
Phylogeny and Diversity of the Genus Pseudohydnum (Auriculariales, Basidiomycota)

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Studies
2.2. DNA Extraction, Amplification, and Sequencing
2.3. Molecular Phylogenetics Analysis
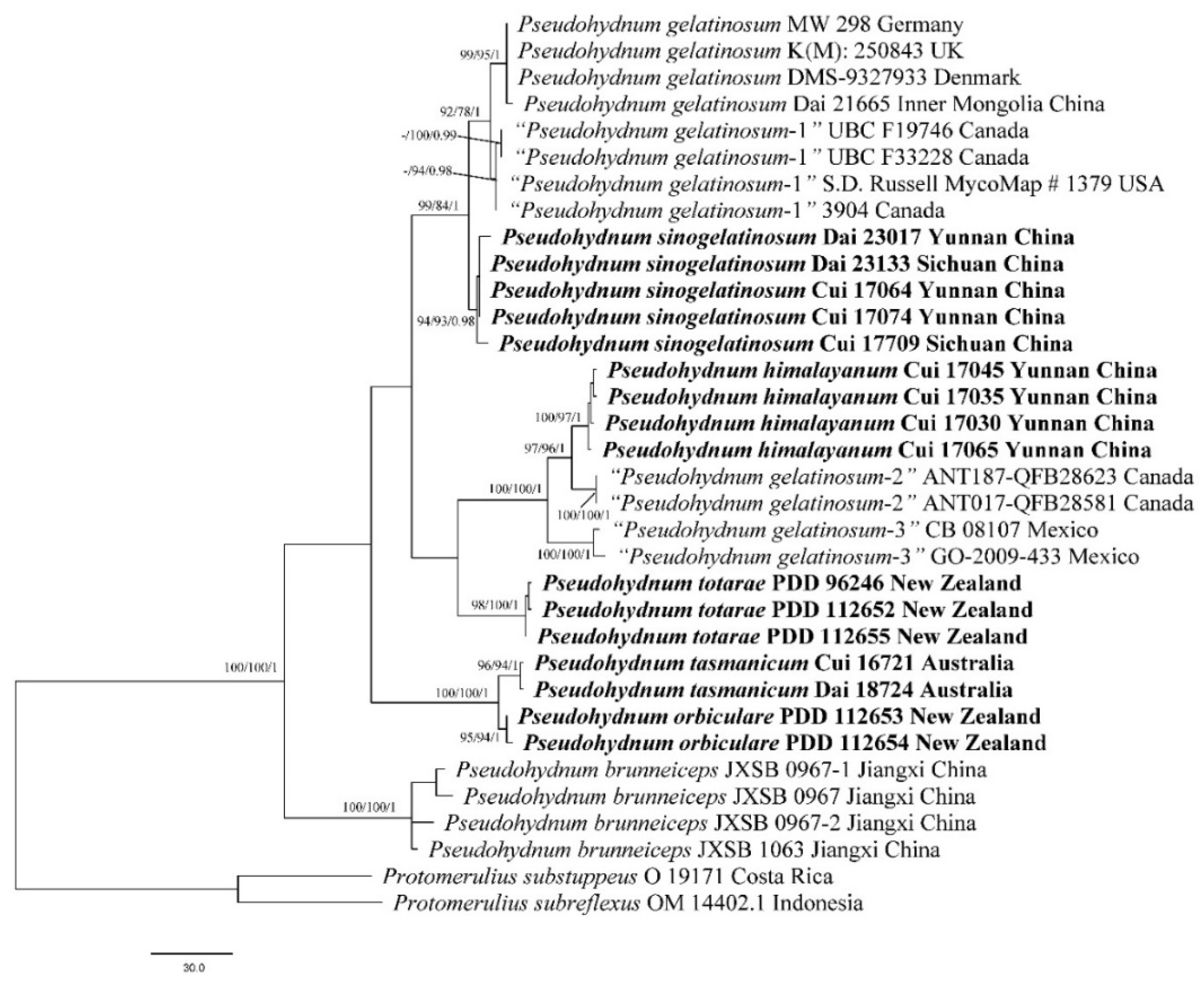
3. Results
3.1. Molecular Phylogeny
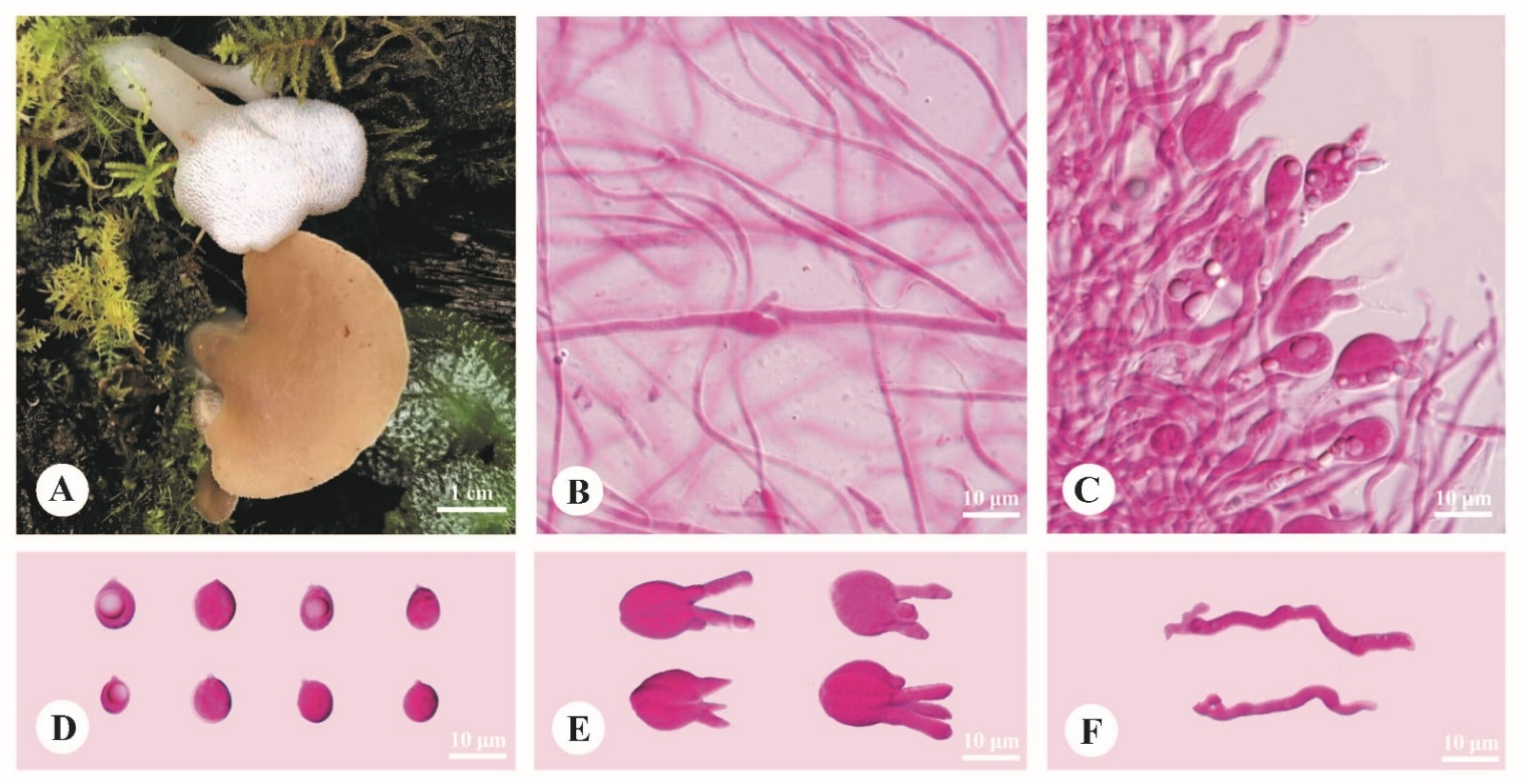
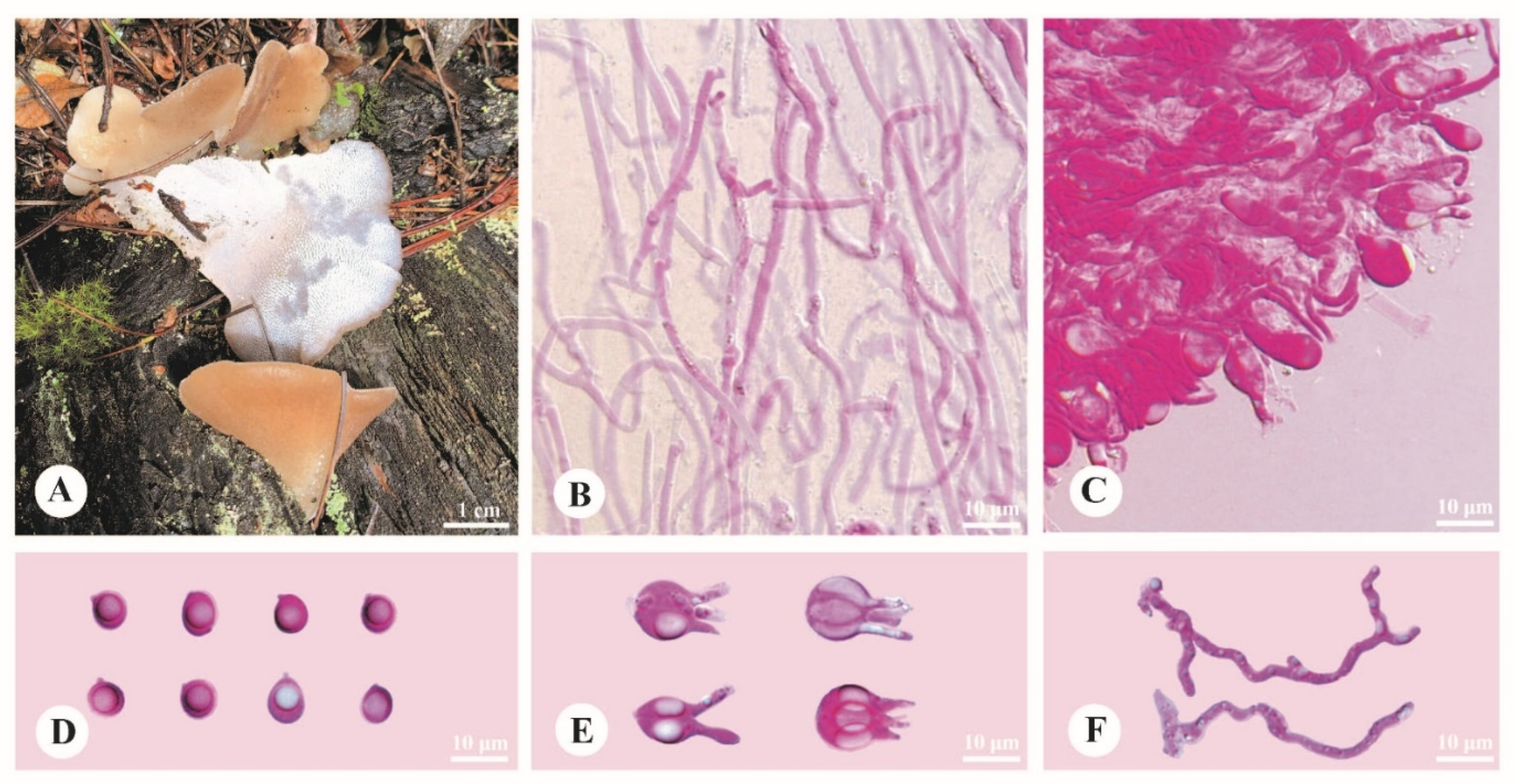
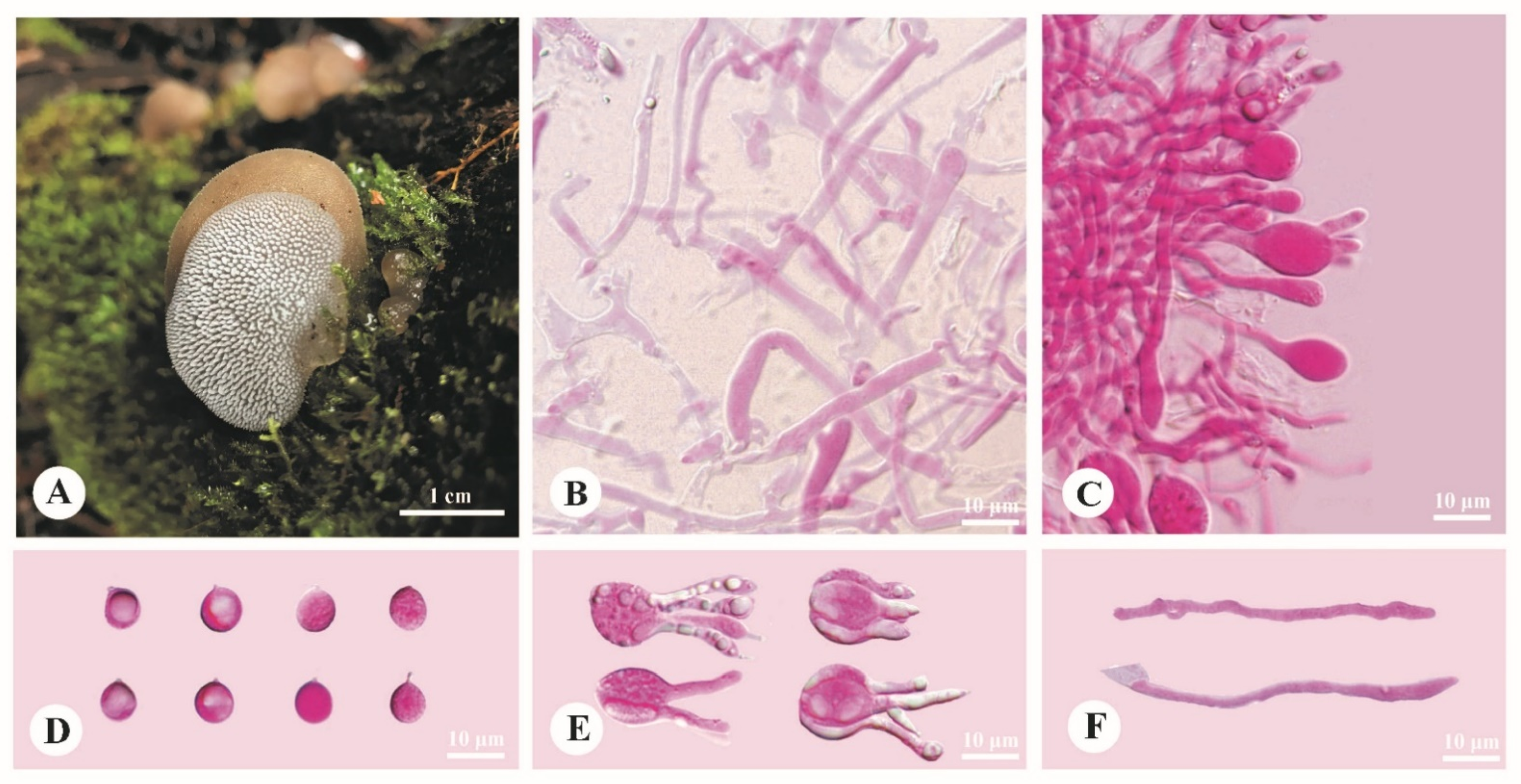
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Karsten, P.A. Auriculariei, Clavariei et Tremellini, in paroecia Tammela crescentes. Not. Sällskapets Fauna Flora Fenn. Förhandlingar 1868, 9, 365–374. [Google Scholar]
- Niveiro, N.; Popoff, O.F. Pseudohydnum gelatinosum (Tremellales, Basidiomycota) in the Argentinean Yungas. Bol. Soc. Argent. Bot. 2011, 46, 223–226. [Google Scholar]
- Breitenbach, J.; Kränzlin, F. Fungi of Switzerland 2. Non-Gilled Fungi, Heterobasidiomycetes, Aphyllophorales, Gasteromycetes; Verlag Mykologia Lucerne Switzerland: Luzern, Switzerland, 1986. [Google Scholar]
- Bandoni, R.J. The Tremellales and Auriculariales: An alternative classification. Trans. Mycol. Soc. Jpn. 1984, 25, 489–530. [Google Scholar]
