
How Do Fungi Survive in the Sea and Respond to Climate Change?

 , and
, and
Abstract
:1. Introduction
2. Growth of Terrestrial and Marine Fungi on Seawater Media
3. Effect of Salinity on Spore Germination
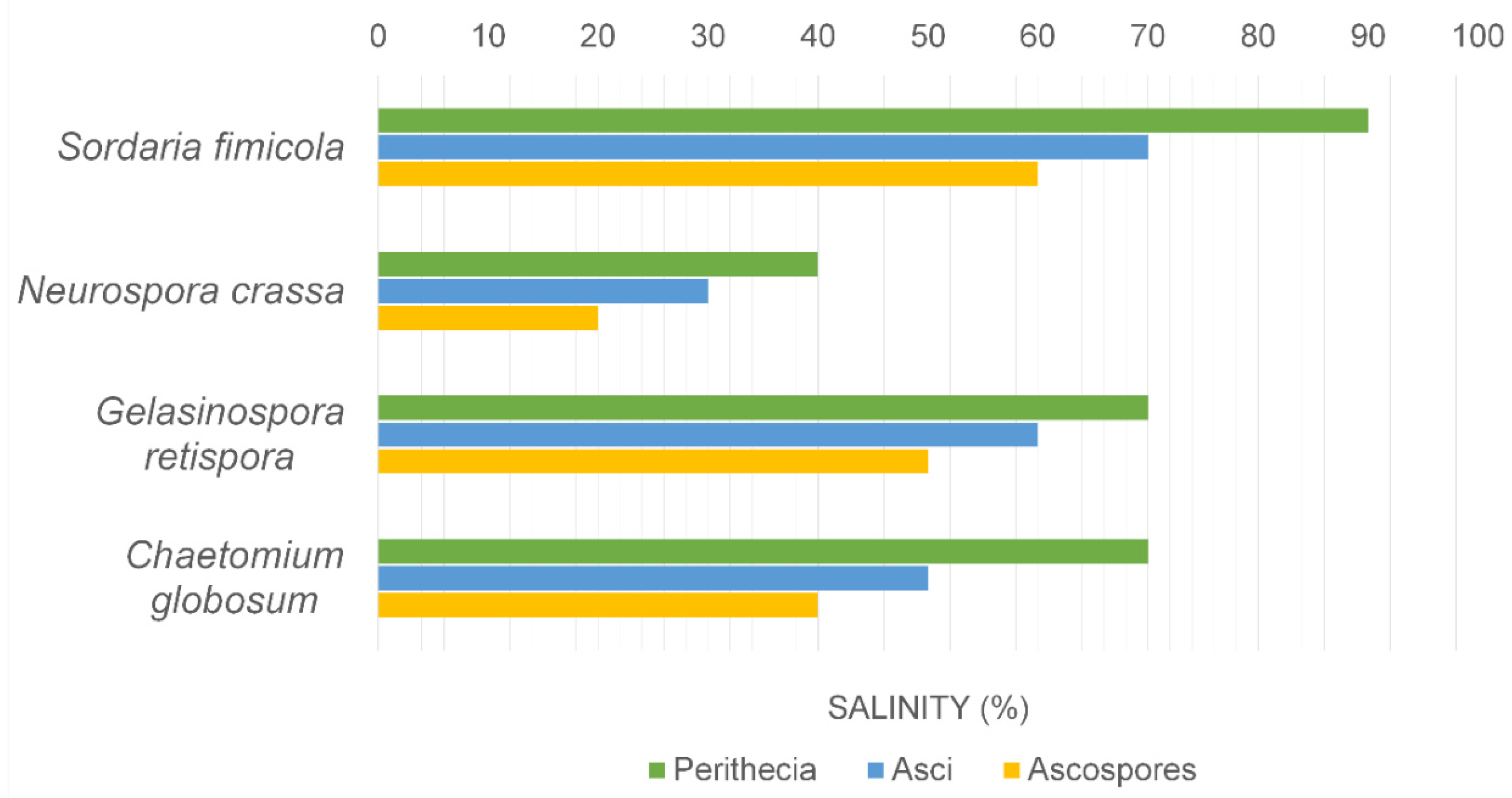
4. Fungal Sporulation with Salinity
5. Fungal-like Organisms and Their Response to Saline Conditions
6. Effect of Temperature and pH on Tolerance to Saline Conditions
7. Can Fungi Be Trained to Tolerate Saline Conditions?
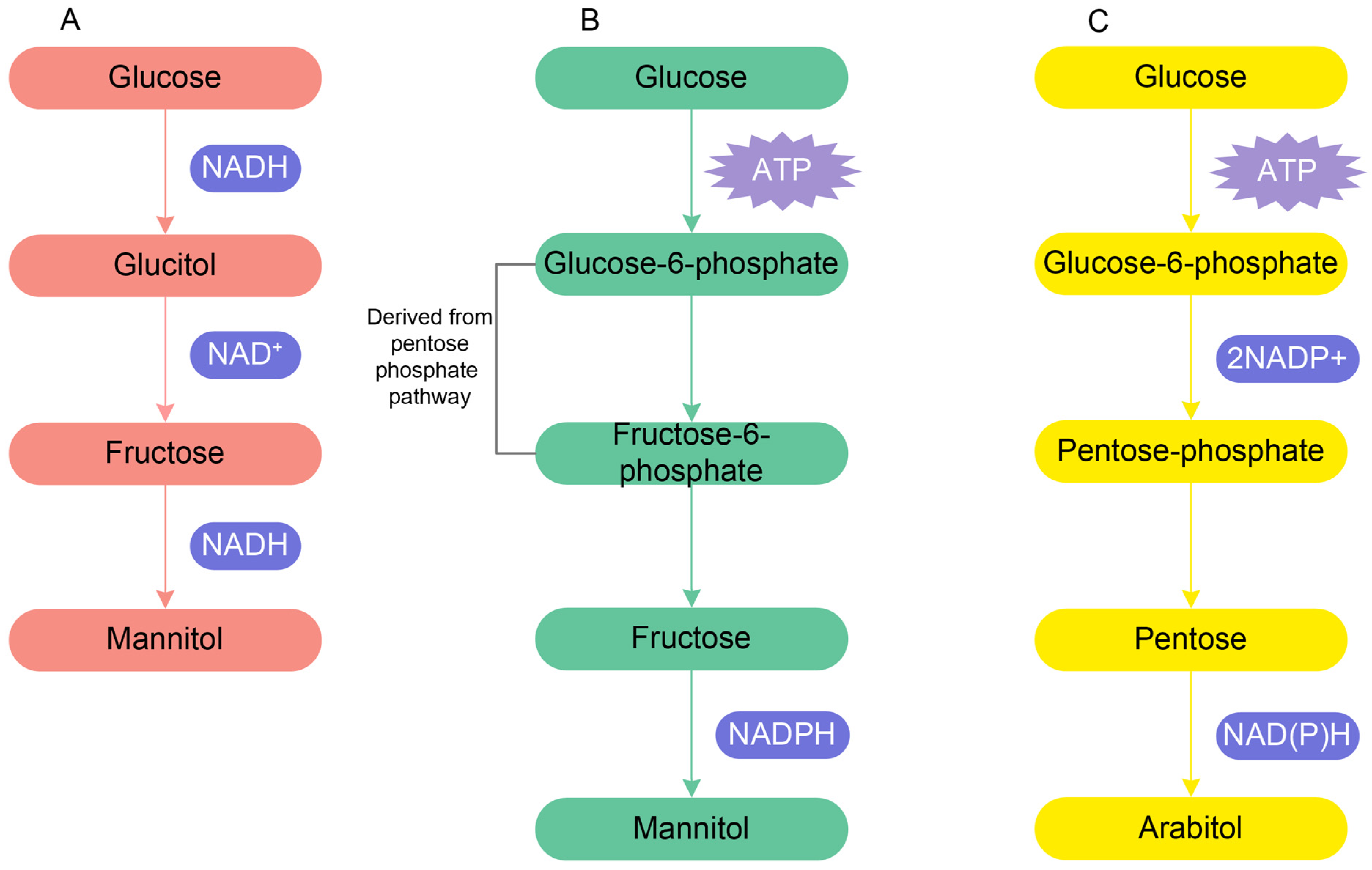
8. Physiological Response to Salinity
9. Salinity Effect on Production of Bioactive Compounds and Other Products
10. Are There Genes That Control the Ability of Fungi to Survive in the Sea?
11. Ecological Occurrence of Marine Fungi
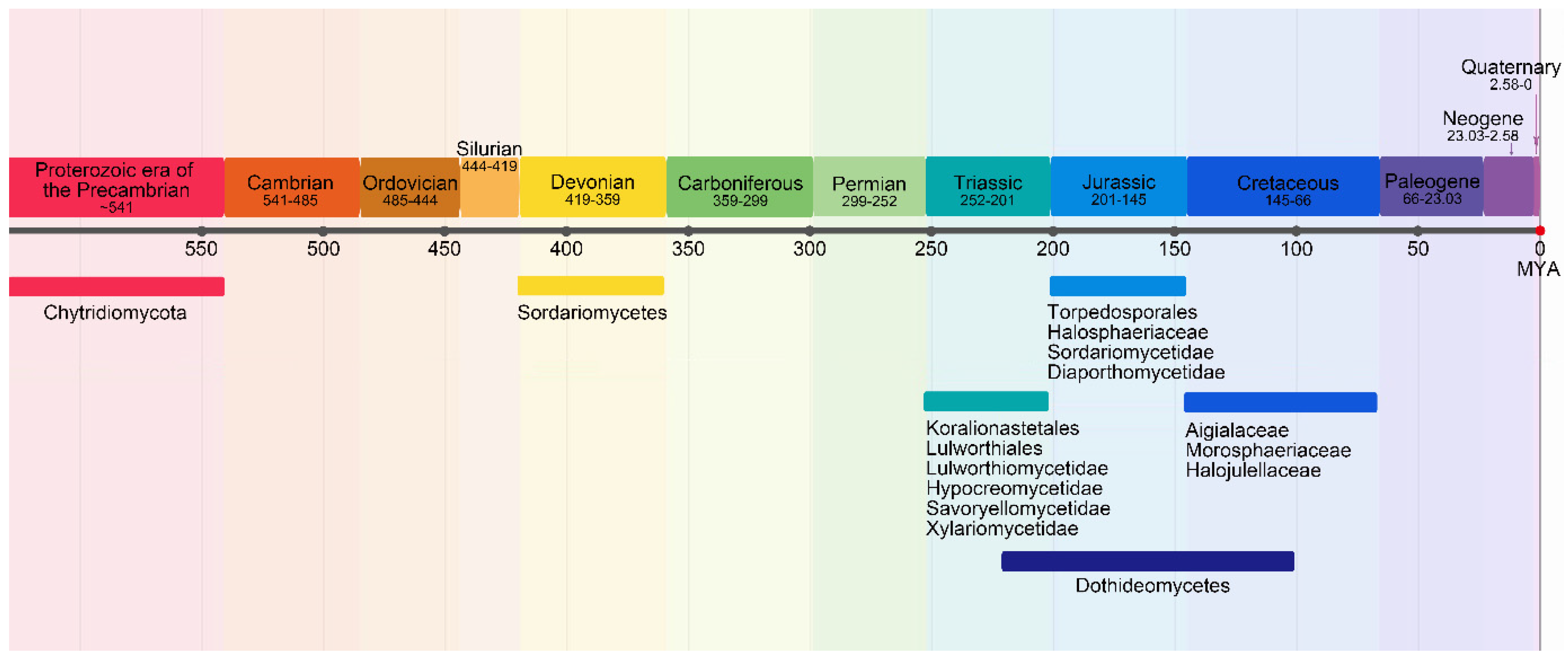
12. Did Marine Fungi Evolve in the Sea?
13. Marine Fungi and Climate Change
14. Unresolved Issues: Endophytes
15. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jones, E.B.G.; Pang, K.L.; Abdel-Wahab, M.A.; Scholz, B.; Hyde, K.D.; Boekhout, T.; Ebel, R.; Rateb, M.E.; Henderson, L.; Sakayaroj, J. An online resource for marine fungi. Fungal Divers. 2019, 96, 347–433. [Google Scholar] [CrossRef]
- Pang, K.L.; Hassett, B.T.; Shaumi, A.; Guo, S.Y.; Sakayaroj, J.; Chiang, M.W.L.; Yang, C.H.; Jones, E.B.G. Pathogenic fungi of marine animals: A taxonomic perspective. Fungal Biol. Rev. 2021, 38, 92–106. [Google Scholar] [CrossRef]
- Devadatha, B.; Jones, E.B.G.; Pang, K.L.; Abdel-Wahab, M.A.; Hyde, K.D.; Sakayaroj, J.; Bahkali, A.H.; Calabon, M.S.; Sarma, V.V.; Suetrong, S.; et al. Occurrence and geographical distribution of mangrove fungi. Fungal Divers. 2021, 106, 137–227. [Google Scholar] [CrossRef]
- Calabon, M.S.; Jones, E.B.G.; Promputtha, I.; Hyde, K.D. Fungal biodiversity in salt marsh ecosystems. J. Fungi 2021, 7, 648. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Suetrong, S.; Sakayaroj, J.; Bahkali, A.H.; Abdel-Wahab, M.A.; Boekhout, T.; Pang, K.-L. Classification of marine Ascomycota, Basidiomycota, Blastocladiomycota and Chytridiomycota. Fungal Divers. 2015, 73, 1–72. [Google Scholar] [CrossRef]
- Jones, E.B.G. Marine fungi: Some factors influencing biodiversity. Fungal Divers. 2000, 4, 53–73. [Google Scholar]
- Burgaud, G.; Hué, N.T.M.; Arzur, D.; Coton, M.; Perrier-Cornet, J.-M.; Jebbar, M.; Barbier, G. Effects of hydrostatic pressure on yeasts isolated from deep-sea hydrothermal vents. Res. Microbiol. 2015, 166, 700–709. [Google Scholar] [CrossRef]
- Zain Ul Arifeen, M.; Chu, C.; Yang, X.; Liu, J.; Huang, X.; Ma, Y.; Liu, X.; Xue, Y.; Liu, C. The anaerobic survival mechanism of Schizophyllum commune 20R-7-F01, isolated from deep sediment 2 km below the seafloor. Environ. Microbiol. 2021, 23, 1174–1185. [Google Scholar] [CrossRef]
- Cai, M.; Fang, Z.; Niu, C.; Zhou, X.; Zhang, Y. Light regulation on growth, development, and secondary metabolism of marine-derived filamentous fungi. Folia Microbiol. 2013, 58, 537–546. [Google Scholar] [CrossRef]
- Bovio, E.; Gnavi, G.; Prigione, V.; Spina, F.; Denaro, R.; Yakimov, M.; Calogero, R.; Crisafi, F.; Varese, G.C. The culturable mycobiota of a Mediterranean marine site after an oil spill: Isolation, identification and potential application in bioremediation. Sci. Total Environ. 2017, 576, 310–318. [Google Scholar] [CrossRef]
- Dell’Anno, F.; Rastelli, E.; Sansone, C.; Brunet, C.; Ianora, A.; Dell’Anno, A. Bacteria, fungi and microalgae for the bioremediation of marine sediments contaminated by petroleum hydrocarbons in the omics era. Microorganisms 2021, 9, 1695. [Google Scholar] [CrossRef]
- De Tender, C.; Devriese, L.I.; Haegeman, A.; Maes, S.; Vangeyte, J.r.; Cattrijsse, A.; Dawyndt, P.; Ruttink, T. Temporal dynamics of bacterial and fungal colonization on plastic debris in the North Sea. Environ. Sci. Technol. 2017, 51, 7350–7360. [Google Scholar] [CrossRef] [PubMed]
- Lacerda, A.L.d.F.; Proietti, M.C.; Secchi, E.R.; Taylor, J.D. Diverse groups of fungi are associated with plastics in the surface waters of the Western South Atlantic and the Antarctic Peninsula. Mol. Ecol. 2020, 29, 1903–1918. [Google Scholar] [CrossRef] [PubMed]
- Barghoorn, E.S.; Linder, D.H. Marine fungi: Their taxonomy and biology. Farlowia 1944, 1, 395–467. [Google Scholar] [CrossRef]
- Pang, K.L.; Overy, D.P.; Jones, E.B.G.; Calado, M.D.; Burgaud, G.; Walker, A.K.; Johnson, J.A.; Kerr, R.G.; Cha, H.J.; Bills, G.F. ‘Marine fungi’ and ‘marine-derived fungi’ in natural product chemistry research: Toward a new consensual definition. Fungal Biol. Rev. 2016, 30, 163–175. [Google Scholar] [CrossRef]
