4.1. Fungal Macroinvaginations of the PM Are Tubular and Lamellar in Nature
As a result of the study, it was found that cells of xylotrophic basidiomycetes actively and abundantly form large PM invaginations. Presumably, filamentous fungi from other taxonomic and ecological-trophic groups also have this property. This issue requires additional research. Judging by the fact that preincubation of the mycelium in a liquid medium enhances the formation of invaginations, as well as by the initial lag of the formation of invaginations in some cases when the mycelium is transferred to the slides from a solid medium, it can be assumed that the PM of the mycelium immersed in the liquid invaginates most actively. However, it is difficult to say how mycelium growing in artificial and natural substrates, with a small amount of free liquid or with a separation of water and air phases, behaves in this respect in the depth of an agar medium or on its surface, in soil, in a woody substrate, etc.
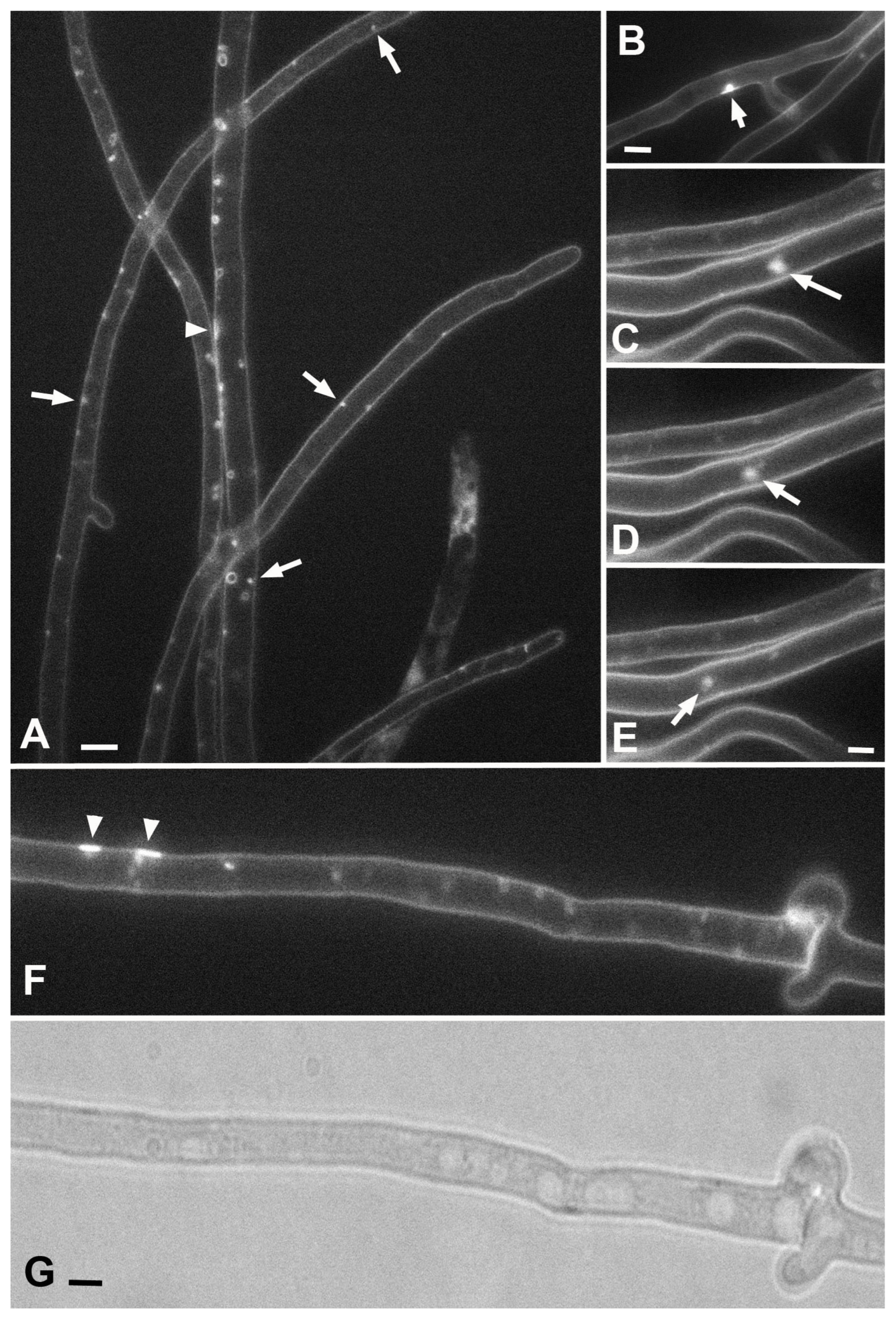
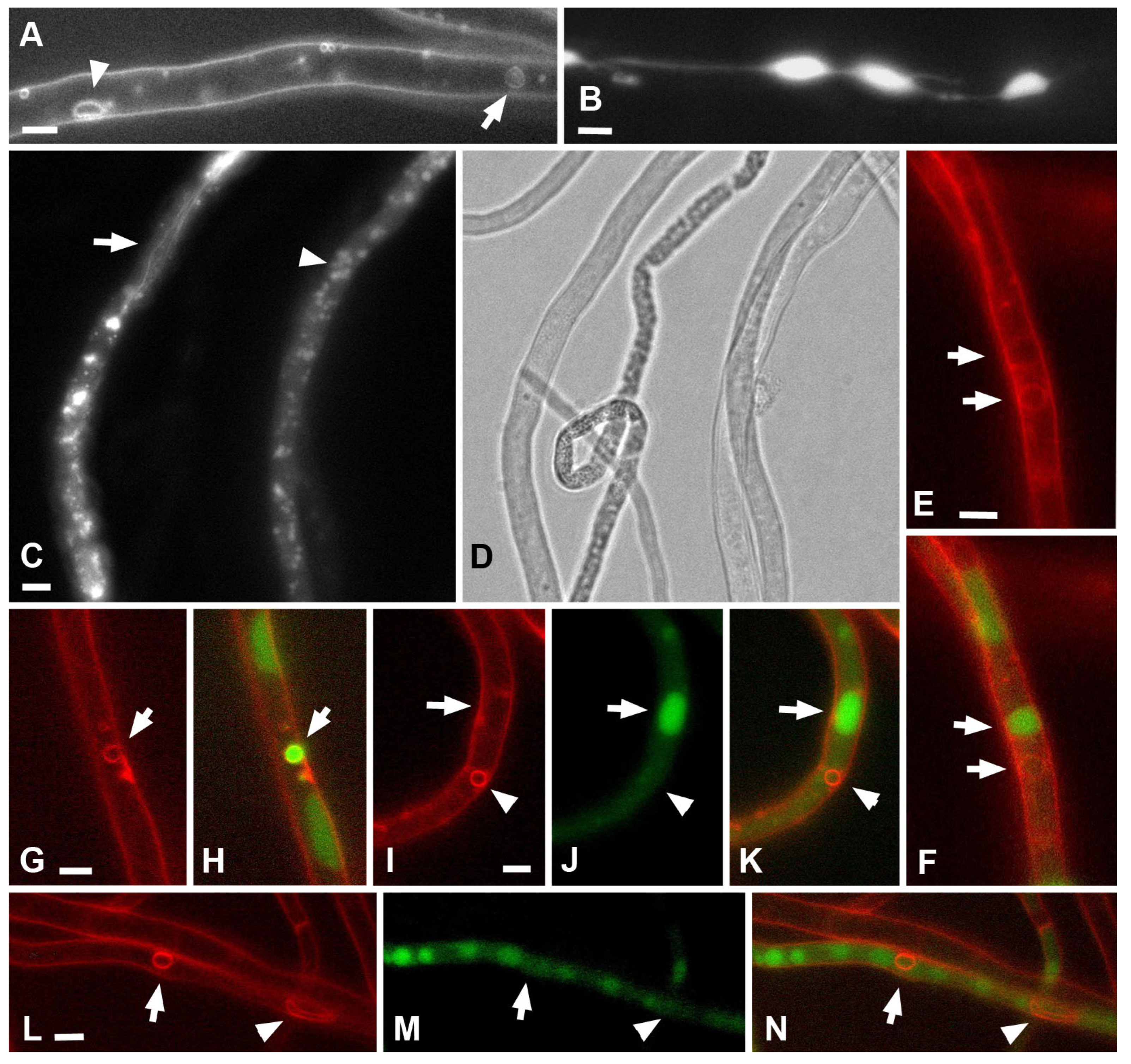
