
Application of Non-Thermal Plasma to Fungal Resources



Abstract
:1. Introduction
2. NTP Technology
3. Inactivation of Fungi Using NTP
3.1. Inactivation of Fungal Spores, Cells, and Biofilms In Vitro
3.2. Inactivation of Fungi in Agriculture and Foods
3.3. Inactivation of Fungi in Medicine
4. Activation of Beneficial Fungi by NTP
4.1. Activation through Non-Mutational Ways
4.2. Activation through Mutagenesis
5. Mechanisms of Fungal Inactivation and Activation by NTP
6. Conclusions and Future Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Purvis, A.; Hector, A. Getting the Measure of Biodiversity. Nature 2000, 405, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S.; et al. The Amazing Potential of Fungi: 50 Ways We Can Exploit Fungi Industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Hussain, M.; Zhang, W.; Stadler, M.; Liu, X.; Xiang, M. Current Insights into Fungal Species Diversity and Perspective on Naming the Environmental DNA Sequences of Fungi. Mycology 2019, 10, 127–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, M.C.; Gow, N.A.R.; Gurr, S.J. Tackling Emerging Fungal Threats to Animal Health, Food Security and Ecosystem Resilience. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20160332. [Google Scholar] [CrossRef] [Green Version]
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and Multi-National Prevalence of Fungal Diseases-Estimate Precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef]
- Benedict, K.; Jackson, B.R.; Chiller, T.; Beer, K.D. Estimation of Direct Healthcare Costs of Fungal Diseases in the United States. Clin. Infect. Dis. 2019, 68, 1791–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konopka, J.; Casadevall, A.; Taylor, J.; Heitman, J.; Cowen, L. One Health: Fungal Pathogens of Humans, Animals, and Plants; Report on an American Academy of Microbiology Colloquium, Washington DC USA, 18 October 2017; America Society for Microbiology: Washington, DC, USA, 2019. [Google Scholar]
- Cairns, T.C.; Nai, C.; Meyer, V. How a Fungus Shapes Biotechnology: 100 Years of Aspergillus niger Research. Fungal Biol. Biotechnol. 2018, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- McKelvey, S.M.; Murphy, R.A. Biotechnological Use of Fungal Enzymes; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 201–225. [Google Scholar]
- Meyer, V.; Andersen, M.R.; Brakhage, A.A.; Braus, G.H.; Caddick, M.X.; Cairns, T.C.; de Vries, R.P.; Haarmann, T.; Hansen, K.; Hertz-Fowler, C.; et al. Current Challenges of Research on Filamentous Fungi in Relation to Human Welfare and a Sustainable Bio-Economy: A White Paper. Fungal Biol. Biotechnol. 2016, 3, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, M.E.; Park, B.J. Think Fungus-Prevention and Control of Fungal Infections. Emerg. Infect. Dis. 2013, 19, 1688–1689. [Google Scholar] [CrossRef] [Green Version]
- Kersey, P.J.; Collemare, J.; Cockel, C.; Das, D.; Dulloo, E.M.; Kelly, L.J.; Lettice, E.; Malécot, V.; Maxted, N.; Metheringham, C.; et al. Selecting for Useful Properties of Plants and Fungi—Novel Approaches, Opportunities, and Challenges. Plants People Planet 2020, 2, 409–420. [Google Scholar] [CrossRef]
- Avramidis, G.; Stüwe, B.; Wascher, R.; Bellmann, M.; Wieneke, S.; von Tiedemann, A.; Viöl, W. Fungicidal Effects of an Atmospheric Pressure Gas Discharge and Degradation Mechanisms. Surf. Coatings Technol. 2010, 205, S405–S408. [Google Scholar] [CrossRef]
- Xiong, Z.; Lu, X.P.; Feng, A.; Pan, Y.; Ostrikov, K. Highly Effective Fungal Inactivation in He+ O2 Atmospheric-Pressure Nonequilibrium Plasmas. Phys. Plasmas 2010, 17, 123502. [Google Scholar] [CrossRef] [Green Version]
- Ambrico, P.F.; Šimek, M.; Rotolo, C.; Morano, M.; Minafra, A.; Ambrico, M.; Pollastro, S.; Gerin, D.; Faretra, F.; De Miccolis Angelini, R.M. Surface Dielectric Barrier Discharge Plasma: A Suitable Measure Against Fungal Plant Pathogens. Sci. Rep. 2020, 10, 3673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnabel, U.; Yarova, K.; Zessin, B.; Stachowiak, J.; Ehlbeck, J. The Combination of Plasma-Processed Air (PPA) and Plasma-Treated Water (PTW) Causes Synergistic Inactivation of Candida albicans SC5314. Appl. Sci. 2020, 10, 3303. [Google Scholar] [CrossRef]
- Shimada, K.; Takashima, K.; Kimura, Y.; Nihei, K.; Konishi, H.; Kaneko, T. Humidification Effect of Air Plasma Effluent Gas on Suppressing Conidium Germination of a Plant Pathogenic Fungus in the Liquid Phase. Plasma Process. Polym. 2020, 17, 1900004. [Google Scholar] [CrossRef]
- Farasat, M.; Arjmand, S.; Ranaei Siadat, S.O.; Sefidbakht, Y.; Ghomi, H. The Effect of Non-Thermal Atmospheric Plasma on the Production and Activity of Recombinant Phytase Enzyme. Sci. Rep. 2018, 8, 16647. [Google Scholar] [CrossRef]
- Gao, X.; Liu, E.; Yin, Y.; Yang, L.; Huang, Q.; Chen, S.; Ho, C.T. Enhancing Activities of Salt-Tolerant Proteases Secreted by Aspergillus oryzae Using Atmospheric and Room-Temperature Plasma Mutagenesis. J. Agric. Food Chem. 2020, 68, 2757–2764. [Google Scholar] [CrossRef] [PubMed]
- Veerana, M.; Mitra, S.; Ki, S.H.; Kim, S.M.; Choi, E.H.; Lee, T.; Park, G. Plasma-Mediated Enhancement of Enzyme Secretion in Aspergillus oryzae. Microb. Biotechnol. 2021, 14, 262–276. [Google Scholar] [CrossRef]
- Fridman, A. Plasma Chemistry; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Domonkos, M.; Tichá, P.; Trejbal, J.; Demo, P. Applications of Cold Atmospheric Pressure Plasma Technology in Medicine, Agriculture and Food Industry. Appl. Sci. 2021, 11, 4809. [Google Scholar] [CrossRef]
- Chen, Z.; Xu, R.G.; Chen, P.; Wang, Q. Potential Agricultural and Biomedical Applications of Cold Atmospheric Plasma-Activated Liquids with Self-Organized Patterns Formed at the Interface. IEEE Trans. Plasma Sci. 2020, 48, 3455–3471. [Google Scholar] [CrossRef]
- Mijatović, S.; Savić-Radojević, A.; Plješa-Ercegovac, M.; Simić, T.; Nicoletti, F.; Maksimović-Ivanić, D. The Double-Faced Role of Nitric Oxide and Reactive Oxygen Species in Solid Tumors. Antioxidants 2020, 9, 374. [Google Scholar] [CrossRef]
- Sakudo, A.; Yagyu, Y.; Onodera, T. Disinfection and Sterilization Using Plasma Technology: Fundamentals and Future Perspectives for Biological Applications. Int. J. Mol. Sci. 2019, 20, 5216. [Google Scholar] [CrossRef] [Green Version]
- Braný, D.; Dvorská, D.; Halašová, E.; Škovierová, H. Cold Atmospheric Plasma: A Powerful Tool for Modern Medicine. Int. J. Mol. Sci. 2020, 21, 2932. [Google Scholar] [CrossRef] [Green Version]
- Julák, J.; Soušková, H.; Scholtz, V.; Kvasničková, E.; Savická, D.; Kříha, V. Comparison of Fungicidal Properties of Non-Thermal Plasma Produced by Corona Discharge and Dielectric Barrier Discharge. Folia Microbiol. 2018, 63, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Ki, S.H.; Noh, H.; Ahn, G.R.; Kim, S.H.; Kaushik, N.K.; Choi, E.H.; Lee, G.J. Influence of Nonthermal Atmospheric Plasma-Activated Water on the Structural, Optical, and Biological Properties of Aspergillus brasiliensis Spores. Appl. Sci. 2020, 10, 6378. [Google Scholar] [CrossRef]
- Intanon, W.; Vichiansan, N.; Leksakul, K.; Boonyawan, D.; Kumla, J.; Suwannarach, N.; Lumyong, S. Inhibition of the Aflatoxin-Producing Fungus Aspergillus flavus by a Plasma Jet System. J. Food Process. Preserv. 2021, 45, e15045. [Google Scholar] [CrossRef]
- Hojnik, N.; Modic, M.; Ni, Y.; Filipič, G.; Cvelbar, U.; Walsh, J.L. Effective Fungal Spore Inactivation with an Environmentally Friendly Approach Based on Atmospheric Pressure Air Plasma. Environ. Sci. Technol. 2019, 53, 1893–1904. [Google Scholar] [CrossRef] [PubMed]
- Nojima, H.; Park, R.E.; Kwon, J.H.; Suh, I.; Jeon, J.; Ha, E.; On, H.K.; Kim, H.R.; Choi, K.; Lee, K.H.; et al. Novel Atmospheric Pressure Plasma Device Releasing Atomic Hydrogen: Reduction of Microbial-Contaminants and OH Radicals in the Air. J. Phys. D Appl. Phys. 2007, 40, 501–509. [Google Scholar] [CrossRef]
- Park, B.J.; Lee, D.H.; Park, J.-C.; Lee, I.-S.; Lee, K.-Y.; Hyun, S.O.; Chun, M.-S.; Chung, K.-H. Sterilization Using a Microwave-Induced Argon Plasma System at Atmospheric Pressure. Phys. Plasmas 2003, 10, 4539–4544. [Google Scholar] [CrossRef]
- Park, J.C.; Park, B.J.; Han, D.W.; Lee, D.H.; Lee, I.S.; Hyun, S.O.; Chun, M.S.; Chung, K.H.; Aihara, M.; Takatori, K. Fungal Sterilization Using Microwave-Induced Argon Plasma at Atmospheric Pressure. J. Microbiol. Biotechnol. 2004, 14, 188–192. [Google Scholar]
- Herceg, Z.; Režek Jambrak, A.; Vukušić, T.; Stulić, V.; Stanzer, D.; Milošević, S. The Effect of High-Power Ultrasound and Gas Phase Plasma Treatment on Aspergillus spp. and Penicillium spp. Count in Pure Culture. J. Appl. Microbiol. 2015, 118, 132–141. [Google Scholar] [CrossRef]
- Soušková, H.; Scholtz, V.; Julák, J.; Kommová, L.; Savická, D.; Pazlarová, J. The Survival of Micromycetes and Yeasts Under the Low-Temperature Plasma Generated in Electrical Discharge. Folia microbiol. 2011, 56, 77–79. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, S.; Kawasaki, Y.; Izawa, S. Ferrous Chloride and Ferrous Sulfate Improve the Fungicidal Efficacy of Cold Atmospheric Argon Plasma on Melanized Aureobasidium pullulans. J. Biosci. Bioeng. 2019, 128, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Tyczkowska-Sieroń, E.; Kałużewski, T.; Grabiec, M.; Kałużewski, B.; Tyczkowski, J. Genotypic and Phenotypic Changes in Candida albicans as a Result of Cold Plasma Treatment. Int. J. Mol. Sci. 2020, 21, 8100. [Google Scholar] [CrossRef] [PubMed]
- Nishime, T.M.C.; Borges, A.C.; Koga-Ito, C.Y.; Machida, M.; Hein, L.R.O.; Kostov, K.G. Non-Thermal Atmospheric Pressure Plasma Jet Applied to Inactivation of Different Microorganisms. Surf. Coatings Technol. 2017, 312, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Kostov, K.G.; Borges, A.C.; Koga-Ito, C.Y.; Nishime, T.M.C.; Prysiazhnyi, V.; Honda, R.Y. Inactivation of Candida albicans by Cold Atmospheric Pressure Plasma Jet. IEEE Trans. Plasma Sci. 2015, 43, 770–775. [Google Scholar] [CrossRef]
- Laurita, R.; Barbieri, D.; Gherardi, M.; Colombo, V.; Lukes, P. Chemical Analysis of Reactive Species and Antimicrobial Activity of Water Treated by Nanosecond Pulsed DBD Air Plasma. Clin. Plasma Med. 2015, 3, 53–61. [Google Scholar] [CrossRef]
- Rahimi-Verki, N.; Shapoorzadeh, A.; Razzaghi-Abyaneh, M.; Atyabi, S.M.; Shams-Ghahfarokhi, M.; Jahanshiri, Z.; Gholami-Shabani, M. Cold Atmospheric Plasma Inhibits the Growth of Candida albicans by Affecting Ergosterol Biosynthesis and Suppresses the Fungal Virulence Factors In Vitro. Photodiagnosis Photodyn. Ther. 2016, 13, 66–72. [Google Scholar] [CrossRef]
- Sedghizadeh, P.P.; Chen, M.; Schaudinn, C.; Gorur, A.; Jiang, C. Inactivation Kinetics Study of an Atmospheric-Pressure Cold-Plasma Jet Against Pathogenic Microorganisms. IEEE Trans. Plasma Sci. 2012, 40, 2879–2882. [Google Scholar] [CrossRef]
- Song, Y.; Liu, D.; Ji, L.; Wang, W.; Zhao, P.; Quan, C.; Niu, J.; Zhang, X. The Inactivation of Resistant Candida albicans in a Sealed Package by Cold Atmospheric Pressure Plasmas. Plasma Processes Polym. 2012, 9, 17–21. [Google Scholar] [CrossRef]
- Shi, X.-M.; Zhang, G.-J.; Yuan, Y.-K.; Ma, Y.; Xu, G.-M.; Yang, Y. Research on the Inactivation Effect of Low-Temperature Plasma on Candida albicans. IEEE Trans. Plasma Sci. 2008, 36, 498–503. [Google Scholar] [CrossRef]
- Siadati, S.; Pet’ková, M.; Kenari, A.J.; Kyzek, S.; Gálová, E.; Zahoranová, A. Effect of a Non-Thermal Atmospheric Pressure Plasma Jet on Four Different Yeasts. J. Phys. D: Appl. Phys. 2021, 54, 025204. [Google Scholar] [CrossRef]
- Lu, Q.; Liu, D.; Song, Y.; Zhou, R.; Niu, J. Inactivation of the Tomato Pathogen Cladosporium fulvum by an Atmospheric-Pressure Cold Plasma Jet. Plasma Process. Polym. 2014, 11, 1028–1036. [Google Scholar] [CrossRef]
