
Reappraisal of the Genus Exsudoporus (Boletaceae) Worldwide Based on Multi-Gene Phylogeny, Morphology and Biogeography, and Insights on Amoenoboletus

 , , ,
, , ,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection Site and Sampling
2.2. Morphological Study
2.3. DNA Extraction, PCR Amplification and DNA Sequencing
2.4. Sequence Alignment, Data Set Assembly and Phylogenetic Analysis
3. Results
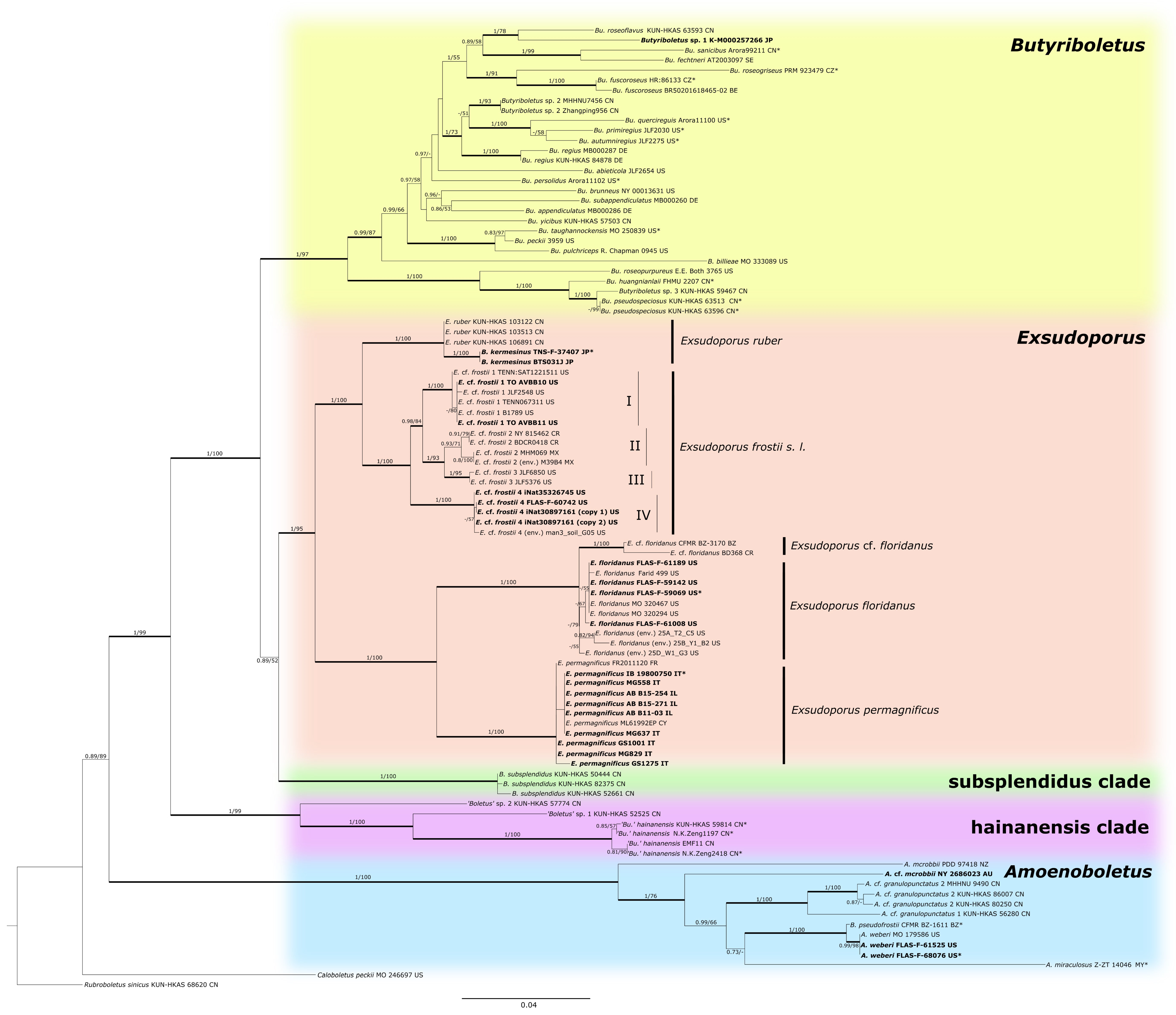
3.1. Molecular Phylogenetic Analysis
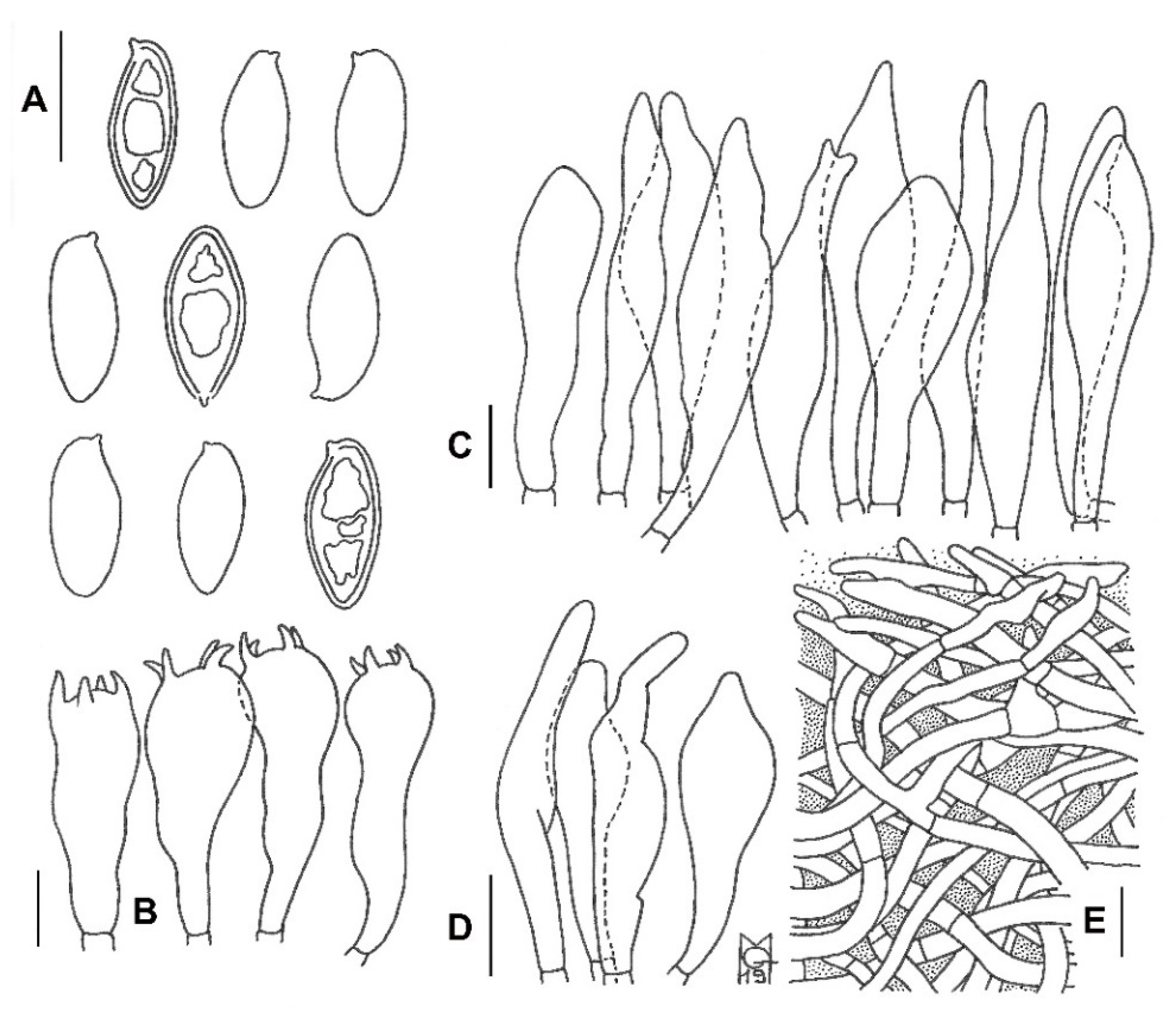
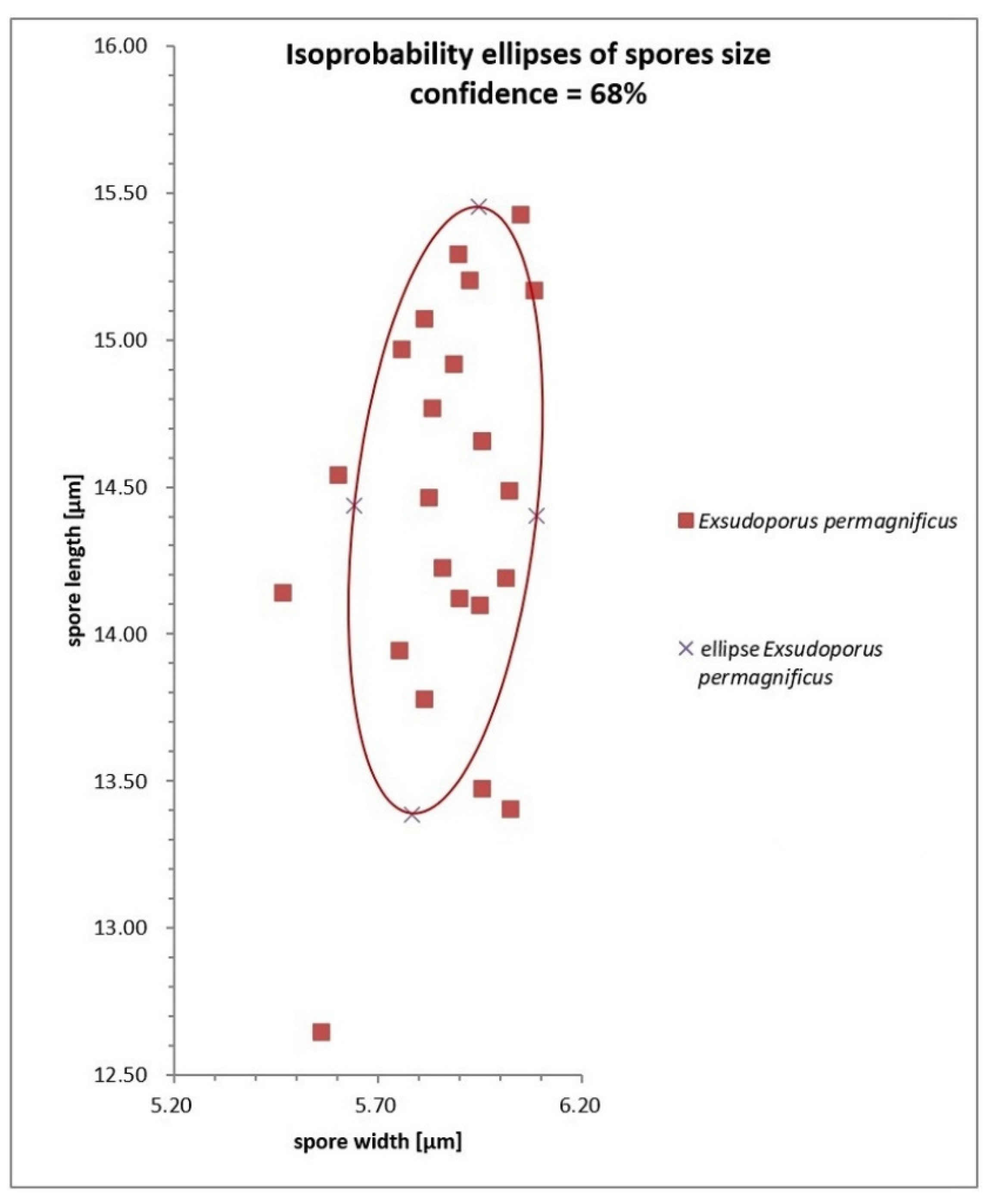
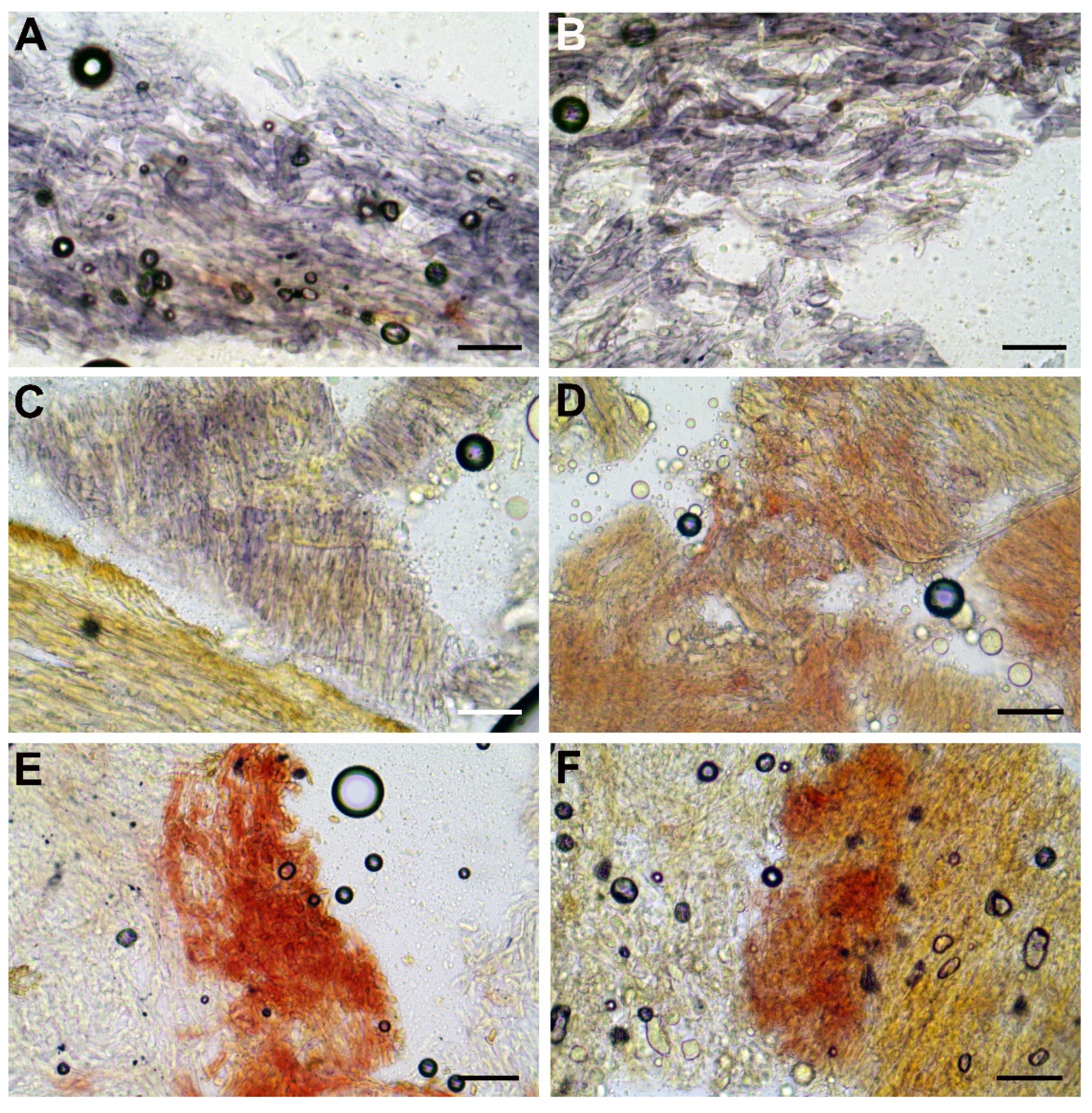
3.2. Taxonomy
Extralimital Taxa
4. Discussion
Key to the Described Species of Exsudoporus
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fries, E.M. Epicrisis Systematis Mycologici Seu Synopsis Hymenomycetum; Typographia Academica: Uppsala, Sweden, 1838. [Google Scholar]
- Horak, E. Synopsis generum Agaricalium (Die Gattungstypen der Agaricales). In Beiträge zur Kryptogamenflora der Schweiz; Waber: Bern, Germany, 1968; Volume 13. [Google Scholar]
- Moser, M.M. Die Röhrlinge und Blätterpilze (Polyporales, Boletales, Agaricales, Russulales). In Kleine Kryptogamenflora IIb/2, Basidiomyceten, XIII, 5th ed.; Gams, H., Ed.; Gustav Fischer: Stuttgart, Germany, 1983. [Google Scholar]
- Singer, R. The Agaricales in Modern Taxonomy, 4th ed.; Koeltz Scientific Books: Koenigstein, Germany, 1986. [Google Scholar]
- Lannoy, G.; Estadès, A. Flore Mycologique d’Europe 6–Les Bolets. In Doc. Mycol. Mém. Hors Série; Association d’Écologie et de Mycologie: Lille, France, 2001; Volume 6. [Google Scholar]
- Watling, R. A manual and source book on the boletes and their allies. In Synopsis Fungorum; Fungiflora: Oslo, Norway, 2008; Volume 24. [Google Scholar]
- Smith, A.H.; Thiers, H.D. The Boletes of Michigan; University of Michigan Press: Ann Arbor, MI, USA, 1971. [Google Scholar]
- Bertéa, P.; Estadès, A. Boletus luteocupreus. Bull. Féd. Micol. Dauphiné-Savoie 1990, 118, 25–31. [Google Scholar]
- Cazzoli, P. Contributo allo studio dei boleti del gruppo purpureus-torosus. In “Il Fungo”, Atti III Seminario Internazionale “Russulales e Boletales”, Castelnovo ne’ Monti, Reggio nell’Emilia, Italy, 6-9 September 1990; Associazione Micologica Bresadola Gruppo “R. Franchi”: Reggio Emilia, Italy, 1990 (“1991”); pp. 29–36. [Google Scholar]
- Andary, C.; Cosson, L.; Bourrier, M.J.; Wylde, R.; Heitz, A. Chimiotaxonomie des bolets de la section Luridi. Cryptogam. Mycol. 1992, 13, 103–114. [Google Scholar]
- Redeuilh, G. Contribution à l’étude des bolets II. Étude critique de Boletus torosus et Boletus xanthocyaneus. Bull. Soc. Mycol. Fr. 1992, 108, 155–172. [Google Scholar]
- Watling, R.; Hills, A.E. Boletes and their allies—Boletaceae, Strobilomycetaceae, Gyroporaceae, Paxillaceae, Coniophoraceae, Gomphidiaceae (revised and enlarged edition). In British Fungus Flora, Agarics and Boleti; Henderson, D.M., Watling, R., Eds.; HMSO: Edinburgh, UK, 2005; Volume 1. [Google Scholar]
- Zang, M. An annotated check-list of the genus Boletus and its sections from China. Fungal Sci. 1999, 14, 79–87. [Google Scholar]
- Galli, R. I Boleti—Atlante Pratico-Monografico per la Determinazione dei Boleti, 3rd ed.; Dalla Natura: Milano, Italy, 2007. [Google Scholar]
