
Morphological and Molecular Evidence Reveal Eight New Species of Gymnopus from Northeast China

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Morphological Studies
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Data Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Country | Voucher/Specimen Numbers | GenBank Accession Numbers | References | |
|---|---|---|---|---|---|
| ITS | LSU | ||||
| Collybiopsis dichroa | USA | TENN56726 | AY256702 | [46] | |
| Co. filamentipes | USA | TENN-F-065861 | MN897832 | MN897832 | [47] |
| Co. furtiva | USA | SFSU: DED4425 | DQ450031 | AF042650 | [47] |
| Co. hasanskyensis | Russia | TENN-F-060730 | MN897829 | [47] | |
| Co. juniperina | USA | TENN59540 | AY256708 | [14] | |
| Co. melanopus | Indonesia | SFSU: A.W. Wilson 54 | NR_137539 | NG_060624 | [48] |
| Co. melanopus | China | LF1758 | KU529307 | [23] | |
| Co. mesoamericana | Costa Rica | TENN 058613 | NR_119583 | KY019632 | [49,50] |
| Co. minor | USA | TENN-F-059993 | MN413334 | MW396880 | [47] |
| Co. parvula | USA | TENN-F-059993 | MN413334 | Unpublished | |
| Co. stenophylla | USA | TENN59449 | DQ450033 | [46] | |
| Gymnopus alkalivirens | USA | TENN51249 | DQ450000 | [46] | |
| G. alliifoetidissimus | China | GDGM76695 | MT023344 | MT017526 | [25] |
| G. alpinus | Latvia | CB16251 | JX536168 | [51] | |
| G. androsaceus | Russia | TENN-F-59594 | KY026663 | KY026663 | [50] |
| G. androsaceus | France | CBS239.53 | MH857174 | MH868713 | [52] |
| G. aquosus | Czech Republic | BRNM665362 | JX536172 | [51] | |
| G. aurantiipes | AWW118 | AY263432 | AY639410 | [48] | |
| G. bicolor | AWW116 | AY263423 | AY639411 | [48] | |
| G. biformis | USA | TENN58541 | DQ450054 | [48] | |
| G. brunneigracilis | AWW01 | AY263434 | AY639412 | [48] | |
| G. changbaiensis | China | HMJAU60300 | OM030272 | OM033387 | this study |
| G. changbaiensis | China | HMJAU60301 | OM030273 | OM033388 | this study |
| G. changbaiensis | China | HMJAU60302 | OM030274 | OM033389 | this study |
| G. collybioides | Costa Rica | TENN58020 | AF505772 | [46] | |
| G. confluens | Sweden | TENN50524 | DQ450044 | [46] | |
| G. confluens | USA | TENN55695 | DQ450050 | [46] | |
| G. cylindricus | Costa Rica | TENN-058097 | NR_119464 | [49] | |
| G. densilamellatus | Republic of Korea | BRNM714984 | KP336686 | KP336695 | [36] |
| G. dryophilus | Czech Republic | BRNM695586 | JX536143 | [51] | |
| G. dryophilus | Germany | BRNM737691 | JX536139 | [51] | |
| G. dryophilus | China | HMAS290095 | MK966542 | Unpublished | |
| G. dryophilus | Japan | Duke31 | DQ480099 | [46] | |
| G. dryophioides | Republic of Korea | BRNM781447 | MH589967 | MH589985 | [53] |
| G. dysodes | USA | TENN59141 | AF505778 | [46] | |
| G. erythropus | Czech Republic | BRNM714784 | JX536136 | [51] | |
| G. erythropus | USA | JFA12910 | DQ449998 | [46] | |
| G. erythropus | Austria | TENN59329 | AF505786 | [46] | |
| G. erythropus | China | HMJAU60313 | OM030281 | OM033395 | this study |
| G. erythropus | China | HMJAU60315 | OM030280 | OM033396 | this study |
| G. fagiphilus | Czech Republic | BRNM707079 | JX536129 | [51] | |
| G. fusipes | Austria | TENN59300 | AF505777 | [46] | |
| G. fusipes | France | TENN59217 | AY256710 | AY256710 | [14] |
| G. globulosus | China | HMJAU60307 | OM030269 | OM033406 | this study |
| G. globulosus | China | HMJAU60308 | OM030270 | OM033407 | this study |
