
Identification and Functional Characterization of a Putative Alternative Oxidase (Aox) in Sporisorium reilianum f. sp. zeae

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
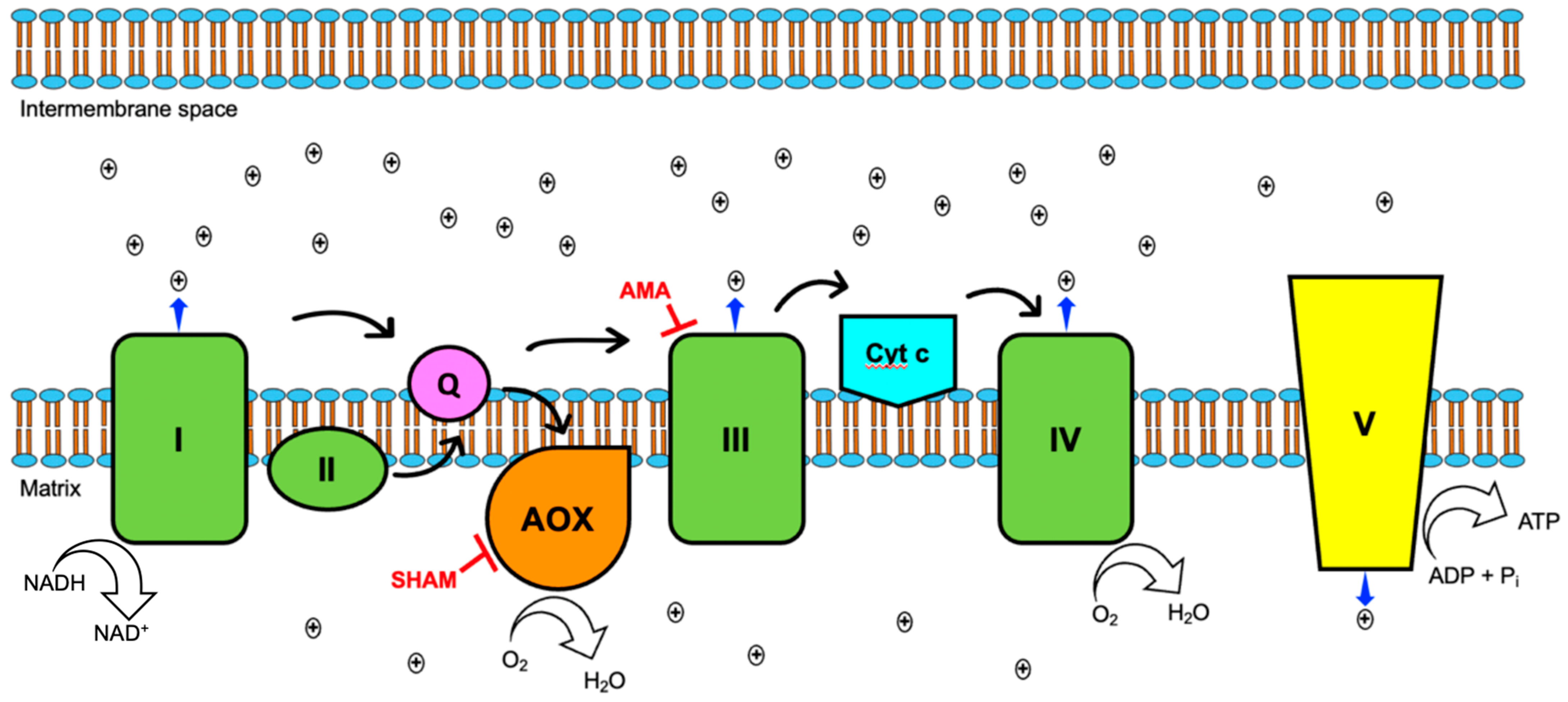{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Bioinformatics
2.3. Molecular Techniques
2.4. Growth Inhibition Assays
2.5. Pathogenicity Assays
2.6. Expression Studies
2.7. Microscopy
3. Results
3.1. Identification of a Putative Alternative Oxidase in SRZ
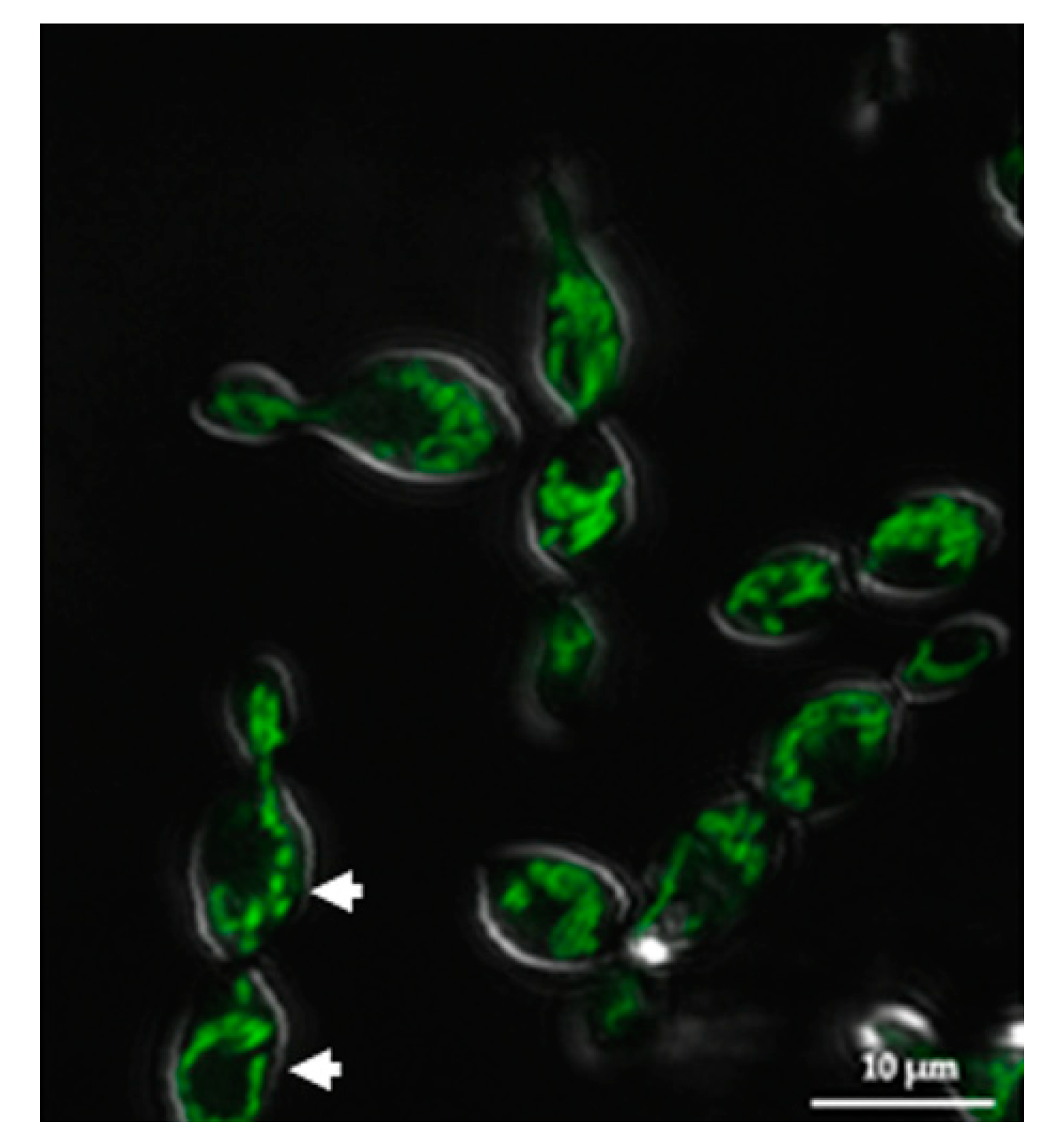
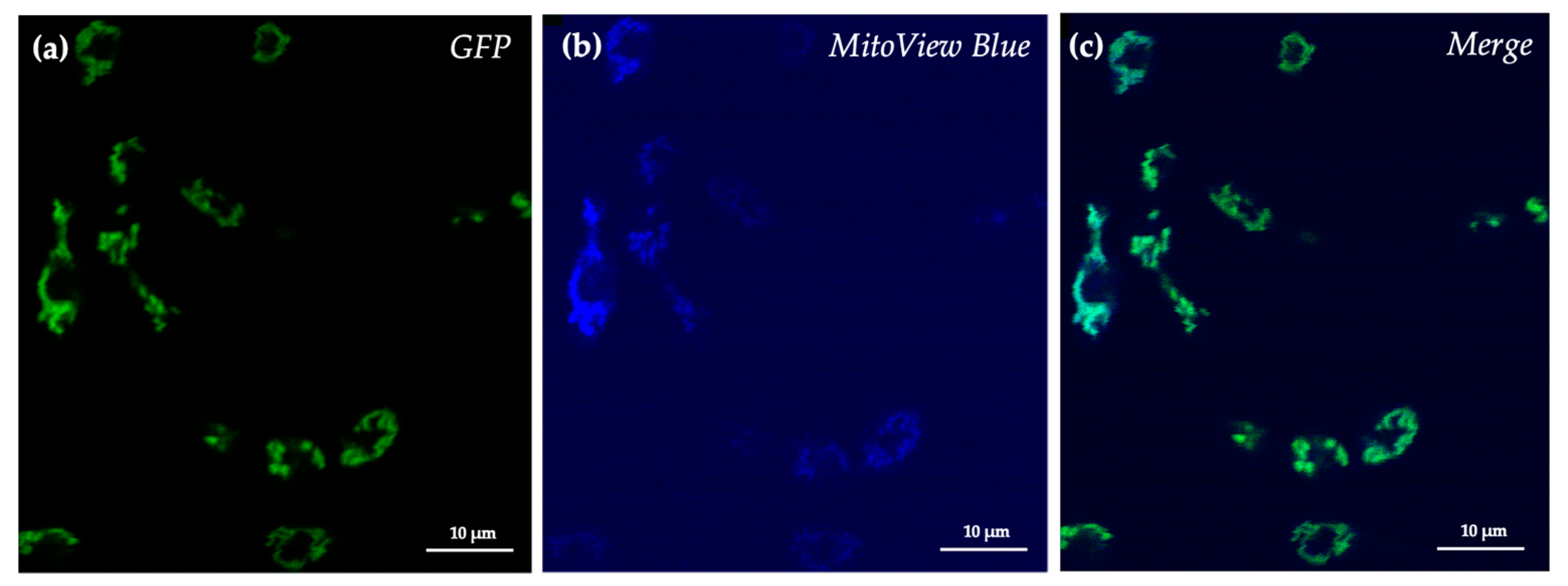
3.2. SRZ Aox Localizes to the Mitochondrial Membrane
3.3. Aox Provides an Alternative Route for the Transport of Electrons during Oxidative Phosphorylation
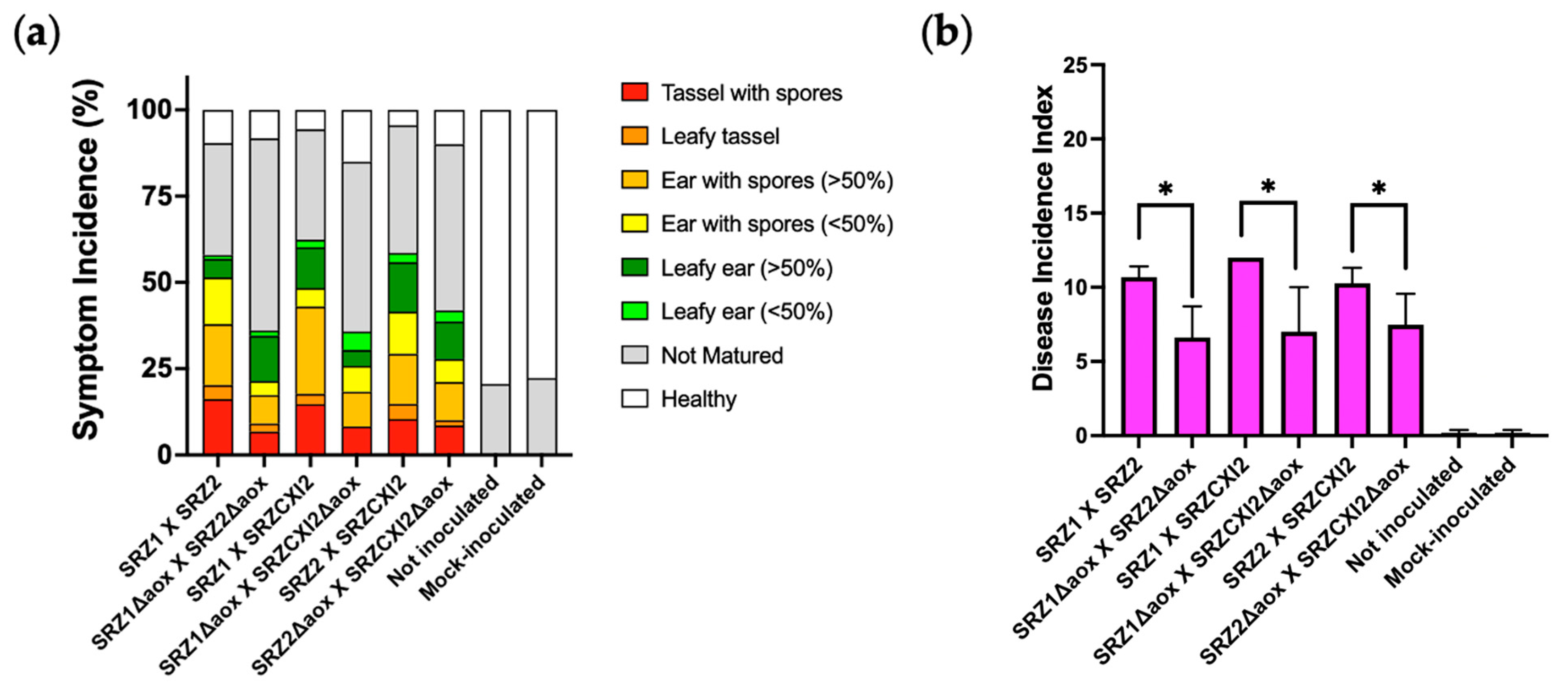
3.4. Alternative Respiration Is Involved in Pathogenicity of SRZ
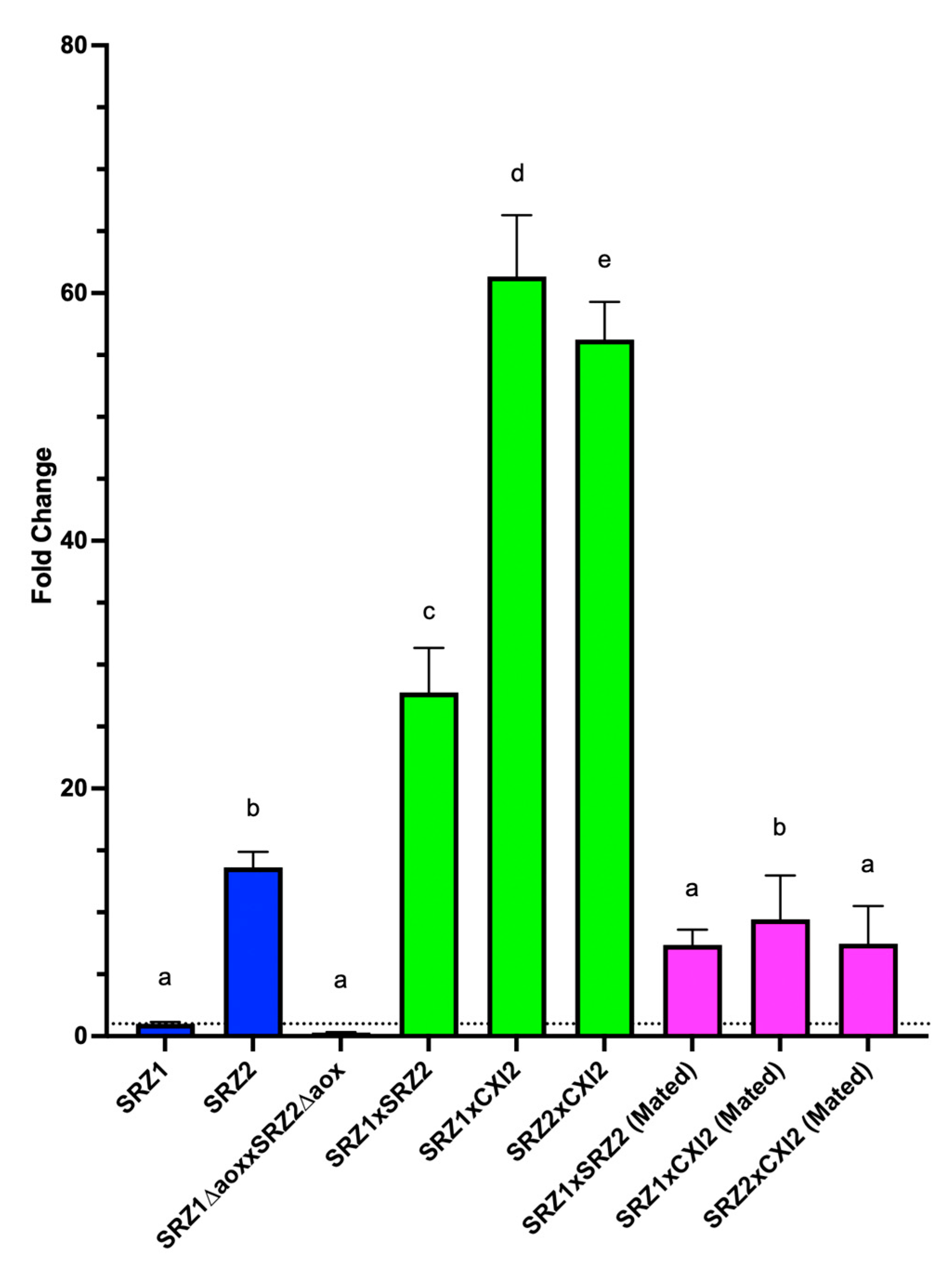
3.5. Aox Expression Is Upregulated during Teliospore Stage of Fungal Life Cycle
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martinez, C.; Roux, C.; Jauneau, A.; Dargent, R. The biological cycle of sporisorium reilianum f.sp. zeae: An overview using microscopy. Mycologia 2002, 94, 505–514. [Google Scholar] [PubMed]
- Schirawski, J.; Heinze, B.; Wagenknecht, M.; Kahmann, R. Mating type loci of Sporisorium reilianum: Novel pattern with three a and multiple b specificities. Eukaryot. Cell 2005, 4, 1317–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghareeb, H.; Becker, A.; Iven, T.; Feussner, I.; Schirawski, J. Sporisorium reilianum infection changes inflorescence and branching architectures of maize. Plant Physiol. 