- Weiß, M.; Oberwinkler, F. Phylogenetic relationships in Auriculariales and related groups—Hypotheses derived from nuclear ribosomal DNA sequences. Mycol. Res. 2001, 105, 103–415. [Google Scholar] [CrossRef]
- Malysheva, V.; Spirin, V. Taxonomy and phylogeny of the Auriculariales (Agaricomycetes, Basidiomycota) with stereoid basidiocarps. Fungal Biol. 2017, 121, 689–715. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, C.G. Mycological notes 75. Mycol. Writ. 1925, 7, 1349–1364. [Google Scholar]
- Chen, Y.L.; Su, M.S.; Zhang, L.P.; Zou, Q.; Wu, F.; Zeng, N.K.; Liu, M. Pseudohydnum brunneiceps (Auriculariales, Basidiomycota), a new species from Central China. Phytotaxa 2020, 441, 87–94. [Google Scholar] [CrossRef]
- Bresadola, G. Iconographia Mycologica; Gruppo Micologico “G. Bresadola”: Trento, Italy, 1932; Volume 23, pp. 1100–1150. [Google Scholar]
- Kobayasi, Y. Monographic studies of Japanese Tremellaceous fungi 6. Nagaoa 1954, 4, 36–47. [Google Scholar]
- Courtecuisse, R.; Lowy, B. Elements for a mycological inventory of the vicinity of ‘Saut Pararé’ (Arataye River) and ‘Nouragues Inselberg’ (French Guiana) 3. Heterobasidiomycetideae. Studies on the flora of the Guianas 52. Mycotaxon 1990, 39, 329–344. [Google Scholar]
- Lowy, B. New or noteworthy Tremellales from Bolivia. Mycologia 1959, 51, 840–850. [Google Scholar] [CrossRef]