- Trisuwan, K.; Rukachaisirikul, V.; Sukpondma, Y.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. Lactone derivatives from the marine-derived fungus Penicillium sp. PSU-F44. Chem. Pharm. Bull. 2009, 57, 1100–1102. [Google Scholar] [CrossRef] [Green Version]
- Chaeprasert, S.; Piapukiew, J.; Whalley, A.J.; Sihanonth, P. Endophytic fungi from mangrove plant species of Thailand: Their antimicrobial and anticancer potentials. Bot. Mar. 2010, 53, 555–564. [Google Scholar] [CrossRef]
- Vargas-Gastélum, L.; Riquelme, M. The mycobiota of the deep sea: What omics can offer. Life 2020, 10, 292. [Google Scholar] [CrossRef]
- Park, D. On the ecology of heterotrophic microorganisms in freshwater. Trans. Br. Mycol. Soc. 1972, 58, 291–299. [Google Scholar] [CrossRef]
- Velez, P. Impact of salinity stress on growth and development of aquatic fungi. In Microorganisms in Saline Environments: Strategies and Functions; Giri, B., Varma, A., Eds.; Springer: Cham, Germany, 2019; pp. 155–168. [Google Scholar]
- Ritchie, D. Salinity optima for marine fungi affected by temperature. Am. J. Bot. 1957, 44, 870–874. [Google Scholar] [CrossRef]
- Gray, W.D.; Pinto, P.V.C.; Pathak, S.G. Growth of fungi in sea water medium. Appl. Microbiol. 1963, 11, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.W., Jr. Morphogenesis and microscopic cytochemistry of marine pyrenomycete ascospores. Nova Hedw. Beih. 1966, 22, 1–128. [Google Scholar]
- Jones, E.B.G.; Jennings, D.H. The effect of salinity on the growth of marine fungi in comparison with non-marine species. Trans. Br. Mycol. Soc. 1964, 47, 619–625. [Google Scholar] [CrossRef]
- Meyers, S.P.; Simms, J. Thalassiomycetes VI. Comparative growth studies of Lindra thalassiae and lignicolous Ascomycete species. Can. J. Bot. 1965, 43, 379–392. [Google Scholar] [CrossRef]
- Meyers, S.P.; Hoyo, L. Observations on the growth of the marine Hyphomycete Varicosporina ramulosa. Can. J. Bot. 1966, 44, 1133–1140. [Google Scholar] [CrossRef]
- Meyers, S.P.; Scott, E.; Thalassiomycetes, X. Variation in growth and reproduction of two isolates of Corollospora maritima. Mycologia 1967, 59, 446–455. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Byrne, P.J.; Alderman, D.J. The response of fungi to salinity. Vie Milieu Suppl. 1971, 22, 265–280. [Google Scholar]
- Davidson, D.E. The effect of salinity on a marine and a freshwater Ascomycete. Can. J. Bot. 1974, 52, 553–563. [Google Scholar] [CrossRef]
- Byrne, P.J.; Jones, E.B.G. Effect of salinity on spore germination of terrestrial and marine fungi. Trans. Brit. Mycol. Soc. 1975, 64, 497–503. [Google Scholar] [CrossRef]
- Byrne, P.J.; Jones, E.B.G. Effect of salinity on the reproduction of terrestrial and marine fungi. Trans. Brit. Mycol. Soc. 1975, 65, 185–200. [Google Scholar] [CrossRef]
- Gessner, R.V. In vitro growth and nutrition of Buergenerula spartinae, a fungus associated with Spartina alterniflora. Mycologia 1976, 68, 583–599. [Google Scholar] [CrossRef]
- Curran, P. The effect of temperature, pH, light and dark on the growth of fungi from Irish coastal waters. Mycologia 1980, 72, 350–358. [Google Scholar] [CrossRef]
- Crabtree, S.L.; Gessner, R.V. Growth and nutrition of the salt marsh fungi Pleospora gaudifroyi and Camarosporium roumeguerii. Mycologia 1982, 74, 640–647. [Google Scholar] [CrossRef]
- Richards, T.A.; Leonard, G.; Mahé, F.; del Campo, J.; Romac, S.; Jones, M.D.M.; Maguire, F.; Dunthorn, M.; De Vargas, C.; Massana, R.; et al. Molecular diversity and distribution of marine fungi across 130 European environmental samples. Proc. R. Soc. B 2015, 282, 20152243. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.B.G.; Jennings, D.H. The effect of cations on the growth of fungi. New Phytol. 1965, 64, 86–100. [Google Scholar] [CrossRef]
- Amon, J.P. An estuarine species of Phlyctochytrium (Chytridiales) having a transient requirement for sodium. Mycologia 1976, 68, 470–480. [Google Scholar] [CrossRef]
- Amon, J.P.; Arthur, R.D. Nutritional studies of a marine Phlyctochytrium sp. Mycologia 1981, 73, 1049–1055. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Ward, A.W. Septate conidia in Asteromyces cruciatus. Trans. Brit. Mycol. Soc. 1973, 61, 181–186. [Google Scholar] [CrossRef]
- Nakagiri, A.; Tubaki, K. Ascocarp peridial wall structure in Corollospora and allied genera of Halosphaeriaceae. In The Biology of Marine Fungi; Moss, S.T., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 245–251. [Google Scholar]
- Lintott, W.H.; Lintott, E.A. Marine fungi from New Zealand. In Fungi in Marine Environments; Hyde, K.D., Ed.; Fungal Diversity Research Series: Hong Kong, China, 2002; pp. 285–292. [Google Scholar]