| G. globulosus | China | HMJAU60308 | OM030271 | OM033408 | this study |
| G. hybridus | Italy | BRNM695773 | JX536177 | [51] | |
| G. inexpectatus | Italy | EU622905 | EU622906 | [54] | |
| G. inusitatus | Spain | SCM B-4058 | JN247553 | JN247557 | [51] |
| G. junquilleus | USA | TENN55224 | NR_119582 | [49] | |
| G. lanipes | Spain | BRNM670686 | JX536137 | [51] | |
| G. longisterigmaticus | China | HMJAU60288 | OM030282 | OM033403 | this study |
| G. longisterigmaticus | China | HMJAU60289 | OM030283 | OM033404 | this study |
| G. longisterigmaticus | China | HMJAU60290 | OM030284 | OM033405 | this study |
| G. longus | China | HMJAU60291 | OM030285 | OM033400 | this study |
| G. longus | China | HMJAU60292 | OM030286 | OM033401 | this study |
| G. longus | China | HMJAU60293 | OM030287 | OM033402 | this study |
| G. macrosporus | China | HMJAU60294 | OM030266 | OM033397 | this study |
| G. macrosporus | China | HMJAU60295 | OM030267 | OM033398 | this study |
| G. macrosporus | China | HMJAU60296 | OM030268 | OM033399 | this study |
| G. ocior | Czech Republic | BRNM699795 | JX536166 | [51] | |
| G. pallipes | China | GDGM81513 | MW582856 | [24] | |
| G. ramulicola | China | GDGM44256 | KU321529 | [27] | |
| G. similis | Republic of Korea | BRNM766739 | KP336692 | KP336699 | [36] |
| G. similis | China | GDGM78308 | MT023352 | MT017530 | [25] |
| G. striatus | China | HMJAU60297 | OM030263 | OM033384 | this study |
| G. striatus | China | HMJAU60298 | OM030264 | OM033385 | this study |
| G. striatus | China | HMJAU60299 | OM030265 | OM033386 | this study |
| G. tiliicola | China | HMJAU60305 | OM030275 | OM033393 | this study |
| G. tiliicola | China | HMJAU60306 | OM030276 | OM033394 | this study |
| G. tiliicola | China | HMJAU60307 | OM030277 | OM033392 | this study |
| G. tomentosus | China | HMJAU60303 | OM030278 | OM033390 | this study |
| G. tomentosus | China | HMJAU60304 | OM030279 | OM033391 | this study |
| Letinula aciculospora | Costa Rica | TENN37996 | AY016443 | [55] | |
| L. boryana | Brazil | TENN58368 | AY016440 | [55] | |
| L. edodes | China | STCL125 | AF031183 | [56] | |
| Marasmius aurantioferrugineus | Republic of Korea | BRNM714752 | FJ904962 | MK278334 | [57] |
| M. brunneospermus | Republic of Korea | KPM-NC0005011 | FJ904969 | FJ904951 | [57] |
| M. maximus | Republic of Korea | BRNM714570 | FJ904976 | FJ904958 | [57] |
| M. nivicola | Republic of Korea | KPM-NC0006038 | FJ904973 | FJ904955 | [57] |
| Marasmiellus ramealis | Sweden | TENN50324 | DQ450030 | [46] | |
| Mycetinis. alliaceus | Russia | TENN-F-55630 | KY696784 | KY696752 | [58] |
| My. curraniae | New Zealand | PDD95301 | KY696778 | [58] | |
| My. opacus | USA | TENN-F-59451 | KY696755 | [58] | |
| My. scorodonius | Switzerland | TENN-F-59451 | KY696725 | [58] | |
| Paragymnopus foliiphilus | USA | TENN-F-68183 | KY026705 | KY026705 | [50] |
| P. perforans | Sweden | TENN-F-50319 | KY026625 | KY026625 | [50] |
| P. pinophilus | USA | TENN-F-69207 | KY026725 | KY026725 | [50] |
| Rhodocollyba butyracea | Sweden | TENN53580 | AY313293 | [46] | |
| R. butyracea | China | HFJAU0269 | MN258680 | Unpublished | |
| R. maculata | Dominican Republic | TFB11720 | KT205402 | [59] | |
| R. maculata | USA | TENN59459 | AY313296 | [46] | |
3. Results
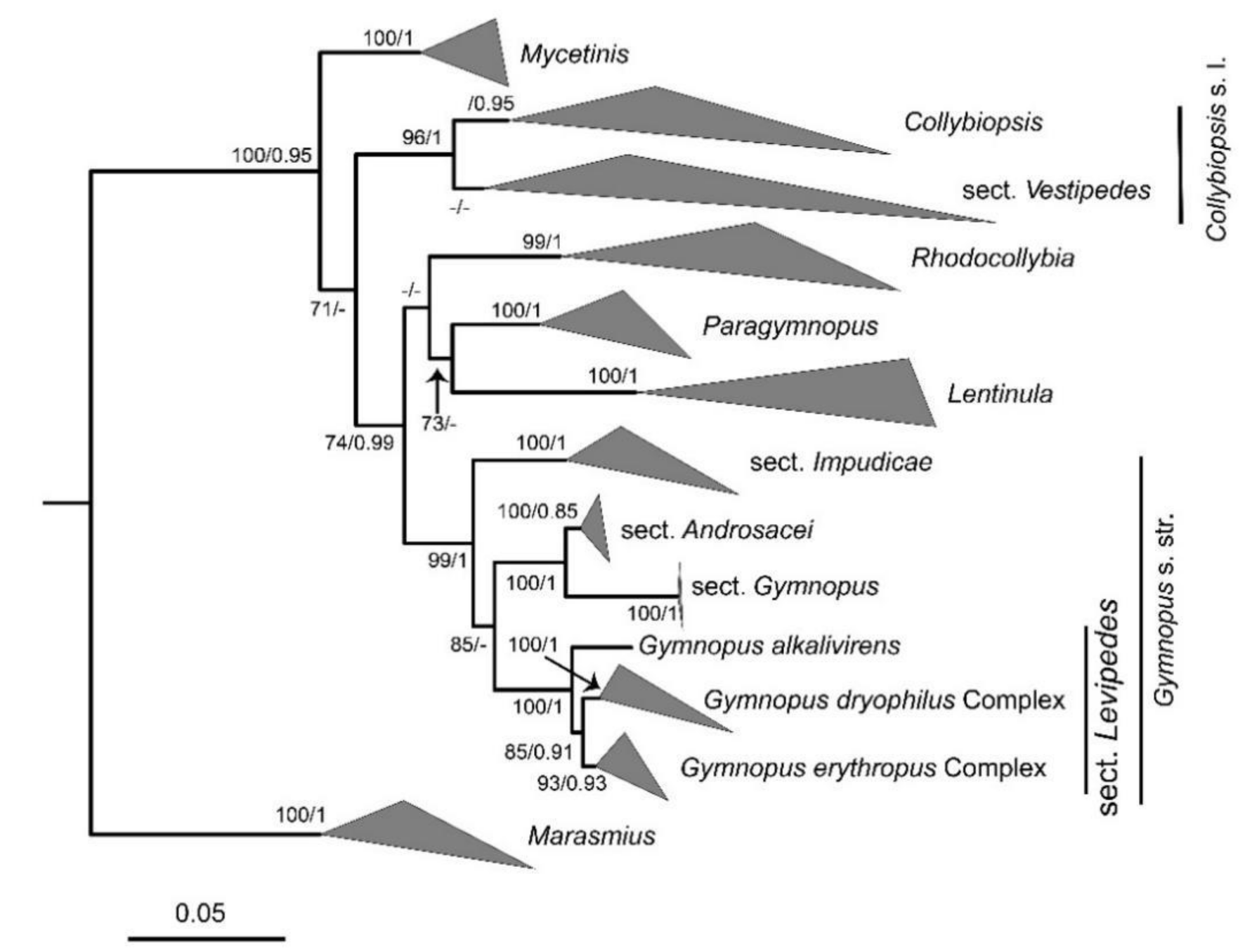
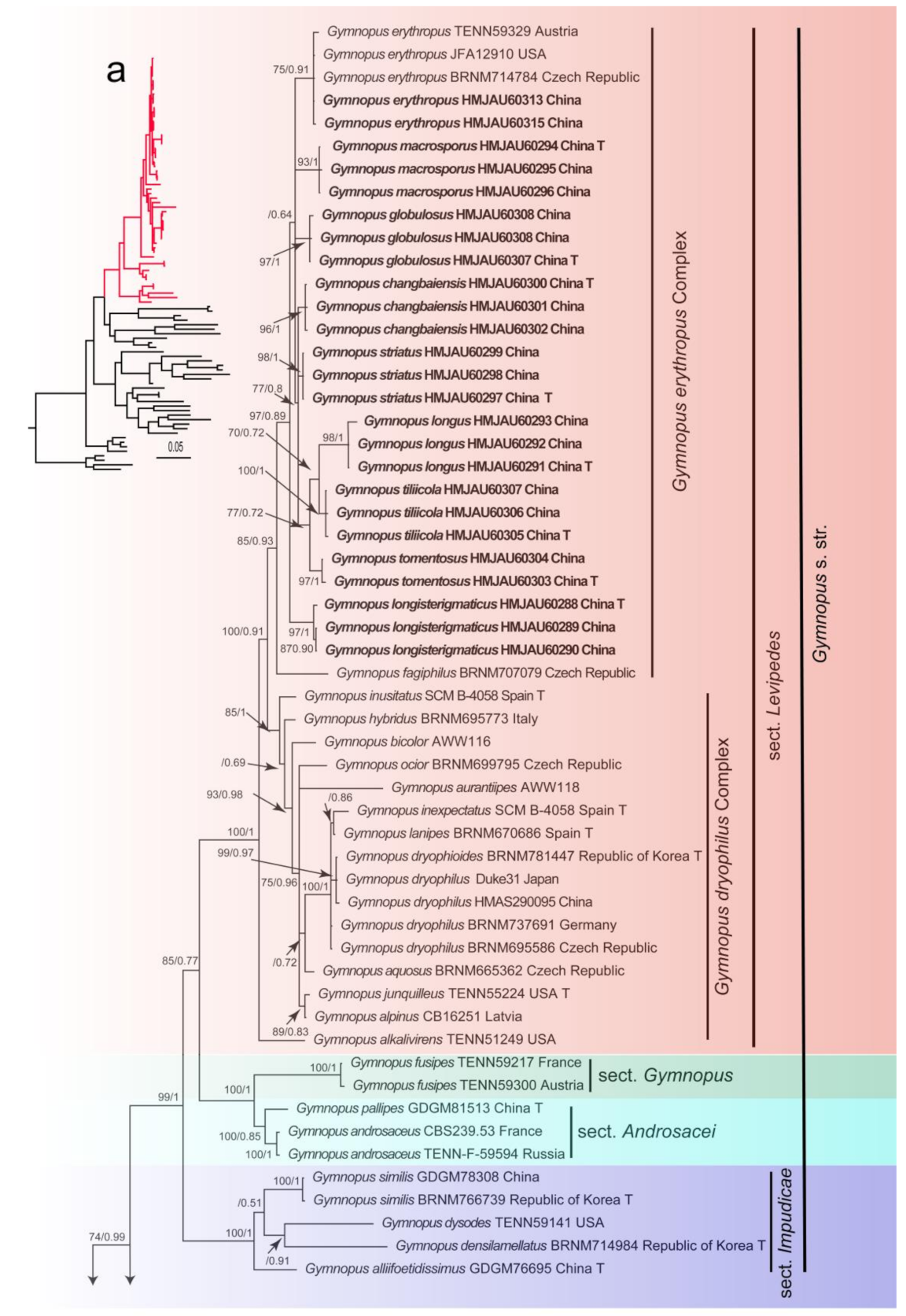