- Lowy, B. Tremellales. Flora Neotrop. 1971, 6, 1–153. [Google Scholar]
- Rick, J.E. Fungi austro-americani exs. Fasc 1. Ann. Mycol. 1904, 2, 406–410. [Google Scholar]
- Léveillé, J.H. Champignons exotiques. Ann. Sci. Nat. Bot. 1844, 2, 167–221. [Google Scholar]
- Ryvarden, L. A note on the genus Hydnodon Banker. Synop. Fungorum 2002, 15, 31–33. [Google Scholar]
- Scopoli, J.A. Flora Carniolica; Impensis Ioannis Pauli Krauss: Vienna, Austria, 1772. [Google Scholar]
- Pemberton, R.T. Agglutinins (lectins) from some British higher fungi. Mycol. Res. 1994, 98, 277–290. [Google Scholar] [CrossRef]
- Salo, P.; Niemela, T.; Salo, U. Suomen Sieniopas; WSOY: Helsinik, Finland, 2005. [Google Scholar]
- Volk, T.J.; Burdsall, H.H.; Reynolds, K. Checklist and host index of wood-inhabiting fungi of Alaska. Mycotaxon 1994, 52, 1–46. [Google Scholar]
- Boulet, B. Les Champignons des Arbres de L’est de I’Amérique de Nord; Les Publications du Québec: Québec, QC, Canada, 2003. [Google Scholar]
- Binion, D.E.; Stephenson, S.L.; Roody, W.C.; Burdsall, H.H.; Vasilyeva, L.N.; Miller, O.K. Macrofungi Associated with Oaks of Eastern North America; West Virginia University Press: Morgantown, WV, USA, 2008. [Google Scholar]
