
The Dermatophyte Trichophyton rubrum Induces Neutrophil Extracellular Traps Release by Human Neutrophils

 , , , , ,
, , , , ,  , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. T. rubrum Induces Neutrophil Extracellular Traps (NETs) Release in a Dose-Dependent Manner
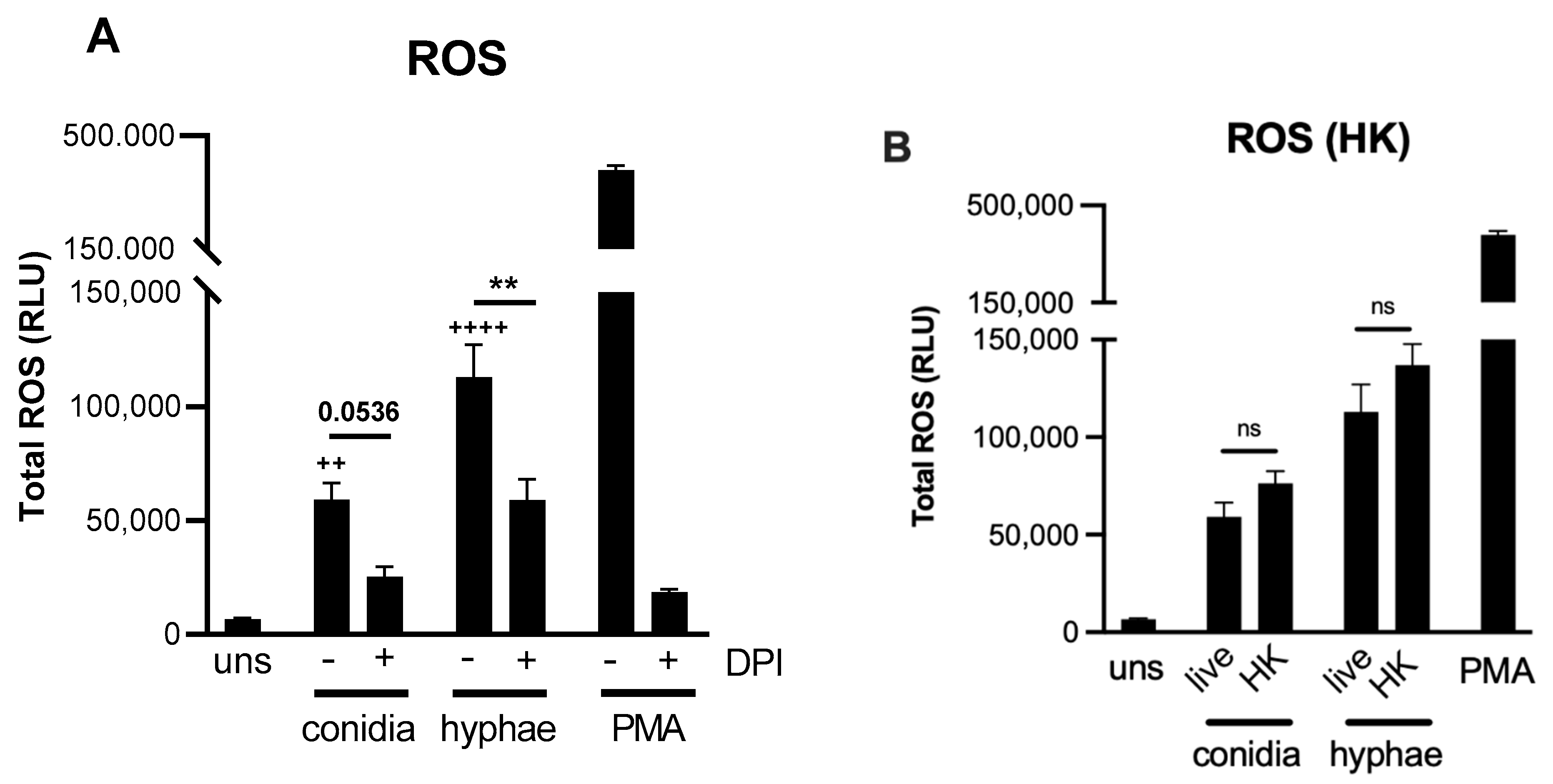
2.2. Reactive Oxygen Species (ROS) Production by Neutrophils against T. rubrum
2.3. NET Induction by T. rubrum Is Dependent on ROS Production
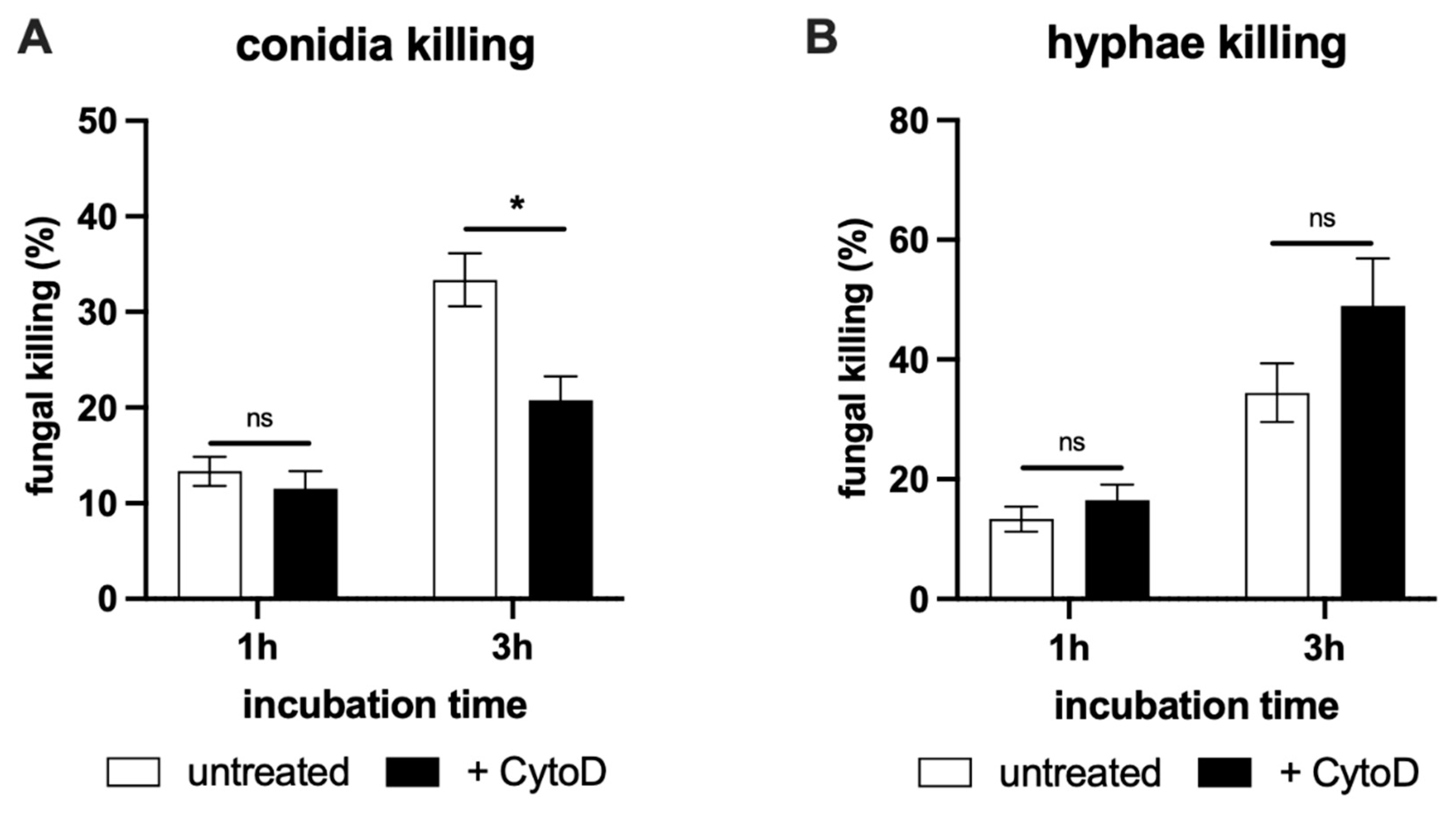
2.4. NET-Mediated Killing of T. rubrum
3. Discussion
4. Material and Methods
4.1. Fungi
4.2. Human Neutrophil Collection
4.3. Immunofluorescence of NETs
4.4. NETs (DNA/MPO) Quantification
4.5. Luminol-Amplified Chemiluminescence Assay
4.6. Assessment of NET-Mediated T. rubrum Killing
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Achterman, R.R.; White, T.C. Dermatophyte virulence factors: Identifying and analyzing genes that may contribute to chronic or acute skin infections. Int. J. Microbiol. 2012, 2012, 358305. [Google Scholar] [CrossRef] [Green Version]
- Nir-Paz, R.; Elinav, H.; Pierard, G.E.; Walker, D.; Maly, A.; Shapiro, M.; Barton, R.C.; Polacheck, I. Deep infection by Trichophyton rubrum in an immunocompromised patient. J. Clin. Microbiol. 2003, 41, 5298–5301. [Google Scholar] [CrossRef] [Green Version]
- Rouzaud, C.; Chosidow, O.; Brocard, A.; Fraitag, S.; Scemla, A.; Anglicheau, D.; Bouaziz, J.D.; Dupin, N.; Bougnoux, M.E.; Hay, R.; et al. Severe dermatophytosis in solid organ transplant recipients: A French retrospective series and literature review. Transpl. Infect. Dis. 2018, 20, e12799. [Google Scholar] [CrossRef] [PubMed]
- Romero, F.A.; Deziel, P.J.; Razonable, R.R. Majocchi’s granuloma in solid organ transplant recipients. Transpl. Infect. Dis. 2011, 13, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Drummond, R.A.; Gaffen, S.L.; Hise, A.G.; Brown, G.D. Innate Defense against Fungal Pathogens. Cold Spring Harb. Perspect. Med. 2014, 5, a019620. [Google Scholar] [CrossRef] [Green Version]