- Wu, M.; Liu, C.; Chiang, C.; Lin, Y.; Lin, Y.; Chang, Y.; Wu, J. Inactivation Effect of Colletotrichum gloeosporioides by Long-Lived Chemical Species Using Atmospheric-Pressure Corona Plasma-Activated Water. IEEE Trans. Plasma Sci. 2019, 47, 1100–1104. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, I.H.; Kim, D.; Kim, S.H.; Kwon, Y.-W.; Han, G.-H.; Cho, G.; Choi, E.H.; Lee, G.J. Effects of Reactive Oxygen Species on the Biological, Structural, and Optical Properties of Cordyceps pruinosa Spores. RSC Adv. 2016, 6, 30699–30709. [Google Scholar] [CrossRef]
- Lee, G.J.; Sim, G.B.; Choi, E.H.; Kwon, Y.-W.; Kim, J.Y.; Jang, S.; Kim, S.H. Optical and Structural Properties of Plasma-Treated Cordyceps bassiana Spores as Studied by Circular Dichroism, Absorption, and Fluorescence Spectroscopy. J. Appl. Phys. 2015, 117, 023303. [Google Scholar] [CrossRef]
- Noh, H.; Kim, J.E.; Kim, J.Y.; Kim, S.H.; Han, I.; Lim, J.S.; Ki, S.H.; Choi, E.H.; Lee, G.J. Spore Viability and Cell Wall Integrity of Cordyceps pruinosa Treated with an Electric Shock-Free, Atmospheric-Pressure Air Plasma Jet. Appl. Sci. 2019, 9, 3921. [Google Scholar] [CrossRef] [Green Version]
- Na, Y.H.; Park, G.; Choi, E.H.; Uhm, H.S. Effects of the Physical Parameters of a Microwave Plasma Jet on the Inactivation of Fungal Spores. Thin Solid Films 2013, 547, 125–131. [Google Scholar] [CrossRef]
- Panngom, K.; Lee, S.H.; Park, D.H.; Sim, G.B.; Kim, Y.H.; Uhm, H.S.; Park, G.; Choi, E.H. Non-Thermal Plasma Treatment Diminishes Fungal Viability and Up-Regulates Resistance Genes in a Plant Host. PLoS ONE 2014, 9, e99300. [Google Scholar] [CrossRef]
- Lee, G.J.; Park, G.; Choi, E.H. Optical and Biological Properties of Plasma-Treated Neurospora crassa Spores as Studied by Absorption, Circular Dichroism, and Raman Spectroscopy. J. Korean Phys. Soc. 2017, 71, 670–678. [Google Scholar] [CrossRef]
- Kang, M.H.; Hong, Y.J.; Attri, P.; Sim, G.B.; Lee, G.J.; Panngom, K.; Kwon, G.C.; Choi, E.H.; Uhm, H.S.; Park, G. Analysis of the Antimicrobial Effects of Nonthermal Plasma on Fungal Spores in Ionic Solutions. Free Radic. Biol. Med. 2014, 72, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Park, G.; Ryu, Y.H.; Hong, Y.J.; Choi, E.H.; Uhm, H.S. Cellular and Molecular Responses of Neurospora crassa to Non-Thermal Plasma at Atmospheric Pressure. Appl. Phys. Lett. 2012, 100, 063703. [Google Scholar] [CrossRef]
- Liu, K.; Wang, C.; Hu, H.; Lei, J.; Han, L. Indirect Treatment Effects of Water–Air MHCD Jet on the Inactivation of Penicillium digitatum Suspension. IEEE Trans. Plasma Sci. 2016, 44, 2729–2737. [Google Scholar] [CrossRef]
- Hashizume, H.; Ohta, T.; Fengdong, J.; Takeda, K.; Ishikawa, K.; Hori, M.; Ito, M. Inactivation Effects of Neutral Reactive-Oxygen Species on Penicillium digitatum Spores Using Non-Equilibrium Atmospheric-Pressure Oxygen Radical Source. Appl. Phys. Lett. 2013, 103, 153708. [Google Scholar] [CrossRef]
- Iseki, S.; Hashizume, H.; Jia, F.; Takeda, K.; Ishikawa, K.; Ohta, T.; Ito, M.; Hori, M. Inactivation of Penicillium digitatum Spores by a High-Density Ground-State Atomic Oxygen-Radical Source Employing an Atmospheric-Pressure Plasma. Appl. Phys. Express 2011, 4, 116201. [Google Scholar] [CrossRef]
- Ishikawa, K.; Mizuno, H.; Tanaka, H.; Tamiya, K.; Hashizume, H.; Ohta, T.; Ito, M.; Iseki, S.; Takeda, K.; Kondo, H.; et al. Real-Time In Situ Electron Spin Resonance Measurements on Fungal Spores of Penicillium digitatum During Exposure of Oxygen Plasmas. Appl. Phys. Lett. 2012, 101, 013704. [Google Scholar] [CrossRef] [Green Version]
- Iseki, S.; Ohta, T.; Aomatsu, A.; Ito, M.; Kano, H.; Higashijima, Y.; Hori, M. Rapid Inactivation of Penicillium digitatum Spores Using High-Density Nonequilibrium Atmospheric Pressure Plasma. Appl. Phys. Lett. 2010, 96, 153704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veremii, Y.; Andriiash, I.; Tsvyd, N.; Chernyak, V.Y.; Sukhomlyn, M.; Martysh, E. Influence of Cold Atmospheric Plasma of Microdischarge on Fungal Mycelium and Spores Growing. Probl. At. Sci. Technol. 2019, 119, 233–236. [Google Scholar]
- Itooka, K.; Takahashi, K.; Izawa, S. Fluorescence Microscopic Analysis of Antifungal Effects of Cold Atmospheric Pressure Plasma in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2016, 100, 9295–9304. [Google Scholar] [CrossRef]
- Ryu, Y.H.; Kim, Y.H.; Lee, J.Y.; Shim, G.B.; Uhm, H.S.; Park, G.; Choi, E.H. Effects of Background Fluid on the Efficiency of Inactivating Yeast with Non-Thermal Atmospheric Pressure Plasma. PLoS ONE 2013, 8, e66231. [Google Scholar]
- Chen, H.; Bai, F.; Xiu, Z. Oxidative Stress Induced in Saccharomyces cerevisiae Exposed to Dielectric Barrier Discharge Plasma in Air at Atmospheric Pressure. IEEE Trans. Plasma Sci. 2010, 38, 1885–1891. [Google Scholar] [CrossRef]
- Feng, H.; Wang, R.; Sun, P.; Wu, H.; Liu, Q.; Fang, J.; Zhu, W.; Li, F.; Zhang, J. A Study of Eukaryotic Response Mechanisms to Atmospheric Pressure Cold Plasma by Using Saccharomyces cerevisiae Single Gene Mutants. Appl. Phys. Lett. 2010, 97, 131501. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Ma, R.; Zhu, Y.; Du, M.; Zhang, H.; Jiao, Z. A Systematic Study of the Antimicrobial Mechanisms of Cold Atmospheric-Pressure Plasma for Water Disinfection. Sci. Total Environ. 2020, 703, 134965. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhu, Y.; Cui, D.; Du, M.; Wang, J.; Ma, R.; Jiao, Z. Evaluating the Roles of OH radicals, H2O2, ORP and pH in the Inactivation of Yeast Cells on a Tissue Model by Surface Micro-Discharge Plasma. J. Phys. D Appl. Phys. 2019, 52, 395201. [Google Scholar] [CrossRef]
- Ma, R.N.; Feng, H.Q.; Liang, Y.D.; Zhang, Q.; Tian, Y.; Su, B.; Zhang, J.; Fang, J. An Atmospheric-Pressure Cold Plasma Leads to Apoptosis in Saccharomyces cerevisiae by Accumulating Intracellular Reactive Oxygen Species and Calcium. J. Phys. D Appl. Phys. 2013, 46, 285401. [Google Scholar] [CrossRef]
- Ma, R.; Feng, H.; Li, F.; Liang, Y.; Zhang, Q.; Zhu, W.; Zhang, J.; Becker, K.H.; Fang, J. An Evaluation of Anti-oxidative Protection for Cells against Atmospheric Pressure Cold Plasma Treatment. Appl. Phys. Lett. 2012, 100, 123701. [Google Scholar] [CrossRef] [Green Version]
- Shapourzadeh, A.; Rahimi-Verki, N.; Atyabi, S.M.; Shams-Ghahfarokhi, M.; Jahanshiri, Z.; Irani, S.; Razzaghi-Abyaneh, M. Inhibitory Effects of Cold Atmospheric Plasma on the Growth, Ergosterol Biosynthesis, and Keratinase Activity in Trichophyton rubrum. Arch. Biochem. Biophys. 2016, 608, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Los, A.; Ziuzina, D.; Boehm, D.; Cullen, P.J.; Bourke, P. Inactivation Efficacies and Mechanisms of Gas Plasma and Plasma-Activated Water Against Aspergillus flavus Spores and Biofilms: A Comparative Study. Appl. Environ. Microbiol. 2020, 86, e02619-19. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Duan, J.; Xu, J.; Ma, M.; Chai, B.; He, G.; Gan, L.; Zhang, S.; Duan, X.; Lu, X.; et al. Candida albicans Biofilm Inactivated by Cold Plasma Treatment In Vitro and In Vivo. Plasma Process. Polym. 2020, 17, 1900068. [Google Scholar] [CrossRef]
- Borges, A.C.; Castaldelli Nishime, T.M.; Kostov, K.G.; de Morais Gouvêa Lima, G.; Lacerda Gontijo, A.V.; de Carvalho, J.N.M.M.; Yzumi Honda, R.; Yumi Koga-Ito, C. Cold Atmospheric Pressure Plasma Jet Modulates Candida Albicans Virulence Traits. Clin. Plasma Med. 2017, 7, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Maisch, T.; Shimizu, T.; Isbary, G.; Heinlin, J.; Karrer, S.; Klämpfl, T.G.; Li, Y.F.; Morfill, G.; Zimmermann, J.L. Contact-Free Inactivation of Candida albicans Biofilms by Cold Atmospheric Air Plasma. Appl. Environ. Microbiol. 2012, 78, 4242–4247. [Google Scholar] [CrossRef] [Green Version]
- Fricke, K.; Koban, I.; Tresp, H.; Jablonowski, L.; Schröder, K.; Kramer, A.; Weltmann, K.-D.; von Woedtke, T.; Kocher, T. Atmospheric Pressure Plasma: A High-Performance Tool for the Efficient Removal of Biofilms. PLoS ONE 2012, 7, e42539. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yu, S.; Sun, P.; Wu, H.; Zhu, W.; Liu, W.; Zhang, J.; Fang, J.; Li, R. Inactivation of Candida Biofilms by Non-Thermal Plasma and Its Enhancement for Fungistatic Effect of Antifungal Drugs. PLoS ONE 2012, 7, e40629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koban, I.; Matthes, R.; Hübner, N.-O.; Welk, A.; Meisel, P.; Holtfreter, B.; Sietmann, R.; Kindel, E.; Weltmann, K.-D.; Kramer, A.; et al. Treatment of Candida albicans Biofilms with Low-Temperature Plasma Induced by Dielectric Barrier Discharge and Atmospheric Pressure Plasma Jet. New J. Phys. 2010, 12, 073039. [Google Scholar] [CrossRef]
- Zahoranová, A.; Hoppanová, L.; Šimončicová, J.; Tučeková, Z.; Medvecká, V.; Hudecová, D.; Kaliňáková, B.; Kováčik, D.; Černák, M. Effect of Cold Atmospheric Pressure Plasma on Maize Seeds: Enhancement of Seedlings Growth and Surface Microorganisms Inactivation. Plasma Chem. Plasma Process. 2018, 38, 969–988. [Google Scholar] [CrossRef]
- Kordas, L.; Pusz, W.; Czapka, T.; Kacprzyk, R. The Effect of Low-Temperature Plasma on Fungus Colonization of Winter Wheat Grain and Seed Quality. Pol. J. Environ. Stud. 2015, 24, 433–438. [Google Scholar]
- Zahoranová, A.; Henselová, M.; Hudecová, D.; Kaliňáková, B.; Kováčik, D.; Medvecká, V.; Černák, M. Effect of Cold Atmospheric Pressure Plasma on the Wheat Seedlings Vigor and on the Inactivation of Microorganisms on the Seeds Surface. Plasma Chem. Plasma Process. 2016, 36, 397–414. [Google Scholar] [CrossRef]
- Devi, Y.; Thirumdas, R.; Sarangapani, C.; Deshmukh, R.R.; Annapure, U.S. Influence of Cold Plasma on Fungal Growth and Aflatoxins Production on Groundnuts. Food Control 2017, 77, 187–191. [Google Scholar] [CrossRef]
- Waskow, A.; Betschart, J.; Butscher, D.; Oberbossel, G.; Klöti, D.; Büttner-Mainik, A.; Adamcik, J.; von Rohr, P.R.; Schuppler, M. Characterization of Efficiency and Mechanisms of Cold Atmospheric Pressure Plasma Decontamination of Seeds for Sprout Production. Front. Microbiol. 2018, 9, 3164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basaran, P.; Basaran-Akgul, N.; Oksuz, L. Elimination of Aspergillus parasiticus from Nut Surface with Low Pressure Cold Plasma (LPCP) Treatment. Food Microbiol. 2008, 25, 626–632. [Google Scholar] [CrossRef]
- Rüntzel, C.L.; Da Silva, J.R.; Da Silva, B.A.; Moecke, E.S.; Scussel, V.M. Effect of Cold Plasma on Black Beans (Phaseolus vulgaris L.), Fungi Inactivation and Micro-Structures Stability. Emir. J. Food Agric. 2019, 31, 864–873. [Google Scholar] [CrossRef]
- Selcuk, M.; Oksuz, L.; Basaran, P. Decontamination of Grains and Legumes Infected with Aspergillus spp. and Penicillum spp. by Cold Plasma Treatment. Bioresour. Technol. 2008, 99, 5104–5109. [Google Scholar] [CrossRef] [PubMed]
- Štěpánová, V.; Slavíček, P.; Kelar, J.; Prášil, J.; Smékal, M.; Stupavská, M.; Jurmanová, J.; Černák, M. Atmospheric Pressure Plasma Treatment of Agricultural Seeds of Cucumber (Cucumis sativus L.) and Pepper (Capsicum annuum L.) with Effect on Reduction of Diseases and Germination Improvement. Plasma Process. Polym. 2018, 15, e1700076. [Google Scholar] [CrossRef]
- Lee, Y.; Lee, Y.Y.; Kim, Y.S.; Balaraju, K.; Mok, Y.S.; Yoo, S.J.; Jeon, Y. Enhancement of Seed Germination and Microbial Disinfection on Ginseng by Cold Plasma Treatment. J. Ginseng Res. 2021, 45, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Puligundla, P.; Kim, J.W.; Mok, C. Effect of Atmospheric Pressure Plasma Treatment on Seed Decontamination and Sprouting of Pak Choi (Brassica rapa L. subsp chinensis (L.) Hanelt). Chiang Mai J. Sci. 2018, 45, 2679–2690. [Google Scholar]
- Ambrico, P.F.; Šimek, M.; Morano, M.; De Miccolis Angelini, R.M.D.; Minafra, A.; Trotti, P.; Ambrico, M.; Prukner, V.; Faretra, F. Reduction of Microbial Contamination and Improvement of Germination of Sweet Basil (Ocimum basilicum L.) Seeds via Surface Dielectric Barrier Discharge. J. Phys. D Appl. Phys. 2017, 50, 305401. [Google Scholar] [CrossRef]