- Binder, M.; Hibbett, D.S. Molecular systematics and biological diversification of Boletales. Mycologia 2006, 98, 971–981. [Google Scholar] [CrossRef]
- Nuhn, M.E.; Binder, M.; Taylor, A.F.S.; Halling, R.E.; Hibbett, D.S. Phylogenetic overview of the Boletineae. Fungal Biol. 2013, 117, 479–511. [Google Scholar] [CrossRef]
- Wu, G.; Feng, B.; Xu, J.P.; Zhu, X.T.; Li, Y.C.; Zeng, N.K.; Hosen, I.; Yang, Z.L. Molecular phylogenetic analyses redefine seven major clades and reveal 22 new generic clades in the fungal family Boletaceae. Fungal Divers. 2014, 69, 93–115. [Google Scholar] [CrossRef]
- Wu, G.; Li, Y.C.; Zhu, X.T.; Zhao, K.; Han, L.H.; Cui, Y.Y.; Li, F.; Xu, J.P.; Yang, Z.L. One hundred noteworthy boletes from China. Fungal Divers. 2016, 81, 25–188. [Google Scholar] [CrossRef]
- Zeng, N.K.; Wu, G.; Li, Y.C.; Liang, Z.Q.; Yang, Z.L. Crocinoboletus, a new genus of Boletaceae (Boletales) with unusual boletocrocin polyene pigments. Phytotaxa 2014, 175, 133–140. [Google Scholar] [CrossRef]
- Vizzini, A. Nomenclatural Novelties. Index Fungorum 2014a. No. 146. Available online: http://www.indexfungorum.org/Names/IndexFungorumPublicationsListing.asp (accessed on 10 December 2020).
- Vizzini, A. Nomenclatural Novelties. Index Fungorum 2014c. No. 188. Available online: http://www.indexfungorum.org/Names/IndexFungorumPublicationsListing.asp (accessed on 10 December 2020).
- Vizzini, A. Nomenclatural Novelties. Index Fungorum 2014d. No. 192. Available online: http://www.indexfungorum.org/Names/IndexFungorumPublicationsListing.asp (accessed on 10 December 2020).
- Zhao, K.; Wu, G.; Yang, Z.L. A new genus, Rubroboletus, to accommodate Boletus sinicus and its allies. Phytotaxa 2014, 188, 61–77. [Google Scholar] [CrossRef]
- Assyov, B.; Bellanger, J.M.; Bertéa, P.; Courtecuisse, R.; Koller, G.; Loizides, M.; Marques, G.; Muñoz, J.A.; Oppicelli, N.; Puddu, D.; et al. Nomenclatural Novelties. Index Fungorum 2015. No. 243. Available online: http://www.indexfungorum.org/Names/IndexFungorumPublicationsListing.asp (accessed on 10 December 2020).
- Gelardi, M.; Simonini, G.; Ercole, E.; Davoli, P.; Vizzini, A. Cupreoboletus (Boletaceae, Boletineae), a new monotypic genus segregated from Boletus sect. Luridi to reassign the Mediterranean species B. poikilochromus. Mycologia 2015, 107, 1254–1269. [Google Scholar] [CrossRef] [PubMed]
- Vizzini, A. Nomenclatural Novelties. Index Fungorum 2014b. No. 183. Available online: http://www.indexfungorum.org/Names/IndexFungorumPublicationsListing.asp (accessed on 10 December 2020).
- Simonini, G.; Vizzini, A. Boletus mendax, una specie recentemente descritta in Italia ed i nuovi orientamenti sulla sistematica della sez. Luridi del genere Boletus. In Proceedings of the XL Mostra Reggiana del Fungo, Reggio Emilia, Italy, 10–11 October 2015; Associazione Micologica Bresadola: Reggio Emilia, Italy, 2015; pp. 3–24. [Google Scholar]
- Bessette, A.E.; Roody, W.C.; Bessette, A.R. Boletes of Eastern North America; Syracuse University Press: Syracuse, NY, USA, 2016. [Google Scholar]
