
Characterization and Functional Analysis of a New Calcium/Calmodulin-Dependent Protein Kinase (CaMK1) in the Citrus Pathogenic Fungus Penicillium italicum

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Cultivation Conditions
2.2. Gene Cloning and Sequence Analysis
2.3. Gene Knockout and Complementation of PiCaMK1
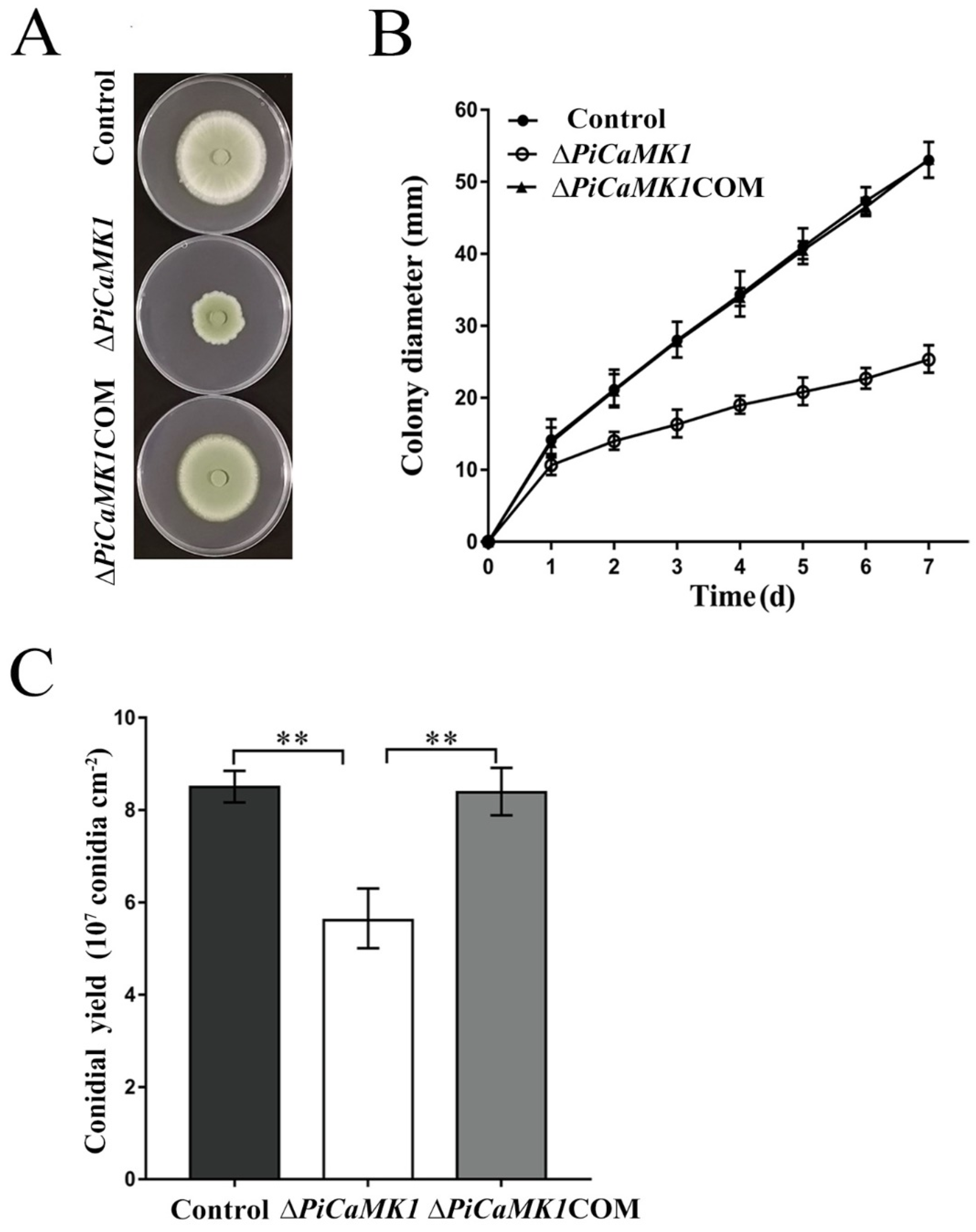
2.4. Vegetative Growth, Conidiation, and Virulence Experiments
2.5. Abiotic Stress Experiments
2.6. Analysis of PiCaMK1-Mediated Transcriptomes
2.7. Real-Time Quantitative PCR (RT-qPCR)
3. Results
3.1. Cloning and Sequence Analysis of PiCaMK1 Gene
3.2. Knockout of PiCaMK1 Gene and Complementation in P. italicum
3.3. PiCaMK1 Is Required for Vegetative Growth and Conidiation
3.4. PiCaMK1 Is Required for Full Virulence
3.5. PiCaMK1 Has No Contribution to DMI-Fungicide Resistance
3.6. The Role of PiCaMK1 in Stress Tolerance of P. italicum
3.7. Transcriptome Analysis and KEGG Enrichment of DEGs
3.8. RT-qPCR Validation of DEGs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Iqbal, Z.; Iqbal, J.; Abbas, I.; Kamran, M. Innovative strategies for eco-friendly management of citrus blue mold disease caused by Penicillium italicum WHEMER. J. Int. Sci. Publ. 2017, 5, 361–365. [Google Scholar]
- Plaza, P.; Usall, J.; Teixidó, N.; Viñas, I. Effect of water activity and temperature on germination and growth of Penicillium digitatum, P. italicum and Geotrichum candidum. J. Appl. Microbiol. 2003, 94, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Kanashiro, A.M.; Akiyama, D.Y.; Kupper, K.C.; Fill, T.P. Penicillium italicum: An underexplored postharvest pathogen. Front. Microbiol. 2020, 11, 606852. [Google Scholar] [CrossRef]
- Louw, J.P.; Korsten, L. Pathogenicity and host susceptibility of Penicillium spp. on citrus. Plant Dis. 2015, 99, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papoutsis, K.; Mathioudakis, M.M.; Hasperué, J.H.; Ziogas, V. Non-chemical treatments for preventing the postharvest fungal rotting of citrus caused by Penicillium digitatum (Green Mold) and Penicillium italicum (Blue Mold). Trends Food Sci. Technol. 2019, 86, 479–491. [Google Scholar] [CrossRef]
- Zhang, T.; Xu, Q.; Sun, X.; Li, H. The calcineurin-responsive transcription factor Crz1 is required for conidation, full virulence and DMI resistance in Penicillium digitatum. Microbiol. Res. 2013, 168, 211–222. [Google Scholar] [CrossRef]
- de Ramón-Carbonell, M.; Sánchez-Torres, P. The transcription factor PdSte12 contributes to Penicillium digitatum virulence during citrus fruit infection. Postharvest Biol. Technol. 2017, 125, 129–139. [Google Scholar] [CrossRef]
- de Ramón-Carbonell, M.; Sánchez-Torres, P. Penicillium digitatum MFS transporters can display different roles during pathogen-fruit interaction. Int. J. Food Microbiol. 2021, 337, 108918. [Google Scholar] [CrossRef]
