
Mucosal-Associated Invariant T Cells Accumulate in the Lungs during Murine Pneumocystis Infection but Are Not Required for Clearance


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. P. murina Infection Model
2.3. Quantitation of P. murina by qPCR
2.4. ELISA
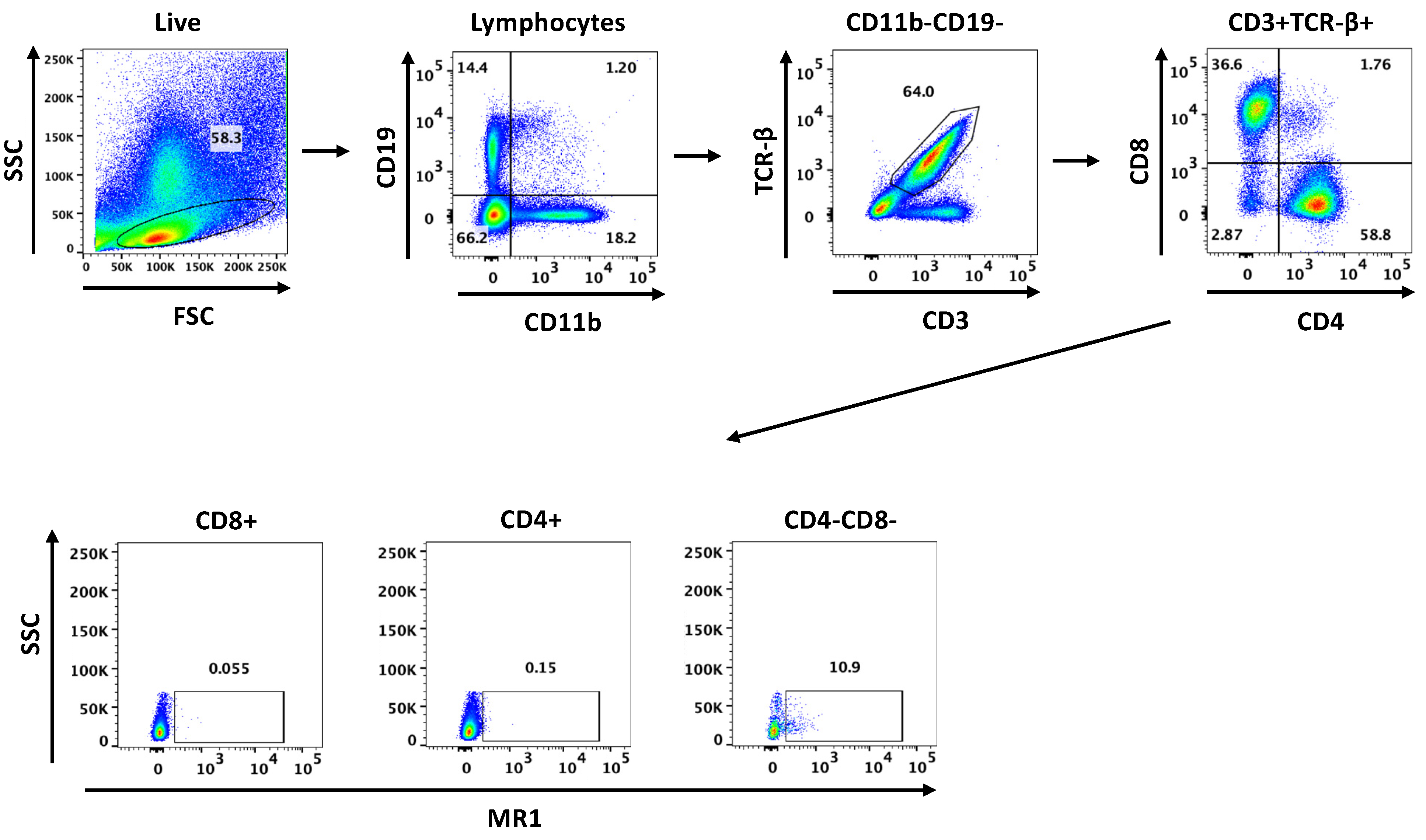
2.5. Flow Cytometry
2.6. Statistics
3. Results
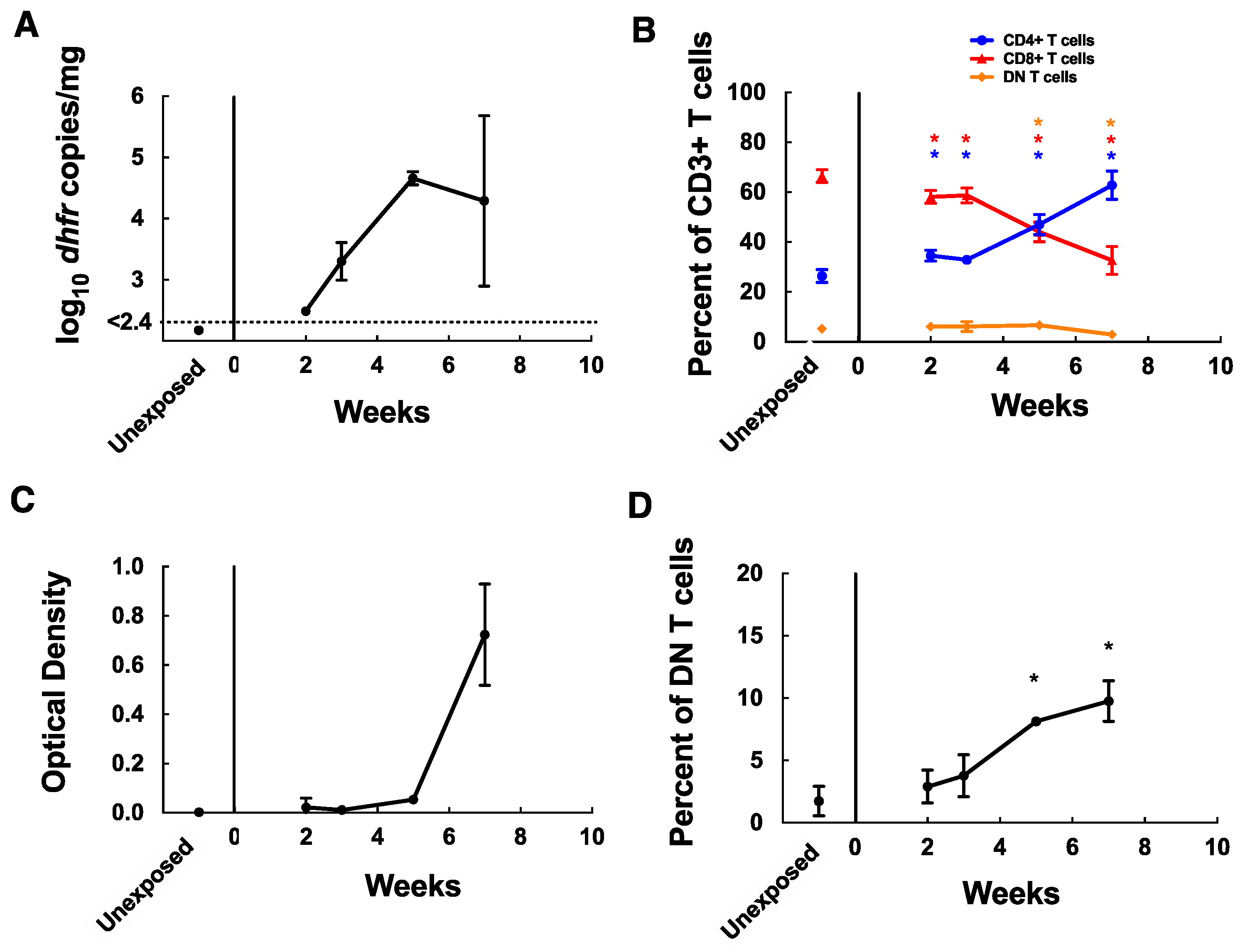
3.1. MAIT Cells Accumulate in the Lungs of C57BL/6 Mice during P. murina Infection
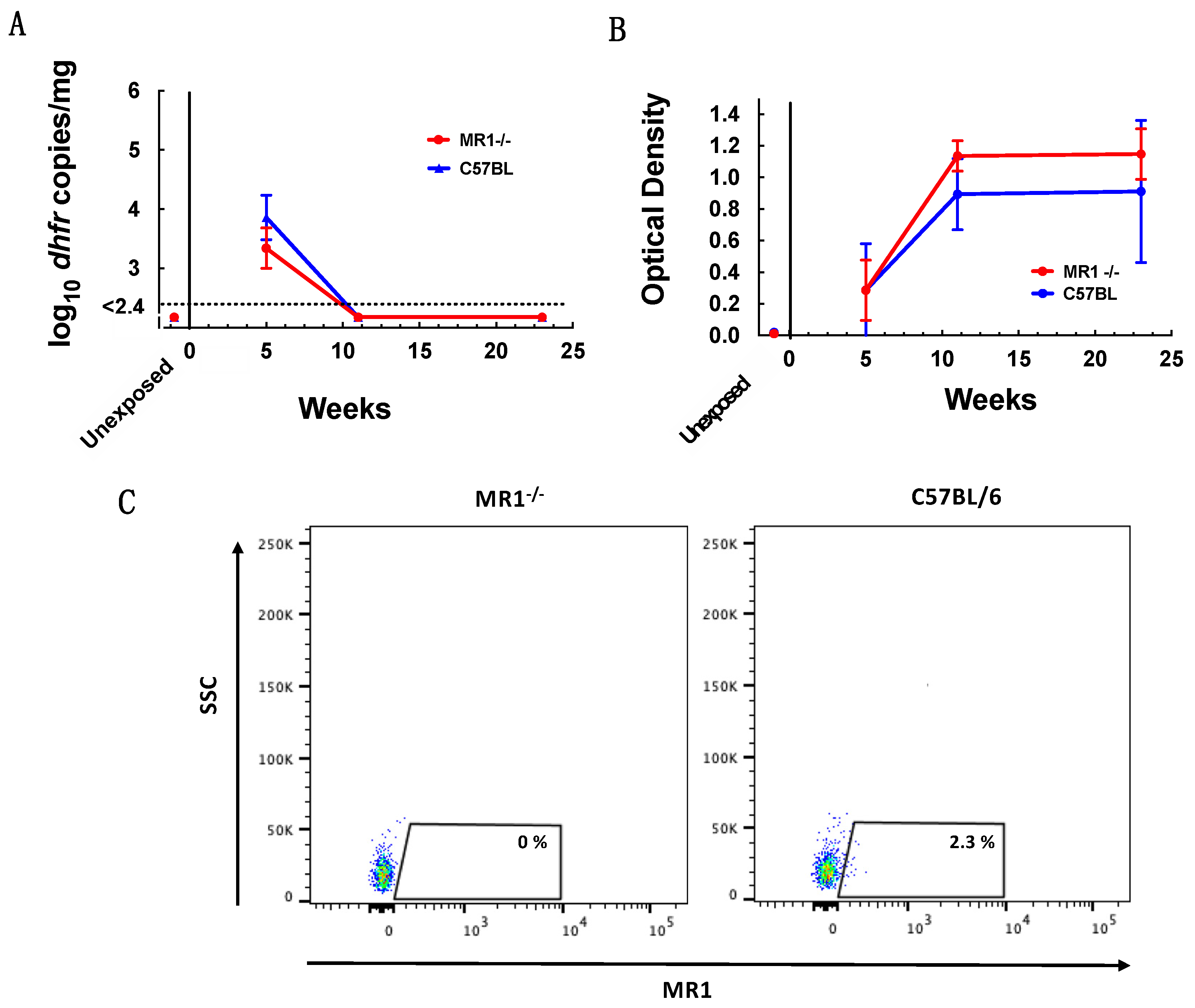
3.2. MAIT Cells Are Not Required for Control of P. murina Infection
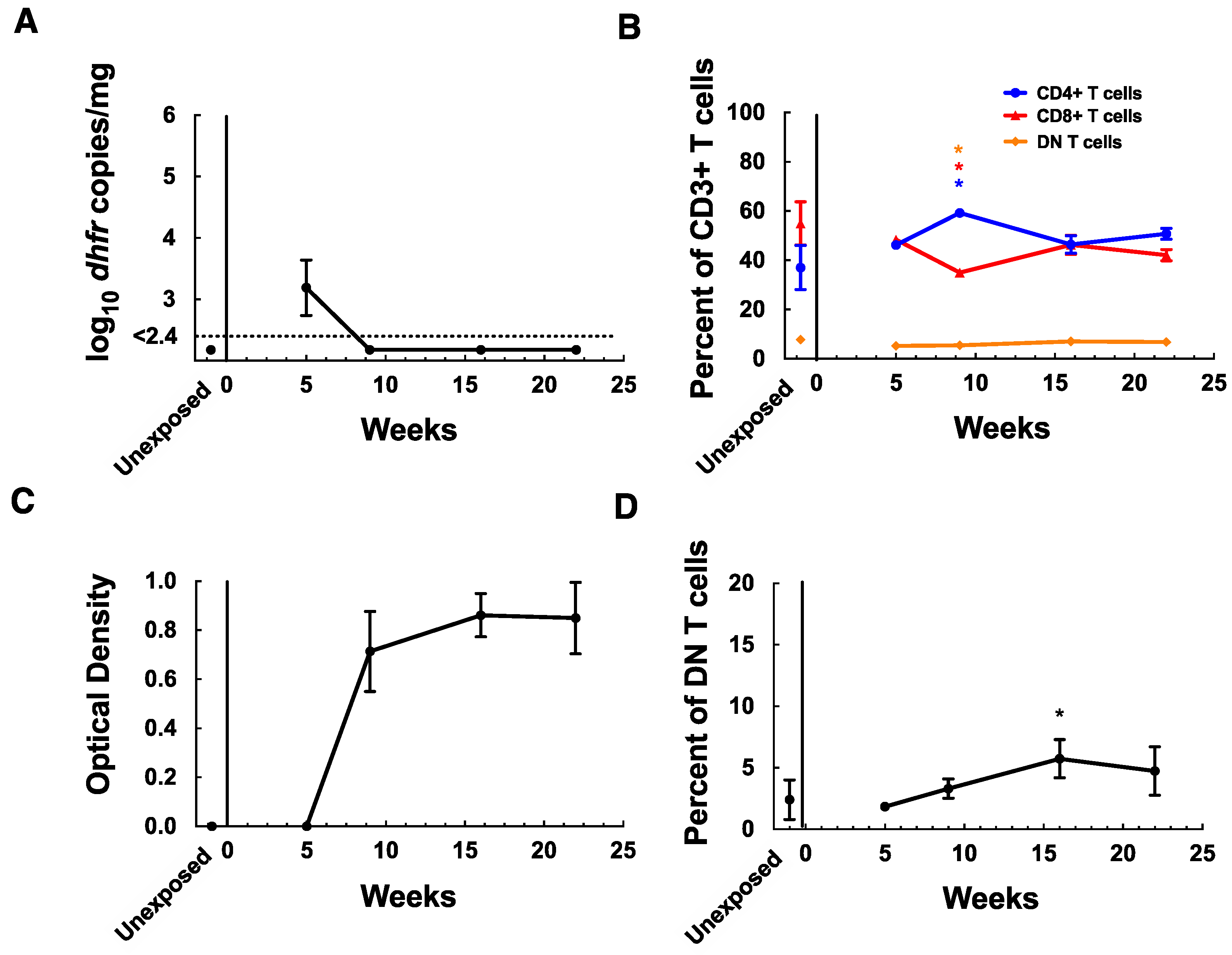
3.3. MAIT Cells Remain Elevated in Lungs after Clearance of P. murina Infection
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kovacs, J.A.; Masur, H. Evolving health effects of Pneumocystis: One hundred years of progress in diagnosis and treatment. JAMA 2009, 301, 2578–2585. [Google Scholar] [CrossRef] [PubMed]
- Carmona, E.M.; Limper, A.H. Update on the diagnosis and treatment of Pneumocystis pneumonia. Ther. Adv. Respir. Dis. 2011, 5, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, D.I.; Koay, H.F.; McCluskey, J.; Gherardin, N.A. The biology and functional importance of MAIT cells. Nat. Immunol. 2019, 20, 1110–1128. [Google Scholar] [CrossRef] [PubMed]