2011, 156, 2037–2052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghareeb, H.; Zhao, Y.; Schirawski, J. Sporisorium reilianum possesses a pool of effector proteins that modulate virulence on maize. Mol. Plant Pathol. 2019, 20, 124–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, S.B.; Bonner, W.D. Studies of electron transport in dry and imbibed peanut embryos. Plant Physiol. 1971, 48, 340–344. [Google Scholar] [CrossRef] [Green Version]
- Nosek, J.; Fukuhara, H. NADH dehydrogenase subunit genes in the mitochondrial DNA of yeasts. J. Bacteriol. 1994, 176, 5622–5630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, A.L.; Siedow, J.N. The regulation and nature of the cyanide-resistant alternative oxidase of plant mitochondria. Biochim. Biophys. Acta 1991, 1059, 121–140. [Google Scholar] [CrossRef]
- Erdal, S.; Genisel, M.; Turk, H.; Dumlupinar, R.; Demir, Y. Modulation of alternative oxidase to enhance tolerance against cold stress of chickpea by chemical treatments. J. Plant Physiol. 2015, 175, 95–101. [Google Scholar] [CrossRef]
- Grabelnych, O.I.; Borovik, O.A.; Tauson, E.L.; Pobezhimova, T.P.; Katyshev, A.I.; Pavlovskaya, N.S.; Koroleva, N.A.; Lyubushkina, I.V.; Bashmakov, V.Y.; Popov, V.N.; et al. Mitochondrial energy-dissipating systems (alternative oxidase, uncoupling proteins, and external NADH dehydrogenase) are involved in development of frost-resistance of winter wheat seedlings. Biochemistry 2014, 79, 506–519. [Google Scholar] [CrossRef]
- Smith, C.A.; Melino, V.J.; Sweetman, C.; Soole, K.L. Manipulation of alternative oxidase can influence salt tolerance in Arabidopsis thaliana. Physiol. Plant. 2009, 137, 459–472. [Google Scholar] [CrossRef]
- Wu, G.; Li, S.; Li, X.; Liu, Y.; Zhao, S.; Liu, B.; Zhou, H.; Lin, H. A functional alternative oxidase modulates plant salt tolerance in Physcomitrella patens. Plant Cell Physiol. 2019, 60, 1829–1841. [Google Scholar] [CrossRef] [PubMed]
- Mhadhbi, H.; Fotopoulos, V.; Mylona, P.V.; Jebara, M.; Aouani, M.E.; Polidoros, A.N. Alternative oxidase 1 (Aox1) gene expression in roots of Medicago truncatula is a genotype-specific component of salt stress tolerance. J. Plant Physiol. 2013, 170, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Dinakar, C.; Vishwakarma, A.; Raghavendra, A.S.; Padmasree, K. Alternative oxidase pathway optimizes photosynthesis during osmotic and temperature stress by regulating cellular ROS, malate valve and antioxidative systems. Front. Plant Sci. 2016, 7, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Oh, Y.; Li, H.; Baldwin, I.T.; Galis, I. Alternative oxidase in resistance to biotic stresses: Nicotiana attenuata AOX contributes to resistance to a pathogen and a piercing-sucking insect but not Manduca sexta larvae. Plant Physiol. 