- Kisimova-Horovitz, L.; Oberwinkler, F.; Gómez, L.D. Basidiomicetos resupinados de Costa Rica. Myxariaceaes. Jülich, Sebacinaceae Wells & Oberwy, Tremellodendropsidaceae Jülich. Rev. Biol. Trop. 2000, 48, 519–538. [Google Scholar]
- Hongo, T.; Izawa, M. Yama-Kei Field Books No.7; Yama-Kei Publishers Co., Ltd.: Tokyo, Japan, 2006. [Google Scholar]
- Dai, Y.C. A revised checklist of corticioid and hydnoid fungi in China for 2010. Mycoscience 2011, 52, 69–79. [Google Scholar] [CrossRef]
- Li, Y.; Li, T.H.; Yang, Z.L.; Tolgor, B.; Dai, Y.C. Atlas of Chinese Macrofungal Resources; Center China Farmer Press: Zhengzhou, China, 2015. [Google Scholar]
- McNabb, R.F.R. New Zealand Tremellales 1. N. Z. J. Bot. 1964, 2, 403–414. [Google Scholar] [CrossRef]
- Fuhrer, B.; Robinson, R. Rainforest Fungi of Tasmania and South-East Australia; CSIRO: Tasmania, Australia, 1992. [Google Scholar]
- Hood, I.A. An Illustrated Guide to Fungi on Wood in New Zealand; Auckland University Press: Auckland, New Zealand, 1992. [Google Scholar]
- Ridley, G.S.; Horne, D. A Photographic Guide to Mushrooms and Other Fungi of New Zealand; New Holland: Auckland, New Zealand, 2006. [Google Scholar]
- Gates, G.; Ratkowsky, D. A Field Guide to Tasmanian Fungi; Tasmanian Field Naturalists Club: Hobart, Australia, 2016. [Google Scholar]