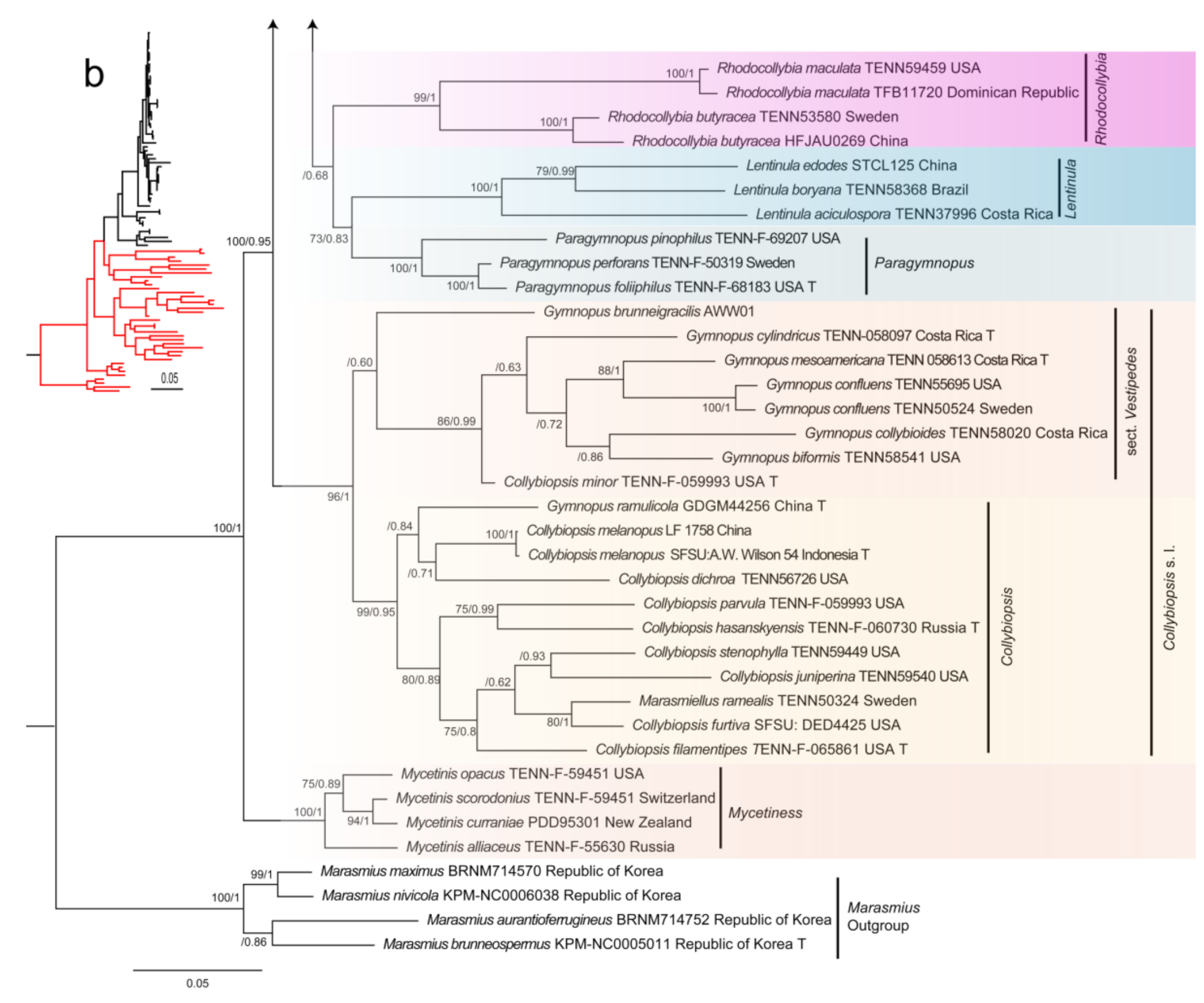
3.1. Phylogenetic Analyses



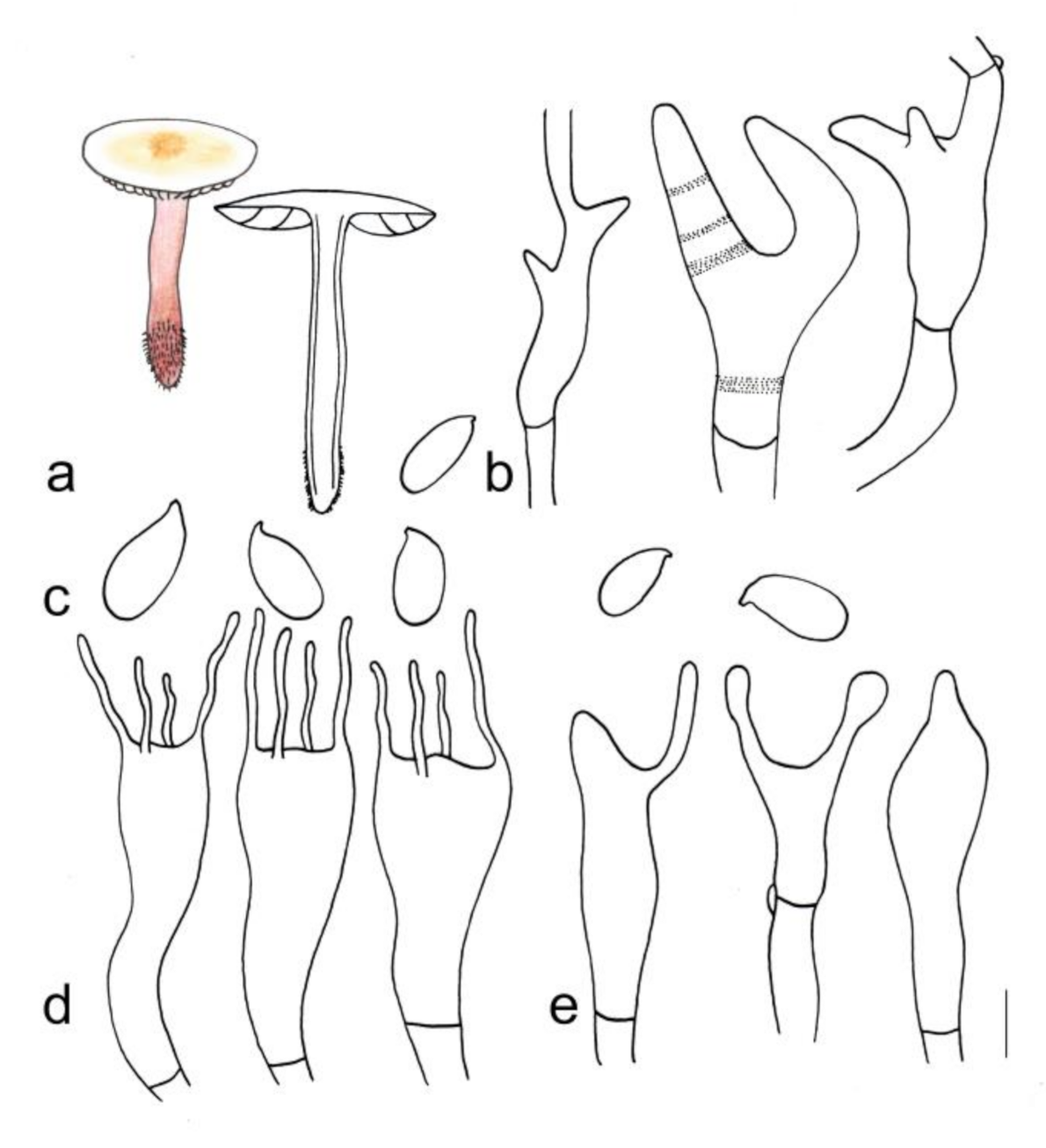
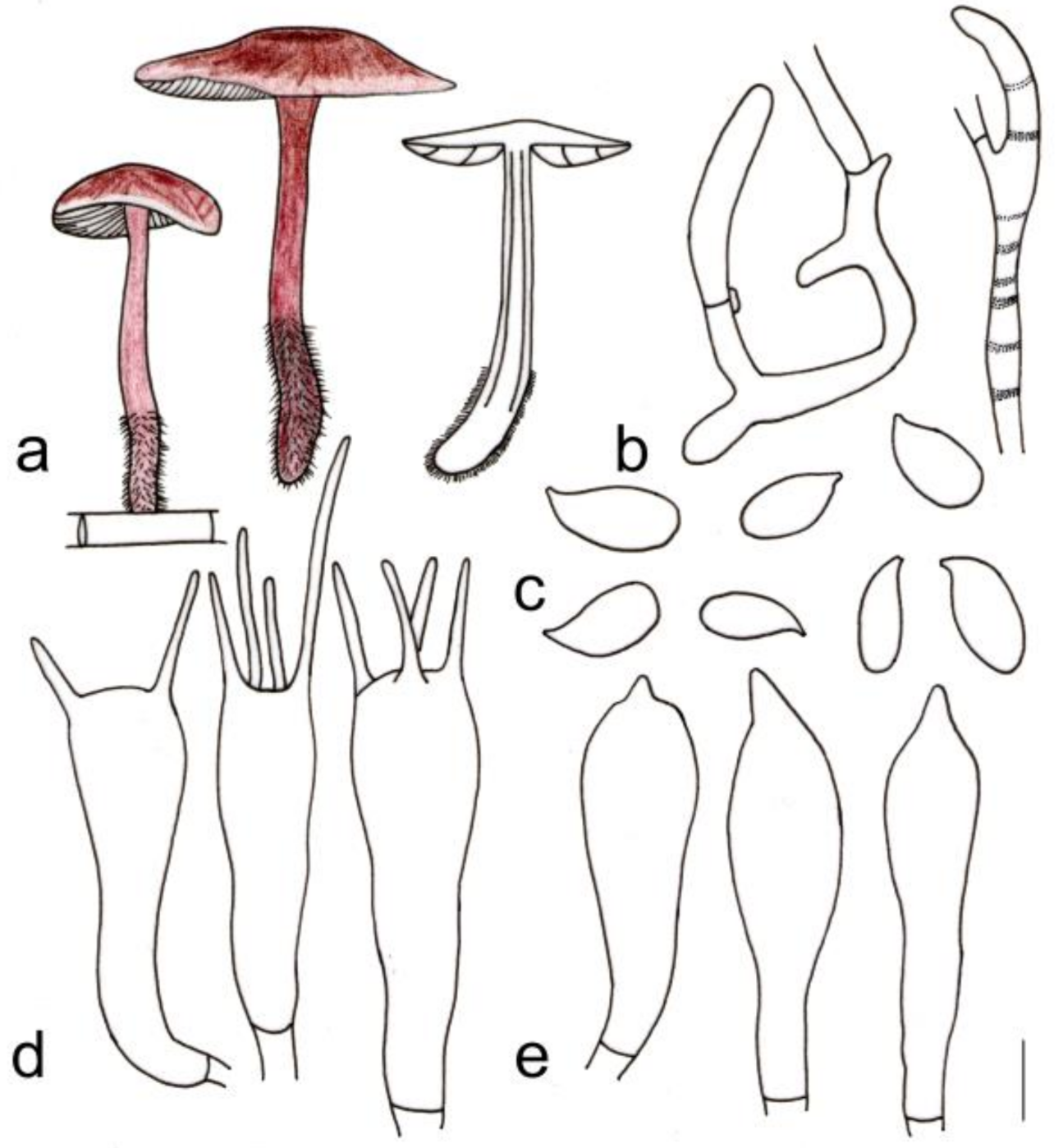
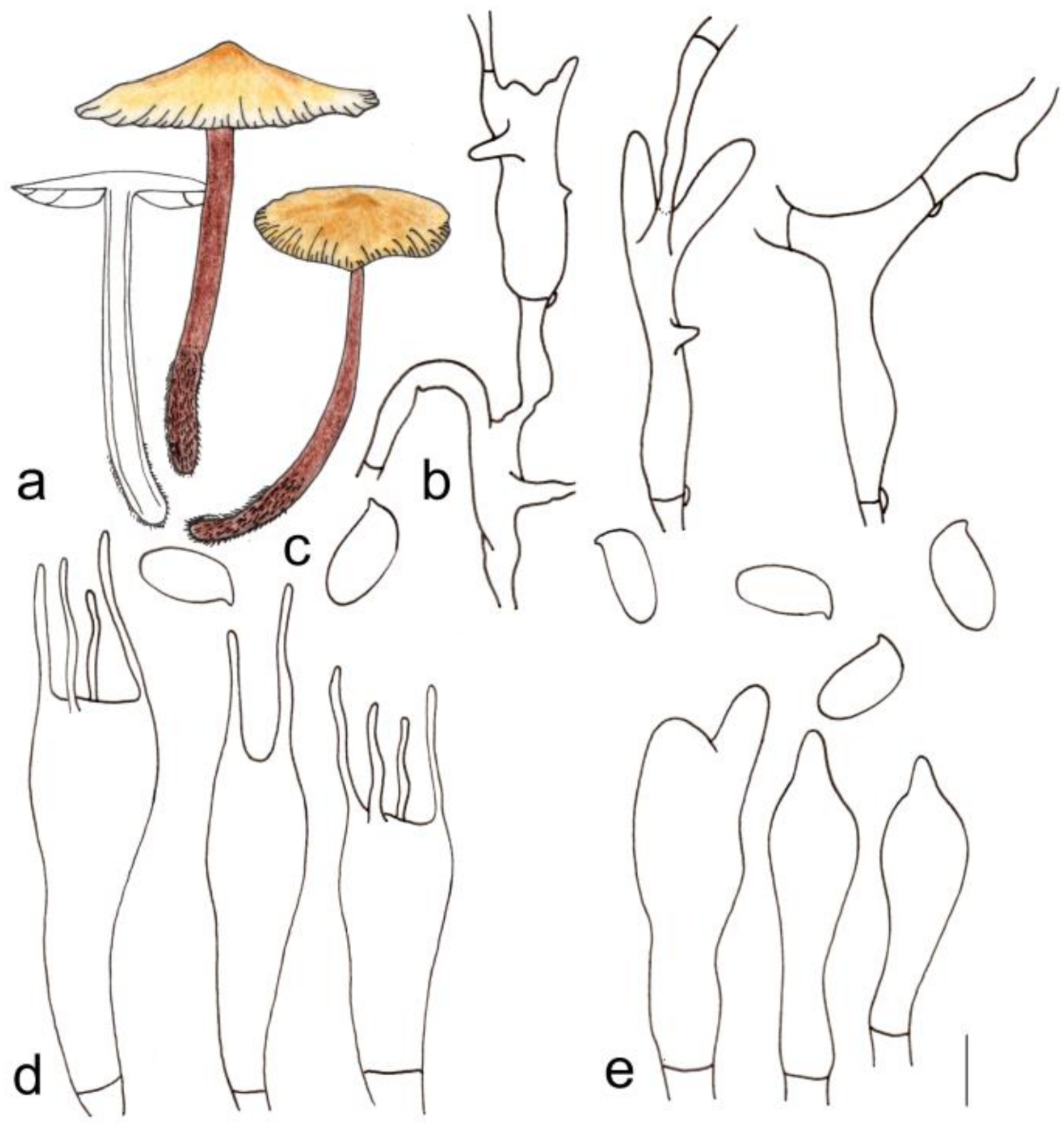
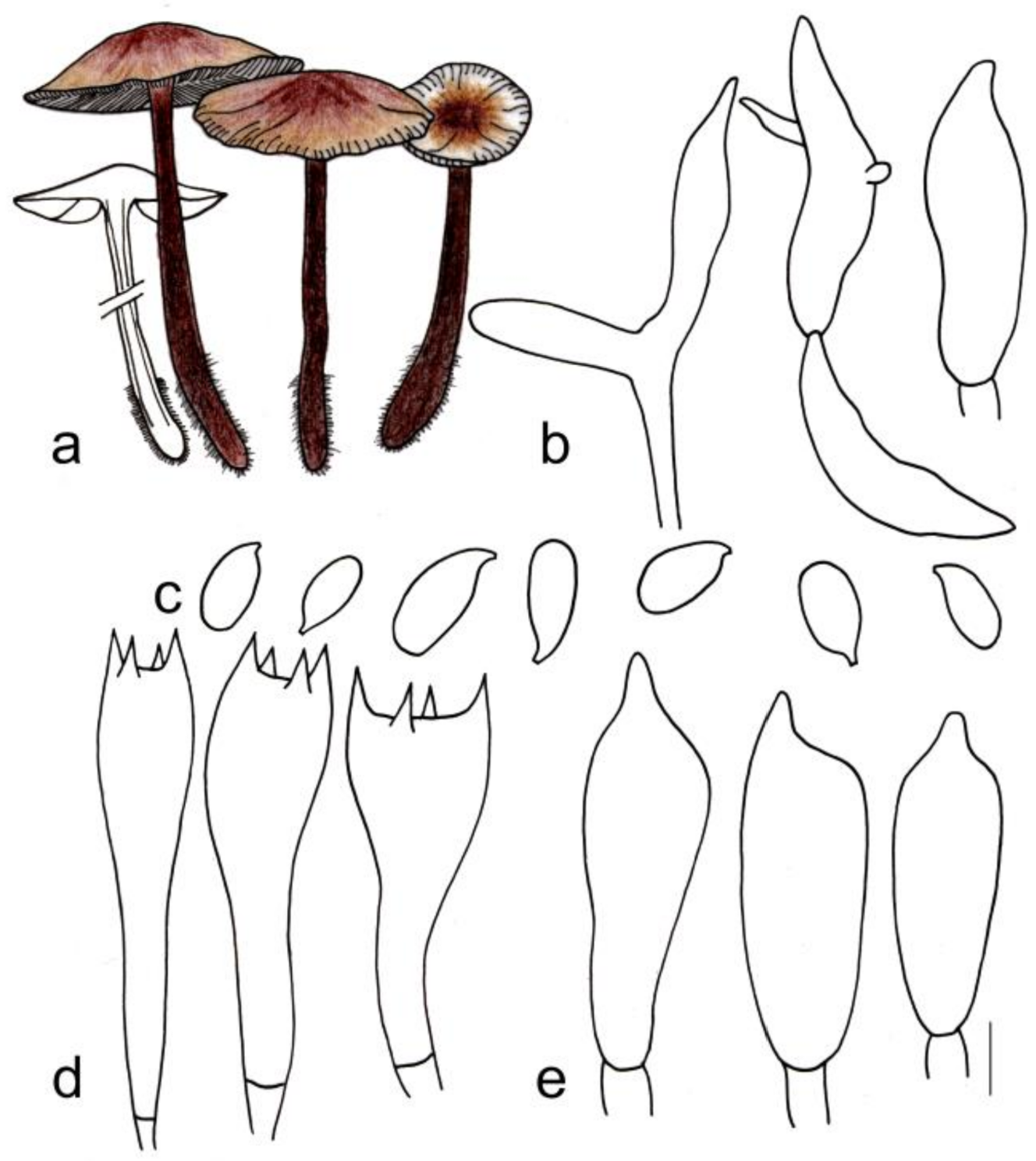
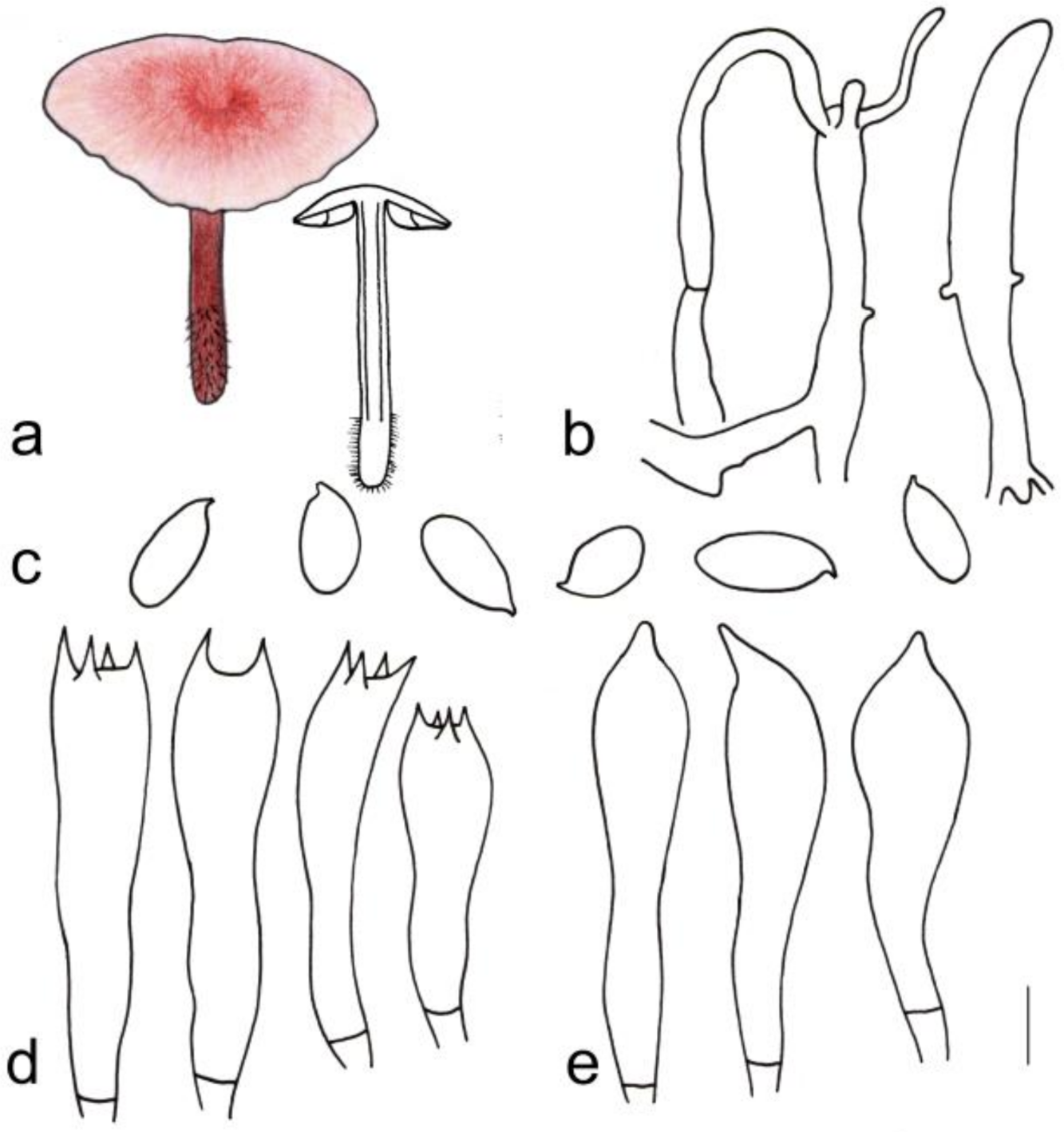
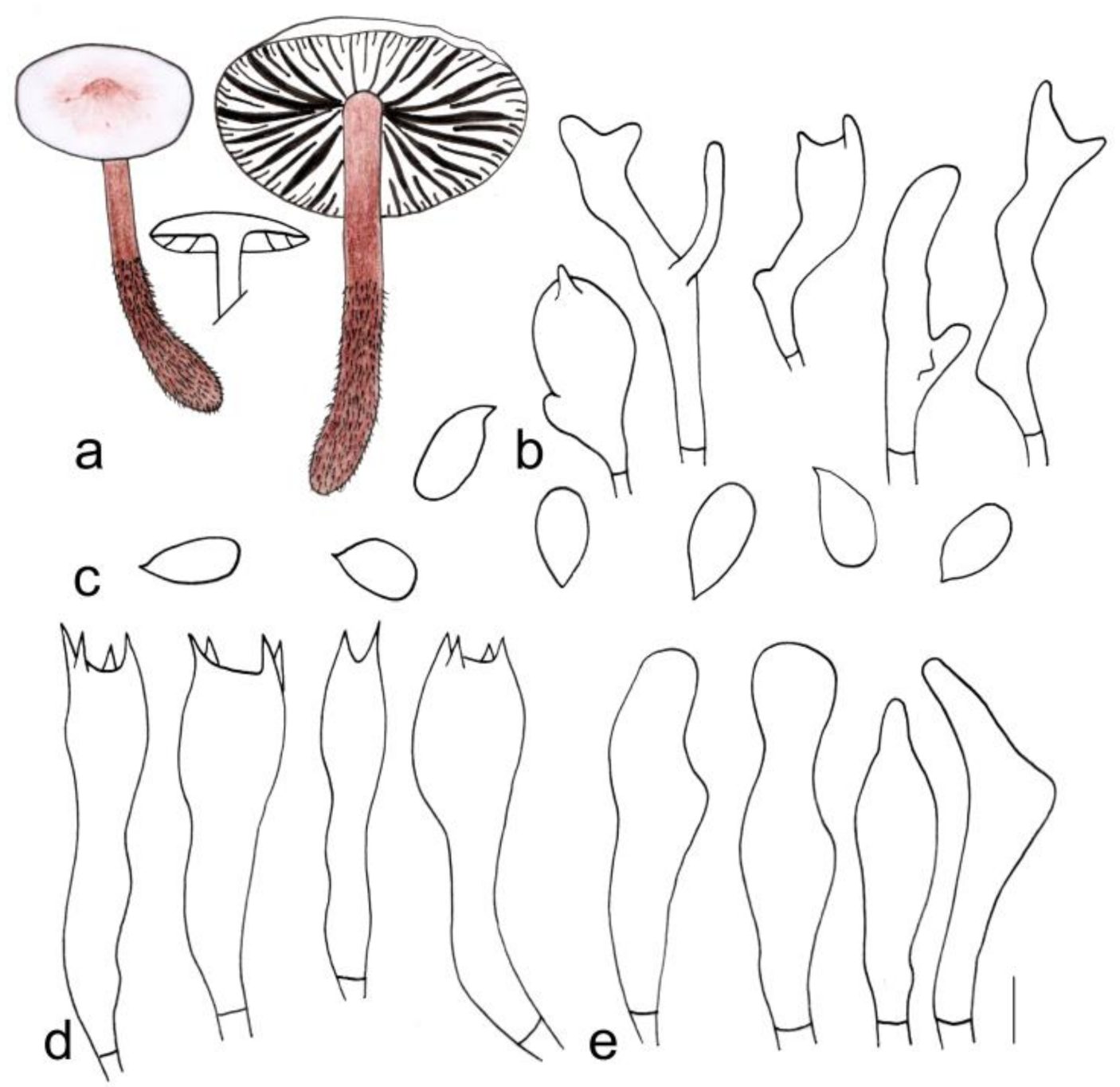
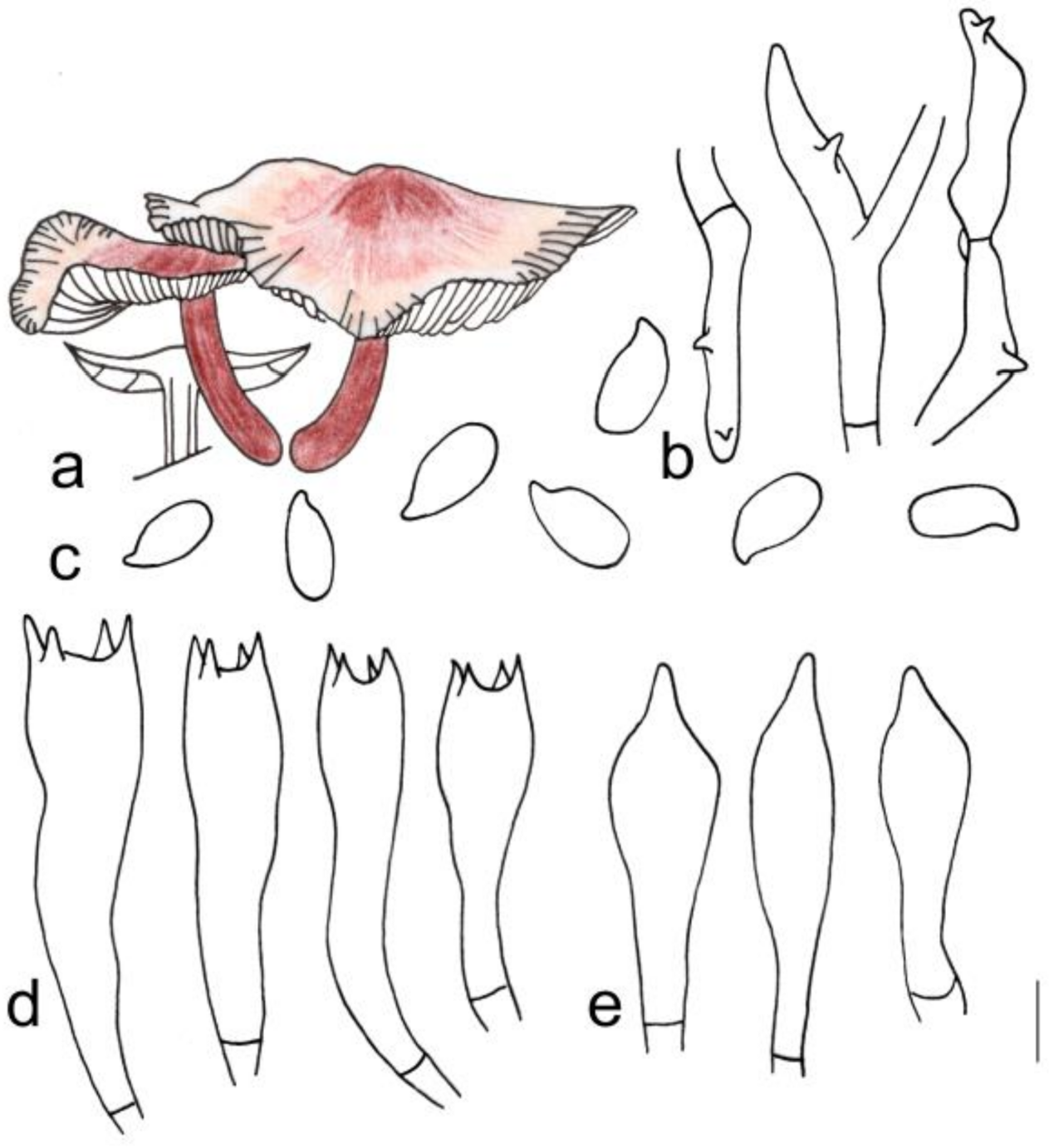
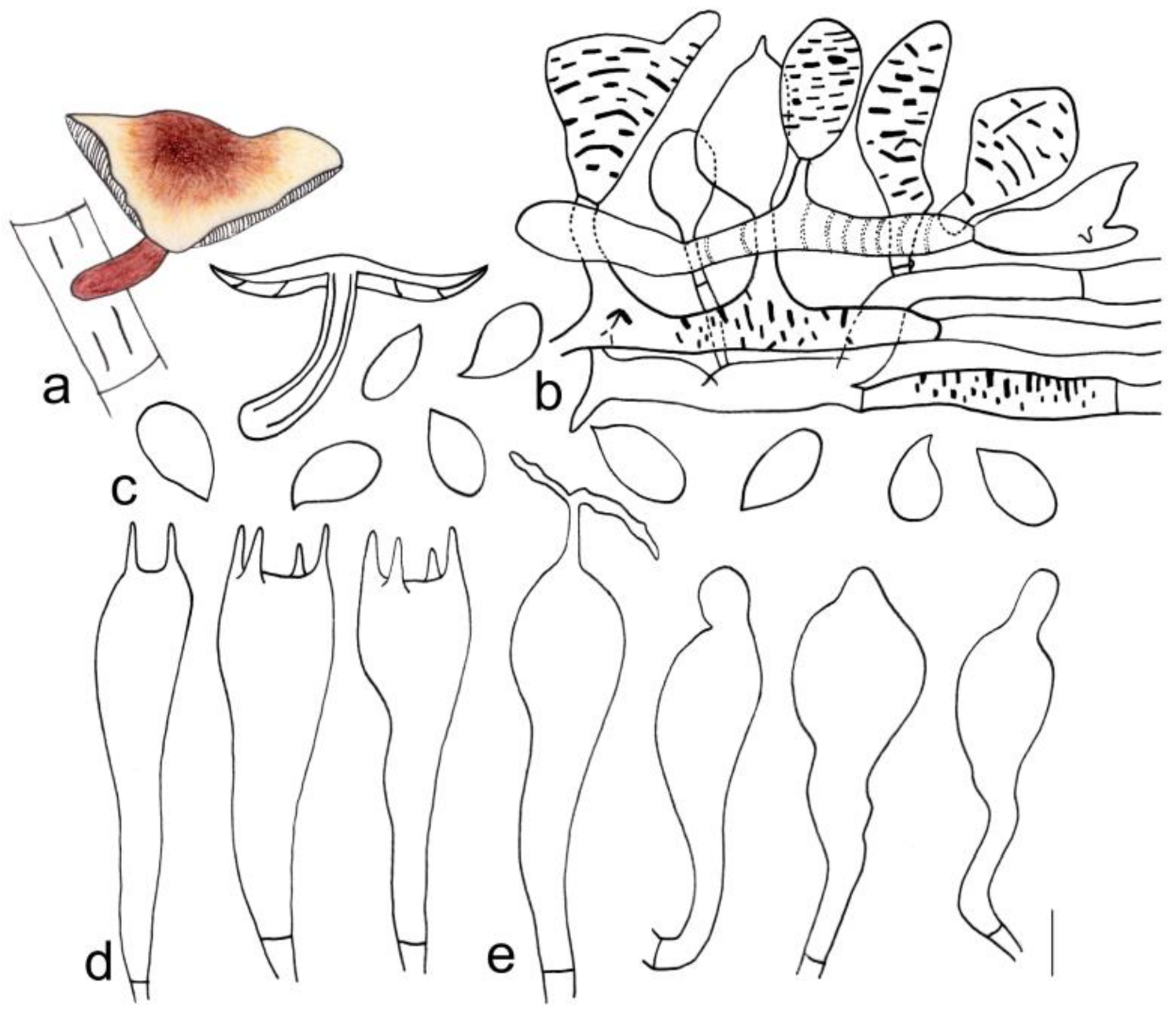
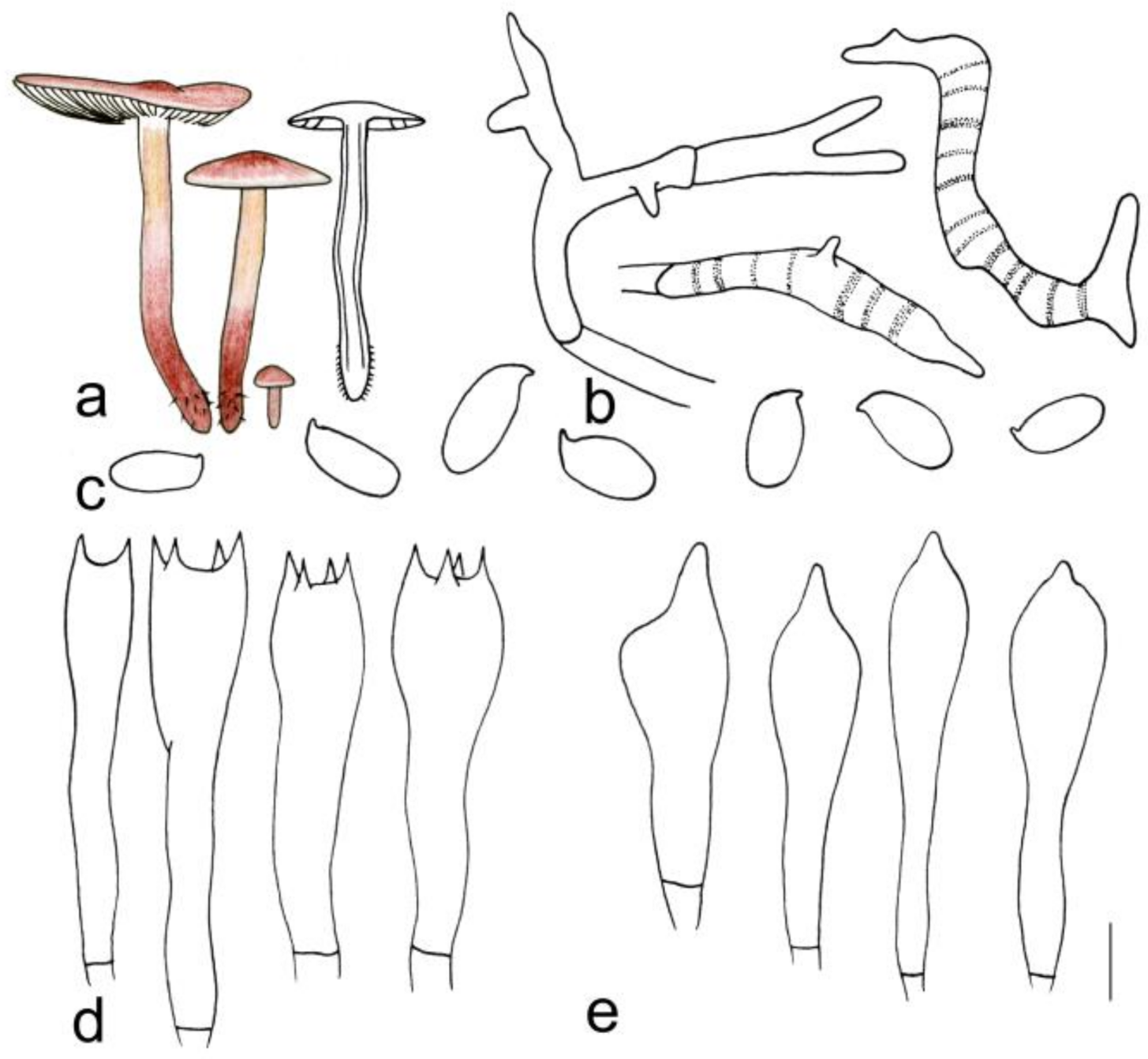
3.2. Taxonomy










| Scientific Name | Pileus | Lamellae | Stipe |
|---|---|---|---|
| G. longisterigmaticus | 1.5–3.2 cm diameter, smooth, brown at center, light brown to yellow towards the margin, margin light yellow to yellowish white, entire | Lamellae subfree to adnate, white to light yellow, I = 1–3, L = 15–18, crowded | 3.2–5.0 × 0.2–0.3 cm, reddish brown, smooth, covered with white to light reddish brown density hairs at base |
| G. longus | 1.7–3.7 cm diameter, smooth, reddish brown at center, towards margin light reddish brown to brown; margin white to light yellow or light brown | Adnate, white to light yellow, I = 5–7, L = 19–24, crowded | 3.7–4.3 cm × 0.3–0.6 cm, reddish brown, with brown farinose on the upper part, and white to light reddish brown tomentose at the base |
| G. macrosporus | 1.2–4.6 cm diameter, smooth, deep reddish brown at center, reddish brown to yellowish brown towards margin; margin beige to light yellow, striped, wavy | Adnexed to adnate or near free, light yellow, I = 1–3, L = 13–17, crowded | 7.8–9.5 × 0.2–0.5 cm, deep reddish brown to reddish brown, smooth, and light yellow to light reddish brown tomentose at the base |
| G. striatus | Depressed when old, 2.3–4.1 cm diameter, smooth, cinnamon at center, brown to light brown towards margin; margin white to light yellow, striped, wavy | Adnate, yellow to light brown, I = 3–9, L = 17–23, crowded | 5.5–7.0 cm × 0.3–0.8 cm, deep reddish brown to reddish brown, longitudinal striped, covered with yellow to light brown hairs up to 1/3 (from the base upwards), fistulose, fibrous |
| G. changbaiensis | 2.1–3.4 cm diameter, smooth, hygrophanus, reddish brown at the center, light pink towards margin; margin white to light pink, striped | Adnate, fresh to pink, I = 1–5, L = 19–24, crowded | 4.2–5.3 cm × 0.2–0.3 cm, deep reddish brown to reddish brown, covered with light yellow to brown hairs up to 1/3 (from the base upwards) |
| G. tomentosus | 1.6–3.0 cm diameter, smooth, tan at the center, light brown towards margin; margin white, tomentose | Adnexed, yellowish green, I = 3–7, L = 19–25, crowded | 3.3–4.3 cm × 0.2–0.5 cm, blackish green at apex, reddish brown below, covered with reddish brown hairs up to 1/4 (from the base upwards) |
| G. tiliicola | 3.0–6.7 cm diameter, smooth, deep rose-red at the center, yellowish pink towards margin; margin white to light yellow, striped | Adnexed to adnate, light pink to fresh, I = 1–3, L = 19–24, crowded | 2.2–4.5 cm × 0.3–0.7 cm, deep reddish brown, smooth |
| G. globulosus | 4.5–5.5 cm diameter, smooth, deep reddish brown at the center, yellowish brown towards margin; margin white to light yellow, striped | Adnexed to adnate, white to light yellowish green, I = 1–3, L = 9–15, crowded | 4.8–6.0 cm × 0.6–0.8 cm, deep reddish brown, paler at apex, smooth |