2012, 160, 1453–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Cui, R.; Xu, X.; Cheng, Q.; Li, X. Jasmonic acid- and ethylene-induced mitochondrial alternative oxidase stimulates Marssonina brunnea defense in poplar. Plant Cell Physiol. 2021, 61, 2031–2042. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, M.; Ajayi, W.; Hill, G.C. Biochemical and molecular properties of the Trypanosoma brucei alternative oxidase. Mol. Biochem. Parasitol. 1998, 95, 53–68. [Google Scholar] [CrossRef]
- Ott, R.; Chibale, K.; Anderson, S.; Chipeleme, A.; Chaudhuri, M.; Guerrah, A.; Colowick, N.; Hill, G.C. Novel inhibitors of the trypanosome alternative oxidase inhibit Trypanosoma brucei growth and respiration. Acta Trop. 2006, 100, 172–184. [Google Scholar] [CrossRef]
- Nakamura, K.; Fujioka, S.; Fukumoto, S.; Inoue, N.; Sakamoto, K.; Hirata, H.; Kido, Y.; Yabu, Y.; Suzuki, T.; Watanabe, Y.; et al. Trypanosome alternative oxidase, a potential therapeutic target for sleeping sickness, is conserved among Trypanosoma brucei subspecies. Parasitol. Int. 2010, 59, 560–564. [Google Scholar] [CrossRef]
- Roberts, C.W.; Roberts, F.; Henriquez, F.L.; Akiyoshi, D.; Samuel, B.U.; Richards, T.A.; Milhous, W.; Kyle, D.; McIntosh, L.; Hill, G.C.; et al. Evidence for mitochondrial-derived alternative oxidase in the apicomplexan parasite Cryptosporidium parvum: A potential anti-microbial agent target. Int. J. Parasitol. 2004, 34, 297–308. [Google Scholar] [CrossRef]