- Wu, F.; Zhou, L.W.; Yang, Z.L.; Bau, T.; Li, T.H.; Dai, Y.C. Resource Diversity of Chinese macrofungi: Edible, medicinal and poisonous species. Fungal Divers. 2019, 98, 1–76. [Google Scholar] [CrossRef]
- Sternisa, M.; Sabotic, J.; Klancnik, A. A novel approach using growth curve analysis to distinguish between antimicrobial and anti-biofilm activities against Salmonella. Int. J. Food Microbiol. 2022, 364, 109520. [Google Scholar] [CrossRef]
- The Royal Botanic Garden Edinburgh. Flora of British Fungi; Colour Identification Chart; Her Majesty’s Stationery Office: London, UK, 1969. [Google Scholar]
- Petersen, J.H. Farvekort. The Danish Mycological Society’s Colour-Chart; Foreningen til Svampekundskabens Fremme: Greve, Italy, 1996; pp. 1–6. [Google Scholar]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed; Eyre Methuen: London, UK, 1978. [Google Scholar]
- Pippola, E.; Kotiranta, H. The genus Tremella (Basidiomycota, Tremellales) in Finland. Ann. Bot. Fenn. 2008, 45, 401–434. [Google Scholar] [CrossRef]
- Malysheva, V.; Malysheva, E.F.; Bulakh, E.M. The genus Tremella (Tremellales, Basidiomycota) in Russia with description of two new species and proposal of one nomenclatural combination. Phytotaxa 2015, 238, 40–70. [Google Scholar] [CrossRef]