- Dentinger, B.T.M.; Ammirati, J.F.; Both, E.E.; Desjardin, D.E.; Halling, R.E.; Henkel, T.W.; Moreau, P.-A.; Nagasawa, E.; Soytong, K.; Taylor, A.F.S.; et al. Molecular phylogenetics of porcini mushrooms (Boletus section Boletus). Mol. Phy. Evol. 2010, 57, 1276–1292. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Xu, J.; Wu, G.; Zeng, N.K.; Li, Y.C.; Tolgor, B.; Kost, G.W.; Yang, Z.L. DNA sequence analyses reveal abundant diversity, endemism and evidence for Asian origin of the porcini mushrooms. PLoS ONE 2012, 7, e37567. [Google Scholar] [CrossRef]
- Zeng, N.K.; Cai, Q.; Yang, Z.L. Corneroboletus, a new genus to accommodate the southeast Asian Boletus indecorus. Mycologia 2012, 104, 1420–1432. [Google Scholar] [CrossRef] [PubMed]
- Arora, D.; Frank, J.L. Clarifying the butter Boletes: A new genus, Butyriboletus, is established to accommodate Boletus sect. Appendiculati, and six new species are described. Mycologia 2014, 106, 464–480. [Google Scholar] [CrossRef] [PubMed]
- Šutara, J.; Janda, V.; Kříž, M.; Graca, M.; Kolařík, M. Contribution to the study of genus Boletus, section Appendiculati: Boletus roseogriseus sp. nov. and neotypification of Boletus fuscoroseus Smotl. Czech Mycol. 2014, 66, 1–37. [Google Scholar] [CrossRef]
- Zhao, K.; Wu, G.; Halling, R.E.; Yang, Z.L. Three new combinations of Butyriboletus (Boletaceae). Phytotaxa 2015, 234, 51–62. [Google Scholar] [CrossRef]
- Smith, M.E.; Amses, K.R.; Elliott, T.F.; Obase, K.; Aime, M.C.; Henkel, T.W. New sequestrate fungi from Guyana: Jimtrappea guyanensis gen. sp. nov., Castellanea pakaraimophila gen. sp. nov., and Costatisporus cyanescens gen. sp. nov. (Boletaceae, Boletales). IMA Fungus 2015, 6, 297–317. [Google Scholar] [CrossRef]
- Henkel, T.W.; Obase, K.; Husbands, D.; Uehling, J.K.; Bonito, G.; Aime, M.C.; Smith, M.E. New Boletaceae taxa from Guyana: Binderoboletus segoi gen. and sp. nov., Guyanaporus albipodus gen. and sp. nov., Singerocomus rubriflavus gen. and sp. nov., and a new combination for Xerocomus inundabilis. Mycologia 2016, 108, 157–173. [Google Scholar] [CrossRef]
- Crous, P.W.; Luangsa-ard, J.J.; Wingfield, M.J.; Carnegie, A.J.; Hernández-Restrepo, M.; Lombard, L.; Roux, J.; Barreto, R.W.; Baseia, I.G.; Cano-Lira, J.F.; et al. Fungal Planet description sheets: 785–867. Persoonia 2018, 41, 238–417. [Google Scholar] [CrossRef]
- Bozok, F.; Assyov, B.; Taşkin, H. First records of Exsudoporus permagnificus and Pulchroboletus roseoalbidus (Boletales) in association with non-native Fagaceae, with taxonomic remarks. Phytol. Balc. 2019, 25, 13–27. [Google Scholar]
- Loizides, M.; Bellanger, J.-M.; Assyov, B.; Moreau, P.-A.; Richard, F. Present status and future of boletoid fungi (Boletaceae) on the island of Cyprus: Cryptic and threatened diversity unravelled by ten-year study. Fungal Ecol. 2019, 41, 65–81. [Google Scholar] [CrossRef]
- He, M.Q.; Zhao, R.L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.; Raspe, O.; Kakishima, M.; Sanchez-Ramirez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef] [Green Version]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.; Saxena, R.K. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Wu, G.; Li, M.X.; Horak, E.; Yang, Z.L. Phylogenetic analysis reveals the new genus Amoenoboletus from Asia and New Zealand. Mycosphere 2021, 12, 1038–1076. [Google Scholar] [CrossRef]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. New York Botanical Garden’s Virtual Herbarium. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 14 December 2021).