- Velez, P.; Alejandri-Ramírez, N.D.; González, M.C.; Estrada, K.J.; Sanchez-Flores, A.; Dinkova, T.D. Comparative transcriptome analysis of the cosmopolitan marine fungus Corollospora maritima under two physiological conditions. G3 Genes Genomes Genet. 2015, 5, 1805–1814. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.B.G.; Oliver, A.C. Occurrence of aquatic hyphomycetes on wood submerged in fresh and brackish water. Trans. Brit. Mycol. Soc. 1964, 47, 45–48. [Google Scholar] [CrossRef]
- Doguet, G. Nia vibrissa Moore et Meyers, Gastéromycète marin. I. Conditions générales de formation des carpophores en culture. Bull. Soc. Mycol. Fr. 1968, 84, 343–351. [Google Scholar]
- Borut, S.Y.; Johnson, T.W., Jr. Some biological observations on fungi in estuarine sediments. Mycologia 1962, 54, 181–193. [Google Scholar] [CrossRef]
- Jennings, D.H. Fungal growth in the sea. In The Biology of Marine Fungi; Moss, S.T., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 1–10. [Google Scholar]
- Galpin, M.F.J.; Jennings, D.H.; Oates, K.; Hobot, J.A. Localisation by X-ray microanalysis of soluble ions, particularly potassium and sodium in fungal hyphae. Exp. Mycol. 1978, 2, 258–269. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Byrne, P.J. Physiology of the higher marine fungi. In Recent Advances in Aquatic Mycology; Jones, E.B.G., Ed.; Wiley: New York, NY, USA, 1976; pp. 135–175. [Google Scholar]
- Jones, E.B.G.; Harrison, J.L. Physiology of marine Phycomycetes. In Recent Advances in Aquatic Mycology; Jones, E.B.G., Ed.; Wiley: New York, NY, USA, 1976; pp. 261–278. [Google Scholar]
- Leaño, E.M.; Vrijmoed, L.L.P.; Jones, E.B.G. Physiological studies on Halophytophthora vesicula (straminipilous fungi) isolated from fallen mangrove leaves from Mai Po, Hong Kong. Bot. Mar. 1998, 41, 411–419. [Google Scholar] [CrossRef]
- Leaño, E.M.; Jones, E.B.G.; Vrijmoed, L.L.P. Why are Halophytophthora species well adapted to mangrove habitats? Fungal Divers. 2000, 5, 131–151. [Google Scholar]
- Su, C.J.; Hsieh, S.Y.; Chiang, M.W.L.; Pang, K.L. Salinity, pH and temperature growth ranges of Halophytophthora isolates suggest their physiological adaptations to mangrove environments. Mycology 2020, 11, 256–262. [Google Scholar] [CrossRef] [Green Version]
- Vishniac, H.S. A new marine Phycomycete. Mycologia 1958, 50, 66–79. [Google Scholar] [CrossRef]
- Goldstein, S. Development and nutrition of new species of Thraustochytrium. Am. J. Bot. 1963, 50, 271–279. [Google Scholar] [CrossRef]
- Goldstein, S.; Belsky, M. Axenic culture studies of a new marine phycomycete possessing an unusual type of asexual reproduction. Am. J. Bot. 1964, 51, 72–78. [Google Scholar] [CrossRef]
- Johnson, T.W.; Sparrow, F.K. Fungi in Oceans and Estuaries; Cramer: Weinheim, Germany, 1961; p. 668. [Google Scholar]
- Harrison, J.L.; Jones, E.B.G. Effect of salinity on sexual and asexual sporulation of members of the Saprolegniaceae. Trans. Br. Mycol. Soc. 1975, 65, 389–394. [Google Scholar] [CrossRef]
- Höhnk, W. Ein Beitrag zur Kenntnis der Phycomyceten des Brackwassers. Kiel. Meeresforsch. 1939, 3, 337–361. [Google Scholar]
- Höhnk, W. Eine neue uferbewohnende Saprolegniazee: Calyptralegnia ripariensis nov. spec. Veröffentlichungen Des Inst. Für Meeresforsch. Bremerhav. 1953, 2, 230–235. [Google Scholar]
- Lorenz, R.; Molitoris, H.P. Combined influence of salinity and temperature (Phoma-pattern) on growth of marine fungi. Can. J. Bot. 1992, 70, 2111–2115. [Google Scholar] [CrossRef]
- Torzilli, A.P.; Vinroot, S.; West, C. Interactive effect of temperature and salinity on growth and activity of a salt marsh isolate of Aureobasidium pullulans. Mycologia 1985, 77, 278–284. [Google Scholar] [CrossRef]
- Pang, K.L.; Chow, R.K.K.; Chan, C.W.; Vrijmoed, L.L.P. Diversity and physiology of marine lignicolous fungi in Arctic waters: A preliminary account. Polar Res. 2011, 30, 5859. [Google Scholar] [CrossRef]
- Pang, K.L.; Chiang, M.W.; Guo, S.Y.; Shih, C.Y.; Dahms, H.U.; Hwang, J.S.; Cha, H.J. Growth study under combined effects of temperature, pH and salinity and transcriptome analysis revealed adaptations of Aspergillus terreus NTOU4989 to the extreme conditions at Kueishan Island Hydrothermal Vent Field, Taiwan. PLoS ONE 2020, 15, e0233621. [Google Scholar] [CrossRef] [PubMed]
- Park, J.C.; Nemoto, Y.; Homma, T.; Jing, W.; Chen, Y.; Matsuoka, H.; Ohno, H.; Takatori, K.; Kurata, H. Adaptation of Aspergillus niger to short-term salt stress. Appl. Microbiol. Biotechnol. 1993, 40, 394–398. [Google Scholar] [CrossRef]
- Sampangi-Ramaiah, M.H.; Jagadheesh; Dey, P.; Jambagi, S.; Vasantha Kumari, M.M.; Oelmüller, R.; Nataraja, K.N.; Venkataramana Ravishankar, K.; Ravikanth, G.; Uma Shaanker, R. An endophyte from salt-adapted Pokkali rice confers salt-tolerance to a salt-sensitive rice variety and targets a unique pattern of genes in its new host. Sci. Rep. 2020, 10, 3237. [Google Scholar] [CrossRef]