- Fan, L.F.; Alvarenga, R.L.M.; Gibertoni, T.B.; Wu, F.; Dai, Y.C. Four new species in the Tremella fibulifera complex (Tremellales, Basidiomycota). MycoKeys 2021, 82, 33–56. [Google Scholar] [CrossRef]
- Sun, Y.-F.; Costa-Rezende, D.; Xing, J.-H.; Zhou, J.-L.; Zhang, B.; Gibertoni, T.; Gates, G.; Glen, M.; Dai, Y.-C.; Cui, B.-K. Multi-gene phylogeny and taxonomy of Amauroderma s.lat. (Ganodermataceae). Persoonia 2020, 44, 206–239. [Google Scholar] [CrossRef]
- Chen, J.J.; Dai, Y.C. Two new species of Physisporinus (Polyporales, Basidiomycota) from Yunnan, Southwest China. Mycol. Prog. 2021, 20, 1–10. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, M.J.T., Eds.; Academic Press: San Diego, FL, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. Bioedit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; Volume 41, pp. 95–98. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), version 4.0 Beta; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Miller, M.A.; Holder, M.T.; Vos, R.; Midford, P.E.; Liebowitz, T.; Chan, L.; Hoover, P.; Warnow, T. The CIPRES Portals. CIPRES. 2009. Available online: http://www.phylo.org/sub_sections/portal (accessed on 4 April 2022).
