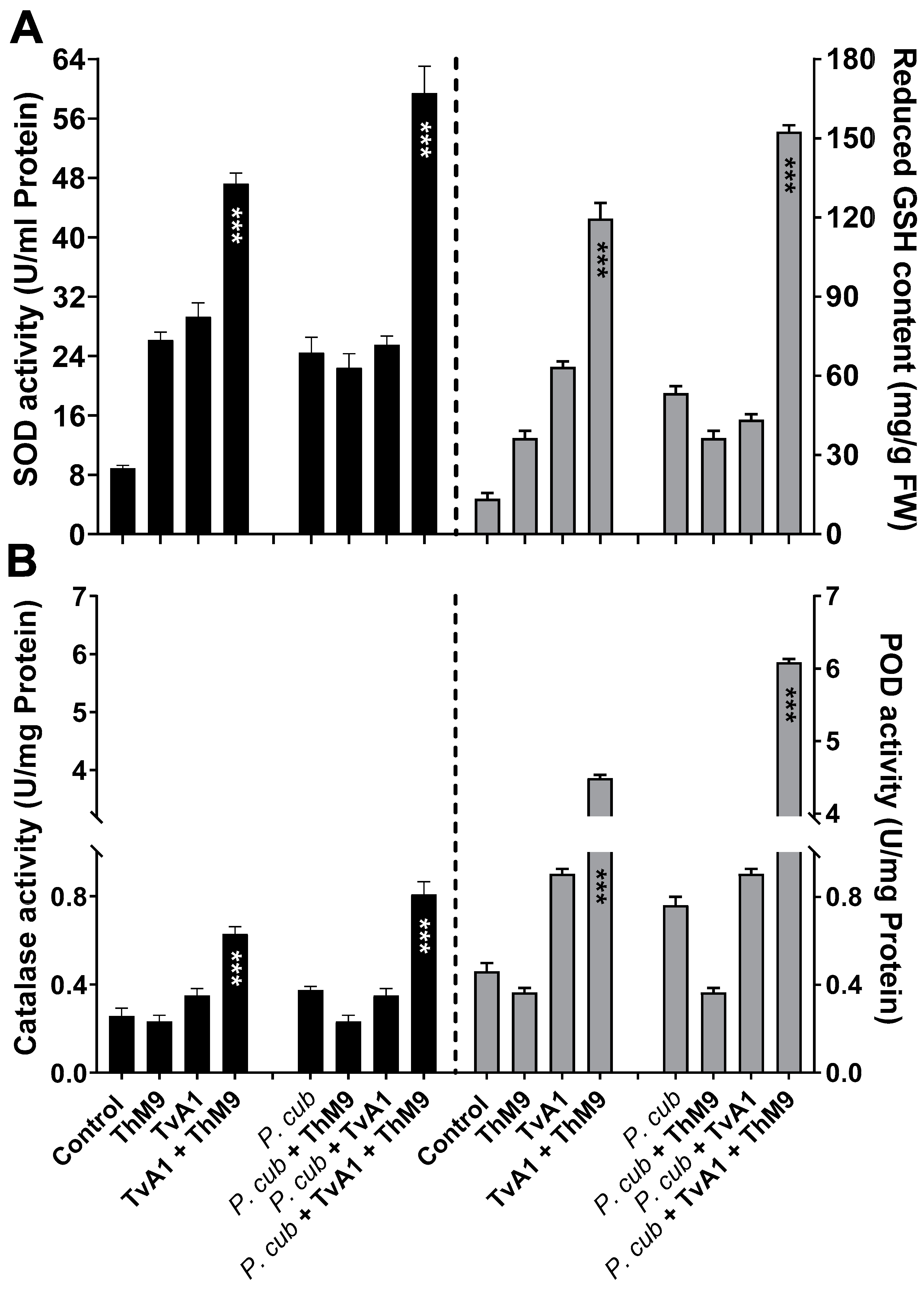
3.10. Effect of ThM9 and TvA1 Isolates on Oxidative Status of Infected Luffa Leaf
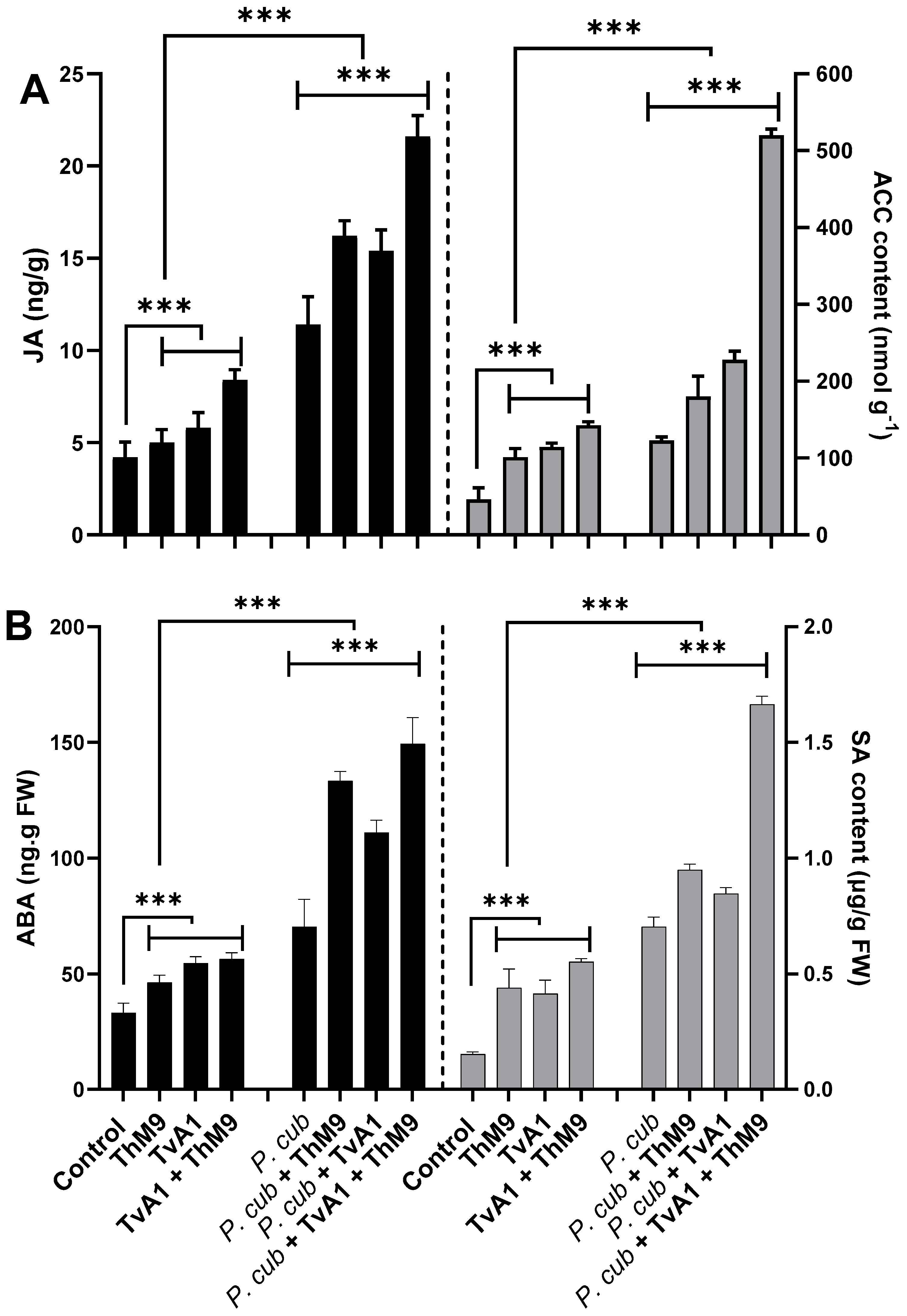
3.11. Effect of ThM9 and TvA1 Isolates on Phytohormonal Content of Infected Luffa Plants
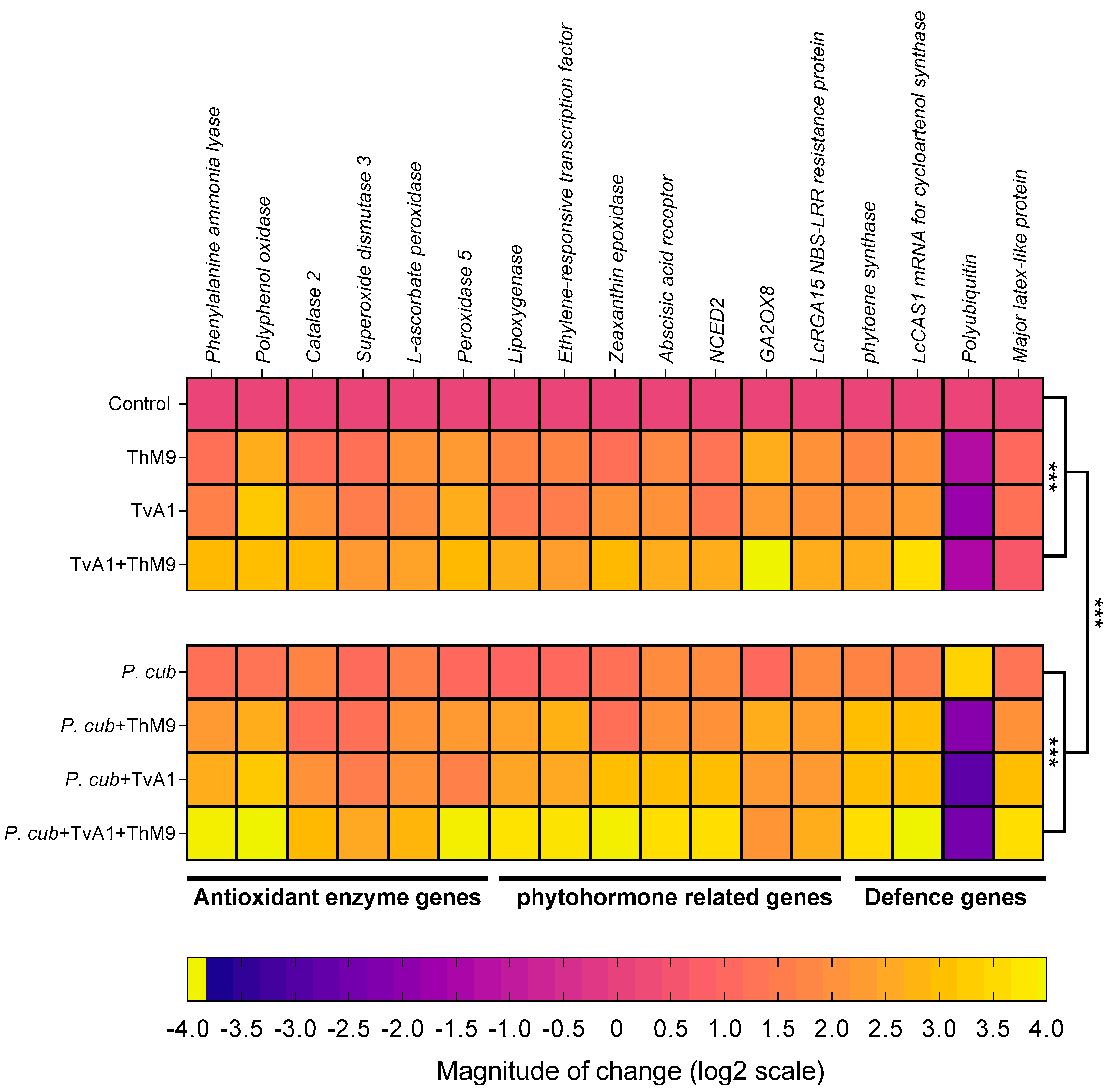
3.12. Effect of ThM9 and TvA1 isolates on gene expression of infected luffa plants
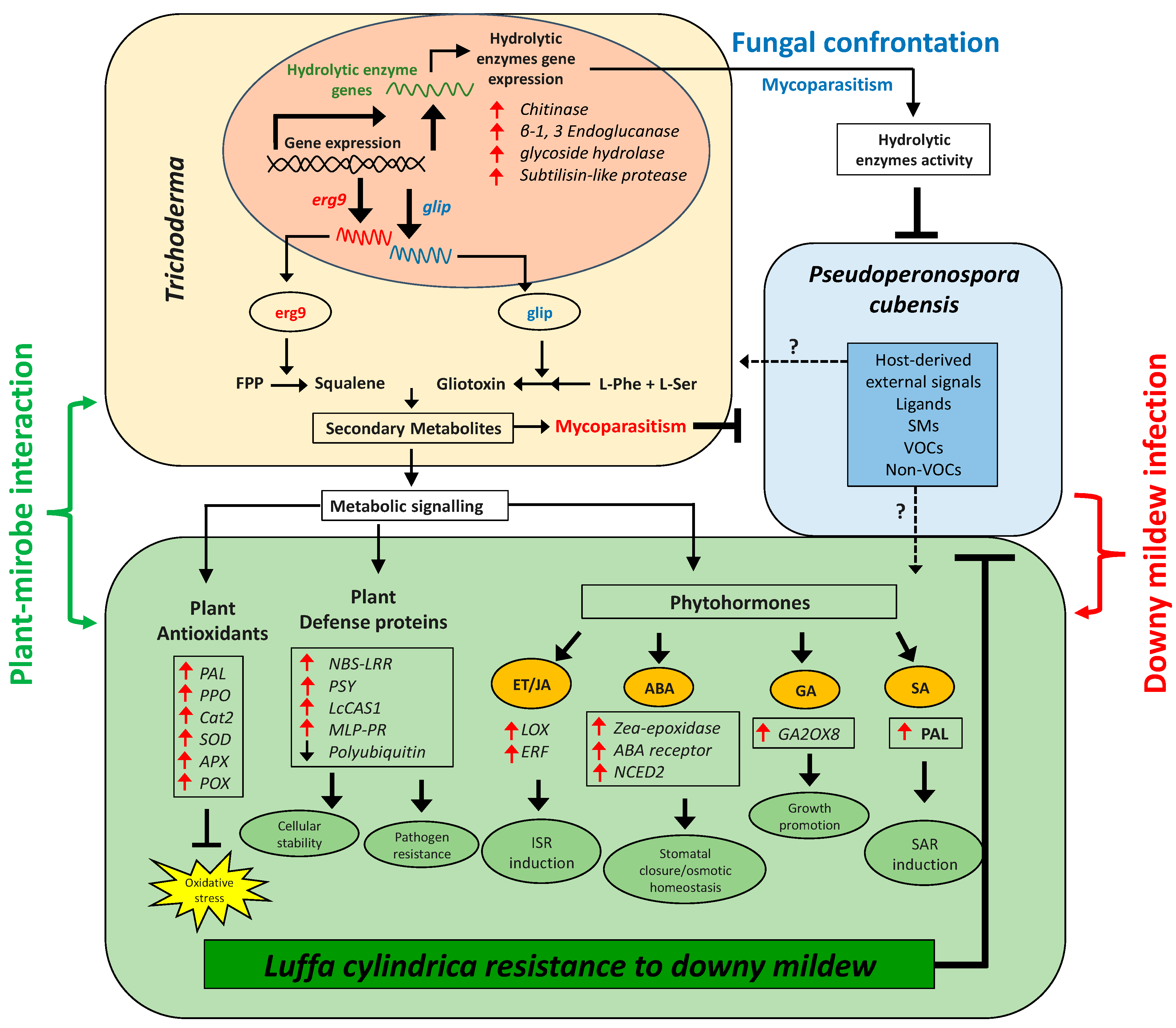
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Ethics Approval and Consent to Participate
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Savory, E.A.; Granke, L.L.; Quesada-ocampo, L.M.; Varbanova, M.; Hausbeck, M.K.; Day, B. The cucurbit downy mildew pathogen Pseudoperonospora cubensis. Mol. Plant Pathol. 2011, 12, 217–226. [Google Scholar] [CrossRef] [PubMed]
- USDA. National Nutrient Database for Standard Reference, Release28. USA Department of Agriculture, Agricultural Research Service. 2015. Available online: http://www.ars.usda.gov/nutrientdata (accessed on 22 April 2022).