- Constantinides, M.G.; Link, V.M.; Tamoutounour, S.; Wong, A.C.; Perez-Chaparro, P.J.; Han, S.-J.; Chen, Y.E.; Li, K.; Farhat, S.; Weckel, A.; et al. MAIT cells are imprinted by the microbiota in early life and promote tissue repair. Science 2019, 366, eaax6624. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.N.; Eckle, S.B.; Xu, W.; Liu, L.; Hughes, V.A.; Mak, J.Y.; Meehan, B.S.; Pediongco, T.; Birkinshaw, R.W.; Chen, Z.; et al. Drugs and drug-like molecules can modulate the function of mucosal-associated invariant T cells. Nat. Immunol. 2017, 18, 402–411. [Google Scholar] [CrossRef]
- Sugimoto, C.; Fujita, H.; Wakao, H. Mucosal-associated invariant T cells from induced pluripotent stem cells: A novel approach for modeling human diseases. World J. Stem Cells 2016, 8, 158–169. [Google Scholar] [CrossRef]
- Cosgrove, C.; Ussher, J.E.; Rauch, A.; Gartner, K.; Kurioka, A.; Huhn, M.H.; Adelmann, K.; Kang, Y.-H.; Fergusson, J.R.; Simmonds, P.; et al. Early and nonreversible decrease of CD161++/MAIT cells in HIV infection. Blood 2013, 121, 951–961. [Google Scholar] [CrossRef]
- Leeansyah, E.; Ganesh, A.; Quigley, M.F.; Sonnerborg, A.; Andersson, J.; Hunt, P.W.; Somsouk, M.; Deeks, S.G.; Martin, J.N.; Moll, M.; et al. Activation, exhaustion, and persistent decline of the antimicrobial MR1-restricted MAIT-cell population in chronic HIV-1 infection. Blood 2013, 121, 1124–1135. [Google Scholar] [CrossRef]