- Liu, S.; Roellig, D.M.; Guo, Y.; Li, N.; Frace, M.A.; Tang, K.; Zhang, L.; Feng, Y.; Xiao, L. Evolution of mitosome metabolism and invasion-related proteins in Cryptosporidium. BMC Genom. 2016, 17, 1006. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Wu, J.; Jamieson, P.A.; Zhang, C. Alternative oxidase is involved in the pathogenicity, development, and oxygen stress response of Botrytis cinerea. Phytopathology 2019, 109, 1679–1688. [Google Scholar] [CrossRef] [PubMed]
- Yukioka, H.; Inagaki, S.; Tanaka, R.; Katoh, K.; Miki, N.; Mizutani, A.; Masuko, M. Transcriptional activation of the alternative oxidase gene of the fungus Magnaporthe grisea by a respiratory-inhibiting fungicide and hydrogen peroxide. Biochim. Biophys. Acta 1998, 1442, 161–169. [Google Scholar] [CrossRef]
- Miguez, M.; Reeve, C.; Wood, P.M.; Hollomon, D.W. Alternative oxidase reduces the sensitivity of Mycosphaerella graminicola to QOI fungicides. Pest Manag. Sci. 2004, 60, 3–7. [Google Scholar] [CrossRef]
- Magnani, T.; Soriani, F.M.; Martins Vde, P.; Policarpo, A.C.; Sorgi, C.A.; Faccioli, L.H.; Curti, C.; Uyemura, S.A. Silencing of mitochondrial alternative oxidase gene of Aspergillus fumigatus enhances reactive oxygen species production and killing of the fungus by macrophages. J. Bioenerg. Biomembr. 