- Jennings, D.H. Cations and filamentous fungi: Invasion of the sea and hyphal functioning. In Ion Transport in Plants; Anderson, W.P., Ed.; Academic Press: London, UK, 1973; pp. 323–335. [Google Scholar]
- Jennings, D.H. Some aspects of the physiology and biochemistry of marine fungi. Biol. Rev. 1983, 58, 423–459. [Google Scholar] [CrossRef]
- Jennings, D.H. Polyol metabolism in fungi. Adv. Microb. Physiol. 1985, 25, 149–193. [Google Scholar]
- Holligan, P.M.; Jennings, D.H. Carbohydrate metabolism in the fungus Dendryphiella salina. I. Changes in the Levels of Soluble Carbohydrates during Growth. New Phytol. 1972, 71, 569–582. [Google Scholar] [CrossRef]
- Holligan, P.M.; Jennings, D.H. Carbohydrate metabolism in the fungus Dendryphiella salina. II. The influence of different carbon and nitrogen sources on the accumulation of mannitol and arabitol. New Phytol. 1972, 71, 583–594. [Google Scholar] [CrossRef]
- Holligan, P.M.; Jennings, D.H. Carbohydrate metabolism in the fungus Dendryphiella salina. III. The effect of the nitrogen source on the metabolism of [I-14C]-and [6-14C]-glucose. New Phytol. 1972, 71, 1119–1133. [Google Scholar] [CrossRef]
- Jennings, D.H.; Austin, S. The stimulatory effect of the non-metabolisable sugar 3-O-methyl glucose on the conversion of mannitol and arabitol to polysaccharide and other insoluble compounds in the fungus Dendryphiella salina. J. Gen. Microbiol. 1973, 75, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Lowe, D.A.; Jennings, D.H. Carbohydrate metabolism in the fungus Dendryphiella salina. V. The pattern of label in arabitol and polysaccharide after growth in the presence of specifically labelled carbon sources. New Phytol. 1975, 74, 67–79. [Google Scholar] [CrossRef]
- Galpin, M.F.J.; Jennings, D.H. Histochemical study of the hyphae and the distribution of adenosine triphosphatase in Dendryphiella salina. Trans. Brit. Mycol. Soc. 1975, 65, 477–483. [Google Scholar] [CrossRef]
- Wethered, J.M.; Metcalf, E.C.; Jennings, D.H. Carbohydrate metabolism in the fungus Dendryphiella salina. VIII. The contribution of polyols and ions to the mycelial solute potential in relation to the external osmoticum. New Phytol. 1985, 101, 631–649. [Google Scholar] [CrossRef]
- Galpin, M.F.J.; Jennings, D.H. A plasma-membrane ATPase from Dendryphiella salina: Cation specificity and interaction with fusicoccin and cyclic AMP. Trans. Br. Mycol. Soc. 1980, 75, 35–46. [Google Scholar] [CrossRef]
- Siegenthaler, P.A.; Belsky, M.M.; Goldstein, S. Phosphate uptake in an obligately marine fungus: A specific requirement for sodium. Science 1967, 155, 93–94. [Google Scholar] [CrossRef]
- Siegenthaler, P.A.; Belsky, M.M.; Goldstein, S.; Menna, M. Phosphate uptake in an obligately marine fungus. II. Role of culture conditions, energy sources and inhibitors. J. Bacteriol. 1967, 93, 1281–1288. [Google Scholar] [CrossRef] [Green Version]
- Wethered, J.M.; Jennings, D.H. The major solutes contributing to the solute potential of Thraustochytrium aureum and T. roseum after growth in media of different salinities. Trans. Br. Myol. Soc. 1985, 85, 439–446. [Google Scholar] [CrossRef]
- Norkrans, B. Studies on marine occurring yeasts. Growth related to pH, NaCl concentrations and temperature. Arch. Mikrobiol. 1966, 54, 374–392. [Google Scholar] [CrossRef]
- Norkrans, B.; Kylin, A. Regulation of potassium to sodium and of the osmotic potential in relation to salt tolerance in yeasts. J. Bacteriol. 1969, 100, 836–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustafsson, L.; Norkrans, B. On the mechanism of salt tolerance. Production of glycerol and heat during growth of Debaryomyces hansenii. Arch. Microbiol. 1976, 110, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Adler, L.; Gustafsson, L. Polyhydric alcohol production and intracellular amino acid pool in relation to halotolerance of the yeast Debaryomyces hansenii. Arch. Microbiol. 1980, 124, 123–130. [Google Scholar] [CrossRef]
- Adler, L. Physiological and biochemical characteristics of the yeast Debaryomyces hansenii in relation to salinity. In The Biology of Marine Fungi; Moss, S.T., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 81–89. [Google Scholar]
- Larsen, H. Halophilic and halotolerant microorganisms- an overview and historical perspective. FEMS Microbiol. Rev. 1986, 39, 3–7. [Google Scholar] [CrossRef]