Undoubtedly, one of the two main conclusions of this study is that large PM invaginations in xylotrophs have a tubular or lamellar nature, and the thickness (lumen diameter) of the tubes/lamellae can be different, and the type of macroinvagination will depend on this. The mechanism for the formation of thin and thick tubes is obvious and is shown in the videos in this article. The formation of small or large vesicles occurs in two main ways. In the first way, vesicles are sections through transverse tubes and are visible as vesicles in a single focal section. In the second way, shown in
Videos S28 and S29, the tubules swell into a true vesicle. Apparently, there are still mechanisms for the formation of PM macroinvaginations of the vesicle type. For example, it can be through the wrapping of the lamella (a plicate of the PM) with the formation of a hollow roller transverse to the cell (perhaps
Video S14 demonstrates this). In any case, the main idea is that any macroinvagination passes through the stage of a tube (or lamella) during its formation. This idea extends to glomeruli and pendants (maybe also plaques), the mechanism of formation of which is not as obvious as that of tubes and vesicles.
We propose three main mechanisms for the formation of glomeruli and pendants (
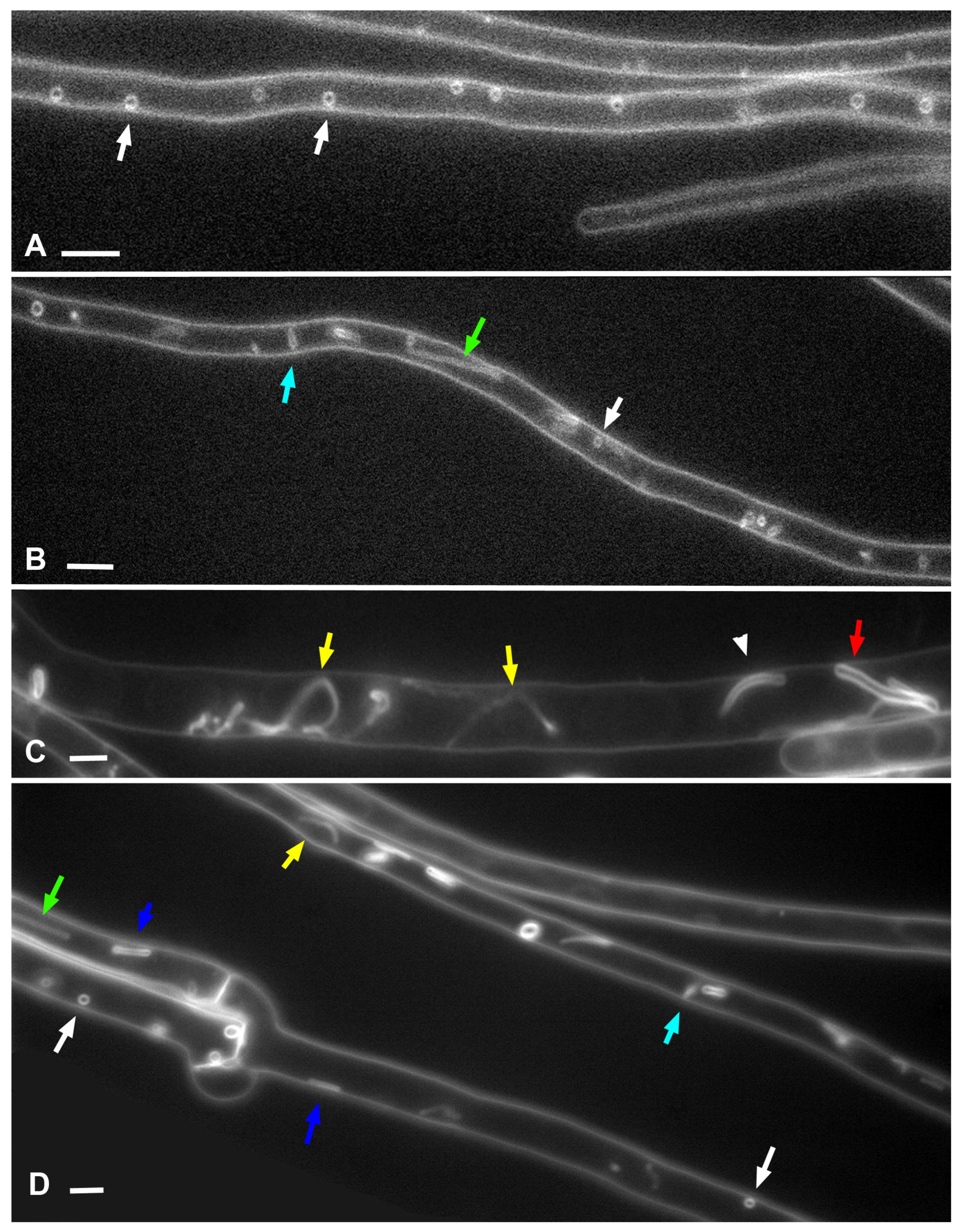
Figure 10). Here, electron microscopic images of lomasomes are important evidence. Precisely lomasomes, apparently, are visible at the fluorescent microscopic level as glomeruli and pendants. The first mechanism: a thin long filamentous tube is drawn into the cell (no more than 50 nm in cross-sectional diameter, most likely). It curls up into a glomerulus near the PM or at some distance (
Figure 10A–C). Accordingly, a true glomerulus or pendant is formed. Similar can be seen in
Figure 9B, as well as in a number of classic works (see Figures in [
14,
23]). The second mechanism: many classic vesicular lomasomes have a common outer membrane—they look like a parietal vacuole with vesicles or tubules inside (
Figure 9A, for example). It can be assumed that, in this case, a PM invagination tube is first formed, which swells into a vesicle (or it is a tube pressed to the PM initially). After that, from one or many sites of the membrane of such invagination, secondary invagination tubes begin to form in the lumen of the maternal invagination (
Figure 10D–F). They fill the lumen of the maternal macroinvagination chaotically (then vesicles and tubules are visible on the section of the lomasome,
Figure 9A top) or with a certain stacking (
Figure 9C,D, then, for example, only vesicles are visible on the section or regular alternation of vesicles and short tubules). The third mechanism: not a filamentous tube is drawn into the cell, but a thin lamella (less than 50 nm thick;
Figure 10G–H). Most often, such a lamella is located across the cell. This lamella then either folds into a tight roll and forms the classic myelin-like body or folds into a looser lomasome (
Figure 9E,G). On ultrathin sections, lamellar lomasomes consist of even concentric tubules (but in reality, they cannot be true tubules—the tubules cannot always be exactly in the plane of the section—they are sliced lamellas,
Figure 9F). In some of its parts, the lamella may also begin to swell and form secondary tubules-invaginations. Then a hybrid lomasome is obtained—vesicular-lamellar (
Figure 9A,C). Accordingly, such lamellar lomasomes are visible at the fluorescent microscopic level as glomeruli pressed against the PM and elongated across the cell.