- de Ramón-Carbonell, M.; Sánchez-Torres, P. Unveiling the role displayed by Penicillium digitatum PdMut3 transcription factor in pathogen-fruit interaction. J. Fungi 2021, 7, 828. [Google Scholar] [CrossRef]
- Costa, J.H.; Bazioli, J.M.; de Moraes-Pontes, J.G.; Fill, T.P. Penicillium digitatum infection mechanisms in citrus: What do we know so far? Fungal Biol. 2019, 123, 584–593. [Google Scholar] [CrossRef]
- Ruan, R.; Chen, Y.; Li, H.; Wang, M. Functional diversification of sterol regulatory element binding proteins following gene duplication in a fungal species. Fungal Genet. Biol. 2019, 131, 103239. [Google Scholar] [CrossRef] [PubMed]
- Gandía, M.; Garrigues, S.; Hernanz-Koers, M.; Manzanares, P.; Marcos, J.F. Differential roles, crosstalk and response to the Antifungal Protein AfpB in the three Mitogen-Activated Protein Kinases (MAPK) pathways of the citrus postharvest pathogen Penicillium digitatum. Fungal Genet. Biol. 2019, 124, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Larralde-Corona, C.P.; Ramírez-González, M.S.; Pérez-Sánchez, G.; Oliva-Hernández, A.A.; Narváez-Zapata, J.A. Identification of differentially expressed genes in the citrus epiphytic-yeast Pichia guilliermondii during interaction with Penicillium digitatum. Biol. Control 2011, 57, 208–214. [Google Scholar] [CrossRef]
- Liu, P.; Cheng, Y.; Yang, M.; Liu, Y.; Chen, K.; Long, C.; Deng, X. Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of Penicillium molds of citrus fruits. BMC Microbiol. 2014, 14, 242. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Sun, X.; Zhu, C.; Xu, Q.; Ruan, R.; Yu, D.; Li, H. PdbrlA, PdabaA and PdwetA control distinct stages of conidiogenesis in Penicillium digitatum. Res. Microbiol. 2015, 166, 56–65. [Google Scholar] [CrossRef]
- Wan, C.; Shen, Y.; Nisar, M.F.; Qi, W.; Chen, C.; Chen, J. The antifungal potential of carvacrol against Penicillium digitatum through 1H-NMR based metabolomics approach. Appl. Sci. 2019, 9, 2240. [Google Scholar] [CrossRef] [Green Version]
- de Ramón-Carbonell, M.; López-Pérez, M.; González-Candelas, L.; Sánchez-Torres, P. PdMFS1 transporter contributes to Penicilliun digitatum fungicide resistance and fungal virulence during citrus fruit infection. J. Fungi 2019, 5, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Wang, Y.; Hu, X.; Lei, M.; Wang, M.; Zeng, J.; Li, H.; Liu, Z.; Zhou, T.; Yu, D. Involvement of LaeA in the regulation of conidia production and stress responses in Penicillium digitatum. J. Basic Microbiol. 2020, 60, 82–88. [Google Scholar] [CrossRef]
- Lin, S.; Wang, Y.; Lu, Q.; Zhang, B.; Wu, X. Combined transcriptome and metabolome analyses reveal the potential mechanism for the inhibition of Penicillium digitatum by X33 antimicrobial oligopeptide. Bioresour. Bioprocess. 2021, 8, 120. [Google Scholar] [CrossRef]
- Gong, L.; Liu, Y.; Xiong, Y.; Li, T.; Yin, C.; Zhao, J.; Yu, J.; Yin, Q.; Gupta, V.K.; Jiang, Y.; et al. New insights into the evolution of host specificity of three Penicillium species and the pathogenicity of P. Italicum involving the infection of Valencia orange (Citrus sinensis). Virulence 2020, 11, 748–768. [Google Scholar] [CrossRef]