- Magalhaes, I.; Pingris, K.; Poitou, C.; Bessoles, S.; Venteclef, N.; Kiaf, B.; Beaudoin, L.; Da Silva, J.; Allatif, O.; Rossjohn, J.; et al. Mucosal-associated invariant T cell alterations in obese and type 2 diabetic patients. J. Clin. Investig. 2015, 125, 1752–1762. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Wang, H.; D’Souza, C.; Sun, S.; Kostenko, L.; Eckle, S.B.; Meehan, B.S.; Jackson, D.C.; Strugnell, R.A.; Cao, H.; et al. Mucosal-associated invariant T-cell activation and accumulation after in vivo infection depends on microbial riboflavin synthesis and co-stimulatory signals. Mucosal Immunol. 2017, 10, 58–68. [Google Scholar] [CrossRef] [Green Version]
- Leeansyah, E.; Svard, J.; Dias, J.; Buggert, M.; Nystrom, J.; Quigley, M.F.; Moll, M.; Sonnerborg, A.; Nowak, P.; Sandberg, J.K. Arming of MAIT Cell Cytolytic Antimicrobial Activity Is Induced by IL-7 and Defective in HIV-1 Infection. PLoS Pathog. 2015, 11, e1005072. [Google Scholar] [CrossRef] [PubMed]
- Leng, T.; Akther, H.D.; Hackstein, C.P.; Powell, K.; King, T.; Friedrich, M.; Christoforidou, Z.; McCuaig, S.; Neyazi, M.; Arancibia-Carcamo, C.V.; et al. TCR and Inflammatory Signals Tune Human MAIT Cells to Exert Specific Tissue Repair and Effector Functions. Cell Rep. 2019, 28, 3077–3091.