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayers, E.W.; Cavanaugh, M.; Clark, K.; Pruitt, K.D.; Schoch, C.L.; Sherry, S.T.; Karsch-Mizrachi, I. GenBank. Nucleic Acids Res. 2021, 49, D92–D96. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Location | Sample | ITS | nLSU |
|---|---|---|---|---|
| Pseudohydnum brunneiceps | Jiangxi, China | JXSB 0967 | MN497254 | MN497259 |
| Pseudohydnum brunneiceps | Jiangxi, China | JXSB 0967-1 | MN497255 | MN497260 |
| Pseudohydnum brunneiceps | Jiangxi, China | JXSB 0967-2 | MN497256 | MN497261 |
| Pseudohydnum brunneiceps | Jiangxi, China | JXSB 1063 | MN497257 | MN497258 |
| Pseudohydnum gelatinosum | Inner Mongolia, China | Dai 21665 | ON243826 | ON243924 |
| Pseudohydnum gelatinosum | Denmark | DMS-9327933 | MT644890 | MT644890 |
| Pseudohydnum gelatinosum | Germany | MW 298 | DQ520094 | DQ520094 |
| Pseudohydnum gelatinosum | UK | K(M): 250843 | MZ159722 | - |
| “Pseudohydnum gelatinosum-1” | Canada | 3904 | KM406980 | - |
| “Pseudohydnum gelatinosum-1” | Canada | UBC F19746 | HQ604801 | HQ604801 |
| “Pseudohydnum gelatinosum-1” | Canada | UBC F33228 | MG953967 | - |
| “Pseudohydnum gelatinosum-1” | USA | S.D. Russell MycoMap # 1379 | MK575262 | - |
| “Pseudohydnum gelatinosum-2” | Canada | ANT017-QFB28581 | MN992496 | - |
| “Pseudohydnum gelatinosum-2” | Canada | ANT187-QFB28623 | MN992495 | - |
| “Pseudohydnum gelatinosum-3” | Mexico | CB 08107 | KT875091 | - |
| “Pseudohydnum gelatinosum-3” | Mexico | GO-2009-433 | KC152166 | - |
| Pseudohydnum himalayanum | Yunnan, China | Cui 17030 | ON243827 | ON243925 |
| Pseudohydnum himalayanum | Yunnan, China | Cui 17035 | ON243828 | ON243926 |
| Pseudohydnum himalayanum | Yunnan, China | Cui 17045 | ON243829 | ON243927 |
| Pseudohydnum himalayanum | Yunnan, China | Cui 17065 | ON243830 | ON243928 |
| Pseudohydnum orbiculare | New Zealand | PDD 112653 | ON243831 | ON243929 |
| Pseudohydnum orbiculare | New Zealand | PDD 112654 | ON243832 | - |
| Pseudohydnum sinogelatinosum | Yunnan, China | Cui 17064 | ON243833 | - |
| Pseudohydnum sinogelatinosum | Yunnan, China | Cui 17074 | ON243834 | ON243930 |
| Pseudohydnum sinogelatinosum | Sichuan, China | Cui 17709 | ON243835 | ON243931 |
| Pseudohydnum sinogelatinosum | Yunnan, China | Dai 23017 | ON243836 | ON243932 |
| Pseudohydnum sinogelatinosum | Sichuan, China | Dai 23133 | ON243837 | ON243933 |
| Pseudohydnum tasmanicum | Australia | Cui 16721 | ON243838 | ON243934 |
| Pseudohydnum tasmanicum | Australia | Dai 18724 | ON243839 | ON243935 |
| Pseudohydnum totarae | New Zealand | PDD 96246 | ON243840 | - |
| Pseudohydnum totarae | New Zealand | PDD 112652 | ON243841 | - |
| Pseudohydnum totarae | New Zealand | PDD 112655 | ON243842 | ON243936 |
| Protomerulius subreflexus | Indonesia | OM 14402.1 | MG757508 | MG757508 |
| Protomerulius substuppeus | Costa Rica | O 19171 | JX134482 | JQ764649 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.-M.; Liu, H.-G.; Gates, G.M.; Wu, F.; Dai, Y.-C.; Cooper, J.A. Phylogeny and Diversity of the Genus Pseudohydnum (Auriculariales, Basidiomycota). J. Fungi 2022, 8, 658. https://doi.org/10.3390/jof8070658
Zhou H-M, Liu H-G, Gates GM, Wu F, Dai Y-C, Cooper JA. Phylogeny and Diversity of the Genus Pseudohydnum (Auriculariales, Basidiomycota). Journal of Fungi. 2022; 8(7):658. https://doi.org/10.3390/jof8070658
Chicago/Turabian StyleZhou, Hong-Min, Hong-Gao Liu, Genevieve M. Gates, Fang Wu, Yu-Cheng Dai, and Jerry A. Cooper. 2022. "Phylogeny and Diversity of the Genus Pseudohydnum (Auriculariales, Basidiomycota)" Journal of Fungi 8, no. 7: 658. https://doi.org/10.3390/jof8070658