2008, 40, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Liu, L.; Zhang, H.; Zhang, L.; Zhang, L.; Zhang, J.; Gao, Q.; Wang, D. Functional analysis of the mitochondrial alternative oxidase gene (aox1) from Aspergillus niger CGMCC 10142 and its effects on citric acid production. Appl. Microbiol. Biotechnol. 2018, 102, 7981–7995. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Lee, S.Y.; Woo, S.Y.; Chun, H.S. Alternative Oxidase: A potential target for controlling aflatoxin contamination and propagation of Aspergillus flavus. Front. Microbiol. 2020, 11, 419. [Google Scholar] [CrossRef]
- Luevano-Martinez, L.A.; Caldeira da Silva, C.C.; Nicastro, G.G.; Schumacher, R.I.; Kowaltowski, A.J.; Gomes, S.L. Mitochondrial alternative oxidase is determinant for growth and sporulation in the early diverging fungus Blastocladiella emersonii. Fungal Biol. 2019, 123, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Juarez, O.; Guerra, G.; Martinez, F.; Pardo, J.P. The mitochondrial respiratory chain of Ustilago maydis. Biochim. Biophys. Acta 2004, 1658, 244–251. [Google Scholar] [CrossRef] [Green Version]
- Juarez, O.; Guerra, G.; Velazquez, I.; Flores-Herrera, O.; Rivera-Perez, R.E.; Pardo, J.P. The physiologic role of alternative oxidase in Ustilago maydis. FEBS J. 2006, 273, 4603–4615. [Google Scholar] [CrossRef]