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinks, T.S.C. Boosting MAIT cells as immunotherapy: Context is everything. Mucosal Immunol. 2021, 14, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Kjer-Nielsen, L.; Shi, M.; D’Souza, C.; Pediongco, T.J.; Cao, H.; Kostenko, L.; Lim, X.Y.; Eckle, S.B.G.; Meehan, B.S.; et al. IL-23 costimulates antigen-specific MAIT cell activation and enables vaccination against bacterial infection. Sci. Immunol. 2019, 4, eaaw0402. [Google Scholar] [CrossRef]
- Bohineust, A.; Tourret, M.; Derivry, L.; Caillat-Zucman, S. Mucosal-associated invariant T (MAIT) cells, a new source of universal immune cells for chimeric antigen receptor (CAR)-cell therapy. Bull. Cancer 2021, 108, S92–S95. [Google Scholar] [CrossRef]
- Masina, N.; Bekiswa, A.; Shey, M. Mucosal-associated invariant T cells in natural immunity and vaccination against infectious diseases in humans. Curr. Opin. Immunol. 2021, 71, 1–5. [Google Scholar] [CrossRef]
- Ma, L.; Chen, Z.; Huang, D.W.; Kutty, G.; Ishihara, M.; Wang, H.; Abouelleil, A.; Bishop, L.; Davey, E.; Deng, R.; et al. Genome analysis of three Pneumocystis species reveals adaptation mechanisms to life exclusively in mammalian hosts. Nat. Commun. 2016, 7, 10740. [Google Scholar] [CrossRef]
- Cisse, O.H.; Ma, L.; Dekker, J.P.; Khil, P.P.; Youn, J.-H.; Brenchley, J.M.; Blair, R.; Pahar, B.; Chabe, M.; Van Rompay, K.K.A.; et al. Genomic insights into the host specific adaptation of the Pneumocystis genus. Commun. Biol. 2021, 4, 305. [Google Scholar] [CrossRef]