- Yin, C.; Zhu, H.; Jiang, Y.; Shan, Y.; Gong, L. Silencing dicer-like genes reduces virulence and sRNA generation in Penicillium italicum, the cause of citrus blue mold. Cells 2020, 9, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, B.D.; Hoch, H.C. Biology of the fungal cell. In The Mycota VIII; Howard, R.J., Gow, N.A.R., Eds.; Springer-Verlag KG: Berlin/Heidelberg, Germany, 2001; pp. 73–89. [Google Scholar]
- Tamuli, R.; Kumar, R.; Deka, R. Cellular roles of neuronal calcium sensor-1 and calcium/calmodulin-dependent kinases in fungi. J. Basic Microbiol. 2011, 51, 120–128. [Google Scholar] [CrossRef]
- Zhen, Z.; Zhang, G.; Yang, L.; Ma, N.; Li, Q.; Ma, Y.; Niu, X.; Zhang, K.; Yang, J. Characterization and functional analysis of calcium/calmodulin-dependent protein kinases (CaMKs) in the nematode-trapping fungus Arthrobotrys oligospora. Appl. Microbiol. Biot. 2019, 103, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Pausch, M.H.; Kaim, D.; Kunisawa, R.; Admon, A.; Thorner, J. Multiple Ca2+/calmodulin-dependent protein kinase genes in a unicellular eukaryote. EMBO J. 1991, 10, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Dayton, S.; Means, A.R. Ca2+/calmodulin-dependent kinase is essential for both growth and nuclear division in Aspergillus nidulans. Mol. Biol. Cell. 1996, 7, 1511–1519. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.D.; Means, A.R. Identification and characterization of two Ca2+/CaM-dependent protein kinases required for normal nuclear division in Aspergillus nidulans. J. Biol. Chem. 2000, 275, 38230–38238. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, C.D. Cloning of a calmodulin kinase I homologue from Schizosaccharomyces pombe. J. Biol. Chem. 2000, 275, 685–690. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, K.; Yamada, Y.; Sueyoshi, N.; Watanabe, A.; Asada, Y.; Kameshita, I. Novel Ca2+/calmodulin-dependent protein kinase expressed in actively growing mycelia of the basidiomycetous mushroom Coprinus cinereus. Biochim. Biophys. Acta 2009, 1790, 71–79. [Google Scholar] [CrossRef]
- Liu, X.; Lu, J.; Dong, B.; Gu, Y.; Lin, F. Disruption of MoCMK1, encoding a putative calcium/calmodulin-dependent kinase, in Magnaporthe oryzae. Microbiol. Res. 2010, 165, 402–410. [Google Scholar] [CrossRef]
- Jiao, M.; Yu, D.; Tan, C.; Guo, J.; Lan, D.; Han, E.; Qi, T.; Voegele, R.T.; Kang, Z.; Guo, J. Basidiomycete-specific PsCaMKL1 encoding a CaMK-like protein kinase is required for full virulence of Puccinia striiformis f. sp. tritici. Environ. Microbiol. 2017, 19, 4177–4189. [Google Scholar] [CrossRef]