- Kim, J.W.; Puligundla, P.; Mok, C. Effect of Corona Discharge Plasma Jet on Surface-Borne Microorganisms and Sprouting of Broccoli Seeds. J. Sci. Food Agric. 2017, 97, 128–134. [Google Scholar] [CrossRef]
- Khamsen, N.; Onwimol, D.; Teerakawanich, N.; Dechanupaprittha, S.; Kanokbannakorn, W.; Hongesombut, K.; Srisonphan, S. Rice (Oryza sativa L.) Seed Sterilization and Germination Enhancement via Atmospheric Hybrid Nonthermal Discharge Plasma. ACS Appl. Mater. Interfaces 2016, 8, 19268–19275. [Google Scholar] [CrossRef]
- Brasoveanu, M.; Nemţanu, M.; Carmen, S.-B.; Karaca, G.; Erper, İ. Effect of Glow Discharge Plasma on Germination and Fungal Load of Some Cereal Seeds. Rom. Rep. Phys. 2015, 67, 617–624. [Google Scholar]
- Pérez Pizá, M.C.; Prevosto, L.; Zilli, C.; Cejas, E.; Kelly, H.; Balestrasse, K. Effects of Non–Thermal Plasmas on Seed-Borne Diaporthe/Phomopsis Complex and Germination Parameters of Soybean Seeds. Innov. Food Sci. Emerg. Technol. 2018, 49, 82–91. [Google Scholar] [CrossRef]
- Sera, B.; Zahoranova, A.; Bujdakova, H.; Sery, M. Disinfection from Pine Seeds Contaminated with Fusarium circinatum Nirenberg & O’Donnell Using Non-Thermal Plasma Treatment. Rom. Rep. Phys. 2019, 71, 701. [Google Scholar]
- Ochi, A.; Konishi, H.; Ando, S.; Sato, K.; Yokoyama, K.; Tsushima, S.; Yoshida, S.; Morikawa, T.; Kaneko, T.; Takahashi, H. Management of Bakanae and Bacterial Seedling Blight Diseases in Nurseries by Irradiating Rice Seeds with Atmospheric Plasma. Plant Pathol. 2017, 66, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.H.; Pengkit, A.; Choi, K.; Jeon, S.S.; Choi, H.W.; Shin, D.B.; Choi, E.H.; Uhm, H.S.; Park, G. Differential Inactivation of Fungal Spores in Water and on Seeds by Ozone and Arc Discharge Plasma. PLoS ONE 2015, 10, e0139263. [Google Scholar] [CrossRef]
- Jo, Y.K.; Cho, J.; Tsai, T.C.; Staack, D.; Kang, M.H.; Roh, J.H.; Shin, D.B.; Cromwell, W.; Gross, D. A Non-Thermal Plasma Seed Treatment Method for Management of a Seedborne Fungal Pathogen on Rice Seed. Crop Sci. 2014, 54, 796–803. [Google Scholar] [CrossRef]
- Świecimska, M.; Tulik, M.; Šerá, B.; Golińska, P.; Tomeková, J.; Medvecká, V.; Bujdáková, H.; Oszako, T.; Zahoranová, A.; Šerý, M. Non-Thermal Plasma Can Be Used in Disinfection of Scots Pine (Pinus sylvestris L.) Seeds Infected with Fusarium oxysporum. Forests 2020, 11, 837. [Google Scholar] [CrossRef]
- Los, A.; Ziuzina, D.; Akkermans, S.; Boehm, D.; Cullen, P.J.; Van Impe, J.; Bourke, P. Improving Microbiological Safety and Quality Characteristics of Wheat and Barley by High Voltage Atmospheric Cold Plasma Closed Processing. Food Res. Int. 2018, 106, 509–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishioka, T.; Takai, Y.; Kawaradani, M.; Okada, K.; Tanimoto, H.; Misawa, T.; Kusakari, S. Seed Disinfection Effect of Atmospheric Pressure Plasma and Low Pressure Plasma on Rhizoctonia solani. Biocontrol Sci. 2014, 19, 99–102. [Google Scholar] [CrossRef] [Green Version]
- Sen, Y.; Onal-Ulusoy, B.; Mutlu, M. Aspergillus Decontamination in Hazelnuts: Evaluation of Atmospheric and Low-Pressure Plasma Technology. Innov. Food Sci. Emerg. Technol. 2019, 54, 235–242. [Google Scholar] [CrossRef]
- Dasan, B.G.; Boyaci, I.H.; Mutlu, M. Nonthermal Plasma Treatment of Aspergillus spp. Spores on Hazelnuts in an Atmospheric Pressure Fluidized Bed Plasma System: Impact of Process Parameters and Surveillance of the Residual Viability of Spores. J. Food Eng. 2017, 196, 139–149. [Google Scholar] [CrossRef]
- Dasan, B.G.; Boyaci, I.H.; Mutlu, M. Inactivation of Aflatoxigenic Fungi (Aspergillus spp.) on Granular Food Model, Maize, in an Atmospheric Pressure Fluidized Bed Plasma System. Food Control 2016, 70, 1–8. [Google Scholar] [CrossRef]
- Dasan, B.G.; Mutlu, M.; Boyaci, I.H. Decontamination of Aspergillus flavus and Aspergillus parasiticus Spores on Hazelnuts via Atmospheric Pressure Fluidized Bed Plasma Reactor. Int. J. Food Microbiol. 2016, 216, 50–59. [Google Scholar] [CrossRef]
- Wiktor, A.; Hrycak, B.; Jasiński, M.; Rybak, K.; Kieliszek, M.; Kraśniewska, K.; Witrowa-Rajchert, D. Impact of Atmospheric Pressure Microwave Plasma Treatment on Quality of Selected Spices. Appl. Sci. 2020, 10, 6815. [Google Scholar] [CrossRef]
- Ouf, S.A.; Basher, A.H.; Mohamed, A.A. Inhibitory Effect of Double Atmospheric Pressure Argon Cold Plasma on Spores and Mycotoxin Production of Aspergillus niger Contaminating Date Palm Fruits. J. Sci. Food Agric. 2015, 95, 3204–3210. [Google Scholar] [CrossRef]
- Ouf, S.A.; Mohamed, A.-A.H.; El-Sayed, W.S. Fungal Decontamination of Fleshy Fruit Water Washes by Double Atmospheric Pressure Cold Plasma. C.L.E.A.N. Soil Air Water 2016, 44, 134–142. [Google Scholar] [CrossRef]
- Hayashi, N.; Yagyu, Y.; Yonesu, A.; Shiratani, M. Sterilization Characteristics of the Surfaces of Agricultural Products Using Active Oxygen Species Generated by Atmospheric Plasma and UV Light. Jpn. J. Appl. Phys. 2014, 53, 05FR03. [Google Scholar] [CrossRef]
- Hu, X.; Sun, H.; Yang, X.; Cui, D.; Wang, Y.; Zhuang, J.; Wnag, X.; Ma, R.; Jiao, Z. Potential Use of Atmospheric Cold Plasma for Postharvest Preservation of Blueberries. Postharvest Biol. Technol. 2021, 179, 111564. [Google Scholar] [CrossRef]
- Phan, K.T.K.; Phan, H.T.; Brennan, C.S.; Regenstein, J.M.; Jantanasakulwong, K.; Boonyawan, D.; Phimolsiripol, Y. Gliding Arc Discharge Non-Thermal Plasma for Retardation of Mango Anthracnose. LWT 2019, 105, 142–148. [Google Scholar] [CrossRef]
- Dong, X.Y.; Yang, Y.L. A Novel Approach to Enhance Blueberry Quality During Storage Using Cold Plasma at Atmospheric Air Pressure. Food Bioprocess Technol. 2019, 12, 1409–1421. [Google Scholar] [CrossRef]
- Xiang, Q.; Liu, X.; Liu, S.; Ma, Y.; Xu, C.; Bai, Y. Effect of Plasma-Activated Water on Microbial Quality and Physicochemical Characteristics of Mung Bean Sprouts. Innov. Food Sci. Emerg. Technol. 2019, 52, 49–56. [Google Scholar] [CrossRef]
- Puligundla, P.; Lee, T.; Mok, C. Effect of Intermittent Corona Discharge Plasma Treatment for Improving Microbial Quality and Shelf Life of Kumquat (Citrus japonica) Fruits. LWT 2018, 91, 8–13. [Google Scholar] [CrossRef]
- Xu, Y.; Tian, Y.; Ma, R.; Liu, Q.; Zhang, J. Effect of Plasma Activated Water on the Postharvest Quality of Button Mushrooms, Agaricus bisporus. Food Chem. 2016, 197, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, A.; Niemira, B.A.; Gurtler, J.B.; Fan, X.; Sites, J.; Boyd, G.; Chen, H. Atmospheric Cold Plasma Inactivation of Aerobic Microorganisms on Blueberries and Effects on Quality Attributes. Food Microbiol. 2015, 46, 479–484. [Google Scholar] [CrossRef]
- Liu, C.-M.; Nishida, Y.; Iwasaki, K.; Ting, K. Prolonged Preservation and Sterilization of Fresh Plants in Controlled Environments Using High-Field Plasma. IEEE Trans. Plasma Sci. 2011, 39, 717–724. [Google Scholar] [CrossRef]
- Go, S.-M.; Park, M.-R.; Kim, H.-S.; Choi, W.S.; Jeong, R.-D. Antifungal Effect of Non-Thermal Atmospheric Plasma and Its Application for Control of Postharvest Fusarium oxysporum Decay of Paprika. Food Control 2019, 98, 245–252. [Google Scholar] [CrossRef]
- Yagyu, Y.; Hatayama, Y.; Hayashi, N.; Mishima, T.; Nishioka, T.; Sakudo, A.; Ihara, T.; Ohshima, T.; Kawasaki, H.; Suda, Y. Direct Plasma Disinfection of Green Mold Spore on Citrus by Atmospheric Pressure Dielectric Barrier Discharge for Agricultural Applications. Trans. Mat. Res. Soc. Japan 2016, 41, 127–130. [Google Scholar] [CrossRef] [Green Version]
- Won, M.Y.; Lee, S.J.; Min, S.C. Mandarin Preservation by Microwave-Powered Cold Plasma Treatment. Innov. Food Sci. Emerg. Technol. 2017, 39, 25–32. [Google Scholar] [CrossRef]
- Sakudo, A.; Yagyu, Y. Application of a Roller Conveyor Type Plasma Disinfection Device with Fungus-Contaminated Citrus Fruits. AMB Express 2021, 11, 16. [Google Scholar] [CrossRef]
- Jerushalmi, S.; Maymon, M.; Dombrovsky, A.; Freeman, S. Effects of Cold Plasma, Gamma and e-Beam Irradiations on Reduction of Fungal Colony Forming Unit Levels in Medical Cannabis Inflorescences. J. Cannabis Res. 2020, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Liu, D.; Zhou, R.; Song, Y.; Sun, Y.; Zhang, Q.; Niu, J.; Fan, H.; Yang, S.-Z. Atmospheric Cold Plasma Jet for Plant Disease Treatment. Appl. Phys. Lett. 2014, 104, 043702. [Google Scholar] [CrossRef]
- Kim, J.E.; Oh, Y.J.; Won, M.Y.; Lee, K.S.; Min, S.C. Microbial Decontamination of Onion Powder Using Microwave-Powered Cold Plasma Treatments. Food Microbiol. 2017, 62, 112–123. [Google Scholar] [CrossRef]
- Yong, H.I.; Lee, H.; Park, S.; Park, J.; Choe, W.; Jung, S.; Jo, C. Flexible Thin-Layer Plasma Inactivation of Bacteria and Mold Survival in Beef Jerky Packaging and Its Effects on the Meat’s Physicochemical Properties. Meat Sci. 2017, 123, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Sohbatzadeh, F.; Mirzanejhad, S.; Shokri, H.; Nikpour, M. Inactivation of Aspergillus flavus Spores in a Sealed Package by Cold Plasma Streamers. J. Theor. Appl. Phys. 2016, 10, 99–106. [Google Scholar] [CrossRef]
- Kim, J.E.; Lee, D.U.; Min, S.C. Microbial Decontamination of Red Pepper Powder by Cold Plasma. Food Microbiol. 2014, 38, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Suhem, K.; Matan, N.; Nisoa, M.; Matan, N. Inhibition of Aspergillus flavus on Agar Media and Brown Rice Cereal Bars Using Cold Atmospheric Plasma Treatment. Int. J. Food Microbiol. 2013, 161, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.I.; Farrokhi, N.; Shokri, K.; Khani, M.R.; Shokri, B. Cold Low Pressure O2 Plasma Treatment of Crocus sativus: An Efficient Way to Eliminate Toxicogenic Fungi with Minor Effect on Molecular and Cellular Properties of Saffron. Food Chem. 2018, 257, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Starek, A.; Sagan, A.; Andrejko, D.; Chudzik, B.; Kobus, Z.; Kwiatkowski, M.; Terebun, P.; Pawłat, J. Possibility to Extend the Shelf Life of NFC Tomato Juice Using Cold Atmospheric Pressure Plasma. Sci. Rep. 2020, 10, 20959. [Google Scholar] [CrossRef]
- Park, S.Y.; Ha, S.-D. Application of Cold Oxygen Plasma for the Reduction of Cladosporium cladosporioides and Penicillium citrinum on the Surface of Dried Filefish (Stephanolepis cirrhifer) Fillets. Int. J. Food Sci. Technol. 2015, 50, 966–973. [Google Scholar] [CrossRef]
- Choi, E.J.; Park, H.W.; Kim, S.B.; Ryu, S.; Lim, J.; Hong, E.J.; Byeon, Y.S.; Chun, H.H. Sequential Application of Plasma-Activated Water and Mild Heating Improves Microbiological Quality of Ready-to-Use Shredded Salted Kimchi Cabbage (Brassica pekinensis L.). Food Control 2019, 98, 501–509. [Google Scholar] [CrossRef]
- Siciliano, I.; Spadaro, D.; Prelle, A.; Vallauri, D.; Cavallero, M.C.; Garibaldi, A.; Gullino, M.L. Use of Cold Atmospheric Plasma to Detoxify Hazelnuts from Aflatoxins. Toxins 2016, 8, 125. [Google Scholar] [CrossRef]
- Sen, Y.; Onal-Ulusoy, B.; Mutlu, M. Detoxification of Hazelnuts by Different Cold Plasmas and Gamma Irradiation Treatments. Innov. Food Sci. Emerg. Technol. 2019, 54, 252–259. [Google Scholar] [CrossRef]
- Puligundla, P.; Lee, T.; Mok, C. Effect of Corona Discharge Plasma Jet Treatment on the Degradation of Aflatoxin B1 on Glass Slides and in Spiked Food Commodities. LWT 2020, 124, 108333. [Google Scholar] [CrossRef]
- Hojnik, N.; Modic, M.; Žigon, D.; Kovač, J.; Jurov, A.; Dickenson, A.; Walsh, J.L.; Cvelbar, U. Cold Atmospheric Pressure Plasma-Assisted Removal of Aflatoxin B1 from Contaminated Corn Kernels. Plasma Process. Polym. 2021, 18, 2000163. [Google Scholar] [CrossRef]
- Makari, M.; Hojjati, M.; Shahbazi, S.; Askari, H. Elimination of Aspergillus flavus from Pistachio Nuts with Dielectric Barrier Discharge (DBD) Cold Plasma and Its Impacts on Biochemical Indices. J. Food Qual. 2021, 2021, 9968711. [Google Scholar] [CrossRef]