- Conrath, U. Molecular aspects of defence priming. Trends Plant Sci. 2011, 16, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.A.A.; Najeeb, S.; Hussain, S.; Xie, B.; Li, Y. Bioactive Secondary Metabolites from Trichoderma spp. against Phytopathogenic Fungi. Microorganisms 2020, 8, 817. [Google Scholar] [CrossRef] [PubMed]
- Nandini, B.; Puttaswamy, H.; Saini, R.K.; Prakash, H.S.; Geetha, N. Trichovariability in rhizosphere soil samples and their biocontrol potential against downy mildew pathogen in pearl millet. Sci. Rep. 2021, 11, 9517. [Google Scholar] [CrossRef] [PubMed]
- Mathys, J.; De Cremer, K.; Timmermans, P.; Van Kerckhove, S.; Lievens, B.; Vanhaecke, M.; Cammue, B.P.A.; De Coninck, B. Genome-Wide Characterization of ISR Induced in Arabidopsis thaliana by Trichoderma hamatum T382 Against Botrytis cinerea Infection. Front. Plant Sci. 2012, 3, 108. [Google Scholar] [CrossRef] [Green Version]
- Alfano, G.; Ivey, M.L.L.; Cakir, C.; Bos, J.I.B.; Miller, S.A.; Madden, L.V.; Kamoun, S.; Hoitink, H.A.J. Systemic Modulation of Gene Expression in Tomato by Trichoderma hamatum 382. Phytopathology 2007, 97, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Studholme, D.J.; Harris, B.; Le Cocq, K.; Winsbury, R.; Perera, V.; Ryder, L.; Beale, M.; Ward, J.; Thornton, C.R.; Grant, M. Investigating the beneficial traits of Trichoderma hamatum GD12 for sustainable agriculture—insights from genomics. Front. Plant Sci. 2013, 4, 258. [Google Scholar] [CrossRef] [Green Version]
- Vitti, A.; Pellegrini, E.; Nali, C.; Lovelli, S.; Sofo, A.; Valerio, M.; Scopa, A.; Nuzzaci, M. Trichoderma harzianum T-22 Induces Systemic Resistance in Tomato Infected by Cucumber mosaic virus. Front. Plant Sci. 2016, 7, 1520. [Google Scholar] [CrossRef] [Green Version]
- Perazzolli, M.; Dagostin, S.; Ferrari, A.; Elad, Y.; Pertot, I. Induction of systemic resistance against Plasmopara viticola in grapevine by Trichoderma harzianum T39 and benzothiadiazole. Biol. Control 2008, 47, 228–234. [Google Scholar] [CrossRef]
- Nagaraju, A.; Sudisha, J.; Murthy, S.M.; Ito, S.I. Seed priming with Trichoderma harzianum isolates enhances plant growth and induces resistance against Plasmopara halstedii, an incitant of sunflower downy mildew disease. Australas. Plant Pathol. 2012, 41, 609–620. [Google Scholar] [CrossRef]
- Szczech, M.; Nawrocka, J.; Felczyński, K.; Małolepsza, U.; Sobolewski, J.; Kowalska, B.; Maciorowski, R.; Jas, K.; Kancelista, A. Trichoderma atroviride TRS25 isolate reduces downy mildew and induces systemic defence responses in cucumber in field conditions. Sci. Hortic. 2017, 224, 17–26. [Google Scholar] [CrossRef]
- Nandini, B.; Puttaswamy, H.; Prakash, H.S.; Adhikari, S.; Jogaiah, S.; Nagaraja, G. Elicitation of Novel Trichogenic-Lipid Nanoemulsion Signaling Resistance Against Pearl Millet Downy Mildew Disease. Biomolecules 2020, 10, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazazzara, V.; Vicelli, B.; Bueschl, C.; Parich, A.; Pertot, I.; Schuhmacher, R.; Perazzolli, M. Trichoderma spp. volatile organic compounds protect grapevine plants by activating defense-related processes against downy mildew. Physiol. Plant. 2021, 172, 1950–1965. [Google Scholar] [CrossRef] [PubMed]
- El-Rahman, A.A.E.-M.A.; El-Shafei, S.M.A.E.-A.; Ivanova, E.V.; Fattakhova, A.N.; Pankova, A.V.; El-Shafei, M.A.E.-A.; El-Morsi, E.-M.A.E.-F.; Alimova, F.K. Cytotoxicity of Trichoderma spp. cultural filtrate against human cervical and breast cancer cell lines. Asian Pac. J. Cancer Prev. 2014, 15, 7229–7234. [Google Scholar] [CrossRef] [Green Version]
- Gomes, E.V.; Ulhoa, C.; Cardoza, R.E.; Silva, R.N.; Gutiérrez, S. Involvement of Trichoderma harzianum Epl-1 Protein in the Regulation of Botrytis Virulence- and Tomato Defense-Related Genes. Front. Plant Sci. 2017, 8, 880. [Google Scholar] [CrossRef]
- Vinale, F.; Nigro, M.; Sivasithamparam, K.; Flematti, G.; Ghisalberti, E.L.; Ruocco, M.; Varlese, R.; Marra, R.; Lanzuise, S.; Eid, A.; et al. Harzianic acid: A novel siderophore from Trichoderma harzianum. FEMS Microbiol. Lett. 2013, 347, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Alkooranee, J.T.; Aledan, T.R.; Xiang, J.; Lu, G.; Li, M. Induced Systemic Resistance in Two Genotypes of Brassica napus (AACC) and Raphanus oleracea (RRCC) by Trichoderma Isolates against Sclerotinia sclerotiorum. Am. J. Plant Sci. 2015, 6, 1662–1674. [Google Scholar] [CrossRef] [Green Version]
- Malmierca, M.G.; McCormick, S.P.; Cardoza, R.E.; Alexander, N.J.; Monte, E.; Gutiérrez, S. Production of trichodiene by Trichoderma harzianum alters the perception of this biocontrol strain by plants and antagonized fungi. Environ. Microbiol. 2015, 17, 2628–2646. [Google Scholar] [CrossRef]
- Malmierca, M.G.; McCormick, S.P.; Cardoza, R.E.; Monte, E.; Alexander, N.J.; Gutiérrez, S. Trichodiene Production in a Trichoderma harzianum erg1-Silenced Strain Provides Evidence of the Importance of the Sterol Biosynthetic Pathway in Inducing Plant Defense-Related Gene Expression. Mol. Plant-Microbe Interact. 2015, 28, 1181–1197. [Google Scholar] [CrossRef] [Green Version]
- Spanova, M.; Zweytick, D.; Lohner, K.; Klug, L.; Leitner, E.; Hermetter, A.; Daum, G. Influence of squalene on lipid particle/droplet and membrane organization in the yeast Saccharomyces cerevisiae. Biochim. Et Biophys. Acta—Mol. Cell Biol. Lipids 2012, 1821, 647–653. [Google Scholar] [CrossRef] [Green Version]