- Ding, X.; Yu, Q.; Zhang, B.; Xu, N.; Jia, C.; Dong, Y.; Chen, Y.; Xing, L.; Li, M. The type II Ca2+/calmodulin-dependent protein kinases are involved in the regulation of cell wall integrity and oxidative stress response in Candida albicans. Biochem. Biophys. Res. Commun. 2014, 446, 1073–1078. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zhu, G.; Ding, Q.; Zhou, P.; Liu, L.; Chen, X. CgCmk1 activates CgRds2 to resist low-pH stress in Candida glabrata. Appl. Environ. Microbiol. 2020, 86, e00302-20. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Shan, A.; Hu, J.; Cao, Z.; Lv, R.; Feng, J. Genetic interaction between Ptc2 and protein phosphatase 4 (PP4) in the regulation of DNA damage response and virulence in Candida albicans. FEMS Yeast Res. 2019, 19, foz075. [Google Scholar] [CrossRef] [PubMed]
- Peroumal, D.; Manohar, K.; Patel, S.K.; Kumari, P.; Sahu, S.R.; Acharya, N. Virulence and pathogenicity of a Candida albicans mutant with reduced filamentation. Cell. Microbiol. 2019, 21, e13103. [Google Scholar] [CrossRef]
- Jenull, S.; Tscherner, M.; Gulati, M.; Nobile, C.J.; Chauhan, N.; Kuchler, K. The Candida albicans HIR histone chaperone regulates the yeast-tohyphae transition by controlling the sensitivity to morphogenesis signals. Sci. Rep. 2017, 7, 8308. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Cao, Q.; Li, N.; Liu, D.; Yuan, Y. Transcriptome analysis of fungicide responsive gene expression profiles in two Penicillium italicum strains with different response to the sterol demethylation inhibitor (DMI) fungicide prochloraz. BMC Genom. 2020, 21, 156. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Wang, S.; Yuan, Y.; Zhang, T.; Liu, J.; Liu, D. A novel major facilitator superfamily transporter in Penicillium digitatum (PdMFS2) is required for prochloraz resistance, conidiation and full virulence. Biotechnol. Lett. 2016, 38, 1349–1357. [Google Scholar] [CrossRef]
- Ballester, A.; Marcet-Houben, M.; Levin, E.; Sela, N.; Selma-Lázaro, C.; Carmona, L.; Wisniewski, M.; Droby, S.; González-Candelas, L.; Gabaldón, T. Genome, transcriptome, and functional analyses of Penicillium expansum provide new insights into secondary metabolism and pathogenicity. Mol. Plant-Microbe Interact. 2015, 28, 232–248. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with bowtie 2. Nat. Methods 2012, 9, 357. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ohya, Y.; Kawasaki, H.; Suzukis, K.; Londesborough, J.; Anraku, Y. Two yeast genes encoding calmodulin-dependent protein kinases. Isolation, sequencing and bacterial expressions of CMK1 and CMK2. J. Biol. Chem. 1991, 266, 12784–12794. [Google Scholar] [CrossRef]
- Melcher, L.; Thorner, J. Identification and characterization of the CLK1 gene product, a novel CaM kinase-like protein kinase from the yeast Saccharomyces cerevisiae. J. Biol. Chem. 1996, 271, 29958–29968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornstein, L.B.; Gaiso, M.L.; Hammell, R.L.; Bartelt, D.C. Cloning and sequence determination of a cDNA encoding Aspergillus nidulans calmodulin-dependent multifunctional protein kinase. Gene 1992, 113, 75–82. [Google Scholar] [CrossRef]