- Ten Bosch, L.; Pfohl, K.; Avramidis, G.; Wieneke, S.; Viöl, W.; Karlovsky, P. Plasma-Based Degradation of Mycotoxins Produced by Fusarium, Aspergillus and Alternaria species. Toxins 2017, 9, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulson, J.M.; Liveris, D.; Derkatch, I.; Friedman, G.; Geliebter, J.; Park, S.; Singh, S.; Zemel, M.; Tiwari, R.K. Non-Thermal Atmospheric Plasma Treatment of Onychomycosis in an In Vitro Human Nail Model. Mycoses 2020, 63, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Lux, J.; Dobiáš, R.; Kuklová, I.; Litvik, R.; Scholtz, V.; Soušková, H.; Khun, J.; Mrázek, J.; Kantorová, M.; Jaworská, P.; et al. Inactivation of Dermatophytes Causing Onychomycosis and Its Therapy Using Non-Thermal Plasma. J. Fungi 2020, 6, 214. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.; Roe, J.; Grammer, T.C.; Graves, D.B. Plasma Treatment of Onychomycosis. Plasma Process. Polym. 2016, 13, 588–597. [Google Scholar] [CrossRef] [Green Version]
- Scholtz, V.; Soušková, H.; Hubka, V.; Švarcová, M.; Julák, J. Inactivation of Human Pathogenic Dermatophytes by Non-Thermal Plasma. J. Microbiol. Methods 2015, 119, 53–58. [Google Scholar] [CrossRef]
- Sun, P.; Sun, Y.; Wu, H.; Zhu, W.; Lopez, J.L.; Liu, W.; Zhang, J.; Li, R.; Fang, J. Atmospheric Pressure Cold Plasma as an Antifungal Therapy. Appl. Phys. Lett. 2011, 98, 021501. [Google Scholar] [CrossRef] [Green Version]
- Ouf, S.A.; El-Adly, A.A.; Mohamed, A.H. Inhibitory Effect of Silver Nanoparticles Mediated by Atmospheric Pressure Air Cold Plasma Jet Against Dermatophyte Fungi. J. Med. Microbiol. 2015, 64, 1151–1161. [Google Scholar] [CrossRef] [Green Version]
- Scholtz, V.; Soušková, H.; Švarcová, M.; Kříha, V.; Živná, H.; Julák, J. Inactivation of Dermatophyte Infection by Nonthermal Plasma on Animal Model. Med. Mycol. 2017, 55, 422–428. [Google Scholar] [CrossRef]
- Borges, A.C.; Lima, G.M.G.; Nishime, T.M.C.; Gontijo, A.V.L.; Kostov, K.G.; Koga-Ito, C.Y. Amplitude-Modulated Cold Atmospheric Pressure Plasma Jet for Treatment of Oral Candidiasis: In Vivo Study. PLoS ONE 2018, 13, e0199832. [Google Scholar] [CrossRef]
- Daeschlein, G.; Scholz, S.; von Woedtke, T.; Niggemeier, M.; Kindel, E.; Brandenburg, R.; Weltmann, K.D.; Junger, M. In Vitro Killing of Clinical Fungal Strains by Low-Temperature Atmospheric-Pressure Plasma Jet. IEEE Trans. Plasma Sci. 2011, 39, 815–821. [Google Scholar] [CrossRef]
- Lee, O.J.; Ju, H.W.; Khang, G.; Sun, P.P.; Rivera, J.; Cho, J.H.; Park, S.J.; Eden, J.G.; Park, C.H. An Experimental Burn Wound-Healing Study of Non-Thermal Atmospheric Pressure Microplasma Jet Arrays. J. Tissue Eng. Regen. Med. 2016, 10, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Hong, Y.; Park, J.; Lee, S.; Choi, E. A Novel Approach to Inactivate the Clinical Isolates of Trichophyton mentagrophytes and Trichophyton rubrum by Using Non-Thermal Plasma. J. Microb. Biochem. Technol. 2014, 6, 314–319. [Google Scholar]
- Han, L.; Patil, S.; Keener, K.M.; Cullen, P.J.; Bourke, P. Bacterial Inactivation by High-Voltage Atmospheric Cold Plasma: Influence of Process Parameters and Effects on Cell Leakage and DNA. J. Appl. Microbiol. 2014, 116, 784–794. [Google Scholar] [CrossRef] [Green Version]
- Boch, T.; Tennert, C.; Vach, K.; Al-Ahmad, A.; Hellwig, E.; Polydorou, O. Effect of Gaseous Ozone on Enterococcus faecalis Biofilm–an In Vitro Study. Clin. Oral Investig. 2016, 20, 1733–1739. [Google Scholar] [CrossRef]
- Thompson, C.M.; Tischler, A.H.; Tarnowski, D.A.; Mandel, M.J.; Visick, K.L. Nitric Oxide Inhibits Biofilm Formation by Vibrio fischeri via the Nitric Oxide Sensor HnoX. Mol. Microbiol. 2019, 111, 187–203. [Google Scholar] [CrossRef] [Green Version]
- Mravlje, J.; Regvar, M.; Starič, P.; Mozetič, M.; Vogel-Mikuš, K. Cold Plasma Affects Germination and Fungal Community Structure of Buckwheat Seeds. Plants 2021, 10, 851. [Google Scholar] [CrossRef] [PubMed]
- Hojnik, N.; Modic, M.; Tavčar-Kalcher, G.; Babič, J.; Walsh, J.L.; Cvelbar, U. Mycotoxin Decontamination Efficacy of Atmospheric Pressure Air Plasma. Toxins 2019, 11, 219. [Google Scholar] [CrossRef] [Green Version]
- Moisan, M.; Barbeau, J.; Moreau, S.; Pelletier, J.; Tabrizian, M.; Yahia, L.H. Low-Temperature Sterilization Using Gas Plasmas: A Review of the Experiments and an Analysis of the Inactivation Mechanisms. Int. J. Pharm. 2001, 226, 1–21. [Google Scholar] [CrossRef]
- Hertwig, C.; Meneses, N.; Mathys, A. Cold Atmospheric Pressure Plasma and Low Energy Electron Beam as Alternative Nonthermal Decontamination Technologies for Dry Food Surfaces: A Review. Trends Food Sci. Technol. 2018, 77, 131–142. [Google Scholar] [CrossRef]
- Liao, X.; Liu, D.; Xiang, Q.; Ahn, J.; Chen, S.; Ye, X.; Ding, T. Inactivation Mechanisms of Non-Thermal Plasma on Microbes: A Review. Food Control 2017, 75, 83–91. [Google Scholar] [CrossRef]
- Scholtz, V.; Šerá, B.; Khun, J.; Šerý, M.; Julák, J. Effects of Nonthermal Plasma on Wheat Grains and Products. J. Food Qual. 2019, 2019, 7917825. [Google Scholar] [CrossRef] [Green Version]
- Gow, N.A.R.; Netea, M.G. Medical Mycology and Fungal Immunology: New Research Perspectives Addressing a Major World Health Challenge. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köhler, J.R.; Casadevall, A.; Perfect, J. The Spectrum of Fungi That Infects Humans. Cold Spring Harb. Perspect. Med. 2014, 5, a019273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.-M.; Nishida, Y.; Iwasaki, K.; Sung, W.; Wu, F.-P. Characteristics of DC or Pulsed-Type High-Electric Field Plasma and Its Application to Air Cleaning System. IEEE Trans. Plasma Sci. 2018, 47, 1121–1128. [Google Scholar] [CrossRef]
- Bernhardt, T.; Semmler, M.L.; Schäfer, M.; Bekeschus, S.; Emmert, S.; Boeckmann, L. Plasma Medicine: Applications of Cold Atmospheric Pressure Plasma in Dermatology. Oxid. Med. Cell. Longev. 2019, 2019, 3873928. [Google Scholar] [CrossRef] [Green Version]
- Borges, A.C.; Kostov, K.G.; Pessoa, R.S.; De Abreu, G.M.A.; Lima, G.M.G.; Figueira, L.W.; Koga-Ito, C.Y. Applications of Cold Atmospheric Pressure Plasma in Dentistry. Appl. Sci. 2021, 11, 1975. [Google Scholar] [CrossRef]
- Heinlin, J.; Isbary, G.; Stolz, W.; Morfill, G.; Landthaler, M.; Shimizu, T.; Steffes, B.; Nosenko, T.; Zimmermann, J.L.; Karrer, S. Plasma Applications in Medicine with a Special Focus on Dermatology. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 1–11. [Google Scholar] [CrossRef]
- von Woedtke, T.; Emmert, S.; Metelmann, H.; Rupf, S.; Weltmann, K. Perspectives on Cold Atmospheric Plasma (CAP) Applications in Medicine. Phys. Plasmas 2020, 27, 070601. [Google Scholar] [CrossRef]
- Ranieri, P.; Sponsel, N.; Kizer, J.; Rojas-Pierce, M.; Hernández, R.; Gatiboni, L.; Grunden, A.; Stapelmann, K. Plasma Agriculture: Review from the Perspective of the Plant and Its Ecosystem. Plasma Process. Polym. 2021, 18, 2000162. [Google Scholar] [CrossRef]
- Koskey, G.; Mburu, S.W.; Awino, R.; Njeru, E.M.; Maingi, J.M. Potential Use of Beneficial Microorganisms for Soil Amelioration, Phytopathogen Biocontrol, and Sustainable Crop Production in Smallholder Agroecosystems. Front. Sustain. Food Syst. 2021, 5, 130. [Google Scholar] [CrossRef]
- Marco, M.L. Defining How Microorganisms Benefit Human Health. Microb. Biotechnol. 2021, 14, 35–40. [Google Scholar] [CrossRef]
- Veerana, M.; Lim, J.S.; Choi, E.H.; Park, G. Aspergillus oryzae Spore Germination Is Enhanced by Non-Thermal Atmospheric Pressure Plasma. Sci. Rep. 2019, 9, 11184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veerana, M.; Choi, E.H.; Park, G. Influence of Non-Thermal Atmospheric Pressure Plasma Jet on Extracellular Activity of α-Amylase in Aspergillus oryzae. Appl. Sci. 2021, 11, 691. [Google Scholar] [CrossRef]
- Hu, Z.C.; Li, W.J.; Zou, S.P.; Niu, K.; Zheng, Y.G. Mutagenesis of Echinocandin B Overproducing Aspergillus nidulans Capable of Using Starch as Main Carbon Source. Prep. Biochem. Biotechnol. 2020, 50, 745–752. [Google Scholar] [CrossRef]
- Shi, F.; Tan, J.; Chu, J.; Wang, Y.; Zhuang, Y.; Zhang, S. A Qualitative and Quantitative High-Throughput Assay for Screening of Gluconate High-Yield Strains by Aspergillus niger. J. Microbiol. Methods 2015, 109, 134–139. [Google Scholar] [CrossRef]
- Zhu, X.; Arman, B.; Chu, J.; Wang, Y.; Zhuang, Y. Development of a Method for Efficient Cost-Effective Screening of Aspergillus niger Mutants Having Increased Production of Glucoamylase. Biotechnol. Lett. 2017, 39, 739–744. [Google Scholar] [CrossRef]
- Feng, W.; Liang, J.; Wang, B.; Chen, J. Improvement of Kojic Acid Production in Aspergillus oryzae AR-47 Mutant Strain by Combined Mutagenesis. Bioprocess Biosyst. Eng. 2019, 42, 753–761. [Google Scholar] [CrossRef]
- Shu, L.; Si, X.; Yang, X.; Ma, W.; Sun, J.; Zhang, J.; Xue, X.; Wang, D.; Gao, Q. Enhancement of Acid Protease Activity of Aspergillus oryzae Using Atmospheric and Room Temperature Plasma. Front. Microbiol. 2020, 11, 1418. [Google Scholar] [CrossRef]
- Li, X.; Zheng, K.; Lai, C.; Ouyang, J.; Yong, Q. Improved Itaconic Acid Production from Undetoxified Enzymatic Hydrolysate of Steam-Exploded Corn Stover Using an Aspergillus terreus Mutant Generated by Atmospheric and Room Temperature Plasma. BioResources 2016, 11, 9047–9058. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, T.; Zuo, H.; Xiao, S.; Guo, M.; Jiang, M.; Li, Z.; Li, Y.; Zou, X. A Novel Rhodamine-Based Fluorescent pH Probe for High-Throughput Screening of High-Yield Polymalic Acid Strains from Random Mutant Libraries. RSC Adv. 2016, 6, 94756–94762. [Google Scholar] [CrossRef]
- Qiang, W.; Ling-ran, F.; Luo, W.; Han-guang, L.; Lin, W.; Ya, Z.; Xiao-Bin, Y. Mutation Breeding of Lycopene-Producing Strain Blakeslea trispora by a Novel Atmospheric and Room Temperature Plasma (ARTP). Appl. Biochem. Biotechnol. 2014, 174, 452–460. [Google Scholar] [CrossRef]
- Luo, Z.; Zeng, W.; Du, G.; Liu, S.; Fang, F.; Zhou, J.; Chen, J. A High-Throughput Screening Procedure for Enhancing Pyruvate Production in Candida Glabrata by Random Mutagenesis. Bioprocess Biosyst. Eng. 2017, 40, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Liu, S.; Du, G.; Zhou, J.; Chen, J. Identification of a Polysaccharide Produced by the Pyruvate Overproducer Candida Glabrata CCTCC M202019. Appl. Microbiol. Biotechnol. 2017, 101, 4447–4458. [Google Scholar] [CrossRef]
- Zheng, S.; Jiang, B.; Zhang, T.; Chen, J. Combined Mutagenesis and Metabolic Regulation to Enhance d-Arabitol Production from Candida parapsilosis. J. Ind. Microbiol. Biotechnol. 2020, 47, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Qin, J.; Dai, Y.; Mu, W.; Zhang, T. Atmospheric and Room Temperature Plasma (ARTP) Mutagenesis Enables Xylitol Over-Production with Yeast Candida tropicalis. J. Biotechnol. 2019, 296, 7–13. [Google Scholar] [CrossRef]
- Huang, W.W.; Ge, X.Y.; Huang, Y.; Chai, X.T.; Zhang, L.; Zhang, Y.X.; Deng, L.N.; Liu, C.Q.; Xu, H.; Gao, J. High-Yield Strain of Fusidic Acid Obtained by Atmospheric and Room Temperature Plasma Mutagenesis and the Transcriptional Changes Involved in Improving Its Production in Fungus Fusidium coccineum. J. Appl. Microbiol. 2021, 130, 405–415. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, Q.; Zhang, Q.; He, H.; Chen, Z.; Zhao, Y.; Wei, D.; Kong, M.; Huang, Q. Improved Production of Polysaccharides in Ganoderma lingzhi Mycelia by Plasma Mutagenesis and Rapid Screening of Mutated Strains Through Infrared Spectroscopy. PLoS ONE 2018, 13, e0204266. [Google Scholar] [CrossRef] [PubMed]