| G. erythropus | 1.1–3.2 cm diameter, smooth, hygrophanus, reddish brown to brown at the center, light reddish brown to yellowish brown towards margin; margin beige to light yellow | Adnate, fresh-pink, I = 3–5, L = 14–27, crowded | 4.1–10.0 cm × 0.2–0.5 cm, deep reddish brown to light reddish brown, paler at apex, smooth, covered with scattered light yellow to brown hairs hairy at base |
| G. fagiphilus | (7) 15–25 (35) mm broad, when moist slightly translucently striate at margin, yellow-brown or reddish brown, paler at margin | Moderately distant, L = 18–21, I = 3–7 (15), free or narrowly adnate, rarely adnate, pinkish brown or pinkish cream, darker with age or tinged gray | 20–40 (70) × 1–3 mm, orange-brown to red-brown, sometimes paler at apex, dark red-brown towards base, covered with fine, white or yellow hairs up to 2/3 of length (from base upwards) |
| Scientific Name | Pileipellis | Q | Qm | Basidiospores | Basidia | Cheilocystidia | Caulocystidia |
|---|---|---|---|---|---|---|---|
| G. longisterigmaticus | Cutis, irregularly branched hyphae, inflated, 10–27 (35) µm wide, hyaline to light yellow, smooth or pigment yellowish brown incrusting in pileipellis | (1.40) 1.67–2.25 (2.26) | 1.93 ± 0.20 | (6.2) 6.7–9.0 × (3.0) 3.1–4.3 (5.0) μm | Clavate, (18) 19–27 (28) × (5) 6–10 µm, four-spored; sterigmata extremely long, up to 40 µm | Clavate, with obtuse on the top, or branched, (16) 18–27 × (4) 5–8 (9) µm | None |
| G. longus | A translation between a cutis and a trichoderm, made up of irregularly interwoven, repent or ascending inflated hyphae with inflated and irregularly branched terminal elements, hyaline to light brown, (6) 7–13 (15) µm wide, smooth or pigment yellowish brown incrusting in pileipellis | (1.27) 1.47–2.19 (2.58) | 1.80 ± 0.24 | (5.6) 6.0–8.0 × (3.0)3.1–4.1 (4.9) μm | (19) 20–28 (29) × 6–9 µm, two- or four-spored, clavate; sterigmata extremely long, up to 33 µm long | Clavate, (21) 22–29 (30) × 5–7 µm, with obtuse on the top | None |
| G. macrosporus | Cutis, made up of irregular branched or weakly coralloid hyphae, inflated, 10–27 (35) µm wide, hyaline to light yellow, smooth | (1.63) 1.67–2.32 (2.37) | 1.88 ± 0.18 | (6.0) 6.8–7.9 (8.3) × (3.0) 3.1–4.2 (4.3) μm | Clavate, 20–29 × 6–9 µm, two- or four-spored, thin-walled; sterigmata extremely long, up to 32 µm | Clavate, with obtuse on the top, 20–28 (30) × 5 (6)–9 µm | None |
| G. striatus | Cutis, made up of irregular branched or weakly coralloid hyphae, inflated, 10–30 (35) µm wide, hyaline to light yellow, smooth | (1.50) 1.58–2.50 (2.60) | 2.01 ± 0.25 | 6.0–8.0 (9.0) × 3.0–4.0 μm | Clavate, 20 (21)–34 (37) × 5–10 µm, two- or four-spored | Clavate, with obtuse on the top, (17) 20–30 × 4–8 (10) µm | None |
| G. changbaiensis | Cutis, made up of irregular branched or weakly coralloid hyphae, inflated, 8–23 (25) µm wide, hyaline to light yellow | (1.41) 1.53–2.40 (2.50) | 1.98 ± 0.24 | (5.8) 6.0–8.1 (9.0) × 3.0–4.1(4.2) μm | Clavate, (19) 20–29 (32) × 5–8 µm, two- or four-spored | Clavate, with obtuse on the top, (23) 24–34 (39) × (5) 6–7 (9) µm | None |
| G. tomentosus | Cutis, made up of irregular branched to weakly coralloid or bulbous hyphae, inflated, 10–18 (21) µm wide, light brown | (1.50) 1.59–2.33 (2.40) | 1.92 ± 0.23 | (6.0) 6.2–8.2 (9.0) × 3.0–4.1 (4.2) μm | Clavate, 20–30 (31) × 5–8 µm, two- or four-spored | Clavate, with obtuse on the top sometimes, (20) 22–30 (32) × 5–7 µm | None |