- Siddaiah, C.N.; Satyanarayana, N.R.; Mudili, V.; Gupta, V.K.; Gurunathan, S.; Rangappa, S.; Huntrike, S.S.; Srivastava, R.K. Elicitation of resistance and associated defense responses in Trichoderma hamatum induced protection against pearl millet downy mildew pathogen. Sci. Rep. 2017, 7, srep43991. [Google Scholar] [CrossRef] [PubMed]
- Anisha, C.; Radhakrishnan, E.K. Gliotoxin-Producing Endophytic Acremonium sp. from Zingiber officinale Found Antagonistic to Soft Rot Pathogen Pythium myriotylum. Appl. Biochem. Biotechnol. 2015, 175, 3458–3467. [Google Scholar] [CrossRef] [PubMed]
- Rauf, M.; Awais, M.; Din, A.U.; Ali, K.; Gul, H.; Rahman, M.M.; Hamayun, M.; Arif, M. Molecular Mechanisms of the 1-Aminocyclopropane-1-Carboxylic Acid (ACC) Deaminase Producing Trichoderma asperellum MAP1 in Enhancing Wheat Tolerance to Waterlogging Stress. Front. Plant Sci. 2021, 11, 2213. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J.; Innis, M.A.; Gelfand, D.H.; Sninsky, J.J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- Hasan, H.A.H. Gibberellin and auxin-indole production by plant root-fungi and their biosynthesis under salinity-calcium interaction. Acta Microbiol. Et Immunol. Hung. 2002, 49, 105–118. [Google Scholar] [CrossRef]
- Benizri, E.; Courtade, A.; Picard, C.; Guckert, A. Role of maize root exudates in the production of auxins by Pseudomonas fluorescens M.3.1. Soil Biol. Biochem. 1998, 30, 1481–1484. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Laoutari, S.; Litskas, V.D.; Stavrinides, M.C.; Tzortzakis, N. Effects of water stress on lavender and sage biomass production, essential oil composition and biocidal properties against Tetranychus urticae (Koch). Sci. Hortic. 2016, 213, 96–103. [Google Scholar] [CrossRef]
- Khatiwora, E.; Adsul, V.B.; Kulkarni, M.; Deshpande, N.R.; Kashalkar, R.V. Antibacterial activity of Dibutyl Phthalate: A secondary metabolite isolated from Ipomoea carnea stem. J. Pharm. Res. 2012, 5, 150–152. Available online: http://jprsolutions.info (accessed on 22 April 2022).
- Hata, K.; Futai, K. Endophytic fungi associated with healthy pine needles and needles infested by the pine needle gall midge, Thecodiplosis japonensis. Can. J. Bot. 1995, 73, 384–390. [Google Scholar] [CrossRef]
- Tinker, N.A.; Fortin, M.G.; Mather, D.E. Random amplified polymorphic DNA and pedigree relationships in spring barley. Theor. Appl. Genet. 1993, 85, 976–984. [Google Scholar] [CrossRef]
- Runge, F.; Choi, Y.-J.; Thines, M. Phylogenetic investigations in the genus Pseudoperonospora reveal overlooked species and cryptic diversity in the P. cubensis species cluster. Eur. J. Plant Pathol. 2011, 129, 135–146. [Google Scholar] [CrossRef]
- Zhang, S.; Gan, Y.; Xu, B. Mechanisms of the IAA and ACC-deaminase producing strain of Trichoderma longibrachiatum T6 in enhancing wheat seedling tolerance to NaCl stress. BMC Plant Biol. 2019, 19, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Pan, S.Q. A Technique for Detection of Chitinase, β-1,3-Glucanase, and Protein Patterns after a Single Separation Using Polyacrylamide Gel Electrophoresis or Isoelectrofocusing. Phytopathology 1991, 81, 970. [Google Scholar] [CrossRef]
- Rodriguez-Kabana, R.; Morgan-Jones, G.; Gintis, B.O. The determination of soil chitinase activity: Conditions for assay and ecological studies. Plant Soil 1983, 75, 95–106. [Google Scholar] [CrossRef]
- Li, X.-H.; Yang, H.-J.; Roy, B.; Park, E.Y.; Jiang, L.-J.; Wang, D.; Miao, Y.-G. Enhanced cellulase production of the Trichoderma viride mutated by microwave and ultraviolet. Microbiol. Res. 2010, 165, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Lourien, R.E.; Gusek, T.; Hart, B. Cellulase and protease specific activities of commercially available cellulase preparations. J. Appl. Biochem. 1985, 7, 258–272. [Google Scholar]
- Cardoza, R.E.; Hermosa, M.R.; Vizcaíno, J.A.; González, F.; Llobell, A.; Monte, E.; Gutiérrez, S. Partial silencing of a hydroxy-methylglutaryl-CoA reductase-encoding gene in Trichoderma harzianum CECT 2413 results in a lower level of resistance to lovastatin and lower antifungal activity. Fungal Genet. Biol. 2007, 44, 269–283. [Google Scholar] [CrossRef]
- Ghimire, G.P.; Hei, C.L.; Jae, K.S. Improved squalene production via modulation of the methylerythritol 4-phosphate pathway and heterologous expression of genes from Streptomyces peucetius ATCC 27952 in Escherichia coli. Appl. Environ. Microbiol. 2009, 75, 7291–7293. [Google Scholar] [CrossRef] [Green Version]
- Pramanik, M.H.R.; Nagai, M.; Asao, T.; Matsui, Y. Effects of Temperature and Photoperiod on Phytotoxic Root Exudates of Cucumber (Cucumis sativus) in Hydroponic Culture. J. Chem. Ecol. 2000, 26, 1953–1967. [Google Scholar] [CrossRef]
- Khalaf, E.M.; Raizada, M.N. Bacterial seed endophytes of domesticated cucurbits antagonize fungal and oomycete pathogens including powdery mildew. Front. Microbiol. 2018, 9, 42, Erratum in Front. Microbiol. 2018, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Moloi, M.; van der Westhuizen, A.J. The reactive oxygen species are involved in resistance responses of wheat to the Russian wheat aphid. J. Plant Physiol. 2006, 163, 1118–1125. [Google Scholar] [CrossRef] [PubMed]