- Qin, T.; Song, P.; Wang, X.; Ji, X.; Ren, L.; Huang, H. Protoplast Mutant Selection of Glarea lozoyensis and Statistical Optimization of Medium for Pneumocandin B0 Yield-Up. Biosci. Biotechnol. Biochem. 2016, 80, 2241–2246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Wu, D.; Zhang, H.; Li, Q.; Zhang, Z.; Liu, Y.; Zhou, S.; Wang, W.; Li, Z.; Yang, Y. Effects of Atmospheric and Room Temperature Plasma (ARTP) Mutagenesis on Physicochemical Characteristics and Immune Activity In Vitro of Hericium erinaceus Polysaccharides. Molecules 2019, 24, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Liu, R.; Li, J.; Chang, M.; Liu, Y.; Jin, Q.; Wang, X. Enhanced Arachidonic Acid Production from Mortierella alpina Combining Atmospheric and Room Temperature Plasma (ARTP) and Diethyl Sulfate Treatments. Bioresour. Technol. 2015, 177, 134–140. [Google Scholar] [CrossRef]
- Gu, L.S.; Tan, M.Z.; Li, S.H.; Zhang, T.; Zhang, Q.Q.; Li, C.X.; Luo, X.M.; Feng, J.X.; Zhao, S. ARTP/EMS-Combined Multiple Mutagenesis Efficiently Improved Production of Raw Starch-Degrading Enzymes in Penicillium oxalicum and Characterization of the Enzyme-Hyperproducing Mutant. Biotechnol. Biofuels 2020, 13, 187. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Lin, Y.; Qi, X.; Wang, L.; He, P.; Wang, Q.; Ma, Y. Genome Shuffling of the Nonconventional Yeast Pichia anomala for Improved Sugar Alcohol Production. Microb. Cell Fact. 2015, 14, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, F.; Kitahara, Y.; Wang, Z.; Zhao, X.; Du, W.; Liu, D. Novel Mutant Strains of Rhodosporidium toruloides by Plasma Mutagenesis Approach and Their Tolerance for Inhibitors in Lignocellulosic Hydrolyzate. J. Chem. Technol. Biotechnol. 2014, 89, 735–742. [Google Scholar] [CrossRef]
- Kitahara, Y.; Yin, T.; Zhao, X.; Wachi, M.; Du, W.; Liu, D. Isolation of Oleaginous Yeast (Rhodosporidium toruloides) Mutants Tolerant of Sugarcane Bagasse Hydrolysate. Biosci. Biotechnol. Biochem. 2014, 78, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Zhao, X.; Kitahara, Y.; Li, T.; Ou, X.; Du, W.; Liu, D.; Huang, J. Integrative Transcriptomic and Proteomic Analysis of the Mutant Lignocellulosic Hydrolyzate-Tolerant Rhodosporidium toruloides. Eng. Life Sci. 2017, 17, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Shen, H.; Zhang, X.; Yu, X.; Wang, H.; Xiao, S.; Wang, J.; Zhao, Z.K. Combined Mutagenesis of Rhodosporidium toruloides for Improved Production of Carotenoids and Lipids. Biotechnol. Lett. 2016, 38, 1733–1738. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, D.; Yang, Q.; Wang, P. Enhancing Carotenoid Production in Rhodotorula mucilaginosa KC8 by Combining Mutation and Metabolic Engineering. Ann. Microbiol. 2017, 67, 425–431. [Google Scholar] [CrossRef]
- Liang, M.H.; Liang, Y.J.; Chai, J.Y.; Zhou, S.S.; Jiang, J.G. Reduction of Methanol in Brewed Wine by the Use of Atmospheric and Room-Temperature Plasma Method and the Combination Optimization of Malt with Different Adjuncts. J. Food Sci. 2014, 79, M2308–M2314. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Jia, H.; Zhang, L.; Wang, H.; Tang, H.; Zhang, L. Effects of GSH1 and GSH2 Gene Mutation on Glutathione Synthetases Activity of Saccharomyces cerevisiae. Protein J. 2017, 36, 270–277. [Google Scholar] [CrossRef]
- Li, T.; Chen, L.; Wu, D.; Dong, G.; Chen, W.; Zhang, H.; Yang, Y.; Wu, W. The Structural Characteristics and Biological Activities of Intracellular Polysaccharide Derived from Mutagenic Sanghuangporous sanghuang Strain. Molecules 2020, 25, 3693. [Google Scholar] [CrossRef]
- Ma, X.J.; Zhang, H.M.; Lu, X.F.; Han, J.; Zhu, H.X.; Wang, H.; Yao, R.S. Mutant Breeding of Starmerella bombicola by Atmospheric and Room-Temperature Plasma (ARTP) for Improved Production of Specific or Total Sophorolipids. Bioprocess Biosyst. Eng. 2020, 43, 1869–1883. [Google Scholar] [CrossRef]
- Zou, Z.; Zhao, Y.; Zhang, T.; Xu, J.; He, A.; Deng, Y. Efficient Isolation and Characterization of a Cellulase Hyperproducing Mutant Strain of Trichoderma reesei. J. Microbiol. Biotechnol. 2018, 28, 1473–1481. [Google Scholar] [CrossRef]
- Xu, F.; Wang, J.; Chen, S.; Qin, W.; Yu, Z.; Zhao, H.; Xing, X.; Li, H. Strain Improvement for Enhanced Production of Cellulase in Trichoderma viride. Prikl. Biokhim. Mikrobiol. 2011, 47, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Jin, H.; Li, H.; Tao, L.; Wang, J.; Lv, J.; Chen, S. Genome Shuffling of Trichoderma viride for Enhanced Cellulase Production. Ann. Microbiol. 2012, 62, 509–515. [Google Scholar] [CrossRef]
- Zeng, W.; Du, G.; Chen, J.; Li, J.; Zhou, J. A High-Throughput Screening Procedure for Enhancing α-Ketoglutaric Acid Production in Yarrowia lipolytica by Random Mutagenesis. Process Biochem. 2015, 50, 1516–1522. [Google Scholar] [CrossRef]
- Zeng, W.; Fang, F.; Liu, S.; Du, G.; Chen, J.; Zhou, J. Comparative Genomics Analysis of a Series of Yarrowia lipolytica WSH-Z06 Mutants with Varied Capacity for α-Ketoglutarate Production. J. Biotechnol. 2016, 239, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lv, J.; Xu, J.; Xia, J.; Dai, B.; Xu, X.; Xu, J. Erythritol Production by Yarrowia lipolytica Mutant strain M53 Generated Through Atmospheric and Room Temperature Plasma Mutagenesis. Food Sci. Biotechnol. 2017, 26, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Arnau, J.; Yaver, D.; Hjort, C.M. Strategies and Challenges for the Development of Industrial Enzymes Using Fungal Cell Factories. In Grand Challenges in Fungal Biotechnology; Nevalainen, H., Ed.; Springer: Cham, Switzerland, 2019; pp. 179–210. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, X.F.; Li, H.P.; Wang, L.Y.; Zhang, C.; Xing, X.H.; Bao, C.Y. Atmospheric and Room Temperature Plasma (ARTP) as a New Powerful Mutagenesis Tool. Appl. Microbiol. Biotechnol. 2014, 98, 5387–5396. [Google Scholar] [CrossRef]
- Wende, K.; Bekeschus, S.; Schmidt, A.; Jatsch, L.; Hasse, S.; Weltmann, K.D.; Masur, K.; von Woedtke, T. Risk Assessment of a Cold Argon Plasma Jet in Respect to Its Mutagenicity. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2016, 798, 48–54. [Google Scholar] [CrossRef]
- Bruggeman, P.J.; Kushner, M.J.; Locke, B.R.; Gardeniers, J.G.E.; Graham, W.G.; Graves, D.B.; Hofman-Caris, R.C.H.M.; Maric, D.; Reid, J.P.; Ceriani, E.; et al. Plasma-Liquid Interactions: A Review and Roadmap. Plasma Sources Sci. Technol. 2016, 25, 053002. [Google Scholar] [CrossRef]
- Kondeti, V.S.S.K.; Phan, C.Q.; Wende, K.; Jablonowski, H.; Gangal, U.; Granick, J.L.; Hunter, R.C.; Bruggeman, P.J. Long-Lived and Short-Lived Reactive Species Produced by a Cold Atmospheric Pressure Plasma Jet for the Inactivation of Pseudomonas aeruginosa and Staphylococcus aureus. Free Radic. Biol. Med. 2018, 124, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Gaunt, L.F.; Beggs, C.B.; Georghiou, G.E. Bactericidal Action of the Reactive Species Produced by Gas-Discharge Nonthermal Plasma at Atmospheric Pressure: A Review. IEEE Trans. Plasma Sci. 2006, 34, 1257–1269. [Google Scholar] [CrossRef]
- Graves, D.B. The Emerging Role of Reactive Oxygen and Nitrogen Species in Redox Biology and Some Implications for Plasma Applications to Medicine and Biology. J. Phys. D Appl. Phys. 2012, 45, 263001. [Google Scholar] [CrossRef]
- Xu, H.; Zhu, Y.; Du, M.; Wang, Y.; Ju, S.; Ma, R.; Jiao, Z. Subcellular Mechanism of Microbial Inactivation During Water Disinfection by Cold Atmospheric-Pressure Plasma. Water Res. 2021, 188, 116513. [Google Scholar] [CrossRef] [PubMed]
- Dobrynin, D.; Fridman, G.; Friedman, G.; Fridman, A. Physical and Biological Mechanisms of Direct Plasma Interaction with Living Tissue. New J. Phys. 2009, 11, 115020. [Google Scholar] [CrossRef]
- Kobzev, E.N.; Kireev, G.V.; Rakitskii, Y.A.; Martovetskaya, I.I.; Chugunov, V.A.; Kholodenko, V.P.; Khramov, M.V.; Akishev, Y.S.; Trushkin, N.I.; Grushin, M.E. Effect of Cold Plasma on the E. coli Cell Wall and Plasma Membrane. Appl. Biochem. Microbiol. 2013, 49, 144–149. [Google Scholar] [CrossRef]



{kind=link}
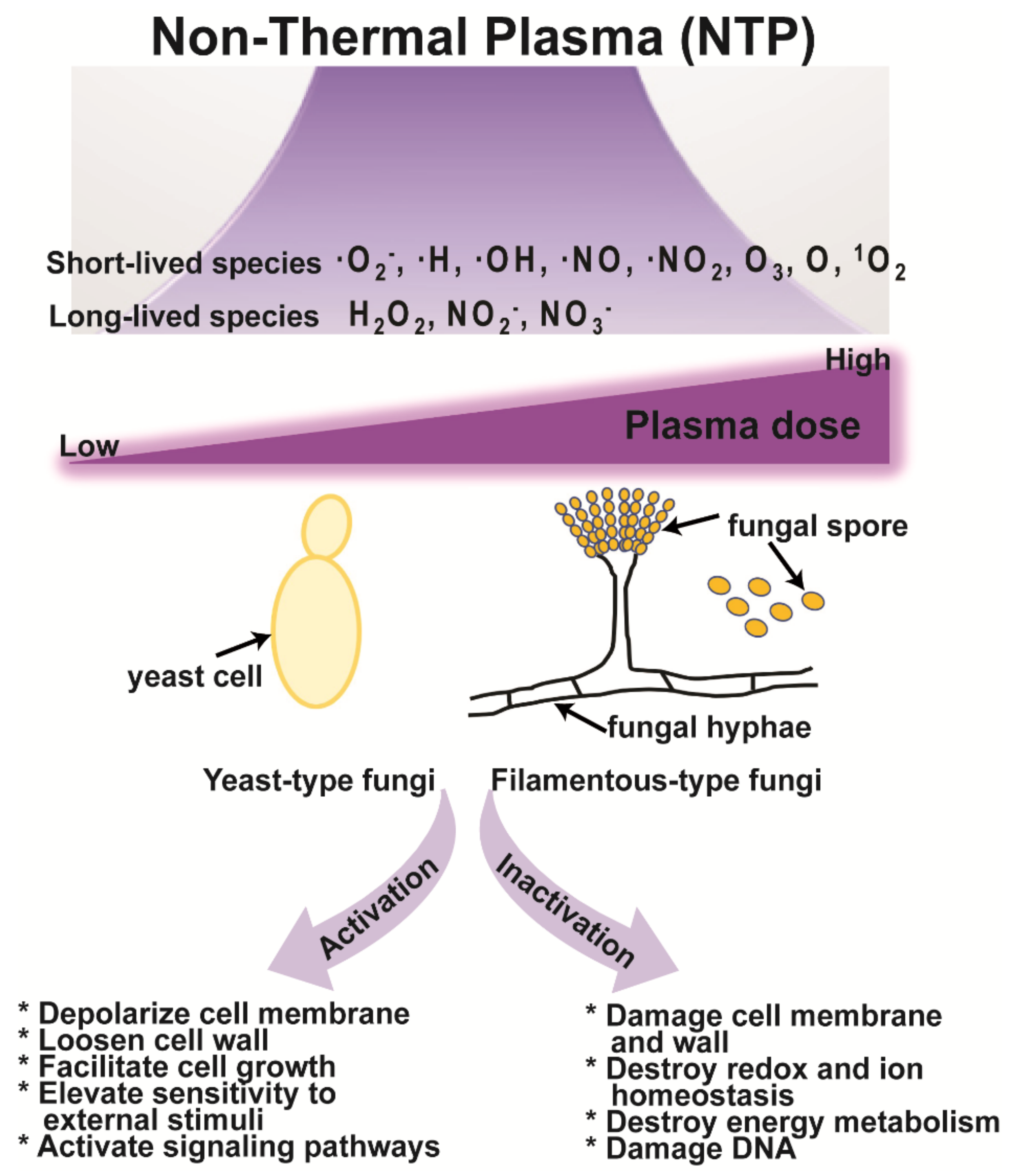{kind=link}
| Application | Target Fungi (Materials) | Plasma Source (Treatment Parameters) | Effects | Ref. |
|---|---|---|---|---|
| Suspension of fungal spores/cells and fungal biofilm | ||||
| Inactivation and inhibition of growth | Alternaria sp. Aspergillus oryzae Byssochlamys nivea Cladosporium sphaerospermum | Corona discharge plasma (9 kV, 300 µA, filtered air) Dielectric barrier discharge plasma (high-voltage, air) | C.sphaerospermum, A. sp. and B. nivea, A. oryzae in order of sensitivity to plasma Spore inactivation time: within 10–40 min | [27] |
| Ascochyta pinodella Fusarium culmorum | Dielectric barrier discharge plasma (20 kV, ~560 W, air) | Complete inhibition of hyphal growth of both fungi after 360 s exposure | [13] | |
| Aspergillus brasiliensis | Plasma activated water (PAW): treated with plasma jet (1.24 kV, 2.96 A, 3.9 W, air) | 15% reduction in spore viability after 30 min in PAW | [28] | |
| Aspergillus flavus | RF plasma jet (80–800 kHz RF power, 100 W, mixture of argon Ar and oxygen O2) | 100% inhibition of growth 48 h after 10 min treatment at 20 W | [29] | |
| Gaseous plasma and plasma-activated aqueous broth (PAB) Plasma source: surface barrier discharge plasma (5–15 kV at 40 kHz, 0.79, 1.24, 1.62 W/cm2, air) | Gaseous plasma treatment: over four log reduction in spore viability after 240 and 480 s treatments under three power conditions PAB treatment: no significant reduction in spore viability | [30] | ||
| Aspergillus niger Penicillium citrinum | Dielectric barrier discharge plasma (3 kV at 230 Hz on dielectric ceramic electrode, −4 kV on needle electrode, helium He) | Maximum 98–99.9% deactivation of fungal spores after 5 h treatment | [31] | |
| Microwave plasma jet (2.45 GHz, 1 kW, Ar) | 100% inactivation of fungal spores after 1 s treatment | [32] | ||
| Aspergillus niger Cladosporium cladosporioides Penicillium citrinum Chaetomium sp. | Microwave plasma jet (2.45 GHz, 1 kW, Ar) | 100% inactivation of fungal spores after 1 s treatment | [33] | |
| Aspergillus ochraceus Penicillium expansum | Plasma jet (2.5 kV at 25 kHz, 3 mA, 4 W, Ar) | A. ochraceus: maximum 3.42 log10 CFU reduction after 5 min treatment P. expansum: maximum 3.11 log10 CFU reduction after 5 min treatment | [34] | |
| Aspergillus oryzae Cladosporium sphaerospermum Penicilium crustosum | Corona discharge plasma (9.7 kV, 400 µA, filtered air) | 99.9–100% spore inactivation after 30 min treatment Significant growth inhibition | [35] | |
| Aureobasidium pullulans | Dielectric barrier discharge plasma (9.3 kV at 11 kHz, Ar) | ~100% and ~30% inactivation of non-melanized and melanized cells after 60 min treatment Improvement of fungicidal efficacy of plasma by using FeCl2 and FeSO4 together | [36] | |
| Candida albicans | Linear microdischarge plasma jet (13.56 MHz, 17 W, He) | Changes in genome sequence, enzyme activity at sublethal dose No change in carbon assimilation and drug susceptibility at sublethal dose | [37] | |
| Plasma jet (1.8 W, He) | 20–30 mm2 inhibition zone after 3 min treatment Maximum 11 mm in diameter of inhibition zone after 3 min treatment on five fungal strains | [38,39] | ||
| Plasma activated water (PAW): treated with nanosecond pulsed DBD plasma (50 mJ per pulse, 20 kV at 1000 Hz, air) | 100% cells killed after 10–30 min incubation in 5 or 10 min-treated PAW | [40] | ||
| RF plasma jet (15 MHz RF power, 10 kV, mixture of 98% He and 2% O2) | 31–82% inhibition of growth, 40–91% reduction in ergosterol biosynthesis, 43–57% reduction in biofilm formation and activities of phospholipase and proteinase | [41] | ||
| High-voltage nanosecond pulse plasma jet (6 kV at 1.5 kHz, mixture of 99% He and 1% O2 | >99.99% inactivation of fungal cells after 30–180 s treatment | [42] | ||
| Dielectric barrier discharge plasma (10 kV at 7.1 kHz, mixture of 99% He and 1% O2) | 100% cells killed after 5 min treatment | [43] | ||
| Plasma jet (8 kV at 8 kHz, mixture of 97% He and 3% O2) | >99.9% inactivation after 3.5 min treatment in the condition of covering A small fraction of fungal cells inactivated even after 8 min treatment without covering | [14] | ||