- Kis-Papo, T.; Oren, A.; Wasser, S.P.; Nevo, E. Survival of filamentous fungi in hypersaline Dead Sea water. Microb. Ecol. 2003, 45, 183–190. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Ramos, J.; Plemenitas, A. Halotolerant and halophilic fungi. Mycol. Res. 2009, 113, 1231–1241. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Butinar, L.; Sonjak, S.; Turk, M.; Uršič, V.; Zalar, P.; Plemenitaš, A. halotolerant and halophilic fungi from coastal environments in the Arctics. In Adaptation to Life at High Salt Concentrations in Archaea, Bacteria, and Eukarya (Cellular Origin, Life in Extreme Habitats and Astrobiology); Gunde-Cimerman, N., Oren, A., Plemenitaš, A., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 397–423. [Google Scholar]
- Zajc, J.; Zalar, P.; Plemenitaš, A.; Gunde-Cimerman, N. The mycobiota of the salterns. In Biology of Marine Fungi; Raghukumar, C., Ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 2012; pp. 133–158. [Google Scholar]
- Abbanat, D.; Leighton, M.; Maiese, W.; Jones, E.B.G.; Pearce, C.; Greenstein, M. Cell wall active antifungal compounds produced by the marine fungus Hypoxylon oceanicum LL-15G256 I. Taxonomy and fermentation. J. Antibiot. 1998, 51, 296–302. [Google Scholar] [CrossRef]
- Bugni, T.S.; Ireland, C.M. Marine-derived fungi: A chemically and biologically diverse group of microorganisms. Nat. Prod. Rep. 2004, 21, 143–163. [Google Scholar] [CrossRef]
- Miller, J.D.; Savard, M.E. Antibiotic activity of the marine fungus Leptosphaeria oraemaris. Proc. Nova Scotia Inst. Sci. 1989, 39, 51–58. [Google Scholar]
- Masuma, R.; Yamaguchi, Y.; Noumi, M.; Omura, S.; Namikoshi, M. Effect of sea water concentration on hyphal growth and antimicrobial metabolite production in marine fungi. Mycoscience 2001, 42, 455–459. [Google Scholar] [CrossRef]
- Chinworrungsee, M.; Kittakoop, P.; Isaka, M.; Rungrod, A.; Tanticharoen, M.; Thebtaranonth, Y. Antimalarial halorosellinic acid from the marine fungus Halorosellinia oceanica. Bioorg. Med. Chem. Lett. 2001, 11, 196–199. [Google Scholar] [CrossRef]
- Vongvilai, P.; Isaka, M.; Kittakoop, P.; Srikitikulchai, P.; Kongsaeree, P.; Thebtaranonth, Y. Ketene acetal and spiroacetal constituents of the marine fungus Aigialus parvus BCC 5311. J. Nat. Prod. 2004, 67, 457–460. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Wu, X.Y.; Feng, S.; Jiang, G.C.; Luo, J.H.; Zhou, S.N.; Vrijmoed, L.L.P.; Jones, E.B.G.; Krohn, K.; Steingröver, K.; et al. Five unique compounds: Xyloketals from mangrove fungus Xylaria sp. from the South China Sea coast. J. Org. Chem. 2001, 66, 6252–6256. [Google Scholar] [CrossRef] [PubMed]
- Toske, S.G.; Jensen, P.R.; Kauffman, C.A.; Fenical, W. Aspergillamides A and B: Modified cytotoxic tripeptides produced by a marine fungus of the genus Aspergillus. Tetrahedron 1998, 54, 13459–13466. [Google Scholar] [CrossRef]
- Janso, J.E.; Bernan, V.S.; Greenstein, M.; Bugni, T.S.; Ireland, C.M. Penicillium dravuni, a new marine-derived species from an alga in Fiji. Mycologia 2005, 97, 444–453. [Google Scholar] [CrossRef]
- Tepšič, K.; Gunde-Cimerman, N.; Frisvad, J.C. Growth and mycotoxin production by Aspergillus fumigatus strains isolated from a saltern. FEMS Microbiol. Lett. 1997, 157, 9–12. [Google Scholar] [CrossRef]
- Overy, D.; Correa, H.; Roullier, C.; Chi, W.C.; Pang, K.L.; Rateb, M.; Ebel, R.; Shang, Z.; Capon, R.; Bills, G. Does osmotic stress affect natural product expression in fungi? Mar. Drugs 2017, 15, 254. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, M.F.M.; Vicente, T.F.L.; Esteves, A.C.; Alves, A. Novel halotolerant species of Emericellopsis and Parasarocladium associated with macroalgae in an estuarine environment. Mycologia 2021, 112, 154–171. [Google Scholar] [CrossRef]
- Aggarwal, M.; Bansal, P.K.; Mondal, A.K. Molecular cloning and biochemical characterization of a 3’(2’), 5’-bisphosphate nucleotidase from Debaryomyces hansenii. Yeast 2005, 22, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Prista, C.; Gonzalez-Hernandez, J.C.; Ramos, J.; Loureiro-Dias, M.C. Cloning and characterization of two K+ transporters of Debaryomyces hansenii. Microbiology 2007, 153, 3034–3043. [Google Scholar] [CrossRef] [Green Version]
- Hagestad, O.C.; Hou, L.; Andersen, J.H.; Hansen, E.H.; Altermark, B.; Li, C.; Kuhnert, E.; Cox, R.J.; Crous, P.W.; Spatafora, J.W.; et al. Genomic characterization of three marine fungi, including Emericellopsis atlantica sp. nov. with signatures of a generalist lifestyle and marine biomass degradation. IMA Fungus 2021, 12, 21. [Google Scholar] [CrossRef] [PubMed]
- Booth, T.; Kenkel, N. Ecological studies of lignicolous marine fungi: A distribution model based on ordination and classification. In The Biology of Marine Fungi; Moss, S.T., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 297–310. [Google Scholar]
- Hughes, G.C. Biogeography and the marine fungi. In The Biology of Marine Fungi; Moss, S.T., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 275–295. [Google Scholar]