- Index Fungorum. Available online: http://www.indexfungorum.org/ (accessed on 24 December 2021).
- Mycobank. Available online: https://www.mycobank.org/ (accessed on 24 December 2021).
- MyCoPortal. Available online: https://www.mycoportal.org/ (accessed on 10 October 2021).
- Ridgway, R. Color Standards and Color Nomenclature; Privately Published: Washington, DC, USA, 1912. [Google Scholar]
- White, T.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Snisky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes. Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubeta, M.A.; Echandi, E.; Abernethy, L.; Vilgalys, R. Characterization of anastomosis groups of binucleate Rhizoctonia species using restriction analysis of an amplified ribosomal RNA gene. Phytopathology 1991, 81, 1395–1400. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–89. [Google Scholar] [CrossRef]
- Matheny, P.B. Improving phylogenetic inference of mushrooms with RPB1 and RPB2 nucleotide sequences (Inocybe; Agaricales). Mol. Phylogenet. Evol. 2005, 35, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetic analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. RaxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Meth. Ecol. Evol. 2020, 12, 373–377. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Suchard, M.; Xie, D.; Drummond, A. Tracer v. 1.6. Institute of Evolutionary Biology, University of Edinburgh, Scotland, 2014. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 16 December 2021).
- Alessio, C.L. Boletus frostii Russell anche in Italia? Micol. Ital. 1980, 9, 15–20. [Google Scholar]
- Angarano, M. Boletus flammans. Boll. Gr. Micol. Bres. 1980, 23. Front cover. [Google Scholar]
- Cazzoli, P.; Consiglio, G. Micologia di Base—Approccio al Genere Boletus—II. Riv. Micol. 2001, 44, 195–213. [Google Scholar]
- Alessio, C.L. Ancora sul presunto Boletus frostii Russel. Micol. Ital. 1981, 10, 39–41. [Google Scholar]
- Alessio, C.L. E’ Boletus siculus Inzenga il già presunto B. frostii Russel rinvenuto in Italia. Micol. Ital. 1981, 10, 40–42. [Google Scholar]
- Inzenga, G. Funghi Siciliani; Centuria 2. F Lao: Palermo, Italy, 1869. [Google Scholar]
- Pöder, R. Boletus permagnificus sp. nov., un appariscente boleto della Sez. Luridi Fr. associato alle querce. Boll. Gr. Micol. Bres. 1983, 26, 82–89. [Google Scholar]
- Bellù, F. Precisazioni e commenti. La questione flammans. Boll. Gr. Micol. Bres. 1981, 24, 50. [Google Scholar]
- Pöder, R. Boletus permagnificus sp. nov., ein Auffallender Röhrling der Sekt. Luridi Fr. assoziiert mit Eichen. Sydowia 1981 (“1982”), 34, 149–156. [Google Scholar]
- Alessio, C.L. Boletus permagnificus Pöder è sinonimo di B. siculus Inzenga. Micol. Ital. 1982, 11, 34. [Google Scholar]
- Alessio, C.L. Boletus siculus Inz. e Boletus permagnificus Pöder. Micol. Ital. 1984, 13, 63–68. [Google Scholar]
- Alessio, C.L. Boletus Dill. ex L. In Fungi Europaei, 1st ed.; Giovanna Biella: Saronno, Italy, 1985; Volume 2. [Google Scholar]
- Alessio, C.L. Revisione dei miei lavori comparsi nei numeri 26–50 di Micologia Italiana. Micol. Ital. 1990, 19, 67–73. [Google Scholar]
- Alessio, C.L. Boletus Dill. ex L. (Supplemento). In Fungi Europaei; Giovanna Biella: Saronno, Italy, 1991; Volume 2. [Google Scholar]
- Bellù, F. Precisazioni e commenti. Ancora su Boletus permagnificus e Boletus siculus. Boll. Gr. Micol. Bres. 1986, 29, 75–79. [Google Scholar]
- Lavorato, C. Chiave analitica e note bibliografiche della micoflora del cisto. Boll. AMER 1991, 24, 16–45. [Google Scholar]
- Alessio, C.L. Un boleto non ancora noto. Xerocomus ichnusanus Alessio, Galli et Littini sp. nov. Boll. Gr. Micol. Bres. 1984, 27, 166–170. [Google Scholar]
- Gelardi, M.; Simonini, G.; Ercole, E.; Vizzini, A. Alessioporus and Pulchroboletus (Boletaceae, Boletineae), two novel genera for Xerocomus ichnusanus and X. roseoalbidus from the European Mediterranean basin: Molecular and morphological evidence. Mycologia 2014, 106, 1168–1187. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Dios, J.B. Nomenclatural Novelties. Index Fungorum. 2015. No. 211. Available online: http://www.indexfungorum.org/Names/IndexFungorumPublicationsListing.asp (accessed on 10 December 2020).