- Keutgen, A.J.; Pawelzik, E. Quality and nutritional value of strawberry fruit under long term salt stress. Food Chem. 2008, 107, 1413–1420. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Analytical biochemistry. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Gorin, N.; Heidema, F.T. Peroxidase activity in Golden Delicious apples as a possible parameter of ripening and senescence. Journal of agricultural and food chemistry 1976, 24, 200–201. [Google Scholar] [CrossRef]
- Asada, K. Ascorbate peroxidase–a hydrogen peroxide-scavenging enzyme in plants. Physiologia Plantarum 1992, 85, 235–241. [Google Scholar] [CrossRef]
- Chandlee, J.M.; Scandalios, J.G. Analysis of variants affecting the catalase developmental program in maize scutellum. Theor. Appl. Genet. 1984, 69, 71–77. [Google Scholar] [CrossRef]
- Yu, Y.; Jin, C.; Sun, C.; Wang, J.; Ye, Y.; Zhou, W.; Lu, L.; Lin, X. Inhibition of ethylene production by putrescine alleviates aluminium-induced root inhibition in wheat plants. Sci. Rep. 2016, 6, 18888. [Google Scholar] [CrossRef] [Green Version]
- Perata, P.; Matsukura, C.; Vernieri, P.; Yamaguchi, J. Sugar Repression of a Gibberellin-Dependent Signaling Pathway in Barley Embryos. Plant Cell 1997, 9, 2197–2208. [Google Scholar] [CrossRef]
- Pellegrini, E.; Trivellini, A.; Campanella, A.; Francini, A.; Lorenzini, G.; Nali, C.; Vernieri, P. Signaling molecules and cell death in Melissa officinalis plants exposed to ozone. Plant Cell Rep. 2013, 32, 1965–1980. [Google Scholar] [CrossRef]
- Rauf, M.; Arif, M.; Fisahn, J.; Xue, G.-P.; Balazadeh, S.; Mueller-Roeber, B. NAC Transcription Factor SPEEDY HYPONASTIC GROWTH Regulates Flooding-Induced Leaf Movement in Arabidopsis. Plant Cell 2013, 25, 4941–4955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.-D.; Wang, B.; Li, Y.-P.; Zeng, M.-J.; Liu, J.-T.; Ye, X.-R.; Zhu, H.-S.; Wen, Q.-F. Reference gene selection for qRT-PCR analyses of luffa (Luffa cylindrica) plants under abiotic stress conditions. Sci. Rep. 2021, 11, 3161. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.-S.; Chu, C.; Cheng, C.-W.; Chen, W.-Y.; Tsay, J.-G. Differentiation of two powdery mildews of sunflower (Helianthus annuus) by a PCR-mediated method based on ITS sequences. Eur. J. Plant Pathol. 2008, 121, 1–8. [Google Scholar] [CrossRef]
- Joshi, D.; Singh, P.; Singh, A.K.; Lal, R.J.; Tripathi, N. Antifungal Potential of Metabolites from Trichoderma sp. Against Colletotrichum falcatum Went Causing Red Rot of Sugarcane. Sugar Tech 2016, 18, 529–536. [Google Scholar] [CrossRef]
- Harman, G.E.; Doni, F.; Khadka, R.B.; Uphoff, N. Endophytic strains of Trichoderma increase plants’ photosynthetic capability. J. Appl. Microbiol. 2021, 130, 529–546. [Google Scholar] [CrossRef]
- Savada, R.P.; Ozga, J.A.; Jayasinghege, C.P.A.; Waduthanthri, K.D.; Reinecke, D.M. Heat stress differentially modifies ethylene biosynthesis and signaling in pea floral and fruit tissues. Plant Mol. Biol. 2017, 95, 313–331. [Google Scholar] [CrossRef] [PubMed]
- Kapri, A.; Tewari, L. Phosphate solubilization potential and phosphatase activity of rhizospheric Trichoderma spp. Brazilian Journal of Microbiology 2010, 41, 787–795. [Google Scholar] [CrossRef] [Green Version]
- Mendoza-Mendoza, A.; Zaid, R.; Lawry, R.; Hermosa, R.; Monte, E.; Horwitz, B.A.; Mukherjee, P.K. Molecular dialogues between Trichoderma and roots: Role of the fungal secretome. Fungal Biol. Rev. 2018, 32, 62–85. [Google Scholar] [CrossRef]
- Rubio, M.B.; De Alba, A.E.M.; Nicolás, C.; Monte, E.; Hermosa, R. Early Root Transcriptomic Changes in Wheat Seedlings Colonized by Trichoderma harzianum under Different Inorganic Nitrogen Supplies. Front. Microbiol. 2019, 10, 2444. [Google Scholar] [CrossRef]
- Ben Amira, M.; Lopez, D.; Mohamed, A.T.; Khouaja, A.; Chaar, H.; Fumanal, B.; Gousset-Dupont, A.; Bonhomme, L.; Label, P.; Goupil, P.; et al. Beneficial effect of Trichoderma harzianum strain Ths97 in biocontrolling Fusarium solani causal agent of root rot disease in olive trees. Biol. Control 2017, 110, 70–78. [Google Scholar] [CrossRef]
- Nawrocka, J.; Małolepsza, U.; Szymczak, K.; Szczech, M. Involvement of metabolic components, volatile compounds, PR proteins, and mechanical strengthening in multilayer protection of cucumber plants against Rhizoctonia solani activated by Trichoderma atroviride TRS25. Protoplasma 2018, 255, 359–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes-Figueroa, O.; Ortiz-García, C.F.; Torres-de la Cruz, M.; Lagunes-Espinoza, L.D.C.; Valdovinos-Ponce, G. Especies de trichoderma del agroecosistema cacao con potencial de biocontrol sobre moniliophthora roreri. Rev. Chapingo Ser. Cienc. For. Y Del Ambiente 2016, 22, 149–163. [Google Scholar] [CrossRef]
- Zhang, J.; Bayram; Akcapinar, G.; Atanasova, L.; Rahimi, M.J.; Przylucka, A.; Yang, D.; Kubicek, C.P.; Zhang, R.; Shen, Q.; et al. The neutral metallopeptidase NMP1 of Trichoderma guizhouense is required for mycotrophy and self-defence. Environ. Microbiol. 2016, 18, 580–597. [Google Scholar] [CrossRef] [PubMed]
- Vicente, I.; Baroncelli, R.; Hermosa, R.; Monte, E.; Vannacci, G.; Sarrocco, S. Role and genetic basis of specialised secondary metabolites in Trichoderma ecophysiology. Fungal Biol. Rev. 2022, 39, 83–99. [Google Scholar] [CrossRef]
- Klemptner, R.L.; Sherwood, J.S.; Tugizimana, F.; Dubery, I.A.; Piater, L.A. Ergosterol, an orphan fungal microbe-associated molecular pattern (MAMP). Mol. Plant Pathol. 2014, 15, 747–761. [Google Scholar] [CrossRef]