- Hughes, G.C. Geographical distribution of the higher marine fungi. Veroff. Inst. Meeresforsch. Bremerh. Suppl. 1974, 5, 419–441. [Google Scholar]
- Hughes, G.C. Studies of fungi in oceans and estuaries since 1961. I. Lignicolous, caulicolous and foliicolous species. Oceanogr. Mar. Biol. A Rev. 1975, 13, 69–180. [Google Scholar]
- Pugh, G.J.F.; Jones, E.B.G. Antarctic marine fungi: A preliminary account. In The Biology of Marine Fungi; Moss, S.T., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 323–330. [Google Scholar]
- Wilson, I. Some new marine Pyrenomycetes on wood or rope: Halophiobolus and Lindra. Trans. Brit. Mycol. Soc. 1956, 39, 401–415. [Google Scholar] [CrossRef]
- Hyde, K.D. Frequency of occurrence of lignicolous marine fungi in the tropics. In The Biology of Marine Fungi; Moss, S.T., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 311–322. [Google Scholar]
- Sridhar, K.R.; Alias, S.A.; Pang, K.L. Mangrove fungi. In Marine Fungi and Fungus-like Organisms; Jones, E.B.G., Pang, K.L., Eds.; De Gruyter: Berlin, Germany, 2012; pp. 253–271. [Google Scholar]
- Maria, G.L.; Sridhar, K.R. Diversity of filamentous fungi on woody litter of five mangrove plant species from the southwest coast of India. Fungal Divers. 2003, 14, 109–126. [Google Scholar]
- Shearer, C.A. Fungi of the Chesapeake Bay and its tributaries. III. The distribution of wood-inhabiting ascomycetes and fungi imperfecti of the Patuxent River. Am. J. Bot. 1972, 59, 961–969. [Google Scholar] [CrossRef]
- Byrne, P.J.; Jones, E.B.G. Lignicolous marine fungi. Veroeff. Inst. Meeresforsch. Bremerhav. Suppl. 1974, 5, 301–320. [Google Scholar]
- Fryar, S.C.; Booth, W.; Davies, J.; Hodgkiss, I.J.; Hyde, K.D. Distribution of fungi on wood in the Tutong River, Brunei. Fungal Divers. 2004, 17, 17–38. [Google Scholar]
- Rojas-Jimenez, K.; Rieck, A.; Wurzbacher, C.; Jurgens, K.; Labrenz, M.; Grossart, H.P. A salinity threshold separating fungal communities in the Baltic Sea. Front. Microbiol. 2019, 10, 680. [Google Scholar] [CrossRef]
- Heckman, D.S.; Geiser, D.M.; Eidell, B.R.; Stauffer, R.L.; Kardos, N.L.; Hedges, S.B. Molecular evidence for the early colonization of land by fungi and plants. Science 2001, 293, 1129–1133. [Google Scholar] [CrossRef] [Green Version]
- Le Calvez, T.; Burgaud, G.; Mahé, S.; Barbier, G.; Vandenkoornhuyse, P. Fungal diversity in deep sea hydrothermal ecosystems. Appl. Environ. Microbiol. 2009, 75, 6415–6421. [Google Scholar] [CrossRef] [Green Version]
- James, T.; Kauff, F.; Schoch, C.L.; Matheny, P.B.; Hofstetter, V.; Cox, C.J.; Celio, G.; Gueidan, C.; Fraker, E.; Miadlikowska, J.; et al. Reconstructing the early evolution of Fungi using a six-gene phylogeny. Nature 2006, 443, 818–822. [Google Scholar] [CrossRef]
- Lücking, R.; Huhndorf, S.; Pfister, D.H.; Plata, E.R.; Lumbsch, H.T. Fungi evolved right on track. Mycologia 2009, 101, 810–822. [Google Scholar] [CrossRef] [Green Version]
- Raghukumar, S. Origin and Evolution of Marine Fungi. In Fungi in Coastal and Oceanic Marine Ecosystems; Raghukumar, S., Ed.; Springer International Publishing AG: Cham, Germany, 2017; pp. 307–321. [Google Scholar]
- Shearer, C.A. The freshwater ascomycetes. Nova Hedw. 1993, 56, 1–33. [Google Scholar]
- Vijaykrishna, D.; Jeewon, R.; Hyde, K.D. Molecular taxonomy, origins and evolution of freshwater ascomycetes. Fungal Divers. 2006, 23, 367–406. [Google Scholar]
- Spatafora, J.W.; Volkmann-Kohlmeyer, B.; Kohlmeyer, J. Independent terrestrial origins of the Halosphaeriales (marine Ascomycota). Am. J. Bot. 1998, 85, 1569–1580. [Google Scholar] [CrossRef]
- Campbell, J.; Anderson, J.L.; Shearer, C.A. Systematics of Halosarpheia based on morphological and molecular data. Mycologia 2003, 95, 530–552. [Google Scholar] [CrossRef]
- Hibbett, D.S.; Binder, M. Evolution of marine mushrooms. Biol. Bull. 2001, 201, 319–322. [Google Scholar] [CrossRef] [Green Version]
- Binder, M.; Hibbett, D.S.; Wang, Z.; Farnham, W.F. Evolutionary relationships of Mycaureola dilseae (Agaricales), a basidiomycete pathogen of a subtidal rhodophyte. Am. J. Bot. 2006, 93, 547–556. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.B.G.; Choeyklin, R. Ecology of marine and freshwater basidiomycetes. Br. Mycol. Soc. Symp. Ser. 2008, 28, 301–324. [Google Scholar]
- Kumar, V.; Sarma, V.V.; Thambugala, K.M.; Huang, J.J.; Li, X.Y.; Hao, G.F. Ecology and evolution of marine fungi with their adaptation to climate change. Front. Microbiol. 2021, 12, 719000. [Google Scholar] [CrossRef]
- Caldeira, K.; Wickett, M.E. Anthropogenic carbon and ocean pH. Nature 2003, 425, 365. [Google Scholar] [CrossRef]