- Icard, C.; Hurtado, C. Boletus permagnificus Pöder au pied des Alpes. Bull. Féd. Ass. Mycol. Médit. 1997, 12, 5–10. [Google Scholar]
- Muñoz Sánchez, J.A.; Cadiñanos Aguirre, J.A. Algunos Boletales interesantes de la Península Ibérica. Belarra 2001, 17–18, 55–64. [Google Scholar]
- Muñoz, J.A. Boletus s.l. (excl. Xerocomus). In Fungi Europaei; Edizioni Candusso: Alassio, Italy, 2005; Volume 2. [Google Scholar]
- Horak, E. Röhrlinge und Blatterpilze in Europa; Elsevier: Munich, Germany, 2005; Volume 6. [Google Scholar]
- Assyov, B. New and rare Bulgarian boletes. Mycol. Balc. 2005, 2, 75–81. [Google Scholar]
- Galli, R. I Boleti—Atlante Pratico-Monografico per la Determinazione dei Boleti, 4th ed.; Micologica: Vergiate, Italy, 2013. [Google Scholar]
- Raya López, L.; Moreno Arroyo, B. Flora Micológica de Andalucía; Consejería de Medio Ambiente y Ordenación del Territorio: Sevilla, Spain, 2018. [Google Scholar]
- Davoli, P.; Weber, R.W.S. Analisi cromatografica del profilo pigmentario di Boletus permagnificus mediante HPLC-MS. Micol. Ital. 2001, 30, 89–95. [Google Scholar]
- Davoli, P.; Weber, R.W.S. Simple method for reversed-phase high-performance liquid chromatographic analysis of fungal pigments in fruit-bodies of Boletales (Fungi). J. Chromatogr. A 2002, 964, 129–135. [Google Scholar] [CrossRef]
- Vasquez, G. Indagini Micologiche sulle Boletales Epigee del Territorio Siciliano—Mappatura e Censimento delle Specie. Ph.D. Thesis, Università degli Studi di Catania, Catania, Italy, 2014. [Google Scholar]
- Di Rita, F.; Atzeni, M.; Tudino, F. The history of conifers in central Italy supports long-term persistence and adaptation of mesophilous conifer fungi in Arbutus-dominated shrublands. Rev. Palaeobot. Palyn. 2020, 282, 104300. [Google Scholar] [CrossRef]
- Cetto, B. I Funghi dal Vero; Saturnia: Trento, Italy, 1983; Volume 4. [Google Scholar]
- Engel, H.; Krieglsteiner, G.; Dermek, A.; Watling, R. Dickröhrlinge. Die Gattung Boletus in Europa; Verlag Heinz Engel: Weidhausen bei Coburg, Germany, 1983. [Google Scholar]
- Lavorato, C. Chiave per la determinazione delle Boletaceae delle foreste della Calabria. Pag. Micol. 1996, 5, 2–27. [Google Scholar]
- Migliozzi, V.; Coccia, M. Segnalazione per il territorio laziale di Boletacee interessanti e descrizione di Boletus poikilochromus Pöder, Cetto & Zuccherelli. Boll. AMER 1991, 24, 9–15. [Google Scholar]
- Brotzu, R. Guida ai Funghi della Sardegna—Parte Prima; Editrice Archivio Fotografico Sardo: Nuoro, Italy, 1993. [Google Scholar]
- Foiera, F.; Lazzarini, E.; Snabl, M.; Tani, O. Funghi boleti; Edizioni Edagricole: Bologna, Italy, 1993. [Google Scholar]
- Galli, R. I Boleti—Atlante Pratico-Monografico per la Determinazione dei Boleti; Edinatura: Milano, Italy, 1998. [Google Scholar]
- Papetti, C.; Consiglio, G.; Simonini, G. Atlante Fotografico dei Funghi d’Italia; AMB Centro Studi Micologici: Vicenza, Italy, 2001. [Google Scholar]
- Lunghini, D.; Perrone, L. Contributo allo studio e al monitoraggio delle Boletacee del litorale laziale. 2. Boll. AMER 2002, 54–55, 39–60. [Google Scholar]
- Migliozzi, V.; Camboni, M. La micoflora del litorale romano. 11° contributo. Descrizione di Helvella juniperi, Boletus comptus, Boletus luridus, Boletus permagnificus e Xerocomus roseoalbidus. Boll. Gr. Micol. Bres. 2002, 45, 7–28. [Google Scholar]
- Napoli, M.; Signorello, P. Contributo alla conoscenza della flora macromicetica del siracusano. Boll. AMER 2004, 62–63, 14–32. [Google Scholar]
- Corea, E. Il genere Boletus L. in Calabria—2a parte. Riv. Micol. 2006, 49, 99–113. [Google Scholar]
- Boccardo, F.; Traverso, M.; Vizzini, A.; Zotti, M. Funghi d’Italia; Zanichelli: Bologna, Italy, 2008. [Google Scholar]
- Brotzu, R.; Colomo, S. I Funghi della Sardegna. Basidiomiceti: Cortinariaceae, Bolbitiaceae, Boletaceae, Gomphidiaceae, Paxillaceae; Editrice Archivio Fotografico Sardo: Nuoro, Italy, 2009; Volume 6. [Google Scholar]
- Oppicelli, N. Funghi in Italia; Erredi Grafiche Editoriali: Genova, Italy, 2020. [Google Scholar]
- Šutara, J.; Mikšík, M.; Janda, V. Hřibovité Houby; Academia: Praha, Czech Republic, 2009. [Google Scholar]
- Mikšík, M. Hřibovité Houby Evropy; Vydalo Nakladatelství Svojtka & Co.: Praha, Czech Republic, 2017. [Google Scholar]
- Moreno, G.; Esteve-Raventós, F. Boletus aemilii Barbier, B. permagnificus Pöder and Xerocomus truncatus Singer, Snell et Dick, in Spain. Lazaroa 1988, 10, 253–258. [Google Scholar]
- Moreno, G.; Esteve-Raventós, F.; Illana, C. Estudios micologicos en el parque natural de Monfragüe y otras zonas de Extremadura (España), 4. Agaricales. Bol. Soc. Micol. Madr. 1989, 14, 115–141. [Google Scholar]
- Mendaza, R.; Diaz, G. Las Setas en la Naturaleza; Iberdrola: Bilbao, Spain, 1996; Tomo 1. [Google Scholar]
- Gelpi, C.; De Castro, J. Boletus permagnificus (Pöeder) Citado por Alessio (1985) como B. siculus (Inzenga 1869). Bol. Soc. Micol. Extrem. 2003, 14, 44. [Google Scholar]
- Moreno Arroyo, B. Inventario Micològico Básico de Andalucía; Consejería de Medio Ambiente: Córdoba, Spain, 2004. [Google Scholar]
- Pardo, F.M.V.; Maqueda, S.M.; Pimienta, A.B.L.; Pacheco, D.P. Aproximación al Catálogo de las especies del Orden Boletales (Basidiomycetes, Fungi) en Extremadura (España). Rev. Estud. Extrem. 2004, 40, 1255–1291. [Google Scholar]
- Calzada Domínguez, A.C. Guía de los boletos de España y Portugal; Náyade Editorial: Medina del Campo, Spain, 2007. [Google Scholar]
- Becerra Parra, M.; Robles Domínguez, E.; Díaz Romera, J.A.; Astete Sánchez, G.; Olivera Amaya, M.; López Pastora, A.; Gaona Ríos, J.M.; Peña Máquez, M.I. Nuevas aportaciones al conocimiento de los Boletales Andaluces. Lactarius 2015, 22, 87–99. [Google Scholar]
- Chevassut, G.; Bertéa, P. La poussée fongique de l’automne 1991 en Languedoc. Bull. Féd. Ass. Mycol. Médit. 1992, 2, 11–16. [Google Scholar]
- Joset, H. Les Bolets à pores rouges en Corse; Soc. Mycol. Ajaccio: Ajaccio, France, 1992. [Google Scholar]
- Roth, A. Boletus permagnificus Pöder, Xerocomus roseoalbidus Alessio & Littini. Bull. Soc. Mycol. Fr. 1994, 110, 292–294. [Google Scholar]
- Estadès, A.; Lannoy, G. Les bolets européens. Bull. Féd. Mycol. Dauphinée-Savoie 2004, 174, 3–79. [Google Scholar]
- Assyov, B. Scanning electron microscopic study of the European members of Baorangia, Exsudoporus and Lanmaoa (Boletales, fungi). Comprend. Acad. Bulg. Sci. 2017, 70, 657–662. [Google Scholar]
- Denchev, C.M.; Assyov, B. Checklist of the larger basidiomycetes in Bulgaria. Mycotaxon 2010, 111, 279–282. [Google Scholar] [CrossRef] [Green Version]
- Jurc, D.; Ogris, N.; Piltaver, A.; Dolenc, A. Seznam vrst in Razširjenost Makromicet v Sloveniji z Analizo Stopnje Ogroženosti: Ciljni Raziskovalni Program “Konkurenčnost Slovenije 2001–2006”; Projekt št. V4-0703; Gozdarski Inštitit Slovenije: Ljubljana, Slovenia, 2004. [Google Scholar]
- Constantinidis, G. Mushrooms, a Photographic Guide for Collectors; Privately Published: Athens, Greece, 2009. [Google Scholar]
- Wagensommer, R.P.; Flores, G.A.; Arcangeli, A.; Bistocchi, G.; Maneli, F.; Materozzi, G.; Perini, C.; Venanzoni, R.; Angelini, P. Application of IUCN red listing criteria at the regional level: A case study with Boletales across the Appennine province ecoregion and EU-habitats of central Italy. Plant Biosyst. 2021. [Google Scholar] [CrossRef]
- Bedenko, E.P. Makromitsety Srednerusskoy Vozvyshennosti [Macromycetes of the Central Russian Upland]. Ph.D. Thesis, Lomonosov Moscow State University, Moscow, Russia, 1989. (In Russian). [Google Scholar]
- Singer, R. New Boletaceae from Florida (a preliminary communication). Mycologia 1945, 37, 797–799. [Google Scholar] [CrossRef]
- Singer, R. The Boletoideae of Florida. The Boletineae of Florida with notes on extralimital species III. Am. Mid. Nat. 1947, 37, 1–135. [Google Scholar] [CrossRef]
- Both, E.E. The Boletes of North America. A Compendium; Buffalo Museum of Science: Buffalo, NY, USA, 1993. [Google Scholar]
- Bessette, A.E.; Roody, W.C.; Bessette, A.R. North American Boletes. A Color Guide to the Fleshy Pored Mushrooms; Syracuse University Press: Syracuse, NY, USA, 2000. [Google Scholar]
- Frost, C.C. Catalogue of boleti of New England, with descriptions of new species. Bull. Buffalo Soc. Nat. Sci. 1874, 2, 100–105. [Google Scholar]
- Krisai-Greilhuber, I. Boletus frostii, Frosts Dickröhrling—Ein prachtvoller nordamerikanischer Röhrling [Boletus frostii, Frost’s Bolete—A splendid North American bolete]. Beih. Zeit. Mykol. 1999, 9, 9–13. [Google Scholar]
- Zang, M. Notes on the Boletales from eastern Himalayas and adjacent areas of China. Acta Bot. Yunnanica 1986, 8, 1–22. (In Chinese) [Google Scholar]
- Zang, M. Fungi of the Hengduan Mountains—The Series of the Scientific Expedition to the Hengduan Mountains, Qinghai-Xizang Plateau; Science Press: Beijing, China, 1996. (In Chinese) [Google Scholar]
- Takahashi, H.; Taneyama, Y.; Koyama, A. Boletus kermesinus, a new species of Boletus section Luridi from central Honshu, Japan. Mycoscience 2011, 52, 419–424. [Google Scholar] [CrossRef]
- Zang, M.; Li, X.J.; He, Y.S. Boletaceae (II). In Flora Fungorum Sinicorum; Science Press: Beijing, China, 2013; Volume 44. (In Chinese) [Google Scholar]
- Wu, K.; Wu, G.; Yang, Z.L. A taxonomic revision of Leccinum rubrum in subalpine coniferous forests, southwestern China. Acta Edulis Fungi 2020, 27, 92–100. [Google Scholar] [CrossRef]
- Murrill, W.A. Florida boletes. Lloydia 1948, 11, 21–35. [Google Scholar]
- Ortiz-Santana, B.; Lodge, D.J.; Baroni, T.J.; Both, E.E. Boletes from Belize and the Dominican Republic. Fungal Div. 2007, 27, 247–416. [Google Scholar]
- Bessette, A.E.; Bessette, A.R.; Lewis, D.P. Mushrooms of the Gulf Coast States: A Field Guide to Texas, Louisiana, Missisipi, Alabama, and Florida; University of Texas Press: Austin, TX, USA, 2019. [Google Scholar]
- GBIF. Available online: https://www.gbif.org/ (accessed on 24 May 2021).