| G. tiliicola | Cutis, made up of irregular branched to weakly coralloid hyphae, inflated, (5)6–15(17) µm wide, light brown, smooth | (1.70) 1.75–2.26 (2.33) | 1.93 ± 0.17 | (6.0) 6.9–8.0 (8.2) × (3.0) 3.1–4.0 (4.2) μm | Clavate, 20–30 × 6–8 µm, two- or four-spored | Clavate, with obtuse on the top, (20) 21–27 (28) × 5–7 µm | None |
| G. globulosus | Layered, the upper layer inflated to spherical to prolate hyphae, 15–33 (47) µm wide, brown, smooth, thin-walled; down layer made up of branched and inflated hyphae, pigment light brown to brown incrusting in pileipellis, thin-to-thick-walled | (1.63) 1.75–2.20 (2.26) | 1.93 ± 0.16 | (6.8) 7.0–8.8(9.0) × (3.1) 3.3–4.2 (4.8) μm | Clavate, (23) 25–32 (33) × 6–9 (11) µm, two- or four-spored | Clavate, with obtuse on the top, (22) 24–38 (39) × 5–9 (10) µm | None |
| G. erythropus | Cutis, made up of irregular branched or weakly coralloid hyphae, inflated, (6) 8–20 (20) µm wide, hyaline to light yellow, smooth | (1.20) 1.48–2.33 (3.00) | 1.87 ± 0.27 | (5.0) 6.0–8.2 (10.0) × (2.1) 3.0–5.0 (6.0) µm | Clavate, (17) 21–33 (38) × (4) 5–9 (10) µm, two- or four-spored | Clavate, with obtuse on the top, (15) 21–33 (39) × (3) 4–8 (9) µm, thin-walled, smooth, hyaline | None |
| G. fagiphilus | Cutis with transitions to a trichoderm, made up of irregularly shaped, 4.0–15 (25) μm–wide coralloid elements (“Dryophila-structure”); pigment brown-yellow, incrusting in pileipellis | 1.7–2.3 | 2.1 | (6.0) 7.0–9.0 × (3.0) 3.5–4.5 μm | 21–31 × 6.0–8.5 μm, 4-spored | 15–40 (60) × 4.0–8.0 (10) μm, irregularly clavate, often with lobed apex or with short to long, up to 10 μm long rostrum, sometimes very slender lageniform | 20–80 (120) × 4.0–12 μm, subcylindrical or sublageniform, numerous |
4. Discussion
4.1. New Sights on Morphological Characteristics
4.2. Phylogenetic Relationships of Gymnopus s.l. with Related Genera
4.3. Nova Suggestions of Phylogenetic Relationships within Gymnopus s. str.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Antonín, V.; Noordeloos, M.E. A Monograph of Marasmioid and Collybioid Fungi in Europ; IHW Verl: Eching, Germany, 2010. [Google Scholar]
- Persoon, C.H. Synopsis Methodica Fungorum 1; Apud Henricum Dieterich: Göttingen, Germany, 1801. (In Latin) [Google Scholar]
- Fries, E.M. Systema Mycologicum: Sistens Fungorum Ordines, Genera Et Species, Huc Usque Cognitas; Ex officina Berlingiana: Lund, Berling, Germany, 1821; Volume 1. (In Latin) [Google Scholar]
- Staude, F. Die Schwämme Mitteldeutschlands, Insbesondere Des Herzogtums Coburg; Dietz: Coburg, Germany, 1857; Volume 1. (In German) [Google Scholar]
- Singer, R. The Agaricales in Modern Taxonomy, 2nd ed.; J. Cramer: Vaduz, Germany, 1962. [Google Scholar]
- Singer, R. The Agaricales in Modern Taxonomy, 3rd ed.; J. Cramer: Vaduz, Germany, 1975. [Google Scholar]
- Singer, R. The Agaricales in Modern Taxonomy, 4th ed.; Koeltz Botanical Books: Koenigstein, Germany, 1986. [Google Scholar]
- Halling, R.E. The Genus Collybia (Agaricales) in the Northeastern United States and Adjacent Canada. Mycol. Mem. 1983, 8, 1–148. [Google Scholar]
- Antonín, V.; Noordeloos, M.E. A Monograph of Marasmius, Collybia and Related Genera in Europe. Part 1: Marasmius, Setulipes, and Marasmiellus; Libri Botanici; IHW-Verlag: Eching, Germany, 1993; Volume 8, pp. 1–229. [Google Scholar]