- Cardenas-Monroy, C.A.; Pohlmann, T.; Pinon-Zarate, G.; Matus-Ortega, G.; Guerra, G.; Feldbrugge, M.; Pardo, J.P. The mitochondrial alternative oxidase Aox1 is needed to cope with respiratory stress but dispensable for pathogenic development in Ustilago maydis. PLoS ONE 2017, 12, e0173389. [Google Scholar] [CrossRef]
- Schulz, B.; Banuett, F.; Dahl, M.; Schlesinger, R.; Schafer, W.; Martin, T.; Herskowitz, I.; Kahmann, R. The b alleles of U. maydis, whose combinations program pathogenic development, code for polypeptides containing a homeodomain-related motif. Cell 1990, 60, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3--new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A., 3rd; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Brachmann, A.; Konig, J.; Julius, C.; Feldbrugge, M. A reverse genetic approach for generating gene replacement mutants in Ustilago maydis. Mol. Genet. Genom. 2004, 272, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Terfruchte, M.; Joehnk, B.; Fajardo-Somera, R.; Braus, G.H.; Riquelme, M.; Schipper, K.; Feldbrugge, M. Establishing a versatile Golden Gate cloning system for genetic engineering in fungi. Fungal Genet. Biol. 2014, 62, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Stewart, C.R.; Martin, B.A.; Reding, L.; Cerwick, S. Seedling growth, mitochondrial characteristics, and alternative respiratory capacity of corn genotypes differing in cold tolerance. Plant Physiol. 1990, 92, 761–766. [Google Scholar] [CrossRef] [Green Version]