- Treiner, E.; Duban, L.; Bahram, S.; Radosavljevic, M.; Wanner, V.; Tilloy, F.; Affaticati, P.; Gilfillan, S.; Lantz, O. Selection of evolutionarily conserved mucosal-associated invariant T cells by MR1. Nature 2003, 422, 164–169. [Google Scholar] [CrossRef]
- Vestereng, V.H.; Bishop, L.R.; Hernandez, B.; Kutty, G.; Larsen, H.H.; Kovacs, J.A. Quantitative real-time polymerase chain-reaction assay allows characterization of Pneumocystis infection in immunocompetent mice. J. Infect. Dis. 2004, 189, 1540–1544. [Google Scholar] [CrossRef] [Green Version]
- Bishop, L.R.; Lionakis, M.S.; Sassi, M.; Murphy, P.M.; Hu, X.; Huang, D.W.; Sherman, B.; Qiu, J.; Yang, J.; Lempicki, R.A.; et al. Characterization of chemokine and chemokine receptor expression during Pneumocystis infection in healthy and immunodeficient mice. Microbes Infect. 2015, 17, 638–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Davis, A.S.; Ma, L.; Bishop, L.; Cisse, O.H.; Kutty, G.; Kovacs, J.A. MUC1 mediates Pneumocystis murina binding to airway epithelial cells. Cell. Microbiol. 2020, 22, e13182. [Google Scholar] [CrossRef] [PubMed]