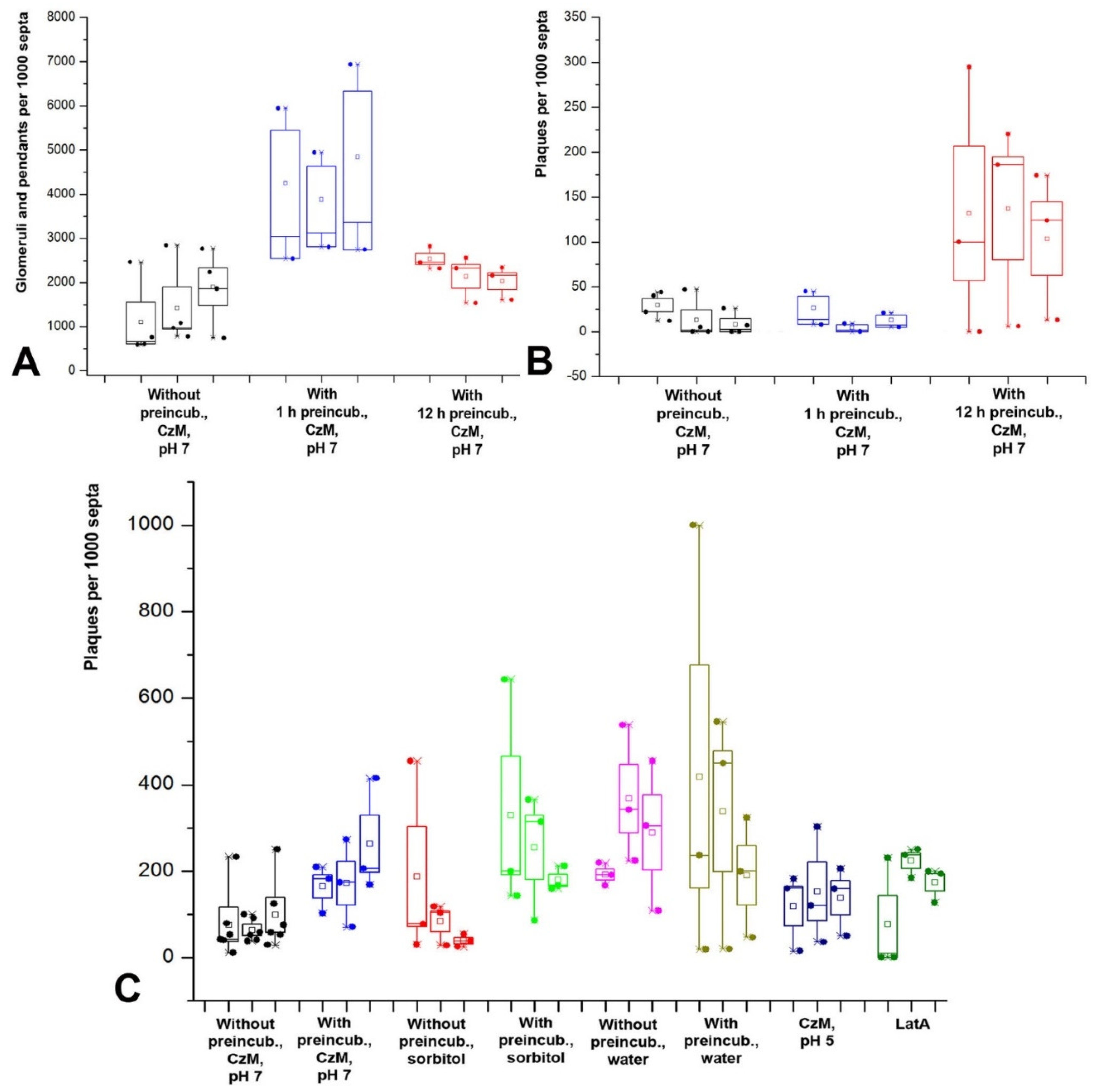
Separately, it is needed to consider the issue of plaques. Judging by the video in which other macroinvaginations flatten into plaques; judging by the increase in the number of plaques with a long incubation of the mycelium in a liquid medium paralleling the decrease in the number of other macroinvaginations. In addition, judging by the increase in the number of plaques in the hypophase, in which the tension of the PM increases, plaques are presumably glomeruli and small vesicles everted into the periplasmic space due to an increase in the PM tension and an increase in the hyphae diameter.