- Murrill, W.A. American Boletes; Privately Published: New York, NY, USA, 1914. [Google Scholar]
- Bessette, A.E.; Roody, W.C.; Bessette, A.R.; Dunaway, D.L. Mushrooms of the Southeastern United States; Syracuse University Press: Syracuse, NY, USA, 2007. [Google Scholar]
- García-Jiménez, J.; Garza-Ocañas, F. Conocimiento de los hongos de la familia Boletaceae de México. Ciencia UANL 2001, 4, 336–343. [Google Scholar]
- García-Jiménez, J. Diversidad de Macromicetos en el Estado de Tamaulipas, México. Ph.D. Thesis, Universidad Autónoma de Nuevo León, San Nicolás de los Garza, Mexico, 2013. [Google Scholar]
- García-Jiménez, J.; Singer, R.; Estrada, E.; Garza-Ocañas, F.; Valenzuela, R. Dos especies nuevas del género Boletus (Boletales: Agaricomycetes) en México. Rev. Mex. Bioxbio 2013, 84, 152–162. [Google Scholar] [CrossRef] [Green Version]
- Gonzáles-Chicas, E.; Cappello, S.; Cifuentes, J.; Torres-De la Cruz, M. New records of Boletales (Basidiomycota) in a tropical oak forest from Mexican Southeast. Bot. Sci. 2019, 97, 423–432. [Google Scholar] [CrossRef]
- Flores Arzù, R. Diversity and importance of edible ectomycorrhizal fungi in Guatemala. In Mushrooms, Humans and Nature in a Changing World—Perspectives from Ecological, Agricultural and Social Sciences; Pérez-Moreno, J., Guerin-Laguette, A., Flores Arzù, R., Yu, F.Q., Eds.; Springer Nature: Cham, Switzerlnd, 2020; Chapter 4; pp. 101–139. [Google Scholar]
- Farid, A.; Bessette, A.E.; Bessette, A.R.; Bolin, J.A.; Kudzma, L.V.; Franck, A.R.; Garey, J.R. Investigations in the boletes (Boletaceae) of southeastern USA: Four novel species, and three novel combinations. Mycosphere 2021, 12, 1038–1076. [Google Scholar] [CrossRef]
- Halling, R.E. Boletes described by Charles, C. Frost. Mycologia 1983, 75, 70–92. [Google Scholar] [CrossRef]
- Curtis, M.A. Contributions to the mycology of North America. Am. J. Sci. 2 Ser. 1848, 6, 349–353. [Google Scholar]
- Peck, C.H. Boleti of the United States. Ann. Rep. N. Y. State Mus. Nat. Hist. 1889, 2, 73–166. [Google Scholar]
- Peck, C.H. Report of the State Botanist, 1906. Bull. N. Y. State Mus. 1907, 116, 1–117. [Google Scholar]
- Murrill, W.A. The Boletaceae of North America—I. Mycologia 1909, 1, 4–18. [Google Scholar] [CrossRef]
- Farlow, W.G.; Burt, E.A. Icones Farlowianae: Illustrations of the Larger Fungi of Eastern North America; The Farlow Library and Herbarium of Harvard University: Cambridge, MA, USA, 1929. [Google Scholar]
- Coker, W.C.; Beers, A.H. The Boletaceae of North Carolina; University of North Carolina Press: Chapel Hill, NC, USA, 1943. [Google Scholar]
- Singer, R. Die Röhrlinge. Teil II. Die Boletoideaea und Strobilomycetaceae. In Die Pilze Mitteleuropas; Verlag Julius Klinkhardt: Bad Heilbrunn, Germany, 1967; Band 6. [Google Scholar]
- Snell, W.H.; Dick, E.A. The Boleti of Northeastern North America; Cramer: Lehre, Germany, 1970. [Google Scholar]
- Miller, O.K., Jr. Mushrooms of North America; E.P. Dutton: New York, NY, USA, 1972. [Google Scholar]
- Smith, A.H.; Weber, N.S. The Mushroom Hunter’s Field Guide, 3rd ed.; University of Michigan Press: Ann Arbor, MI, USA, 1980. [Google Scholar]
- Lincoff, G.H. The Audubon Society Field Guide to North American Mushrooms; Alfred, A. Knopf: New York, NY, USA, 1981. [Google Scholar]
- Imler, L. Icones Mycologicae; Jardin Botanique National de Belgique: Meise, Belgium, 1985; p. 96. [Google Scholar]
- Weber, N.S.; Smith, A.H. A Field Guide to Southern Mushrooms; University of Michigan Press: Ann Arbor, MI, USA, 1985. [Google Scholar]
- Arora, D. Mushrooms Demystified, 2nd ed.; Ten Speed Press: Berkeley, CA, USA, 1986. [Google Scholar]
- Bessette, A.E.; Sundberg, W.J. Mushrooms: A Quick Reference Guide to Mushrooms of North America; McMillian Field Guides: New York, NY, USA, 1987. [Google Scholar]
- McKnight, K.H.; McKnight, B.B. A Field Guide to Mushrooms of North America: Peterson Field Guides; Houghton Mifflin: Boston, MA, USA, 1987; Volume 34. [Google Scholar]
- Phillips, R. Mushrooms of North America; Little Brown Company: Boston, MA, USA, 1991. [Google Scholar]
- Metzler, S.; Metzler, V. Texas Mushrooms: A Field Guide; University of Texas Press: Austin, TX, USA, 1992. [Google Scholar]
- Bessette, A.E.; Bessette, A.R.; Fischer, E.W. Mushrooms of Northeastern North America; Syracuse University Press: Syracuse, NY, USA, 1997. [Google Scholar]
- Kibby, G.G. Editorial. Field Mycol. 2005, 6, 38. [Google Scholar] [CrossRef]
- Elliott, T.F.; Stephenson, S.L. Mushrooms of the Southeast; Timber Press: Portland, OR, USA, 2018. [Google Scholar]
- Ibarra, L.V. Etnomicología de la Etnia Wirrárixa (Huichol), Jalisco, México. Master’s Thesis, Universidad de Guadaljara, Guadalajara, Mexico, 1999. [Google Scholar]
- Morris, M.H.; Perez-Perez, M.A.; Smith, M.E.; Bledsoe, C.S. Influence of host species on ectomycorrhizal communities associated with two co-occurring oaks (Quercus spp.) in a tropical cloud forest. FEMS Microbiol. Ecol. 2009, 69, 274–287. [Google Scholar] [CrossRef] [Green Version]
- García-Jiménez, J. Estudio Sobre la Taxonomía, Ecología y Distributión de Algunos Hongos de la Familia Boletaceae (Agaricales, Basidiomycetes) de México. Master’s Thesis, Universidad Autónoma de Nuevo León, San Nicolás de los Garza, Mexico, 1999. [Google Scholar]
- Gándara, E.; Guzmán-Dávalos, L.; Guzmán, G.; Rodríguez, O. Inventario micobiótico de la región de Tapalpa, Jalisco, México. Acta Bot. Mex. 2014, 107, 165–185. [Google Scholar] [CrossRef]
- Robles-García, D.; Suzán-Azpiri, H.; Montoya-Esquivel, A.; García-Jiménez, J.; Esquivel-Naranjo, E.U.; Yahia, E.; Landeros-Jaime, F. Ethnomycological knowledge in three communities in Amealco, Quéretaro, México. J. Ethnobiol. Ethnomed. 2018, 14, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saldivar, Á.E.; García Jiménez, J.; Herrera Fonseca, M.J.; Rodríguez Alcántar, O. Listado actualizado y nuevos registros de Boletaceae (Fungi, Basidiomycota, Boletales) en Jalisco, México. Polibotánica 2021, 52, 25–49. [Google Scholar] [CrossRef]
- Halling, R.E.; Mueller, G.M. Agarics and boletes of neotropical oakwoods. In Tropical Mycology. Macromycetes; Watling, R., Frankland, J.C., Ainsworth, A.M., Isaac, S., Robinson, C.H., Eds.; CABInternational: Wallingford, UK, 2002; Volume 1, pp. 1–10. [Google Scholar]