- Kim, Y.; Li, D.; Kolattukudy, P.E. Induction of Ca2+-Calmodulin signaling by hard-surface contact primes Colletotrichum gloeosporioides conidia to germinate and form appressoria. J. Bacteriol. 1998, 180, 5144–5150. [Google Scholar] [CrossRef] [Green Version]
- Valle-Aviles, L.; Valentin-Berrios, S.; Gonzalez-Mendez, R.R.; Rodriguez-del, V.N. Functional, genetic and bioinformatic characterization of a calcium/calmodulin kinase gene in Sporothrix schenckii. BMC Microbiol. 2007, 7, 107. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Cheng, P.; Zhi, G.; Liu, Y. Identification of a calcium/calmodulin-dependent protein kinasethat phosphorylates the Neurospora circadian clock protein FREQUENCY. J. Biol. Chem. 2001, 276, 41064–41072. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Tamuli, R. Calcium/calmodulin-dependent kinases are involved in growth, thermotolerance, oxidative stress survival, and fertility in Neurospora crassa. Arch. Microbiol. 2014, 196, 295–305. [Google Scholar] [CrossRef]
- Braun, A.P.; Schulman, H. The multifunctional calcium/calmodulin-dependent protein kinase: From form to function. Annu. Rev. Physiol. 1995, 57, 417–445. [Google Scholar] [CrossRef]
- Hook, S.S.; Means, A.R. Ca2+/CaM-dependent kinases: From activation to function. Annu. Rev. Pharmacol. 2001, 41, 471–505. [Google Scholar] [CrossRef]
- Dayton, J.S.; Sumi, M.; Nanthakumar, N.N.; Means, A.R. Expression of a constitutively active Ca2+/calmodulin-dependent kinase in Aspergillus nidulans spores prevents germination and entry into the cell cycle. J. Biol. Chem. 1997, 272, 3223–3230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crespo, A.; Gavaldà, J.; Julián, E.; Torrents, E. A single point mutation in class III ribonucleotide reductase promoter renders Pseudomonas aeruginosa PAO1 inefficient for anaerobic growth and infection. Sci. Rep. 2017, 7, 13350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenorio-Gómez, M.; de Sena-Tomás, C.; Pérez-Martín, J. MRN- and 9-1-1-independent activation of the ATR-Chk1 pathway during the Induction of the virulence program in the phytopathogen Ustilago maydis. PLoS ONE 2015, 10, e0137192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelliher, C.M.; Haase, S.B. Connecting virulence pathways to cell-cycle progression in the fungal pathogen Cryptococcus neoformans. Curr. Genet. 2017, 63, 803–811. [Google Scholar] [CrossRef]
- González-Rodríguez, V.E.; Liñeiro, E.; Colby, T.; Harzen, A.; Garrido, C.; Cantoral, J.M.; Schmidt, J.; Fernández-Acero, F.J. Proteomic profiling of Botrytis cinerea conidial germination. Arch. Microbiol. 2015, 197, 117–133. [Google Scholar] [CrossRef]
- Wang, Z.; Ma, T.; Huang, Y.; Wang, J.; Chen, Y.; Kistler, H.C.; Ma, Z.; Yin, Y. A fungal ABC transporter FgAtm1 regulates iron homeostasis via the transcription factor cascade FgAreA-HapX. PLoS Pathog. 2019, 15, e1007791. [Google Scholar] [CrossRef]