- Ruan, J.; Zhou, Y.; Zhou, M.; Yan, J.; Khurshid, M.; Weng, W.; Cheng, J.; Zhang, K. Jasmonic Acid Signaling Pathway in Plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef] [Green Version]
- Garaiová, M.; Zambojová, V.; Šimová, Z.; Griač, P.; Hapala, I. Squalene epoxidase as a target for manipulation of squalene levels in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2013, 14, 310–323. [Google Scholar] [CrossRef] [Green Version]
- Payyavula, R.S.; Navarre, D.A.; Kuhl, J.C.; Pantoja, A.; Pillai, S.S. Differential effects of environment on potato phenylpropanoid and carotenoid expression. BMC Plant Biol. 2012, 12, 39. [Google Scholar] [CrossRef] [Green Version]
- Balibar, C.J.; Walsh, C.T. GliP, a Multimodular Nonribosomal Peptide Synthetase in Aspergillus fumigatus, Makes the Diketopiperazine Scaffold of Gliotoxin. Biochemistry 2006, 45, 15029–15038. [Google Scholar] [CrossRef]
- Howell, C.R. Mechanisms Employed by Trichoderma Species in the Biological Control of Plant Diseases: The History and Evolution of Current Concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Dhingra, S.; Andes, D.; Calvo, A.M. VeA Regulates Conidiation, Gliotoxin Production, and Protease Activity in the Opportunistic Human Pathogen Aspergillus fumigatus. Eukaryot. Cell 2012, 11, 1531–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermosa, R.; Rubio, M.B.; Cardoza, R.E.; Nicolás, C.; Monte, E.; Gutiérrez, S. The contribution of Trichoderma to balancing the costs of plant growth and defence. Int. Microbiol. 2013, 16, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, K.; Fan, L.; Fu, K.; Yu, C.; Wang, M.; Xia, H.; Sun, J.; Li, Y.; Chen, J. Cellulase from Trichoderma harzianum interacts with roots and triggers induced systemic resistance to foliar disease in maize. Sci. Rep. 2016, 6, 35543. [Google Scholar] [CrossRef] [PubMed]
- Pocurull, M.; Fullana, A.M.; Ferro, M.; Valero, P.; Escudero, N.; Saus, E.; Gabaldón, T.; Sorribas, F.J. Commercial Formulates of Trichoderma Induce Systemic Plant Resistance to Meloidogyne incognita in Tomato and the Effect Is Additive to That of the Mi-1.2 Resistance Gene. Front. Microbiol. 2020, 10, 3042. [Google Scholar] [CrossRef]
- Kinkema, M.; Fan, W.; Dong, X. Nuclear localization of NPR1 is required for activation of PR gene expression. Plant Cell 2000, 12, 2339–2350. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.J.; Leon-Reyes, A.; Van der Ent, S.; Van Wees, S.C.M. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Niu, D.-D.; Liu, H.-X.; Jiang, C.-H.; Wang, Y.-P.; Wang, Q.-Y.; Jin, H.-L.; Guo, J.-H. The Plant Growth–Promoting Rhizobacterium Bacillus cereus AR156 Induces Systemic Resistance in Arabidopsis thaliana by Simultaneously Activating Salicylate- and Jasmonate/Ethylene-Dependent Signaling Pathways. Mol. Plant-Microbe Interact. 2011, 24, 533–542. [Google Scholar] [CrossRef] [Green Version]
- Elsharkawy, M.M.; Shimizu, M.; Takahashi, H.; Ozaki, K.; Hyakumachi, M. Induction of Systemic Resistance against Cucumber mosaic virus in Arabidopsis thaliana by Trichoderma asperellum SKT-1. Plant Pathol. J. 2013, 29, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Hao, Q.; Wang, W.; Han, X.; Wu, J.; Lyu, B.; Chen, F.; Caplan, A.; Li, C.; Wu, J.; Wang, W.; et al. Isochorismate-based salicylic acid biosynthesis confers basal resistance to Fusarium graminearum in barley. Mol. Plant Pathol. 2018, 19, 1995–2010. [Google Scholar] [CrossRef] [Green Version]
- Shine, M.B.; Yang, J.W.; El-Habbak, M.; Nagyabhyru, P.; Fu, D.Q.; Navarre, D.; Ghabrial, S.; Kachroo, P.; Kachroo, A. Cooperative functioning between phenylalanine ammonia lyase and isochorismate synthase activities contributes to salicylic acid biosynthesis in soybean. New Phytol. 2016, 212, 627–636. [Google Scholar] [CrossRef] [Green Version]
- Nugroho, L.H.; Verberne, M.C.; Verpoorte, R. Activities of enzymes involved in the phenylpropanoid pathway in constitutively salicylic acid-producing tobacco plants. Plant Physiol. Biochem. 2002, 40, 755–760. [Google Scholar] [CrossRef]
- Chaman, M.E.; Copaja, S.V.; Argandoña, V.H. Relationships between Salicylic Acid Content, Phenylalanine Ammonia-lyase (PAL) Activity, and Resistance of Barley to Aphid Infestation. J. Agric. Food Chem. 2003, 51, 2227–2231. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Yao, Y.; Cai, Y.; Lin, Y. Molecular Cloning and Sequence Analysis of a Phenylalanine Ammonia-Lyase Gene from Dendrobium. PLoS ONE 2013, 8, e62352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fesel, P.H.; Zuccaro, A. β-glucan: Crucial component of the fungal cell wall and elusive MAMP in plants. Fungal Genet. Biol. FG & B 2016, 90, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Wanke, A.; Malisic, M.; Wawra, S.; Zuccaro, A. Unraveling the sugar code: The role of microbial extracellular glycans in plant–microbe interactions. J. Exp. Bot. 2021, 72, 15–35. [Google Scholar] [CrossRef]
- El-Katatny, M.H.; Gudelj, M.; Robra, K.-H.; Elnaghy, M.A.; Gübitz, G.M. Characterization of a chitinase and an endo-β-1,3-glucanase from Trichoderma harzianum Rifai T24 involved in control of the phytopathogen Sclerotium rolfsii. Appl. Microbiol. Biotechnol. 2001, 56, 137–143. [Google Scholar] [CrossRef]
- Sivan, A.; Chet, I. Degradation of Fungal Cell Walls by Lytic Enzymes of Trichoderma harzianum. Microbiology 1989, 135, 675–682. [Google Scholar] [CrossRef] [Green Version]