- Hogan, D.; Wheeler, R.T. The complex roles of NADPH oxidases in fungal infection. Cell Microbiol. 2014, 16, 1156–1167. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Rocha, J.D.; Nascimento, M.T.; Decote-Ricardo, D.; Côrte-Real, S.; Morrot, A.; Heise, N.; Nunes, M.P.; Previato, J.O.; Mendonça-Previato, L.; DosReis, G.A.; et al. Capsular polysaccharides from Cryptococcus neoformans modulate production of neutrophil extracellular traps (NETs) by human neutrophils. Sci. Rep. 2015, 5, 8008. [Google Scholar] [CrossRef]
- Guimaraes-Costa, A.B.; Nascimento, M.T.; Froment, G.S.; Soares, R.P.; Morgado, F.N.; Conceicao-Silva, F.; Saraiva, E.M. Leishmania amazonensis promastigotes induce and are killed by neutrophil extracellular traps. Proc. Natl. Acad. Sci. USA 2009, 106, 6748–6753. [Google Scholar] [CrossRef] [Green Version]
- Urban, C.F.; Nett, J.E. Neutrophil extracellular traps in fungal infection. Semin. Cell Dev. Biol. 2019, 89, 47–57. [Google Scholar] [CrossRef]
- Della Coletta, A.M.; Bachiega, T.F.; de Quaglia e Silva, J.C.; Victoriano de Campos Soares, A.M.; De Faveri, J.; Marques, S.A.; Alencar Marques, M.E.; Ximenes, V.F.; Dias-Melicio, L.A. Neutrophil Extracellular Traps Identification in Tegumentary Lesions of Patients with Paracoccidioidomycosis and Different Patterns of NETs Generation In Vitro. PLoS Negl. Trop. Dis. 2015, 9, e0004037. [Google Scholar] [CrossRef] [Green Version]
- Alflen, A.; Aranda Lopez, P.; Hartmann, A.K.; Maxeiner, J.; Bosmann, M.; Sharma, A.; Platten, J.; Ries, F.; Beckert, H.; Ruf, W.; et al. Neutrophil extracellular traps impair fungal clearance in a mouse model of invasive pulmonary aspergillosis. Immunobiology 2020, 225, 151867. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.S.; O’Brien, X.M.; Johnson, C.M.; Lavigne, L.M.; Reichner, J.S. An extracellular matrix-based mechanism of rapid neutrophil extracellular trap formation in response to Candida albicans. J. Immunol. 2013, 190, 4136–4148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejía, S.P.; Cano, L.E.; López, J.A.; Hernandez, O.; González, Á. Human neutrophils produce extracellular traps against Paracoccidioides brasiliensis. Microbiology 2015, 161, 1008–1017. [Google Scholar] [CrossRef]
- de Sousa, M.d.G.T.; Santana, G.B.; Criado, P.R.; Benard, G. Chronic widespread dermatophytosis due to Trichophyton rubrum: A syndrome associated with a Trichophyton-specific functional defect of phagocytes. Front. Microbiol. 2015, 6, 801. [Google Scholar] [CrossRef]