- Krause, E.; Wichels, A.; Gimenez, L.; Gerdts, G. Marine fungi may benefit from ocean acidification. Aquat. Microb. Ecol. 2013, 69, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Jones, E.B.G. Marine mangrove fungi. Mar. Ecol. 1988, 9, 15–33. [Google Scholar] [CrossRef]
- Schmit, J.P.; Shearer, C.A. A checklist of mangrove associated fungi, their geographical distribution and known host plants. Mycotaxon 2003, 85, 423–477. [Google Scholar]
- Jones, E.B.G.; Pang, K.L. Tropical aquatic fungi. Biodivers. Conserv. 2012, 21, 2403–2423. [Google Scholar] [CrossRef]
- Kingham, D.L.; Evans, L.V. The Pelvetia–Mycosphaerella interrelationship. In The Biology of Marine Fungi; Moss, S.T., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 177–187. [Google Scholar]
- Stanley, S.J. Observations on the seasonal occurrence of marine endophytic and parasitic fungi. Can. J. Bot. 1992, 70, 2089–2096. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Pang, K.L.; Stanley, S.J. Fungi form marine algae. In Marine Fungi and Fungus-Like Organisms; Jones, E.B.G., Pang, K.L., Eds.; De Gruyter: Berlin, Germany, 2012; pp. 329–344. [Google Scholar]
- Panno, L.; Bruno, M.; Voyron, S.; Anastasi, A.; Gnavi, G.; Miserere, L.; Varese, G.C. Diversity, ecological role and potential biotechnological applications of marine fungi associated to the seagrass Posidonia oceanica. New Biotechnol. 2013, 30, 685–694. [Google Scholar] [CrossRef]
- Gnavi, G.; Ercole, E.; Panno, L.; Vizzini, A.; Varese, G.C. Dothideomycetes and Leotiomycetes sterile mycelia isolated from the Italian seagrass Posidonia oceanica based on rDNA data. Springerplus 2014, 3, 508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poli, A.; Bovio, E.; Ranieri, L.; Varese, G.C.; Prigione, V. Fungal diversity in the neptune forest: Comparison of the mycobiota of Posidonia oceanica, Flabellia petiolata, and Padina pavonica. Front. Microbiol. 2020, 11, 933. [Google Scholar] [CrossRef] [PubMed]
- Garzoli, L.; Gnavi, G.; Tamma, F.; Tosi, S.; Varese, G.C.; Picco, A.M. Sink or swim: Updated knowledge on marine fungi associated with wood substrates in the Mediterranean Sea and hints about their potential to remediate hydrocarbons. Progr. Oceanog. 2015, 137, 140–148. [Google Scholar] [CrossRef]
- Cha, H.J.; Chiang, M.W.L.; Guo, S.Y.; Lin, S.M.; Pang, K.L. Culturable fungal community of Pterocladiella capillacea at Keelung, Taiwan: Effects of surface sterilization method and isolation medium. J. Fungi 2021, 7, 651. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Shenoy, B.D.; Li, W.; Cai, L. Molecular phylogeny of Neodevriesia, with two new species and several new combinations. Mycologia 2017, 109, 965–974. [Google Scholar] [CrossRef]
- Bovio, E.; Garzoli, L.; Poli, A.; Prigione, V.; Firsova, D.; McCormack, G.; Varese, G.C. The culturable mycobiota associated with three Atlantic sponges, including two new species: Thelebolus balaustiformis and T. spongiae. Fungal Syst. Evol. 2018, 1, 141–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bovio, E.; Garzoli, L.; Poli, A.; Luganini, A.; Villa, P.; Musumeci, R.; McCormack, G.P.; Cocuzza, C.E.; Gribaudo, G.; Mehiri, M.; et al. Marine fungi from the sponge Grantia compressa: Biodiversity, chemodiversity, and biotechnological potential. Mar. Drugs 2019, 17, 220. [Google Scholar] [CrossRef] [Green Version]
- Marchese, P.; Garzoli, L.; Gnavi, G.; O’Connell, E.; Bouraoui, A.; Mehiri, M.; Murphy, J.M.; Varese, G.C. Diversity and bioactivity of fungi associated with the marine sea cucumber Holothuria poli: Disclosing the strains potential for biomedical applications. J. Appl. Microbiol. 2020, 129, 612–625. [Google Scholar] [CrossRef]
- Davies, C.E.; Malkin, S.H.; Thomas, J.E.; Batista, F.M.; Rowley, A.F.; Coates, C.J. Mycosis is a disease state encountered rarely in shore crabs, Carcinus maenas. Pathogens 2020, 9, 462. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Li, B.; Zheng, C.; Wang, G. Molecular detection of fungal communities in the Hawaiian marine sponges Suberites zeteki and Mycale armata. Appl. Environ. Microbiol. 2008, 74, 6091–6101. [Google Scholar] [CrossRef] [Green Version]
- Hurtado-McCormick, V.; Kahlke, T.; Petrou, K.; Jeffries, T.; Ralph, P.; Seymour, J. Regional and microenvironmental scale characterization of the seagrass microbiome. Front. Microbiol. 2019, 10, 1011. [Google Scholar] [CrossRef]
- Nguyen, M.T.; Thomas, T. Diversity, host-specificity and stability of sponge-associated fungal communities of co-occurring sponges. PeerJ 2018, 6, e4965. [Google Scholar] [CrossRef] [Green Version]
- Stanley, S.J. The Autecology and Ultrastructure Interaction between Mycosphaerella ascophylli Cotton, Lautitia danica (Berlese) Schatz, Mycaureola dilsea Maire et Chemin and Their Respective Marine Algal Hosts. Ph.D. Thesis, University of Portsmouth, Portsmouth, UK, 1991. [Google Scholar]