- Antonín, V.; Noordeloos, M.E. A Monograph of Marasmius, Collybia and Related Genera in Europe. Part 2: Collybia, Gymnopus, Rhodocollybia, Crinipellis, Chaetocalathus, and Additions to Marasmiellus; Libri Botanici; IHW-Verlag: Eching, Germany, 1997; Volume 17, pp. 1–256. [Google Scholar]
- Antonín, V.; Halling, R.; Noordeloos, M. Generic Concepts within the Groups of Marasmius and Collybia Sensu Lato. Mycotaxon 1997, 63, 359–368. [Google Scholar]
- Wilson, A.W.; Desjardin, D.E. Phylogenetic Relationships in the Gymnopoid and Marasmioid Fungi (Basidiomycetes, Euagarics Clade). Mycologia 2005, 97, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Moncalvo, J.M.; Vilgalys, R.; Redhead, S.A.; Johnson, J.E.; James, T.Y.; Catherine Aime, M.; Hofstetter, V.; Verduin, S.J.W.; Larsson, E.; Baroni, T.J.; et al. One Hundred and Seventeen Clades of Euagarics. Mol. Phylogenet. Evol. 2002, 23, 357–400. [Google Scholar] [CrossRef]
- Mata, J.L.; Hughes, K.W.; Petersen, R.H. Phylogenetic Placement of Marasmiellus juniperinus. Mycoscience 2004, 45, 214–221. [Google Scholar] [CrossRef]
- Oliveira, J.J.; Vargas-Isla, R.; Cabral, T.S.; Rodrigues, D.P.; Ishikawa, N.K. Progress on the Phylogeny of the Omphalotaceae: Gymnopus S. Str., Marasmiellus S. Str., Paragymnopus gen. nov. and Pusillomyces gen. nov. Mycol. Prog. 2019, 18, 713–739. [Google Scholar] [CrossRef]
- Petersen, R.H.; Hughes, K.W. Two New Genera of Gymnopoid/Marasmioid Euagarics. Mycotaxon 2020, 135, 1–95. [Google Scholar] [CrossRef]
- Halling, R.E. Notes on Collybia V. Gymnopus Section Levipedes in Tropical South America, with Comments on Collybia. Brittonia 1996, 48, 487–494. [Google Scholar] [CrossRef]
- Singer, R. Type Studies in Basidiomycetes. X. Pers. Mol. Phylogeny Evol. Fungi 1961, 2, 1–62. [Google Scholar]
- Jansen, A. Het Geslacht Collybia (De Fungi Van Nederland); Stichting Uitgeverij Koninklijke Nederlandse Natuurhistorische Vereniging: Zeist, Nederlands, 1991. (In Dutch) [Google Scholar]
- Kirk, P.; Cannon, P.; Minter, D.; Stalpers, J. Dictionary of the Fungi, 10th ed.; CABI International: Wallingford, UK, 2008. [Google Scholar]
- Teng, S.Q. Fungi of China; Science Press, Academic Sinica: Beijing, China, 1963. [Google Scholar]
- Tai, F.L. Sylloge Fungorum Sinicorum; Science Press, Academic Sinica: Beijing, China, 1979. (In Latin) [Google Scholar]
- Deng, S.F. Taxonomy of Gymnopus and Preliminary Study of Marasmiaceae Resource in South China. Master’s Thesis, South China Agricultural University, Guangzhou, China, 2016. [Google Scholar]
- Li, J.-P.; Song, B.; Feng, Z.; Wang, J.; Deng, C.-Y.; Yang, Y.-H. A New Species of Gymnopus Sect. Androsacei (Omphalotaceae, Agaricales) from China. Phytotaxa 2021, 521, 1–14. [Google Scholar] [CrossRef]
- Li, J.-P.; Li, Y.; Li, T.-H.; Antonín, V.; Hosen, I.; Song, B.; Xie, M.-L.; Feng, Z. A Preliminary Report of Gymnopus Sect. Impudicae (Omphalotaceae) from China. Phytotaxa 2021, 497, 263–276. [Google Scholar] [CrossRef]
- Mešić, A.; Tkalčec, Z.; Deng, C.-Y.; Li, T.-H.; Pleše, B.; Ćetković, H. Gymnopus Fuscotramus (Agaricales), a New Species from Southern China. Mycotaxon 2011, 117, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.-F.; Li, T.-H.; Jiang, Z.-D.; Song, B. Gymnopus Ramulicola sp. nov., a Pinkish Species from Southern China. Mycotaxon 2016, 131, 663–670. [Google Scholar] [CrossRef]