- Gazendam, R.P.; van Hamme, J.L.; Tool, A.T.; Hoogenboezem, M.; van den Berg, J.M.; Prins, J.M.; Vitkov, L.; van de Veerdonk, F.L.; van den Berg, T.K.; Roos, D.; et al. Human Neutrophils Use Different Mechanisms To Kill Aspergillus fumigatus Conidia and Hyphae: Evidence from Phagocyte Defects. J. Immunol. 2016, 196, 1272–1283. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, M.; Hakkim, A.; Brinkmann, V.; Siler, U.; Seger, R.A.; Zychlinsky, A.; Reichenbach, J. Restoration of NET formation by gene therapy in CGD controls aspergillosis. Blood 2009, 114, 2619–2622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heddergott, C.; Bruns, S.; Nietzsche, S.; Leonhardt, I.; Kurzai, O.; Kniemeyer, O.; Brakhage, A.A. The Arthroderma benhamiae hydrophobin HypA mediates hydrophobicity and influences recognition by human immune effector cells. Eukaryot. Cell 2012, 11, 673–682. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Yi, W.; Jiang, S.; Song, J.; Liang, P. Neutrophil Extracellular Traps Serve as Key Effector Molecules in the Protection Against Phialophora verrucosa. Mycopathologia 2021, 186, 367–375. [Google Scholar] [CrossRef]
- Branzk, N.; Lubojemska, A.; Hardison, S.E.; Wang, Q.; Gutierrez, M.G.; Brown, G.D.; Papayannopoulos, V. Neutrophils sense microbe size and selectively release neutrophil extracellular traps in response to large pathogens. Nat. Immunol. 2014, 15, 1017–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel cell death program leads to neutrophil extracellular traps. J. Cell Biol. 2007, 176, 231–241. [Google Scholar] [CrossRef]
- Parker, H.; Dragunow, M.; Hampton, M.B.; Kettle, A.J.; Winterbourn, C.C. Requirements for NADPH oxidase and myeloperoxidase in neutrophil extracellular trap formation differ depending on the stimulus. J. Leukoc. Biol. 2012, 92, 841–849. [Google Scholar] [CrossRef]