- Stewart, C.R.; Martin, B.A.; Reding, L.; Cerwick, S. Respiration and alternative oxidase in corn seedling tissues during germination at different temperatures. Plant Physiol. 1990, 92, 755–760. [Google Scholar] [CrossRef] [Green Version]
- Polidoros, A.N.; Mylona, P.V.; Pasentsis, K.; Scandalios, J.G.; Tsaftaris, A.S. The maize alternative oxidase 1a (Aox1a) gene is regulated by signals related to oxidative stress. Redox Rep. 2005, 10, 71–78. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendoza, H.; Culver, C.D.; Lamb, E.A.; Schroeder, L.A.; Khanal, S.; Müller, C.; Schirawski, J.; Perlin, M.H. Identification and Functional Characterization of a Putative Alternative Oxidase (Aox) in Sporisorium reilianum f. sp. zeae. J. Fungi 2022, 8, 148. https://doi.org/10.3390/jof8020148
Mendoza H, Culver CD, Lamb EA, Schroeder LA, Khanal S, Müller C, Schirawski J, Perlin MH. Identification and Functional Characterization of a Putative Alternative Oxidase (Aox) in Sporisorium reilianum f. sp. zeae. Journal of Fungi. 2022; 8(2):148. https://doi.org/10.3390/jof8020148
Chicago/Turabian StyleMendoza, Hector, Caroline D. Culver, Emma A. Lamb, Luke A. Schroeder, Sunita Khanal, Christian Müller, Jan Schirawski, and Michael H. Perlin. 2022. "Identification and Functional Characterization of a Putative Alternative Oxidase (Aox) in Sporisorium reilianum f. sp. zeae" Journal of Fungi 8, no. 2: 148. https://doi.org/10.3390/jof8020148