- Edlind, T.; Smith, L.; Henry, K.; Katiyar, S.; Nickels, J. Antifungal activity in Saccharomyces cerevisiae is modulated by calcium signalling. Mol. Microbiol. 2002, 46, 257–268. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; Lu, L. Calcium signaling pathway is involved in non-CYP51 azole resistance in Aspergillus fumigatus. Med. Mycol. 2019, 57, S233–S238. [Google Scholar] [CrossRef]
- Squizani, E.D.; Reuwsaat, J.C.V.; Motta, H.; Tavanti, A.; Kmetzsch, L. Calcium: A central player in Cryptococcus biology. Fungal Biol. Rev. 2021, 36, 27–41. [Google Scholar] [CrossRef]
- Märquez, J.A.; Serrano, R. Multiple transduction pathways regulate the sodium-extrusion gene PMR2/ENA1 during salt stress in yeast. FEBS Lett. 1996, 382, 89–92. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Fang, T.; Yan, H.; Jiang, L. The protein kinase Cmk2 negatively regulates the calcium/calcineurin signalling pathway and expression of calcium pump genes PMR1 and PMC1 in budding yeast. Cell Commun. Signal. 2019, 17, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, P.S.; Waters, O.D.; Jörgens, C.I.; Lowe, R.G.; Rechberger, J.; Trengove, R.D.; Oliver, R.P. Mannitol is required for asexual sporulation in the wheat pathogen Stagonospora nodorum (glume blotch). Biochem. J. 2006, 399, 231–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Long, N.N.; Vasseur, V.; Coroller, L.; Dantigny, P.; Le Panse, S.; Weill, A.; Mounier, J.; Rigalma, K. Temperature, water activity and pH during conidia production affect the physiological state and germination time of Penicillium species. Int. J. Food Microbiol. 2017, 241, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Calmes, B.; Guillemette, T.; Teyssier, L.; Siegler, B.; Pigné, S.; Landreau, A.; Iacomi, B.; Lemoine, R.; Richomme, P.; Simoneau, P. Role of mannitol metabolism in the pathogenicity of the necrotrophic fungus Alternaria brassicicola. Front. Plant Sci. 2013, 13, 131. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KEGG Pathway (ID) | Gene ID | Gene Function | Log2 * FC | FDR |
|---|---|---|---|---|
| DNA replication (ko03030) | EKV13547 | DNA primase (large subunit) | −7.57 | 1.06 × 10−5 |
| EKV11776 | DNA replication licensing factor Mcm6 | −2.13 | 3.01 × 10−7 | |
| EKV18089 | DNA polymerase δ (catalytic subunit) | −1.53 | 5.95 × 10−5 | |
| EKV08814 | DNA polymerase ε (catalytic subunit) | −1.49 | 2.40 × 10−4 | |
| EKV11198 | DNA replication licensing factor Mcm3 | −1.19 | 3.90 × 10−5 | |
| EKV17606 | Replication factor-a protein | −1.14 | 2.22 × 10−3 | |
| EKV16128 | DNA replication licensing factor Mcm2 | −1.08 | 2.62 × 10−5 | |
| Cell cycle (ko04111) | EKV11033 | Condensin complex subunit (HEAT-like repeat) | −4.11 | 1.19 × 10−3 |
| EKV11776 | DNA replication licensing factor Mcm6 | −2.13 | 3.01 × 10−7 | |
| EKV17483 | Cell-cycle checkpoint protein kinase (DNA damage response protein kinase) | −2.12 | 1.27 × 10−10 | |