- Wu, S.Y.; Weng, C.L.; Jheng, M.J.; Kan, H.W.; Hsieh, S.T.; Liu, F.T.; Wu-Hsieh, B.A. Candida albicans triggers NADPH oxidase-independent neutrophil extracellular traps through dectin-2. PLoS Pathog. 2019, 15, e1008096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilsczek, F.H.; Salina, D.; Poon, K.K.; Fahey, C.; Yipp, B.G.; Sibley, C.D.; Robbins, S.M.; Green, F.H.; Surette, M.G.; Sugai, M.; et al. A novel mechanism of rapid nuclear neutrophil extracellular trap formation in response to Staphylococcus aureus. J. Immunol. 2010, 185, 7413–7425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz Caro, T.; Hermosilla, C.; Silva, L.M.; Cortes, H.; Taubert, A. Neutrophil extracellular traps as innate immune reaction against the emerging apicomplexan parasite Besnoitia besnoiti. PLoS ONE 2014, 9, e91415. [Google Scholar] [CrossRef]
- Swanson, K.V.; Deng, M.; Ting, J.P. The NLRP3 inflammasome: Molecular activation and regulation to therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Quach, A.; Ferrante, A. The Application of Dextran Sedimentation as an Initial Step in Neutrophil Purification Promotes Their Stimulation, due to the Presence of Monocytes. J. Immunol. Res. 2017, 2017, 1254792. [Google Scholar] [CrossRef]
- Vong, L.; Sherman, P.M.; Glogauer, M. Quantification and visualization of neutrophil extracellular traps (NETs) from murine bone marrow-derived neutrophils. Methods Mol. Biol. 2013, 1031, 41–50. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reis, A.P.C.; Celestrino, G.A.; Igoa, M.V.B.; Jesus, T.M.; França, T.T.; Moreira, D.V.S.; Rigato, P.O.; Sato, P.K.; Condino-Neto, A.; Noronha, I.L.; et al. The Dermatophyte Trichophyton rubrum Induces Neutrophil Extracellular Traps Release by Human Neutrophils. J. Fungi 2022, 8, 147. https://doi.org/10.3390/jof8020147
Reis APC, Celestrino GA, Igoa MVB, Jesus TM, França TT, Moreira DVS, Rigato PO, Sato PK, Condino-Neto A, Noronha IL, et al. The Dermatophyte Trichophyton rubrum Induces Neutrophil Extracellular Traps Release by Human Neutrophils. Journal of Fungi. 2022; 8(2):147. https://doi.org/10.3390/jof8020147
Chicago/Turabian StyleReis, Ana Paula Carvalho, Giovanna Azevedo Celestrino, Mariana Villas Bôas Igoa, Thais Martins Jesus, Tábata Takahashi França, Daniel Valério Silva Moreira, Paula Ordonhez Rigato, Paula Keiko Sato, Antonio Condino-Neto, Irene L. Noronha, and et al. 2022. "The Dermatophyte Trichophyton rubrum Induces Neutrophil Extracellular Traps Release by Human Neutrophils" Journal of Fungi 8, no. 2: 147. https://doi.org/10.3390/jof8020147