- Halling, R.E.; Mueller, G.M. Common mushrooms of the Talamanca mountains, Costa Rica. In Memoirs of the New York Botanical Garden; New York Botanical Garden Press: Bronx, NY, USA, 2005; Volume 90. [Google Scholar]
- Sharma, A.D.; Jandaik, C.L.; Munjal, R.L.; Seth, P.K. Some fleshy fungi from Himachal Pradesh–I. Indian J. Mush. 1978, 4, 1–4. [Google Scholar]
- Verma, B.; Pandro, V. Distribution of boletaceous mushrooms in India, some new records from sal forest of central India. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1694–1713. [Google Scholar] [CrossRef]
- Yoneyama, T. Mushrooms of Mt. Fuji; Gotenba Food Hygiene Association: Shizuoka, Japan, 2007. (In Japanese) [Google Scholar]
- Yuan, M.S.; Sun, P.Q. The Pictorial Book of Mushrooms of China; Sichuan Science and Technology Press: Chengdu, China, 2007. (In Chinese) [Google Scholar]
- Li, T.H.; Song, B. Chinese boletes: A comparison of boreal and tropical elements. In Tropical Mycology 2000; The Millenium Meeting on Tropical Mycology (Main Meeting 2000); Walley, A.J.S., Ed.; Liverpool John Moores University: Liverpool, UK, 2000; pp. 1–10. [Google Scholar]
- Fu, S.Z.; Wang, Q.B.; Yao, Y.J. An annotated checklist of Leccinum in China. Mycotaxon 2006, 96, 47–50. [Google Scholar]
- Zhang, Y.; Zhou, D.Q.; Zhou, T.S.; Ou, X.K. New records and distribution of macrofungi in Laojun Mountain, northwest Yunnan, China. Mycosystema 2012, 31, 196–212. [Google Scholar]
- McNabb, R.F.R. The boletaceae of New Zealand. N. Z. J. Bot. 1968, 6, 137–176. [Google Scholar] [CrossRef]
- Stevenson, G. Field Guide to Fungi; University of Canterbury: Canterbury, UK, 1982. [Google Scholar]
- New Zealand Provinces 1848–77. Available online: http://rulers.org/newzprov.html (accessed on 17 December 2021).
- Mushroom Observer. Available online: https://mushroomobserver.org/ (accessed on 10 September 2021).
- Liang, Z.Q.; An, D.Y.; Jiang, S.; Su, M.S.; Zeng, N.K. Butyriboletus hainanensis (Boletaceae, Boletales), a new species from tropical China. Phytotaxa 2016, 267, 256–262. [Google Scholar] [CrossRef]
- Zhang, M.; Li, T.H. Erythrophylloporus (Boletaceae, Boletales), a new genus inferred from morphological and molecular data from subtropical and tropical China. Mycosystema 2018, 37, 1111–1126. [Google Scholar] [CrossRef]
- Janda, V.; Kříž, M.; Kolařík, M. Butyriboletus regius and Butyriboletus fechtneri: Typification of two well-known species. Czech Mycol. 2019, 71, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Kuo, M.; Ortiz-Santana, B. Revision of leccinoid fungi, with emphasis on North American taxa, based on molecular and morphological data. Mycologia 2020, 112, 197–211. [Google Scholar] [CrossRef]
- Chiu, W.F. The Boletes of Yunnan. Mycologia 1948, 40, 199–231. [Google Scholar] [CrossRef]
- Chiu, W.F. Atlas of the Yunnan Bolets; Science Press: Beijing, China, 1957. (In Chinese) [Google Scholar]
- Šutara, J. Anatomical structure of pores in European species of genera Boletus s.str. and Butyriboletus (Boletaceae). Czech Mycol. 2014, 66, 157–170. [Google Scholar] [CrossRef]
- Janda, V.; Kříž, M. Evropské druhy hřibů rodu Butyriboletus [European representatives of genus Butyriboletus]. Mykol. Listy 2016, 135, 11–51. [Google Scholar]
- Morris, M.H.; Perez-Perez, M.A.; Smith, M.E.; Bledsoe, C.S. Multiple species of ectomycorrhizal fungi are frequently detected on individual oak root tips in a tropical cloud forest. Mycorrhiza 2008, 18, 375–383. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher | Locality | GenBank Accession Number | Notes | |||
|---|---|---|---|---|---|---|---|
| ITS | nrLSU | tef1-α | rpb2 | ||||
| Amoenoboletus cf. granulopunctatus 1 | KUN-HKAS 56280 | China | MZ708840 | KF112418 | KF112265 | KF112708 | - |
| A. cf. granulopunctatus 2 | MHHNU 9490 | China | MW520189 | MW520186 | MW566747 | MW560081 | - |
| A. cf. granulopunctatus 2 | KUN-HKAS 80250 | China | MW520191 | MW520185 | MW566746 | MW560080 | - |
| A. cf. granulopunctatus 2 | KUN-HKAS 86007 | China | MW520190 | MW520187 | MZ741478 | MW560079 | - |
| A. mcrobbii | PDD 97418 | New Zealand | MZ708841 | JQ924329 | MZ708841 | - | - |
| A.cf. mcrobbii | NY 2686023 (Halling 9916) | Australia | OL960511 | - | - | - | |
| A.miraculosus | Z-ZT 14046 | Malaysia | MZ708842 | MZ708842 | MZ708842 | - | holotype |
| A. weberi | FLAS-F-61525 | USA | MH211950 | - | - | - | - |
| A. weberi | FLAS-F-68076 | USA | OL960512 | - | - | - | topotype |
| A. weberi | MO 179586 | USA | MH251719 | MH249987 | - | - | - |
| MH249988 | |||||||
| Boletus billieae | MO 333089 | USA | MK542835 | MK542836 | - | - | - |
| B. kermesinus | BTS031J | Japan | OL960532 | - | - | - | |
| B. kermesinus | TNS-F-37407 | Japan | OL960531 | - | - | holotype | |
| B. pseudofrostii | CFMR BZ-1611 (BOS-266) | Belize | MN250201 | MN250176 | - | - | holotype |
| B. subsplendidus | KUN-HKAS 50444 | China | KM388725 | KT990540 | KT990742 | KT990379 | - |
| B. subsplendidus | KUN-HKAS 52661 | China | KM388726 | KF112339 | KF112169 | KF112676 | - |
| B. subsplendidus | KUN-HKAS 82375 | China | KM388727 | - | - | - | - |
| “Boletus” sp. 1 | KUN-HKAS 52525 | China | KU317760 | KF112337 | KF112163 | KF112671 | - |
| “Boletus” sp. 2 | KUN-HKAS 57774 | China | KU317761 | KF112330 | KF112155 | KF112670 | - |
| Butyriboletus abieticola | JLF2654 | USA | KC184418 | - | - | - | |
| Bu. appendiculatus | MB000286 | Germany | KT002599 | KT002610 | KT002634 | - | - |
| Bu. autumniregius | JLF2275 | USA | KC184430 | - | - | paratype | |
| Bu. brunneus | NY 00013631 | USA | KT002600 | KT002611 | KT002635 | - | - |
| Bu. fechtneri | AT2003097 | USA | KC584784 | KF030270 | - | - | - |
| Bu. fuscoroseus | BR50201618465-02 | Belgium | KT002602 | KT002613 | KT002637 | - | - |
| Bu. fuscoroseus | HR:86133 | Czech Rep. | KJ419926 | - | - | neotype | |
| “Bu.” hainanensis | N.K.Zeng1197 | China | KU961653 | KU961651 | - | KU961658 | paratype |
| “Bu.” hainanensis | N.K.Zeng2418 | China | KU961654 | KU961652 | KU961656 | KX453856 | paratype |
| “Bu.” hainanensis | KUN-HKAS 59814 | China | KU317762 | KF112336 | KF112199 | KF112699 | paratype |
| “Bu.” hainanensis | EMF11 | China | JF273514 | - | - | - | - |