| EKV19093 | Nuclear condensin complex Smc2 (structural maintenance of chromosome) | −1.51 | 7.96 × 10−8 | |
| EKV04214 | Replication checkpoint protein (MRC1-like domain) | −1.38 | 1.92 × 10−3 | |
| EKV16186 | Mitotic spindle checkpoint protein (Mad2) | −1.27 | 3.21 × 10−3 | |
| EKV11198 | DNA replication licensing factor Mcm3 | −1.19 | 3.90 × 10−5 | |
| EKV16749 | Condensin complex subunit 1 | −1.10 | 3.82 × 10−4 | |
| EKV16128 | DNA replication licensing factor Mcm2 | −1.08 | 2.62 × 10−5 | |
| Purine metabolism (ko00230) | EKV04683 | Xanthine dehydrogenase HxA | −7.57 | 1.06 × 10−5 |
| EKV13547 | DNA primase (large subunit) | −7.57 | 1.06 × 10−5 | |
| EKV18089 | DNA polymerase δ (catalytic subunit) | −1.53 | 5.95 × 10−5 | |
| EKV19574 | Ribonucleoside-diphosphate reductase | −1.53 | 2.12 × 10−5 | |
| EKV08814 | DNA polymerase ε (catalytic subunit) | −1.49 | 2.40 × 10−4 | |
| EKV15599 | Ribonucleotide reductase RnrA | −1.13 | 1.08 × 10−5 | |
| EKV16890 | DNA-directed RNA polymerase III | −1.12 | 1.81 × 10−4 | |
| EKV07940 | Adenylate cyclase | −1.01 | 2.19 × 10−4 | |
| Pyrimidine metabolism (ko00240) | EKV07761 | Uracil phosphoribosyltransferase | −10.48 | 1.79 × 10−32 |
| EKV13547 | DNA primase (large subunit) | −7.57 | 1.06 × 10−5 | |
| EKV18089 | DNA polymerase δ (catalytic subunit) | −1.53 | 5.95 × 10−5 | |
| EKV19574 | Ribonucleoside-diphosphate reductase | −1.53 | 2.12 × 10−5 | |
| EKV08814 | DNA polymerase ε (catalytic subunit) | −1.49 | 2.40 × 10−4 | |
| EKV15599 | Ribonucleotide reductase RnrA | −1.13 | 1.08 × 10−5 | |
| EKV16890 | DNA-directed RNA polymerase III | −1.12 | 1.81 × 10−4 | |
| Base excision repair (ko03410) | EKV18089 | DNA polymerase δ (catalytic subunit) | −1.53 | 5.95 × 10−5 |
| EKV08814 | DNA polymerase ε (catalytic subunit) | −1.49 | 2.40 × 10−4 | |
| EKV07371 | Formamidopyrimidine-DNA glycosylase | −1.33 | 5.95 × 10−5 | |
| Nucleotide excision repair (ko03420) | EKV18089 | DNA polymerase δ (catalytic subunit) | −1.53 | 5.95 × 10−5 |
| EKV08814 | DNA polymerase ε (catalytic subunit) | −1.49 | 2.40 × 10−4 | |
| EKV15299 | DNA repair protein RAD1 | −1.39 | 6.80 × 10−7 | |
| EKV17606 | Replication factor-a protein | −1.14 | 1.22 × 10−3 | |
| Mismatch repair (ko03430) | EKV18089 | DNA polymerase δ (catalytic subunit) | −1.53 | 5.95 × 10−5 |
| EKV17606 | Replication factor-a protein | −1.14 | 1.22 × 10−3 | |
| Meiosis (ko04113) | EKV11776 | DNA replication licensing factor Mcm6 | −2.13 | 3.01 × 10−7 |
| EKV16186 | Mitotic spindle checkpoint protein (Mad2) | −1.27 | 3.21 × 10−3 | |
| EKV11198 | DNA replication licensing factor Mcm3 | −1.19 | 3.90 × 10−5 | |
| EKV16128 | DNA replication licensing factor Mcm2 | −1.08 | 2.62 × 10−5 | |
| EKV07940 | Adenylate cyclase | −1.01 | 2.19 × 10−4 | |
| MAPK signaling pathway (ko04011) | EKV17484 | Phosphatidylinositol 4-kinase | −1.62 | 3.94 × 10−6 |
| Oxidative phosphorylation (ko00190) | EKV05405 | Cytochrome c oxidase assembly protein | −7.43 | 3.13 × 10−5 |