- Wang, X.S.; Bao, J.S.; Bao, H.; Feng, J. Macrofungal Diversity in Hanwula National Nature Reserve, Inner Mongolia. Mycosystema 2020, 39, 695–706. [Google Scholar] [CrossRef]
- Sun, Y.L.; Luo, Y.; Bau, T. Notes on Basidiomycetes of Jilin Province (X). J. Fungal Res. 2021, 19, 139–147. [Google Scholar] [CrossRef]
- Royal Botanic Garden, E. Flora of British Fungi: Colour Identification Chart; HM Stationery Office: Edinburgh, UK, 1969. [Google Scholar]
- César, E.; Bandala, V.M.; Montoya, L.; Ramos-Fernández, A. A New Gymnopus Species with Rhizomorphs and Its Record as Nesting Material by Birds (Tyrannideae) in the Subtropical Cloud Forest from Eastern Mexico. MycoKeys 2018, 42, 21–34. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. Its Primers with Enhanced Specificity for Basidiomycetes-Application to the Identification of Mycorrhizae and Rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Cubeta, M.A.; Echandi, E.; Abernethy, T.; Vilgalys, R. Characterization of Anastomosis Groups of Binucleate Rhizoctonia Species Using Restriction Analysis of an Amplified Ribosomal Rna Gene. Phytopathology 1991, 81, 1395–1400. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid Genetic Identification and Mapping of Enzymatically Amplified Ribosomal DNA from Several Cryptococcus Species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Coimbra, V.R.M.; Pinheiro, F.G.B.; Wartchow, F.; Gibertoni, T.B. Studies on Gymnopus Sect. Impudicae (Omphalotaceae, Agaricales) from Northern Brazil: Two New Species and Notes on G. Montagnei. Mycol. Prog. 2015, 14, 110. [Google Scholar] [CrossRef]
- Ryoo, R.; Antonín, V.; Ka, K.-H.; Tomšovský, M. Marasmioid and Gymnopoid Fungi of the Republic of Korea. 8. Gymnopus Section Impudicae. Phytotaxa 2016, 286, 75–88. [Google Scholar] [CrossRef]
- Thonpson, J. The Clustal X Windows Interface: Flexible Strategies for Multiple Sequence Alignment Aided by Quality Analysis Tools. Nucl. Acids Res. 1997, 24, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Ranwez, V.; Douzery, E.J.P.; Cambon, C.; Chantret, N.; Delsuc, F. Macse V2: Toolkit for the Alignment of Coding Sequences Accounting for Frameshifts and Stop Codons. Mol. Biol. Evol. 2018, 35, 2582–2584. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. Mafft Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Hall, T. Bioedit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/Nt. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zhou, H.; Zhang, J.; Li, W.X.; Wang, G.T. Phylosuite: An Integrated and Scalable Desktop Platform for Streamlined Molecular Sequence Data Management and Evolutionary Phylogenetics Studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. Modelfinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. Mrbayes 3: Bayesian Phylogenetic Inference under Mixed Models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. Raxmlgui 2.0: A Graphical Interface and Toolkit for Phylogenetic Analyses Using Raxml. Methods Ecol. Evol. 2021, 12, 373–377. [Google Scholar] [CrossRef]
- Vizzini, A.; Antonín, V.; Sesli, E.; Contu, M. Gymnopus Trabzonensis sp. nov. Omphalotaceae and Tricholoma Virgatum var. Fulvoumbonatum var. nov. Tricholomataceae, Two New White-Spored Agarics from Turkey. Phytotaxa 2015, 226, 119–130. [Google Scholar] [CrossRef]
- Mata, J.L.; Hughes, K.W.; Petersen, R.H. An Investigation of/Omphalotaceae (Fungi: Euagarics) with Emphasis on the Genus Gymnopus. SYDOWIA-HORN- 2006, 58, 191–289. [Google Scholar]