| Bu. huangnianlaii | FHMU 2207 (N.K.Zeng3246) | China | MH885351 | MH879689 | MH879718 | MH879741 | holotype |
| Bu. peckii | 3959 | USA | - | JQ326999 | JQ327026 | - | - |
| Bu. persolidus | Arora11102 | USA | KC184441 | - | - | paratype | |
| Bu. primiregius | JLF2030 | USA | KC184455 | - | - | holotype | |
| Bu. pseudospeciosus | KUN-HKAS 63596 | China | - | KT990542 | KT990744 | KT990381 | paratype |
| Bu. pseudospeciosus | KUN-HKAS 63513 | China | KM388728 | KT990541 | KT990743 | - | holotype |
| Bu. pulchriceps | R. Chapman 0945 | USA | KT002604 | KT002615 | KT002639 | - | - |
| Bu. quercireguis | Arora11100 | USA | KC184461 | - | - | holotype | |
| Bu. regius | KUN-HKAS 84878 | Germany | - | MT264910 | MT269659 | MT269661 | - |
| Bu. regius | MB000287 | Germany | KT002605 | KT002616 | KT002640 | - | - |
| Bu. roseoflavus | KUN-HKAS 63593 | China | KJ909517 | KJ184559 | KJ184571 | - | - |
| Bu. roseogriseus | PRM 923479 | Czech Rep. | KJ419928 | - | - | paratype | |
| Bu. roseopurpureus | E.E. Both 3765 | USA | KT002606 | KT002617 | KT002641 | - | - |
| Bu. sanicibus | Arora99211 | China | KC184469 | KC184470 | - | - | holotype |
| Bu. subappendiculatus | MB000260 | Germany | KT002607 | KT002618 | KT002642 | - | - |
| Bu. taughannockensis | MO 250839 | USA | MH234472 | MH234473 | - | - | holotype |
| Bu. yicibus | KUN-HKAS 57503 | China | KT002608 | KT002620 | KT002644 | - | - |
| Butyriboletus sp. 1 | K-M000257266 | Japan | OL960513 | - | - | - | - |
| Butyriboletus sp. 2 | MHHNU7456 | China | - | KT990539 | KT990741 | KT990378 | - |
| Butyriboletus sp. 2 | Zhangping956 | China | KU317759 | KU317764 | - | - | - |
| Butyriboletus sp. 3 | KUN-HKAS 59467 | China | KM388729 | - | - | - | - |
| Caloboletus peckii | MO 246697 | USA | - | MH220330 | MH318614 | - | - |
| Exsudoporus floridanus | MO 320467 | USA | - | MN114633 | - | - | - |
| E. floridanus | MO 320294 | USA | - | MK533764 | - | - | - |
| E. floridanus | 25A_T2_C5 | USA | KX899261 | - | - | - | environmental |
| E. floridanus | 25B_Y1_B2 | USA | KX899270 | - | - | - | environmental |
| E. floridanus | 25D_W1_G3 | USA | KX899279 | - | - | - | environmental |
| E. floridanus | FLAS-F-59069 | USA | OL960514 | OL960488 | OL960496 | OL960503 | epitype |
| E. floridanus | FLAS-F-59142 | USA | OL960515 | - | - | - | - |
| E. floridanus | FLAS-F-61008 | USA | OL960516 | OL960489 | OL960497 | OL960504 | - |
| E. floridanus | FLAS-F-61189 | USA | MH211799 | OL960490 | OL960498 | - | - |
| E. floridanus | Farid 499 | USA | - | - | MW737484 | MW737459 | - |
| E. cf. floridanus | BD368 | Costa Rica | JN020981 | HQ161859 | - | - | - |
| E. cf. floridanus | CFMR BZ-3170 | Belize | MN250222 | MK601725 | MK721079 | MK766287 | - |
| E. cf. frostii 1 | JLF2548 | USA | KC812303 | KC812304 | - | - | - |
| E. cf. frostii 1 | TENN067311 | USA | KT002601 | KT002612 | KT002636 | - | - |
| E. cf. frostii 1 | TENN:SAT1221511 | USA | - | KP055021 | KP055018 | KP055027 | - |
| E. cf. frostii 1 | B1789 | USA | KY826056 | - | - | - | - |
| E. cf. frostii 1 | TO AVBB10 | USA | OL960517 | OL960491 | - | - | - |
| E. cf. frostii 1 | TO AVBB11 | USA | OL960518 | OL960492 | - | OL960505 | - |
| E. cf. frostii 2 | MHM069 | Mexico | EU569285 | - | - | - | - |
| E. cf. frostii 2 | BDCR0418 | Costa Rica | - | HQ161855 | - | - | - |
| E. cf. frostii 2 | NY 815462 | Costa Rica | - | JQ924342 | KF112164 | KF112675 | - |
| E. cf. frostii 2 | M39B4 | Mexico | FJ196902 | - | - | environmental | |
| E. cf. frostii 3 | JLF6850 | USA | MN263010 | MN258884 | - | - | - |
| E. cf. frostii 3 | JLF5376 | USA | MN263009 | MN258883 | - | - | - |
| E. cf. frostii 4 | man3_soil_G05 | USA | GU328546 | - | - | - | environmental |
| E. cf. frostii 4 | FLAS-F-60742 | USA | MH016833 | OL960493 | OL960499 | OL960506 | - |
| E. cf. frostii 4 | iNat35326745 | USA | OL960519 | OL960494 | OL960500 | OL960507 | - |
| E. cf. frostii 4 | iNat30897161 | USA | OL960520 | OL960495 | OL960501 | OL960508 | - |
| OL960521 | |||||||
| E. permagnificus | IB 19800750 | Italy | OL960522 | - | - | - | holotype |
| E. permagnificus | AB B11-03 | Israel | OL960523 | - | - | - | |
| E. permagnificus | AB B15-254 | Israel | OL960524 | - | - | - | - |
| E. permagnificus | AB B15-271 | Israel | OL960525 | - | OL960509 | - | |
| E. permagnificus | GS1001 | Italy | OL960526 | - | - | - | - |
| E. permagnificus | GS1275 | Italy | OL960527 | - | - | - | - |
| E. permagnificus | MG558 | Italy | OL960528 | - | - | - | - |
| E. permagnificus | MG637 | Italy | OL960529 | - | - | - | - |
| E. permagnificus | MG829 | Italy | OL960530 | OL960502 | OL960510 | - | |
| E. permagnificus | ML61992EP | Cyprus | MH011858 | - | - | - | - |
| E. permagnificus | FR2011120 | France | KR782301 | - | - | - | - |
| E. ruber | KUN-HKAS 106891 | China | - | MN930518 | MT063123 | MT063120 | - |
| E. ruber | KUN-HKAS 103513 | China | - | MN930519 | MT063124 | MT063121 | - |
| E. ruber | KUN-HKAS 103122 | China | - | MN930520 | - | MT063122 | - |
| Rubroboletus sinicus | KUN-HKAS 68620 | China | KJ951991 | KF112319 | KF112146 | KF112661 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biketova, A.Y.; Gelardi, M.; Smith, M.E.; Simonini, G.; Healy, R.A.; Taneyama, Y.; Vasquez, G.; Kovács, Á.; Nagy, L.G.; Wasser, S.P.; et al. Reappraisal of the Genus Exsudoporus (Boletaceae) Worldwide Based on Multi-Gene Phylogeny, Morphology and Biogeography, and Insights on Amoenoboletus. J. Fungi 2022, 8, 101. https://doi.org/10.3390/jof8020101
Biketova AY, Gelardi M, Smith ME, Simonini G, Healy RA, Taneyama Y, Vasquez G, Kovács Á, Nagy LG, Wasser SP, et al. Reappraisal of the Genus Exsudoporus (Boletaceae) Worldwide Based on Multi-Gene Phylogeny, Morphology and Biogeography, and Insights on Amoenoboletus. Journal of Fungi. 2022; 8(2):101. https://doi.org/10.3390/jof8020101
Chicago/Turabian StyleBiketova, Alona Yu., Matteo Gelardi, Matthew E. Smith, Giampaolo Simonini, Rosanne A. Healy, Yuichi Taneyama, Gianrico Vasquez, Ádám Kovács, László G. Nagy, Solomon P. Wasser, and et al. 2022. "Reappraisal of the Genus Exsudoporus (Boletaceae) Worldwide Based on Multi-Gene Phylogeny, Morphology and Biogeography, and Insights on Amoenoboletus" Journal of Fungi 8, no. 2: 101. https://doi.org/10.3390/jof8020101