| EKV18906 | Mitochondrial F1/F0-ATP synthase | −4.60 | 3.19 × 10−21 | |
| Carotenoid biosynthesis (ko00906) | EKV07272 | Aldehyde dehydrogenase (β-apo-4′-carotenal oxygenase) | −7.28 | 9.59 × 10−5 |
| Glutathione metabolism (ko00480) | EKV19574 | Ribonucleoside-diphosphate reductase | −1.53 | 2.12 × 10−5 |
| EKV15599 | Ribonucleotide reductase RnrA | −1.13 | 1.08 × 10−5 | |
| Cysteine and methionine metabolism (ko00270) | EKV06483 | Cysteine synthase A | −9.77 | 2.13 × 10−22 |
| Sulfur metabolism (ko00920) | EKV06483 | Cysteine synthase A | −9.77 | 2.13 × 10−22 |
| EKV18475 | Assimilatory sulfite reductase | −1.73 | 5.56 × 10−3 | |
| Starch and sucrose metabolism (ko00500) | EKV04855 | Oligo-1,6-glucosidase (α-amylase or maltase) | −3.09 | 3.67 × 10−6 |
| Amino sugar and nucleotide sugar metabolism (ko00520) | EKV05685 | Glucosamine-6-phosphate deaminase | −12.86 | 1.32 × 10−83 |
| EKV11299 | NADH-cytochrome b5 reductase | −1.59 | 1.52 × 10−8 | |
| EKV15950 | Chitin synthase A/B | −1.19 | 5.23 × 10−4 | |
| Biosynthesis of amino acids (ko01230) | EKV06483 | Cysteine synthase A | −9.77 | 2.13 × 10−22 |
| EKV17406 | Catabolic 3-dehydroquinase | −1.26 | 9.29 × 10−5 | |
| Protein processing in endoplasmic reticulum (ko04141) | EKV13467 | DnaJ-related protein SCJ1 | −8.53 | 1.19 × 10−10 |
| EKV14522 | Polyubiquitin binding protein (Doa1/Ufd3) | −1.68 | 1.34 × 10−9 | |
| EKV14057 | Heat shock protein 90 (HSP90) | −1.33 | 8.03 × 10−7 | |
| EKV13033 | Heat shock 70 kDa protein (HSP70) | −1.09 | 2.44 × 10−6 | |
| EKV13686 | Nuclear protein localization protein (NPL4 family) | −1.04 | 6.08 × 10−4 | |
| Endocytosis (ko04144) | EKV18650 | Phospholipase D | −9.35 | 1.16 × 10−17 |
| PHI26_NewGene_31 | Vacuolar protein sorting-associated protein (VHS domain) | −1.57 | 4.21 × 10−8 | |
| PHI26_NewGene_32 | Vacuolar protein sorting-associated protein (FYVE-like protein) | −1.10 | 5.12 × 10−3 | |
| EKV13033 | Heat shock 70 kDa protein (HSP70) | −1.09 | 2.44 × 10−6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, G.; Liu, S.; Wu, L.; Wang, X.; Cuan, R.; Zheng, Y.; Liu, D.; Yuan, Y. Characterization and Functional Analysis of a New Calcium/Calmodulin-Dependent Protein Kinase (CaMK1) in the Citrus Pathogenic Fungus Penicillium italicum. J. Fungi 2022, 8, 667. https://doi.org/10.3390/jof8070667
Li G, Liu S, Wu L, Wang X, Cuan R, Zheng Y, Liu D, Yuan Y. Characterization and Functional Analysis of a New Calcium/Calmodulin-Dependent Protein Kinase (CaMK1) in the Citrus Pathogenic Fungus Penicillium italicum. Journal of Fungi. 2022; 8(7):667. https://doi.org/10.3390/jof8070667
Chicago/Turabian StyleLi, Guoqi, Shaoting Liu, Lijuan Wu, Xiao Wang, Rongrong Cuan, Yongliang Zheng, Deli Liu, and Yongze Yuan. 2022. "Characterization and Functional Analysis of a New Calcium/Calmodulin-Dependent Protein Kinase (CaMK1) in the Citrus Pathogenic Fungus Penicillium italicum" Journal of Fungi 8, no. 7: 667. https://doi.org/10.3390/jof8070667