- Aletaha, R.; Sinegani, S.A.; Zafari, D. A survey on endophytic fungi within roots of Chenopodiaceae species under different environmental conditions. Mycosphere 2018, 9, 618–634. [Google Scholar] [CrossRef]
- Torres, M.S.; White, J.F.; Zhang, X.; Hinton, D.M.; Bacon, C.W. Endophyte-mediated adjustments in host morphology and physiology and effects on host fitness traits in grasses. Fungal Ecol. 2012, 5, 322–330. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fungi | Temperature (°C) | Culture in Days | |||||
|---|---|---|---|---|---|---|---|
| 5 °C | 15 °C | 25 °C | 30 °C | 35 °C | 40 °C | ||
| Group I (high salinity optima) | |||||||
| Arenariomyces trifucatus | - | 80 | 80 | 80 | 100 | - | 35 |
| Corollospora besarispora | - | 80 | 80 | 60 | 80 | - | 42 |
| Corollospora cinnamomea | - | 20 | 100 | 100 | 100 | 100 | 12 |
| Corollospora colossa | - | 40 | 100 | 80 | 100 | 100 | 30 |
| Corollospora maritima CM1 | - | - | 100 | 80 | 80 | 100 | 9 |
| Corollospora maritima PP4169 | - | 80 | 100 | 80 | - | nt | 13 |
| Corollospora maritima PP5089 | - | 20 | 100 | 80 | 100 | nt | 13 |
| Corollospora maritima PP5197 | - | 100 | 100 | 100 | - | nt | 49 |
| Corollospora novofusca | - | 20 | 40 | 100 | 100 | - | 20 |
| Corollospora pulchella | - | 80 | 60 | 60 | 80 | 100 | 15 |
| Corollospora gracilis | - | 100 | 80 | 80 | 100 | nt | 13 |
| Savoryella appendiculata | - | 100 | 60 | 80 | 80 | - | 48 |
| Torpedospora radiata | - | 80 | 60 | 60 | 200 | - | 8 |
| Asteromyces cruciatus | 80 | 80 | 100 | 100 | - | nt | 7 |
| Lulworthia crassa | - | 80 | 80 | 60 | - | - | 16 |
| Group II (low salinity optima) | |||||||
| Carbosphaerella leptosphaerioides | - | 60 | 40 | 60 | 60 | - | 28 |
| Corollospora lacera | 20 | 0 | 20 | 100 | - | - | 25 |
| Species | Growth Optimum | Remark | Reference |
|---|---|---|---|
| Oomycota | |||
| Haliphthoros milfordensis | 2.5–3.0% | Little or no growth at 0–1.5% | [53] |
| Labyrinthulomycota | |||
| Oblongichytrium multirudimentale | 2.5–3.0% | No growth at 0% or above 5.0% | [54] |
| Thraustochytrium motivum | 2.5–3.0% | No growth at 0% or above 5.0% | [54] |
| Thraustochytrium roseum | 2.5–5.0% | Little or no growth at 0.1–0.5% | [54] |
| Schizochytrium aggregatum | 2.5–3.0% | Little or no growth at 0.5–1.0% | [55] |
| Species | Growth Optimum | Sporulation Optimum |
|---|---|---|
| Halophytophthora avicennae | 10–20 (up to 60) | 10–30 (none above 35) |
| Halophytophthora vesicula | 15–25 (up to 60) | 10–15 (none above 35) |
| Phytopythium kandeliae | 10–35 (none above 35) | 15–35 (none above 35) |
| Salispina lobata | 20–40 (none above 40) | 30–40 (none above 40) |
| Salisapilia masteri | 20–35 (up to 60) | 30 (none above 40) |
| Hosts | Dominant (Most Speciose) Fungal Taxa | Reference |
|---|---|---|
| Fucus, Ulva, Enteromorpha (macroalgae) | Emericellopsis, Parasarocladium | [101] |
| Posidonia oceanica (seagrass) | Penicillium, Cladosporium, Acremonium | [139] |
| Posidonia oceanica (seagrass) | Dothideomycetes (Pleosporales and Capnodiales) and Leotiomycetes (Helotiales) | [140] |
| Posidonia oceanica (seagrass), Flabellia petiolata, Padina pavonica (macroalgae) | Penicillium, Cladosporium | [141] |
| Pterocladiella capillacea (macroalgae) | Aspergillus, Cladosporium, Penicillium, Rhodosporidium | [143] |
| Dysidea fragilis, Pachymatisma johnstonia, Sycon ciliatum (marine sponges) | Cladosporium, Penicillium | [145] |
| Grantia compressa (marine sponge) | Cladosporium, Penicillium | [146] |
| Holothuria poli (sea cucumber) | Aspergillus, Penicillium | [147] |
| Carcinus maenas (marine crab) | Ophiocordyceps | [148] |
| Suberites zeteki, Mycale armata (marine sponges) | Malasseziales | [149] |
| Zostera muelleri (seagrass) | Pleosporales, Wallemia ichthyophaga | [150] |
| Cymbastela concentrica, Scopalina sp., Tedania anhelans (marine sponges) | Epicoccum, Cladosporium | [151] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, E.B.G.; Ramakrishna, S.; Vikineswary, S.; Das, D.; Bahkali, A.H.; Guo, S.-Y.; Pang, K.-L. How Do Fungi Survive in the Sea and Respond to Climate Change? J. Fungi 2022, 8, 291. https://doi.org/10.3390/jof8030291
Jones EBG, Ramakrishna S, Vikineswary S, Das D, Bahkali AH, Guo S-Y, Pang K-L. How Do Fungi Survive in the Sea and Respond to Climate Change? Journal of Fungi. 2022; 8(3):291. https://doi.org/10.3390/jof8030291
Chicago/Turabian StyleJones, E. B. Gareth, Sundari Ramakrishna, Sabaratnam Vikineswary, Diptosh Das, Ali H. Bahkali, Sheng-Yu Guo, and Ka-Lai Pang. 2022. "How Do Fungi Survive in the Sea and Respond to Climate Change?" Journal of Fungi 8, no. 3: 291. https://doi.org/10.3390/jof8030291