- Bishop, L.R.; Helman, D.; Kovacs, J.A. Discordant antibody and cellular responses to Pneumocystis major surface glycoprotein variants in mice. BMC Immunol. 2012, 13, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbett, A.J.; Eckle, S.B.; Birkinshaw, R.W.; Liu, L.; Patel, O.; Mahony, J.; Chen, Z.; Reantragoon, R.; Meehan, B.; Cao, H.; et al. T-cell activation by transitory neo-antigens derived from distinct microbial pathways. Nature 2014, 509, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Novoa, B.; Bishop, L.; Logun, C.; Munson, P.J.; Elnekave, E.; Rangel, Z.G.; Barb, J.; Danner, R.L.; Kovacs, J.A. Immune responses to Pneumocystis murina are robust in healthy mice but largely absent in CD40 ligand-deficient mice. J. Leukoc. Biol. 2008, 84, 420–430. [Google Scholar] [CrossRef] [Green Version]
- Ripamonti, C.; Bishop, L.R.; Kovacs, J.A. Pulmonary Interleukin-17-Positive Lymphocytes Increase during Pneumocystis murina Infection but Are Not Required for Clearance of Pneumocystis. Infect. Immun. 2017, 85, e00434-16. [Google Scholar] [CrossRef] [Green Version]
- Gherardin, N.A.; Souter, M.N.; Koay, H.-F.; Mangas, K.M.; Seemann, T.; Stinear, T.P.; Eckle, S.B.; Berzins, S.P.; d’Udekem, Y.; Konstantinov, I.E.; et al. Human blood MAIT cell subsets defined using MR1 tetramers. Immunol. Cell Biol. 2018, 96, 507–525. [Google Scholar] [CrossRef] [Green Version]