- Rombouts, F.M.; Phaff, H.J. Lysis of Yeast Cell Walls Lytic β-(1→6)-Glucanase from Bacillus circulans WL-12’. Eur. J. Biochem. 1976, 63, 109–120. [Google Scholar] [CrossRef]
- Ramada, M.H.S.; Steindorff, A.S.; Bloch, C., Jr.; Ulhoa, C.J. Secretome analysis of the mycoparasitic fungus Trichoderma harzianum ALL 42 cultivated in different media supplemented with Fusarium solani cell wall or glucose. Proteomics 2015, 16, 477–490. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.A.D.; Suzuki, N.; Ciftci-Yilmaz, S.U.L.T.A.N.; Mittler, R.O.N. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Floryszak-Wieczorek, J.; Górski, Z.; Arasimowicz-Jelonek, M. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Eur. J. Plant Pathol. 2011, 1302, 373–399. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Martínez, A.; Fortea, M.I.; Del Amor, F.M.; Núñez-Delicado, E. Kinetic characterisation and thermal inactivation study of partially purified red pepper (Capsicum annuum L.) peroxidase. Food Chem. 2008, 107, 193–199. [Google Scholar] [CrossRef]
- Bielski, B.H.J.; Cabelli, D.E.; Arudi, R.L.; Ross, A.B. Reactivity of HO2/O−2 Radicals in Aqueous Solution. J. Phys. Chem. Ref. Data 1985, 14, 1041–1100. [Google Scholar] [CrossRef]
- Wu, H.; Zhao, G.; Gong, H.; Li, J.; Luo, C.; He, X.; Luo, S.; Zheng, X.; Liu, X.; Guo, J.; et al. A high-quality sponge gourd (Luffa cylindrica) genome. Hortic. Res. 2020, 7, 128. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.; Ewan, R.; Mesmar, J.; Gudipati, V.; Sadanandom, A. E3 ubiquitin ligases and plant innate immunity. J. Exp. Bot. 2009, 60, 1123–1132. [Google Scholar] [CrossRef] [Green Version]
- Hua, Z.; Vierstra, R.D. The cullin-RING ubiquitin-protein ligases. Annu. Rev. Plant. Biol 2011, 62, 299–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, D.R. E3 Ubiquitin Ligases: Key Regulators of Hormone Signaling in Plants. Mol. Cell. Proteom. 2018, 17, 1047–1054. [Google Scholar] [CrossRef] [Green Version]
- Miricescu, A.; Goslin, K.; Graciet, E. Ubiquitylation in plants: Signaling hub for the integration of environmental signals. J. Exp. Bot. 2018, 69, 4511–4527. [Google Scholar] [CrossRef]
- Wang, X.; Huang, M.; Zhou, Q.; Cai, J.; Dai, T.; Cao, W.; Jiang, D. Physiological and proteomic mechanisms of waterlogging priming improves tolerance to waterlogging stress in wheat (Triticum aestivum L.). Environ. Exp. Bot. 2016, 132, 175–182. [Google Scholar] [CrossRef]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749–759, Erratum in Nat. Rev. Microbiol. 2011, 9, 896. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Gas-Pascual, E.; Berna, A.; Bach, T.J.; Schaller, H. Plant Oxidosqualene Metabolism: Cycloartenol Synthase–Dependent Sterol Biosynthesis in Nicotiana benthamiana. PLoS ONE 2014, 9, e109156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babiychuk, E.; Bouvier-Navé, P.; Compagnon, V.; Suzuki, M.; Muranaka, T.; Van Montagu, M.; Kushnir, S.; Schaller, H. Allelic mutant series reveal distinct functions for Arabidopsis cycloartenol synthase 1 in cell viability and plastid biogenesis. Proc. Natl. Acad. Sci. USA 2008, 105, 3163–3168. [Google Scholar] [CrossRef] [Green Version]
- Morita, M.; Shibuya, M.; Kushiro, T.; Masuda, K.; Ebizuka, Y. Molecular cloning and functional expression of triterpene synthases from pea (Pisum sativum): New α-amyrin-producing enzyme is a multifunctional triterpene synthase. Eur. J. Biochem. 2000, 267, 3453–3460. [Google Scholar] [CrossRef]
- Hayashi, H.; Huang, P.; Inoue, K.; Hiraoka, N.; Ikeshiro, Y.; Yazaki, K.; Tanaka, S.; Kushiro, T.; Shibuya, M.; Ebizuka, Y. Molecular cloning and characterization of isomultiflorenol synthase, a new triterpene synthase from Luffa cylindrica, involved in biosynthesis of bryonolic acid. Eur. J. Biochem. 2001, 268, 6311–6317. [Google Scholar] [CrossRef]
- Meyer, M.M.; Xu, R.; Matsuda, S.P.T. Directed Evolution To Generate Cycloartenol Synthase Mutants that Produce Lanosterol. Org. Lett. 2002, 4, 1395–1398. [Google Scholar] [CrossRef]
- Bode, H.; Zeggel, B.; Silakowski, B.; Wenzel, S.C.; Reichenbach, H.; Müller, R. Steroid biosynthesis in prokaryotes: Identification of myxobacterial steroids and cloning of the first bacterial 2,3(S)-oxidosqualene cyclase from the myxobacterium Stigmatella aurantiaca. Mol. Microbiol. 2003, 47, 471–481. [Google Scholar] [CrossRef]
- Deng, Y.; Ning, Y.; Yang, D.-L.; Zhai, K.; Wang, G.-L.; He, Z. Molecular Basis of Disease Resistance and Perspectives on Breeding Strategies for Resistance Improvement in Crops. Mol. Plant 2020, 13, 1402–1419. [Google Scholar] [CrossRef]
- Zhou, B.; Zeng, L. The Tomato U-Box Type E3 Ligase PUB13 Acts With Group III Ubiquitin E2 Enzymes to Modulate FLS2-Mediated Immune Signaling. Front. Plant Sci. 2018, 9, 615. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Song, Y.; Liu, J.; Xu, G.; Dou, D. Ubiquitination of Receptorsomes, Frontline of Plant Immunity. Int. J. Mol. Sci. 2022, 9, 2937. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Accession | Gene Description | Primer Sequences |
|---|---|---|
| P. cubensisPrimers for PCR analysis | ||
| JF414553.1 | Cytochrome c oxidase subunit II (cox2) | F-TAATTGTAGTTACAGTATTC |
| Clade 1 R-GTAATTAATACTCGAATATGG | ||
| Clade 1/2 R GTAAAACATCAGAAGCTGTG | ||
| T. virens Primers for PCR analysis | ||