- Petersen, R.H.; Hughes, K.W. Collybiopsis and Its Type Species, Co. Ramealis. Mycotaxon 2021, 136, 263–349. [Google Scholar] [CrossRef]
- Wilson, A.; Desjardin, D.; Horak, E. Agaricales of Indonesia: 5. The Genus Gymnopus from Java and Bali. Sydowia 2004, 56, 137–210. [Google Scholar]
- Schoch, C.L.; Robbertse, B.; Robert, V.; Vu, D.; Cardinali, G.; Irinyi, L.; Meyer, W.; Nilsson, R.H.; Hughes, K.; Miller, A.N.; et al. Finding Needles in Haystacks: Linking Scientific Names, Reference Specimens and Molecular Data for Fungi. Database 2014, 2014, bau061. [Google Scholar] [CrossRef]
- Petersen, R.H.; Hughes, K.W. Micromphale Sect. Perforantia (Agaricales, Basidiomycetes); Expansion and Phylogenetic Placement. MycoKeys 2016, 18, 1–122. [Google Scholar] [CrossRef]
- Antonín, V.; Sedlák, P.; Tomšovský, M. Taxonomy and Phylogeny of European Gymnopus Subsection Levipedes (Basidiomycota, Omphalotaceae). Persoonia 2013, 31, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Vu, D.; Groenewald, M.; De Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-Scale Generation and Analysis of Filamentous Fungal DNA Barcodes Boosts Coverage for Kingdom Fungi and Reveals Thresholds for Fungal Species and Higher Taxon Delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Ryoo, R.; Antonín, V.; Ka, K.H. Marasmioid and Gymnopoid Fungi of the Republic of Korea. 8. Gymnopus Section Levipedes. Mycobiology 2020, 48, 252–262. [Google Scholar] [CrossRef]
- Vizzini, A.; Consiglio, G.; Antonin, V.; Contu, M. A New Species within the Gymnopus Dryophilus Complex (Agaricomycetes, Basidiomycota) from Italy. Mycotaxon 2008, 105, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Mata, J.L.; Petersen, R.H.; Hughes, K.W. The Genus Lentinula in the Americas. Mycologia 2001, 93, 1102–1112. [Google Scholar] [CrossRef]
- Hibbett, D.S.; Hansen, K.; Donoghue, M.J. Phylogeny and Biogeography of Lentinula Inferred from an Expanded Rdna Dataset. Mycol. Res. 1998, 102, 1041–1049. [Google Scholar] [CrossRef] [Green Version]
- Antonín, V.; Ryoo, R.; Shin, H.-D. Marasmioid and Gymnopoid Fungi of the Republic of Korea. 2. Marasmius Sect. Globulares. Persoonia 2010, 24, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Petersen, R.H.; Hughes, K.W. An Investigation on Mycetinis (Euagarics, Basidiomycota). MycoKeys 2017, 24, 1–139. [Google Scholar] [CrossRef] [Green Version]
- Mata, J.L.; Ovrebo, C.L.; Baroni, T.J.; Hughes, K.W. New Species of Neotropical Rhodocollybia. Mycotaxon 2016, 131, 235–245. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Zhao, G.; Tuo, Y.; Rao, G.; Zhang, Z.; Qi, Z.; Yue, L.; Liu, Y.; Zhang, T.; Li, Y.; et al. Morphological and Molecular Evidence Reveal Eight New Species of Gymnopus from Northeast China. J. Fungi 2022, 8, 349. https://doi.org/10.3390/jof8040349
Hu J, Zhao G, Tuo Y, Rao G, Zhang Z, Qi Z, Yue L, Liu Y, Zhang T, Li Y, et al. Morphological and Molecular Evidence Reveal Eight New Species of Gymnopus from Northeast China. Journal of Fungi. 2022; 8(4):349. https://doi.org/10.3390/jof8040349
Chicago/Turabian StyleHu, Jiajun, Guiping Zhao, Yonglan Tuo, Gu Rao, Zhenhao Zhang, Zhengxiang Qi, Lei Yue, Yajie Liu, Tong Zhang, Yu Li, and et al. 2022. "Morphological and Molecular Evidence Reveal Eight New Species of Gymnopus from Northeast China" Journal of Fungi 8, no. 4: 349. https://doi.org/10.3390/jof8040349