| Dielectric barrier discharge plasma (30 kV at 60 kHz, air) | 100% inactivation of cells after 30 s treatment | [44] | ||
| Candida parapsilosis Magnusiomyces magnusii Saccharomyces cerevisiae Schizosaccharomyces pombe | Plasma jet (13 kV at 5 kHz, Ar) | Less than 10% survival of cells after 10 min treatment and 2 h incubation Maximum 0.76% survival of S. pombe cells | [45] | |
| Cladosporium fulvum | Plasma jet (5–12 kV at 5–13 kHz, mixture of 99% Ar and 1% O2) | Complete killing of fungal spores after 60 s treatment Disruption of membrane and leaking of cytoplasm DNA and protein damage | [46] | |
| Colletotrichum gloeosporioides | Plasma-activated water (PAW): treated with corona discharge plasma jet (3–4 kV at 20 kHz, mixture of 21% O2 and 79% N2 or 99.99% O2) | 96% and 56% spore inactivation after 30 min and 10 min incubation, respectively, in PAW generated using air plasma 55% and 15% spore inactivation after 30 min and 10 min incubation, respectively, in PAW generated using oxygen plasma | [47] | |
| Cordyceps bassiana | Plasma jet (22 kHz, 9 W, Ar) | 16.4% spore survival after 5 min treatment Reduction in DNA content and alteration to cell wall | [48,49] | |
| Electric shock-free plasma jet (67 kHz, air) | 0.8% spore survival after 6 min treatment Plasma generated H2O2 and NOx as major players in antifungal activity | [50] | ||
| Fusarium graminearum Fusarium oxysporum Neurospora crassa | Microwave plasma jet (2.45 GHz, 1.6 W, mixtures of 83% Ar and 17% O2, 83% Ar and 17% N2, or 83% Ar and 17% air, 100% N2) | Dramatic reduction in fungal hyphal growth when O2 is used in plasma generation Increased growth inhibition with increased power and pulse length | [51] | |
| Fusarium oxysporum f. sp. lycopersici | Dielectric barrier discharge plasma (discharge at 0.75 kV and 80 mA, 7.5 W, air or Ar) | <10% survival of fungal spores after treated in saline for 10 min and then incubated for 6 h Increase in size of lipid droplets inside spore cell and number of apoptotic spores | [52] | |
| Neurospora crassa | Plasma jet (4 kV at 22 kHz, 13 mA, Ar) | ~80% reduction in spore viability after 3 min treatment in water Less effective when treated in saline and culture media Crushed spores, cell wall damage, degradation of β-carotene | [53,54,55] | |
| Penicillium digitatum | Plasma jet (discharge at −500–−1000 V and 3–16 mA, 5 W, humid air) | Maximum 91% spore inactivation after 9 min treatment and moisture was added in working gas | [56] | |
| Plasma generated oxygen radical source (Tough Plasma; Fuji Machine MFG Co. Ltd., Chiryu, Japan) | >2 log10 CFU reduction in spore viability after 5 min treatment at distance of 10 mm | [57,58] | ||
| Microwave plasma jet (2.45 GHz, 50 W, O2) | ~2 log10 CFU reduction in spore viability after 10 min treatment Decay of electron spin resonance (ESR) signal in situ from plasma treated spores | [59] | ||
| Plasma jet (6 kV at 60 Hz, Ar) | >3 log10 CFU reduction in spore viability after 7 min treatment at distance of 10 mm | [60] | ||
| Penicillium sp. | Plasma microdischarge torch (discharge at 5–10 kV and 15 mA, 7.5–15 W, air) | Inhibition of fungal growth after treatment at distance of 3 cm Partial spore damage after plasma treatment | [61] | |
| Saccharomyces cerevisiae | Dielectric barrier discharge plasma (Ar) | ~99% reduction in cell viability after 10–15 min treatment Accumulation of oxidative stress responsive transcription factor, mitochondrial fragmentation, enhanced intracellular oxidation | [62] | |
| Plasma jet (4 kV at 22 kHz, 13 mA, Ar) | ~100% reduction in cell viability after 2 min treatment in water Less effective when treated in saline and culture media Crushed cells, increased lipid peroxidation, and DNA degradation after plasma treatment in water and saline | [63] | ||
| Dielectric barrier discharge plasma (12 kV at 20 kHz, 3.8 mA, 26 W, air) | Maximum ~2 log10 CFU reduction in cell viability after 5 min treatment Release of protein and nucleic acids, cell cycle arrest at G1 phase | [64] | ||
| Plasma microjet (0.56 kV, 30 mA, mixture of 98% He and 2% O2) | Maximum 100% reduction in cell viability after 5 min treatment | [65] | ||
| Plasma microjet (0.56 kV, 30 mA, mixture of 98% He and 2% O2) | >2 log10 CFU reduction in cell viability after 5 min treatment in water ROS and acidic pH exert a synergistic antimicrobial effect | [66] | ||
| Surface micro-discharge plasma (8 kV at 8 kHz, 0.07 W/cm2, He) | Inactivation pattern of cells is dependent on distribution and concentration of OH radical | [67] | ||
| Plasma microjet (0.56 kV, 30 mA, mixture of 98% He and 2% O2) | Plasma-generated ROS leads to the accumulation of intracellular ROS and Ca2+, which in turn cause apoptosis of yeast cells | [68] | ||
| Plasma microjet (discharge at 0.56 kV and 30 mA, mixture of 98% He and 2% O2) | Evaluated the protection effects of gene manipulation and reactive species scavengers against plasma-induced oxidative stresses: overexpression of superoxide dismutases reduces plasma oxidative stress | [69] | ||
| Trichophyton rubrum | Plasma jet (10 kV at 15 MHz, 10 W, mixture of 98% He and 2% O2) | ~91% spore inactivation, >50% reduction in fungal dry weight, and 53% inhibition in ergosterol synthesis after 3 min treatment | [70] | |
| Inhibition of biofilm formation | Aspergillus flavus | Gaseous plasma and plasma activated water (PAW) Plasma source: dielectric barrier discharge plasma (80 kV, air) | Maximum 2.2 and 0.6 log10 CFU reduction in spore viability after treatment with gaseous plasma and plasma activated water, respectively ~50% reduction in biofilm biomass after gaseous plasma treatment | [71] |
| Candida albicans | Plasma jet (15 kV at 1 kHz, mixture of 99.5% He and 0.5% O2) | Reduction from 35.6 × 102 CFU/mL to 4.6 × 102 CFU/mL after 8 min treatment of suspension Complete killing of fungal cells in biofilm after 8 min treatment | [72] | |
| Plasma jet (1.8 W, He) | 40 times reduction in filamentation Reduction in fungal adherence and biofilm viability (~1 log10 CFU reduction in cell viability within biofilm) No effect on exoenzyme production | [73] | ||
| Surface dielectric barrier microdischarge plasma (9 kV at 1 kHz, 0.02 W/cm2, air) | 3–5 log10 CFU reduction in cell viability within biofilm | [74] | ||
| Plasma jet (kINPen08; 2–6 kV at 1.7 MHz, 65 W, Ar, mixture of 99% Ar and 1% O2) | Removal of biofilm with a thickness of 10 to 20 µm within 300 s plasma treatment using mixture of Ar and O2 as working gas Insufficient removal of biofilm using Ar plasma | [75] | ||
| Plasma microjet (discharge at 0.56 kV and 30 mA, mixture of 98% He and 2% O2) | Complete removal of biofilms after 1 min treatment Severe deformation of fungal elements | [76] | ||
| Plasma jet (kINPen09; 2–6 kV at 1.82 MHz, Ar, mixture of Ar and O2) Hollow electrode dielectric barrier discharge (HDBD) plasma (37.6 kHz RF power, 9 W, 9 kV, Ar, mixture of Ar and O2) Volume dielectric barrier discharge (VDBD) plasma (10 kV at 40 kHz, 16 W, Ar) | kINPen09; maximum 1 log10 CFU reduction in cell viability within biofilm HDBD; maximum 3.3 log10 CFU reduction in cell viability within biofilm VDBD; maximum 5.2 log10 CFU reduction in cell viability within biofilm | [77] | ||
| Fungal contamination in agriculture and foods | ||||
| Disinfection of seeds | Alternaria alternata Aspergillus flavus Fusarium culmorum (maize seeds) | Diffuse coplanar surface barrier discharge plasma (80 W/cm3, air) | Reduction of 3.79 log10 CFU/g in F. culmorum after 60 s treatment Reduction of 4.21 log10 CFU/g in A. flavus and 3.22 log10 CFU/g in A. alternata after 300 s treatment Increase in seed surface wettability Enhancement of seedling growth | [78] |
| Alternaria alternata Alternaria botrytis Aspergillus brasiliensis Epicoccum nigrum Fusarium culmorum Fusarium poae Gibberella zeae Mucor hiemalis Penicillium sp. Rhizopus stolonifer Trichoderma sp. (winter wheat) | Plane-type plasma (8 kV at 0.1–83 kHz, air) | Reduction in number of fungal colonies on seeds after 10 s treatment Positive effect on seed germination and initial seedling development | [79] | |
| Aspergillus clavatus Aspergillus flavus Fusarium culmorum Fusarium nivale Trichothecium roseum (wheat) | Diffuse coplanar surface barrier discharge plasma (100 W/cm3, air) | Order of efficiency in fungal decrease after plasma treatment; F. nivale > F. culmorum > T. roseum > A. flavus > A. clavatus | [80] | |
| Aspergillus flavus Aspergillus parasiticus (groundnuts) | RF plane-type plasma (13.56 MHz RF power, 40 W and 60 W, air) | 97.9% and 99.3% reduction in CFU of A. parasiticus and A. flavus, respectively, when treated at 60 W | [81] | |
| Aspergillus niger Penicillium decumbens (lentil seeds) | Diffuse coplanar surface barrier discharge plasma (RPS400; 400 W, air) | 1.6 and 3.1 log10 CFU/g reduction for A. niger and P. decumbens, respectively, after 10 min treatment No significant effect on germination | [82] | |
| Aspergillus parasiticus (hazelnuts, peanuts, pistachio nuts) | Low-pressure plasma (20 kV at 1 kHz, 300 W, 100 or 500 mTorr, air or SF6) | 1 and 5 log10 CFU reduction after 5 min treatment with air and SF6 plasma, respectively | [83] | |
| Aspergillus sp. Penicillium sp. (black beans) | Dielectric barrier discharge plasma (8 kV, 510 W, air) | Complete fungal disinfection after treatment for at least 10 min Wrinkling on seed surface and change in cotyledon color | [84] | |
| (seeds of tomato, wheat, bean, chickpea, soybean, barley, oat, rye, lentil, corn) | Low-pressure plasma (20 kV at 1 kHz, 300 W, air or SF6 at low pressure) | Fungal decontamination below 1% of initial load 3 log10 CFU reduction after 15 min treatment with SF6 plasma No significant change in seed germination quality | [85] | |
| Cladosporium cucumerinum Didymella bryoniae Didymella licopersici (cucumber and pepper seeds) | Surface dielectric barrier discharge plasma (20 kV at 15 kHz, 400 W, air) | No presence of C. cucumerinum and 60–80% reduction in D. bryoniae spore viability on cucumber seeds after 20 s treatment 50–80% reduction in D. lycopersici spore viability on pepper seeds after 4 s treatment Improvement in seed germination | [86] | |
| Cladosporiumfulvum (tomato seeds) | Plasma jet (5–12 kV at 5–13 kHz, mixture of 99% Ar and 1% O2) | Maximum 14% reduction in seed rotting caused by fungal infection after 60 s treatment | [46] | |
| Clonostachys rossmaniae Coniochaeta fasciculat Cylindrocarpon destructans Fusarium proliferatum Humicola fuscoatra Mortierella hyalina Pyrenochaeta sp. (ginseng seeds) | Dielectric barrier discharge plasma (120 V at 60 Hz, Ar or mixture of 80% Ar and 20% O2) | 27.7% and 40% survival of fungal spores on seeds after Ar and Ar/O2 plasma treatments, respectively | [87] | |
| Contaminated fungi (Pak-Choi seeds) | Corona discharge plasma jet (20 kV at 58 kHz, 1.5 A, air) | 1.3–2.1 log10 CFU/g reduction after 3 min treatment Positive effect on seed germination after treatment for up to 2 min | [88] | |
| (sweet basil seeds) | Surface dielectric barrier discharge plasma (8.6 kV at 500 Hz, 6.5 W, air) | ~30% reduction in number of seeds naturally contaminated with fungi after 300 s treatment Improvement in growth of seedlings | [89] | |
| (broccoli seeds) | Corona discharge plasma jet (20 kV at 58 kHz, 1.5 A, air) | 1.5 log10 CFU/g reduction of natural fungal flora on seeds after 3 min treatment Positive effect on seed germination and seedling growth after treatment for up to 2 min | [90] | |
| (rice seeds) | Microcorona dielectric barrier discharge plasma (~14 kV at ~700 Hz, air) | Complete removal of fungal contamination from seeds after 1 min treatment and then incubation for 14 days Enhancement of seed germination | [91] | |