| JN039096.1 | Translation elongation factor 1-like (tef1) | F-ACACAGCTAACCATTCGCCA |
| R-TCCTTGGTTTAGCACTGGGC | ||
| T. harzianum Primers for PCR analysis | ||
| MW407164.1 | RNA polymerase II (RPB2) | F-GCAGGAAGATGACCCGGAAA |
| R-TGGAAGGGTGGACAACATGC | ||
| T. virens Primers for RT-qPCR | ||
| KP641615.1 | Chitinase (ech42) | F-GATGACACTCAGGCCACCAA |
| R-TGGCAAACAAGTTGGCATCG | ||
| MG702349.1 | Endoglucanase (EG4) | F-GGTCTCATCAGTGGCGGAAA |
| R-AATGAGTTCGTGGCGTAGCA | ||
| XM_014103795.1 | Subtilisin-like protease | F-CATCCGATACCGAGCACGAA |
| R-TGGCCTTGGTTCCAAAGGTT | ||
| NW_014013747.1 | Cellulase/glycoside hydrolase | F-CCAGCGAGCATAGTTGTGGA |
| R-ACTCTGGACGCCACCAATTT | ||
| XM_014100723.1 | Non-ribosomal peptide synthetase GliP(NRPS dioxopiperazine synthetase) | F-CAAGGAAAACTTGTGGGCCG |
| R-AGGGAGGAGAGCTGGTAGTG | ||
| FJ442590.1 | Actin (ACT) | F-GTCCTTGGTCTTGAGAGCGG |
| R-GAATGCAATTAGCGCCACTG | ||
| T. harzianum Primers for RT-qPCR | ||
| MG601052.1 | Chitinase | F-TAACTACTCCAAGCTGCGCC |
| R-GAATCGGTGTTGAAGGGGGT | ||
| MG702349.1 | Endoglucanase (EG4) | F-GGTCTCATCAGTGGCGGAAA |
| R-AATGAGTTCGTGGCGTAGCA | ||
| KC876057.1 | Subtilisin-like serine protease | F-CATCAACGACGTCCAGACCA |
| R-TCTTGGTTCTCGTTACCGGC | ||
| NW_020209251.1 | Cellulase/glycoside hydrolase | F-TGTCATGGGGTAGCAAAACGA |
| R-ACTCGATGTGGAAGACAGGC | ||
| XM_024916014.1 | Squalene synthase (SQS1/ERG9) | F-TGGAATGGCAACAAAGTTGA |
| R-AAGGAGATTTGGAGCAAGCA | ||
| FJ442452.1 | Actin (ACT) | F-TGTCCTTGGTCTTGAGAGCG |
| R-CGTAAAGCTGTGCGACTCAAT | ||
| Luffaaegyptiaca Primers for RT-qPCR | ||
| MN548044 | Elongation factor 1α (EF-1α) | F-TCAAGAAGGTCGGATACA |
| F-ACAGGGACAGTTCCAATAC | ||
| KP341758.1 | Phenylalanine ammonia-lyase; PAL | F-CTGGGTGATGGAGAGCATGA |
| R-CTCTTGTTGCTGAGTGAGGC | ||
| KM506755.1 | Peroxidase; POD | F-CGCTCTATCTGGGGCACATA |
| R-AATGTGTCCGGTGTTGTTGG | ||
| KR819890.1 | Polyphenol oxidase; PPO2 | F-AAGACGTCGCAGCTCAAATC |
| R-CGTGCCATCATCAAGAACGT | ||
| KR184674.1 | Catalase 2; CAT2 | F-GCGATTTGTGGAAGCGTTAT |
| R-ATCCAATCACGTTGGCTTTC | ||
| KX092448.1 | Superoxide dismutase 3; SOD3 | F-CCGAGCAGAGTGTTTTGTGA |
| RACCCTACAGGGGGATTTGAC | ||
| KX092439.1 | L-ascorbate peroxidase; APX | F-GGGGTTGTTGCTGTTGAAGT |
| R-GATTGGTTGTCCAAGCACCT | ||
| KX092434.1 | Peroxidase 5; POD5 | F-CCCCTGGAGTTGTTGCTTTA |
| R-GCAACCCCTTGTTGTCAAGT | ||
| MF678593.1 | Gibberellin 2-beta-dioxygenase 8-like protein; GA2OX8 (Gibberellin 2-beta-dioxygenase 8—Arabidopsis, involved in the pathway gibberellin biosynthesis) | F-GCGGATTCCAACAGAGAGAG |
| R-CGGTAATCAGCCAATCTGGT | ||
| MF678591.1 | Ethylene-responsive transcription factor; ERF | F-CCGGAGCTGGTAAAATCAAA |
| R-CTCGTGGGGTTTGAAAAAGA | ||
| KX092444.1 | Lipoxygenase; LOX (jasmonic acid biosynthesis) | F-ATTCGGGGTAATGGAAAAGG |
| R-GGAACATAAACCGGATGTGG | ||
| MK649987.1 | Zeaxanthin epoxidase; ZEP (ABA biosynthesis enzyme) | F-AACCGACCGAGTTTATCACG |
| R-AATCCAGCGGAGGAAAGATT | ||
| KX092441.1 | 9-cis-epoxycarotenoid dioxygenase; NCED2 (ABA biosynthesis) | F-AAGACGATTTGCCGTACCAC |
| R-TTCGAGGTCTTTGGATTTGG | ||
| MF678592.1 | Abscisic acid receptor | F-GTCGTTTGTTGTGGATGTGC |
| R-TGGAAAAATCGATTGGGAAG | ||
| LC177373.1 | Major latex-like protein 3; MLP3 (positive regulator of ABA responses and induces drought tolerance and pathogen resistance) | F-TTTCGGGGATTTGGTCAATA |
| R-GGCTTCACCAGATCTCCATC | ||
| JN230655.1 | NBS-LRR resistance protein gene (pathogen resistant) | F-GGCAAGAAGTCGTGATTGGT |
| R-CTAAGGGCAATCCTCCACAG | ||
| KX092450.1 | Phytoene synthase; PSY (phytoene synthase is a transferase enzyme involved in the iosynthesis of carotenoids) | F-TTCAAATTCCCGATTCTTCG |
| R-TTCACTAGAGCCGCCTGTTT | ||
| AB033334.1 | LcCAS1 mRNA for cycloartenol synthase (biosynthesis of steroids) | F-AATCCACTTCCTCCCGAACT |
| R-CACACTCATTGCGAGCCTTA | ||
| KR349345.1 | Polyubiquitin; Ubi | F-GTCAACCCTCCACCTTGTGT |
| R-TCCAGCGAAGATCAACCTCT | ||
| Inoculations | Inoculum Concentration | Germination (%) | Seedling Vigor |
|---|---|---|---|
| Control | Non-inoculated | 84 ± 2.013 a | 1865 ± 2.013 a |
| ThM9 | 1 × 108 CFU mL−1 | 88 ± 2.014 ab | 1895 ± 2.013 ab |
| TvA1 | 1 × 108 CFU mL−1 | 89 ± 2.014 ab | 1896 ± 2.013 ab |
| ThM9 + TvA1 | (1 × 108 CFU mL−1) + (1 × 108 CFU mL−1) | 97 ± 2.014 b | 1983 ± 2.013 b |
| Inoculations | Releasing Ratio (%) | Releasing Inhibition Ratio (%) * |
|---|---|---|
| Control | 89.3 | --- |
| ThM9 | 22.6 | 79.1 a |
| TvA1 | 24.3 | 81.4 b |
| ThM9 + TvA1 | 11.1 | 95.1 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rauf, M.; Ur-Rahman, A.; Arif, M.; Gul, H.; Ud-Din, A.; Hamayun, M.; Lee, I.-J. Immunomodulatory Molecular Mechanisms of Luffa cylindrica for Downy Mildews Resistance Induced by Growth-Promoting Endophytic Fungi. J. Fungi 2022, 8, 689. https://doi.org/10.3390/jof8070689
Rauf M, Ur-Rahman A, Arif M, Gul H, Ud-Din A, Hamayun M, Lee I-J. Immunomodulatory Molecular Mechanisms of Luffa cylindrica for Downy Mildews Resistance Induced by Growth-Promoting Endophytic Fungi. Journal of Fungi. 2022; 8(7):689. https://doi.org/10.3390/jof8070689
Chicago/Turabian StyleRauf, Mamoona, Asim Ur-Rahman, Muhammad Arif, Humaira Gul, Aziz Ud-Din, Muhammad Hamayun, and In-Jung Lee. 2022. "Immunomodulatory Molecular Mechanisms of Luffa cylindrica for Downy Mildews Resistance Induced by Growth-Promoting Endophytic Fungi" Journal of Fungi 8, no. 7: 689. https://doi.org/10.3390/jof8070689