- Le Bourhis, L.; Martin, E.; Peguillet, I.; Guihot, A.; Froux, N.; Core, M.; Levy, E.; Dusseaux, M.; Meyssonnier, V.; Premel, V.; et al. Antimicrobial activity of mucosal-associated invariant T cells. Nat. Immunol. 2010, 11, 701–708. [Google Scholar] [CrossRef] [Green Version]
- Beard, C.B.; Fox, M.R.; Lawrence, G.G.; Guarner, J.; Hanzlick, R.L.; Huang, L.; del Rio, C.; Rimland, D.; Duchin, J.S.; Colley, D.G. Genetic differences in Pneumocystis isolates recovered from immunocompetent infants and from adults with AIDS: Epidemiological Implications. J. Infect. Dis. 2005, 192, 1815–1818. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the NIH. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bishop, L.R.; Curran, S.J.; Kovacs, J.A. Mucosal-Associated Invariant T Cells Accumulate in the Lungs during Murine Pneumocystis Infection but Are Not Required for Clearance. J. Fungi 2022, 8, 645. https://doi.org/10.3390/jof8060645
Bishop LR, Curran SJ, Kovacs JA. Mucosal-Associated Invariant T Cells Accumulate in the Lungs during Murine Pneumocystis Infection but Are Not Required for Clearance. Journal of Fungi. 2022; 8(6):645. https://doi.org/10.3390/jof8060645
Chicago/Turabian StyleBishop, Lisa R., Shelly J. Curran, and Joseph A. Kovacs. 2022. "Mucosal-Associated Invariant T Cells Accumulate in the Lungs during Murine Pneumocystis Infection but Are Not Required for Clearance" Journal of Fungi 8, no. 6: 645. https://doi.org/10.3390/jof8060645