| (barley and corn seeds) | Glow discharge low pressure plasma (100 or 200 W, 15 Pa, air) | Barley: 25% reduction in fungal contamination on seeds after 20 min treatment, retardation of seed germination and no influence on seedling growth Corn: ~40% reduction in fungal contamination on seeds after 10 min treatment, no influence on seed germination and slight improvement of seedling growth | [92] | |
| Diaporthe/Phomopsis (D/P) complex (soybean seeds) | Dielectric barrier discharge plasma (65 or 85 W, 50 Hz, ~50 mA, N2 or O2) | ~49–81% disinfection of seeds Significant stimulating effects on seed germination and vigor | [93] | |
| Fusariumcircinatum (pine seeds) | Diffuse coplanar surface barrier discharge plasma (10 kV at 14 kHz, 400 W, air) | 14–100% disinfection of seeds after treatment up to 300 s Reduction in seed germination percentage | [94] | |
| Fusarium fujikuroi (rice seeds) | Plasma jet (20 kV at 10 kHz, humid air) | Reduction to 7.8% of non-treated control in the percentage of plants with disease symptoms after 10 min treatment of seeds in water No adverse effect on seed germination and seedling growth | [95] | |
| Underwater arc discharge plasma (10 kV at 12 Hz, air in water) | ~80% disinfection of seeds after 20 min treatment | [96] | ||
| Dielectric barrier discharge plasma (30 kV at 22 kHz, air) | >92% disinfection of seeds after 120 s treatment Significant reduction in disease development after 10 min treatment on seeds No adverse effect on seed germination and seedling growth | [97] | ||
| Fusarium oxysporum (Scots pine seeds) | Diffuse coplanar surface barrier discharge plasma (20 kV at 14 kHz, 400 W, air) | ~6% disinfection of seeds after 3 s treatment Slight increase in seed germination percentage | [98] | |
| Penicillium verrucosum (wheat and barley seeds) | Dielectric barrier discharge plasma (80 kV at 50 Hz, air) | Maximum 2.1 and 2.5 log10 CFU/g reduction in barley and wheat seeds, respectively, after 20 min treatment followed by incubation for 24 h No significant effect on seed germination | [99] | |
| Rhizoctonia solani (brassicaceous seeds) | Atmospheric pressure plasma (10 kV at 10 kHz, Ar) Low pressure plasma (5.5 kV at 10 kHz, 80 torr, Ar) | Atmospheric-pressure plasma: 97% reduction in fungal survival on seeds after 10 min treatment, delay of seed germination Low-pressure plasma: 81% reduction in fungal survival on seeds after 10 min treatment, no change in seed germination rate | [100] | |
| Disinfection of post-harvest vegetables and fruits | Aspergillus flavus Aspergillus parasiticus (hazelnut) | Atmospheric pressure plasma jet (25 kHz, 655 W, air or N2) Low pressure RF plasma (13.56 MHz RF power, 100 W, 0.25 mbarr, air, N2 or O2) | Atmospheric-pressure plasma: 5.5 and 5.4 log10 CFU/g reduction in A. parasiticus and A. flavus on hazelnuts, respectively, after 1.7 min treatment Low-pressure plasma: 5.6 and 4.7 log10 CFU/g reduction in A. parasiticus and A. flavus on hazelnuts, respectively, after 30 min treatment | [101] |
| (hazelnut, maize) | Fluidized bed plasma (5–10 kV at 18–25 kHz, 655 W, air or N2) | Maximum 4.09–4.19 and 4.17–4.50 log10 CFU/g reduction in A. parasiticus and A. flavus on hazelnuts, respectively, after 5 min treatment with air plasma, no or little fungal regrowth during storage for 30 days Maximum 5.20 and 5.48 log10 CFU/g reduction in A. parasiticus and A. flavus on maize, respectively, after 5 min treatment with air plasma, no fungal regrowth during storage for 30 days | [102,103,104] | |
| Aspergillus niger (black pepper, allspice berry, juniper berry) | Microwave plasma (2.45 GHz, 600 W, Ar) | Partial inactivation of A. niger Reduction in water activity Enhancement of extractability of phenolics or piperine from black pepper | [105] | |
| (date palm fruit discs) | Plasma jet (25 kV at ~25 kHz, Ar) | Complete removal of fungal spores on fruit discs after 7.5 min treatment with Ar flow 3.5 L/min | [106] | |
| Aspergillus niger Penicillium italicum (fruit washwater) | Plasma jet (25 kV at ~25 kHz, Ar) | 74.7–100% removal of fungal contamination in the washwater of cherries after 7.5 min treatment | [107] | |
| Aspergillus oryzae Penicillium digitatum (rice, lemon) | Surface dielectric barrier discharge (7–10 kV at 10 kHz, air) | ~90% and ~100% removal of fungal contamination on rice and lemon surface, respectively, after 20 min treatment | [108] | |
| Botrytis cinerea (blueberry) | Dielectric barrier surface discharge plasma (4 kV at 8 kHz, 5 W, air) | Inhibition of native microbial growth and natural decay of blueberries after plasma treatment Maximum ~40% reduction in decay incidence in blueberries inoculated with B. cinerea after 20 min treatment and 10-day storage Minor effects on blueberry quality after less than 15 min treatment but severe oxidative damage to the blueberry peels after 20 min treatment | [109] | |
| Botrytis cinerea Monilinia fructicola (cherry) | Surface dielectric barrier discharge plasma (8.6 kV at 500 Hz, 6.5 W, air) | >50% reduction in number of infected fruits after 5 min treatment in earlier days Pre-treatment of fruits by plasma before inoculation enhances the resistance to infections | [15] | |
| Colletotrichum gloeosporioides (mango) | Gliding arc discharge (discharge at 8 kV and 0.6 A, 600 W, humid Ar) | Significant inhibition of mycelium growth Significant delay in disease development in mango after 7 min treatment during storage for 12 days at 30 °C | [110] | |
| Contaminated fungi (blueberry) | Dielectric barrier discharge plasma (discharge at 36 V and 1.8 A, air) | 25.8% decrease in fungal contamination on blueberry and 5.2% blueberry decay rate after 10 min treatment during storage for 20 days | [111] | |
| (mung bean sprout) | Plasma-activated water (PAW): treated with plasma jet (5 kV at 40 kHz, 750 W, air) | 2.84 log10 CFU/g reduction in yeasts and molds on mung bean after 30 min treatment in PAW No significant change in total phenolic and flavonoid contents and sensory characteristics of mung bean | [112] | |
| (kumquat) | Corona discharge plasma jet (8 kV at 20 kHz, air) | 0.77–1.57 log10 CFU/g reduction in yeasts and molds on kumquat after 2 min treatment No significant change in taste, flavor, color, texture, and total acceptance | [113] | |
| (button mushroom) | Plasma activated water (PAW): treated with plasma jet (18 kV at 10 kHz, mixture of 98% Ar and 2% O2) | 0.5 log10 CFU reduction in fungi on mushroom during storage for over 7 days Delay in mushroom softening No significant change in color, pH, antioxidant properties | [114] | |
| (blueberry) | Plasma jet (47 kHz, 549 W, air) | 1.5–2.0 log10 CFU/g reduction in yeasts and molds on blueberries after 7 days Significant reduction in firmness and anthocyanin content after treatment for over 60 s | [115] | |
| (banana, grape) | High-field plasma system (2 kV at 500 Hz, 20–30 µA, 3–4 × 106 V/m electric field, air) | No increase in mold load on surface of fruits during storage in high-filed plasma system Lower amount of ethylene gas emitted during storage in high-field plasma system | [116] | |
| Fusarium oxysporum (paprika) | Plasma jet (28 kHz, 1000 W, air) | 50% inhibition of fungal growth on paprika after 90 s treatment No significant change in color and hardness during 14 days of storage | [117] | |
| Penicillium digitatum (citrus) | Dielectric barrier discharge plasma (10 kV at ~10 kHz, air) | ~90% and ~99% reduction in CFU number of fungal spores on citrus surface after 1 s and 3 s treatments, respectively | [118] | |
| Penicillium italium (mandarin) | Microwave plasma jet (2.45 GHz, 900 W, 500–30,000 Pa, N2) | 84% reduction in disease incidence after 10 min treatment Significant increase in total phenolic content and antioxidant activity of mandarin peel | [119] | |
| Penicillium venetum (citrus) | Roller conveyor type dielectric barrier discharge plasma (11.87 kV at 8.85 kHz, air) | ~0.7–1 log10 CFU/mL reduction in viable spore number after 2 min treatment | [120] | |
| Disinfection of pre-harvest plants | Botrytis cinerea (cannabis influorescence) | RF plasma (6 kV, low pressure air with the addition of H2O2 (35%)) | 5 log10 CFU reduction in viable fungal spores on influorescence | [121] |
| Colletotrichum gloeosporioides (Green Emerald leaves) | Plasma jet (5 kV, 11 W, mixture of 97% He and 3% O2) | Complete recovery of leaves with black spot diameter of <2 mm after plasma treatment for 3 weeks (twice a day and 10 s per each treatment) | [122] | |
| Food sanitation | Aspergillus brasiliensis (oninon powder) | Microwave plasma (2.45 GHz, 900 W, He) | 1.6 log10 CFU/cm2 reduction after 40 min treatment at 400 W | [123] |
| Aspergillus flavus (in-package beef jerky) | Flexible thin-layer plasma (15 kHz, air) | 2 -3 log10 CFU/g reduction in number of viable fungal spores on beef jerky after 10 min treatment No significant change in metmyoglobin content, shear force, myofibrillar gragmentation index Negative effects on flavor, off-color, and overall acceptability | [124] | |
| (in-package pistachio) | Dielectric barrier discharge plasma (12.5 kHz suppressed by a modulated pulsed signal at 110 Hz, 2.49 W/cm3, air) | 4 log10 CFU/sample reduction in number of viable fungal spores on pistachio after 18 min treatment Slight reduction in moisture content of pistachio and no change in pH of pistachio | [125] | |
| (red pepper powder) | Microwave plasma (2.45 GHz, 50–1000 W, N2, mixture of N2 and O2, He, or mixture of He and O2) | 2.5 log10 CFU/g reduction in number of viable fungal spores in red pepper powder after 20 min treatment with N2 plasma | [126] | |
| (brown rice cereal bar) | RF plasma jet (50–600 kHz RF power, 0–40 W, Ar) | No fungal growth on cereal bars for up to 20 days under 25 °C and 100% relative humidity after 20 min treatment at 40 W | [127] | |
| Aspergillus sp. Rhizopus sp. Penicillium sp. (saffron) | Low-pressure RF oxygen plasma (10–90 W, 8.5 mTorr system pressure, 13.5 mTorr working pressure, O2) | Complete inactivation of fungi after 15 min treatment at 60 W | [128] | |
| Candida albicans Saccharomyces cerevisiae (tomato juice) | AC gliding arc plasma (3.8 kV at 50 Hz, 40 W, N2) | ~4 log10 CFU/g reduction in fungal cell viability in tomato juice after 600 s treatment followed by storage for 10 days No substantial change in the physicochemical properties of tomato juice | [129] | |
| Cladosporium cladosporioides Penicillium citrinum (dried filefish fillets) | Oxygen plasma (photoplasma; Model InDuct, ID 60, BioZone Scientific International Inc., Orlando, FL, USA) | 0.91 and 1.04 log10 CFU/g reduction in number of C. cladosporioides and P. citrinum on fillets, respectively, after 3–20 min treatment Increase in the level of thiobarbituric acid reactive substance (TBARS) and decrease in overall sensory acceptance after 20 min treatment | [130] | |
| Contaminated fungi (shredded salted kimchi cabbage) | Plasma-activated water (PAW): treated with a plasma system (18 kV at 14.3 kHz, air) | 1.8 log10 CFU/g reduction in yeasts and molds associated with kimchi cabbages after submerging in PAW treated with plasma for 120 min Combined treatment with mild heating can enhance fungal inactivation | [131] | |
| Mycotoxin degradation | Aflatoxin (hazelnuts) | Dielectric barrier discharge plasma (100–150 kHz, 0.4–2 kW, N2 or mixture of N2 and air) | >70% reduction in the content of total aflatoxins and aflatoxin B1 on hazelnuts after 12 min treatment at 1000 W | [132] |
| (groundnuts) | RF-plane-type plasma (13.56 MHz RF power, 40 W and 60 W, air) | >70% and 90% reduction in the content of aflatoxin B1 on groundnuts after treatment for 50 min at 40 W and 12 min at 60 W, respectively | [81] | |
| (hazelnuts) | Atmospheric pressure plasma jet (25 kHz, 655 W, air)Low-pressure RF plasma (13.56 MHz RF power, 100 W, <0.25 mbar, air) | 72–73% reduction in the amount of aflatoxin B1 spiked on hazelnuts after treatment with both plasmas | [133] | |
| (rice and wheat) | Corona discharge plasma jet (20 kV at 58 kHz, air) | 45–56% reduction in the level of aflatoxin B1 on rice and wheat after 30 min treatment | [134] | |
| (corn kernels) | DC surface barrier discharge plasma (0.18–0.31 W/cm, air) | Complete degradation of aflatoxin B1 after 480 s treatment | [135] | |
| (slideglass, pistachio nuts) | Dielectric barrier discharge plasma (15 kV at 20 kHz, 130 W, air) | Maximum 64.63% and 52.42% reduction in the level of aflatoxin B1 on slideglass and pistachio nuts, respectively, after 180 s treatment | [136] | |
| Deoxynivalenol, zearalenone, enniatins, fumonisin B1, T2 toxin, sterigmatocystin, AAL toxin (coverglass, rice extracts) | High-voltage pulsed atmospheric-pressure-plasma (~19 kV at 17 kHz, air) | Complete removal of all mycotoxins on coverglass after 60 s treatment; fumon: fumonisin B1 is most sensitive and sterigmatocystin is most resistant Various degradation rates of mycotoxins in extracts of fungal cultures on rice | [137] | |
| Fungal contamination in medicine | ||||
| Prevention of onychomycosis | Candida albicans Trichophyton mentagrophytes (fungal suspension, infected nail) | Dielectric barrier discharge (5–20 kV at 1 Hz–1 kHz, air) | Complete killing of C. albicans and T. mentagrophytes in suspension after 12 min treatment at dose of 30 and 15 kPulses, respectively 100× reduction in viable cell number of C. albicans on nail after treatment at dose of 550 kPulses | [138] |
| Trichophyton benhamiae Trichophyton interdigital Trichophyton rubrum (fungal suspension, patients with infected nails) | Negative DC corona discharge (7 kV, 150 µA, air) | Complete inactivation of all fungal species in vitro straight after plasma treatment More than 70% of onychomycosis patients are cured after the combined treatment of plasma and nail plate abrasion and refreshment | [139] | |
| Trichophyton rubrum (infected sliced hoof discs) | Plasma jet (8 kV at 4 kHz, mixture of 99.5% He and 0.5% O2) Surface microdischarge (SMD) plasma (2.5 kV at ~25 kHz, air) Floating electrode (FE) dielectric barrier discharge (DBD) plasma (6 kV at 4 kHz, air) | 1 and ~3 log10 CFU reduction in viable cell number of T. rubrum infected in hoof discs after 45 min treatment with SMD plasma and 10 min treatment with FE-DBD plasma, respectively | [140] | |
| Prevention of dermatophytosis | Arthroderma benhamiae Microsporum gypseu Trichophyton interdigitale Trichophyton rubrum (fungal suspension in water, fungal spores on agar plates) | Positive and negative point-to-plane corona discharge plasma (10 kV, 0.5 mA, air) Cometary discharge plasma (10 kV at 20 kHz, air) | In suspension: significant decrease in number of viable spores of all fungal species after 15 min treatment, complete killing of T. interdigitale and T. rubrum spores after 25 min treatment On agar plates: complete killing of all fungal species, except M. gypseu, after 25 min treatment | [141] |
| Candida albicans Candida glabrata Candida krusei (fungal suspension in water, fungal spores on sabouraud dextrose agar plates) | Plasma microjet (400 V, 35 mA, mixture of 98% He and 2% O2) | >90% inactivation of fungal spores after 10 min on agar plates and 1 min in suspension | [142] | |
| Epidermophyton floccosum Microsporum canis Microsporum gypseum Trichophyton mentagrophytes, Trichophyton rubrum (fungal suspension, infected guinea pig) | Plasma jet (0.6 kV, 15 mA, 21 kHz, air) in combination with silver nanoparticles | Reduction in values of minimum inhibitory concentration (MIC) of silver nanoparticles after the combined treatment with plasma Enhancement of fungal mycelium permeability of nanoparticles after the combined treatment with plasma Increase in efficiency of healing and suppressing disease symptoms of guinea pig skin after the combined treatment of nanoparticles and plasma | [143] | |
| Prevention of dermatophytosis | Trichophyton mentagrophytes (Infected guinea pig) | Cometary discharge plasma (5 kV, discharge at 50–100 µA, air) | A week shorter and milder infection in guinea pigs treated with plasma Significant reduction in number of viable fungal cells in guinea pigs treated with plasma No adverse effects on guinea pigs | [144] |
| Prevention of oral candidiasis | Candida albicans (Fungal biofilm, infected mouse tung) | Amplitude-modulated cold atmospheric-pressure plasma jet (13 kV, 32 kHz, He) | Significant reduction in the viability of C. albicans biofilms after 5 min treatment No significant difference in values of CFU/tongue but marked reduction in candidal tissue invasion after plasma treatment No adverse effects on mouse cells | [145] |
| Killing of clinical fungal strains | Candida albicans Microsporum canis Trichophyton interdigitale Trichophyton rubrum (fungal cells on agar plates, dandruffs, shoes from a patient with chronic tinea pedis) | Plasma jet (1–5 kV and 1.5 MHz RF power, Ar) | The largest growth inhibition zone on C. albicans agar plate and the smallest zone on M. canis agar plate after 15 s treatment Complete removal of viable fungal elements of T. interdigitale in dandruffs and contaminated shoes after plasma treatment | [146] |
| Candida albicans (fungal cells on agar plates) | Glow discharge microplasma jet (1 kV at 20 kHz, 860 Torr, He) | Increase in growth inhibition zone in sabouraud dextrose agar plates after 1.5 min treatment | [147] | |
| Trichophyton mentagrophytes Trichophyton rubrum (Fungal suspension, infected skin model) | Floating electrode-dielectric barrier discharge plasma jet (8 kV, ~33 mA, 49 W, Ar) | ~96% and 90% reduction in CFU number of T. mentagrophytes and T. rubrum, respectively, after 5 min treatment Significant inhibition of hyphal growth of both fungal species in infected skin mimicking model after plasma treatment | [148] | |
| Application | Fungi | Plasma Source (Treatment Parameters) | Effects | Ref. |
|---|---|---|---|---|
| Enhancement of spore germination and protein secretion | Aspergillus oryzae | Micro-dielectric barrier discharge plasma (1.2 kV, 50–63 mA, 28.8 ms on and 160 ms off pulse times, N2) | Significant increase in percentage of spore germination in phosphate buffered saline (PBS) and potato dextrose broth (PDB) after 2 min and 5 min treatments, respectively 7.4–9.3% increase in activity of α-amylase in PDB after 24 and 48 h of plasma treatment (5 min) | [20,168] |
| Plasma jet (~0.68 kv at ~83 kHz, ~77 mA, air) | ~10% increase in spore germination after 5 min and 10 min treatments Significant elevation of α-amylase activity in PDB after 24–96 h of plasma treatment (10 and 15 min) | [169] | ||
| Pichia pastoris | Plasma jet (0–15 kV at 10 kHz, He) | Increased production of recombinant phytase by P. pastoris after plasma treatment 125% increase in activity of phytase in commercial enzyme solution after plasma treatment | [18] | |
| Mutagenesis | Aspergillus nidulans | Atmospheric and room temperature plasma (ARTP) mutation system: radio-frequency atmospheric-pressure glow discharge (RF APGD) plasma jet (150–300 V, 15–50 MHz RF power, 40–120 W, He) Commercial product from Siqingyuan Biotechnology Co., Ltd., Beijing or Wuxi, China) | Mutant: echinocandin B production of 1.3-fold higher than that of the parental strain | [170] |
| Aspergillus niger | ARTP mutation system | Four mutants: gluconate production of 15.5%, 32.8%, 12.1%, and 70% higher than that of the parental strain | [171,172] | |
| Aspergillus oryzae | ARTP mutation system | Mutants: 54.7, 17.3, and 8.5% increase in activities of acid protease, neutral protease, and total protease, respectively, 292.3% increase in kojic acid production, enhanced activities of salt-tolerant proteases | [19,173,174] | |
| Aspergillus terreus | ARTP mutation system | Mutant: growth and secretion of itaconic acid in undetoxified enzymatic hydrolysate | [175] | |
| Auerobasidium pullulans | ARTP mutation system | Mutant: 13.8% increase in polymalic acid production | [176] | |
| Blakeslea trispora | ARTP mutation system | Mutant: 55% increase in lycopene production and requirement of 10% less (than that of parent strain) dissolved oxygen for maximum production | [177] | |
| Candida glabrata | ARTP mutation system | Mutant: 32.2–35.4% increase in pyruvate production | [178,179] | |
| Candida parapsilosis | ARTP mutation system | Mutant: 53.98% increase in D-arabitol production | [180] | |
| Candida tropicalis | ARTP mutation system | Mutant: 22% increase in xylitol production, increase in gene expression and activity of xylose reductase | [181] | |
| Fusidium coccineum | ARTP mutation system | Mutant: 59.4% increase in fusidic acid production | [182] | |
| Ganoderma lingzhi | Dielectric barrier discharge plasma (10–15 kV, Ar or He) | Mutant: 25.6% increase in polysaccharides production | [183] | |
| Glarea lozoyensis | ARTP mutation system | Mutant: 1.39 fold increase in pneumocandin B0 production | [184] | |
| Hericium erinaceus | ARTP mutation system | Mutant: 22% and 16% increase in the yield of fruiting body and polysaccharide production, respectively | [185] | |
| Mortierella alpina | ARTP mutation system | Mutant: 40.61% increase in arachidonic acid production | [186] | |
| Penicillium oxalicum | Combined treatment with ARTP mutation system and ethylmethanesulfonate | Mutant: 61.1% increase in production of raw starch-degrading enzymes | [187] | |
| Pichia anomala | ARTP mutation system | Mutant: 32.3% increase in sugar alcohol production | [188] | |
| Rhodosporidium toruloides | ARTP mutation system | Mutants: improvement in tolerance to the inhibitory compounds in lignocellulosic hydrolysate and producing lipids with sugarcane bagasse hydrolysate as carbon source, improvement in production of lipids and carotenoids Enhanced expression of four genes is related to the tolerance to lignocellulosic hydrolyzate | [189,190,191,192] | |
| Rhodotorula mucilaginosa | ARTP mutation system | Mutant: 67% increase in carotenoids production | [193] | |
| Saccharomyces cerevisiae | ARTP mutation system | Mutant: 72.54% decrease in production of methanol, which is a toxic by-product of brewing wine Mutant: 56.76% increase in glutathione production, improvement of the activity of glutathione synthetases | [194,195] | |
| Sanghuangporous sanghuang | ARTP mutation system | Mutant: 1.2–1.5 fold increase in polysaccharides production | [196] | |
| Starmerella bombicola | ARTP mutation system | Mutants: over 30% increase in lactonic, acidic, or total sophorolipid production | [197] | |
| Trichoderma reesei | ARTP mutation system | Mutant: increase in cellulase production Mutation in galactokinase gene may be related to improvement of cellulase production Up-regulation of cellulase and hemicellulose genes | [198] | |
| Trichoderma viride | ARTP mutation system | Mutant: 2.18–2.61 fold increase in activities of cellulases Mutant: 1.97 fold increase in total cellulase activity | [199,200] | |
| Yarrowia lipolytica | ARTP mutation system | Mutant: 45.4–51.8% increase in α-ketoglutaric acid production Mutations in genes regulating mitochondrial biogenesis and energy metabolism and a gene associated with cell cycle control are responsible for improvement of α-ketoglutaric acid production Mutant: the highest yield of erythritol production (64.8 g/L erythritol from 100 g/L glycerol) | [201,202,203] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veerana, M.; Yu, N.; Ketya, W.; Park, G. Application of Non-Thermal Plasma to Fungal Resources. J. Fungi 2022, 8, 102. https://doi.org/10.3390/jof8020102
Veerana M, Yu N, Ketya W, Park G. Application of Non-Thermal Plasma to Fungal Resources. Journal of Fungi. 2022; 8(2):102. https://doi.org/10.3390/jof8020102
Chicago/Turabian StyleVeerana, Mayura, Nannan Yu, Wirinthip Ketya, and Gyungsoon Park. 2022. "Application of Non-Thermal Plasma to Fungal Resources" Journal of Fungi 8, no. 2: 102. https://doi.org/10.3390/jof8020102