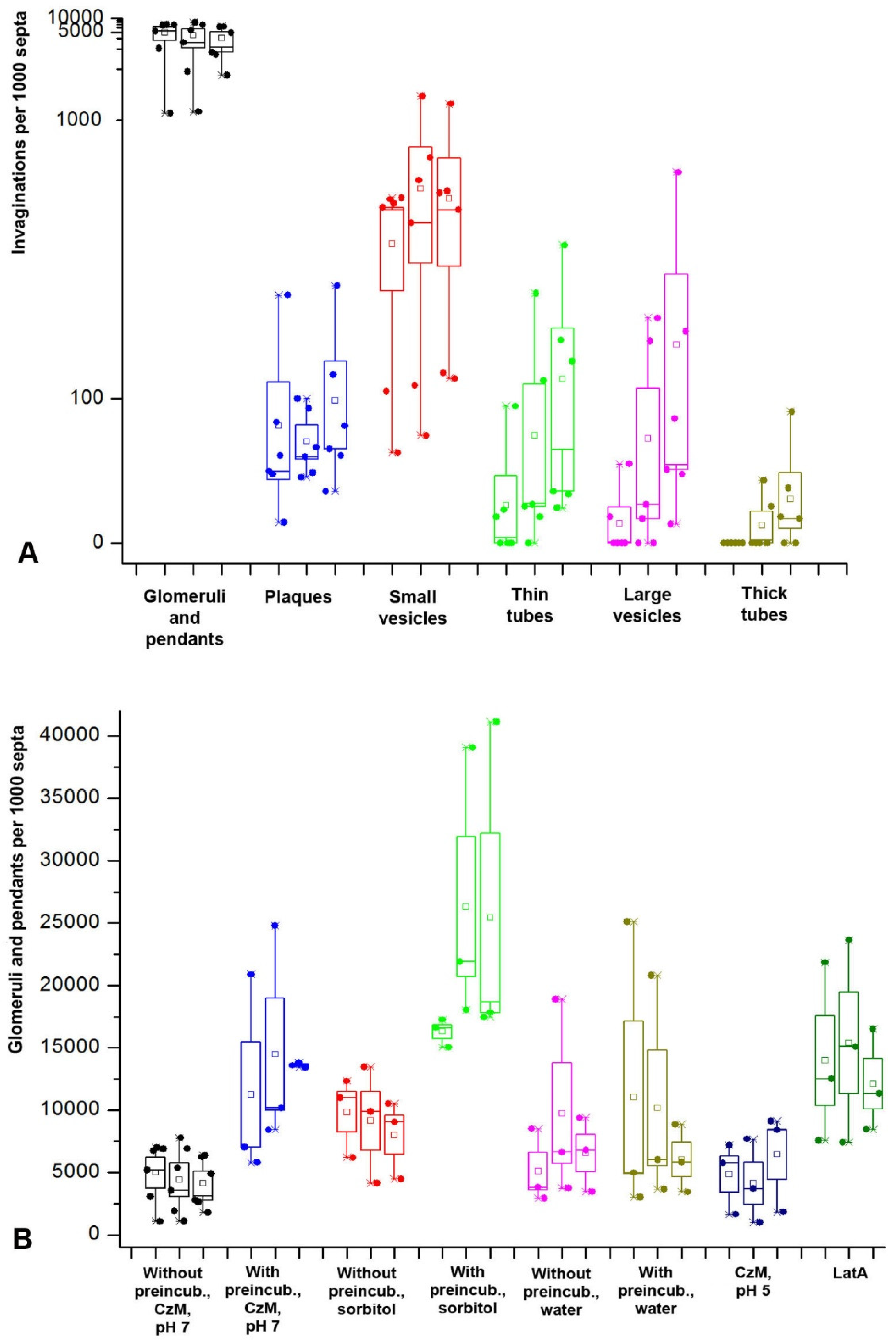
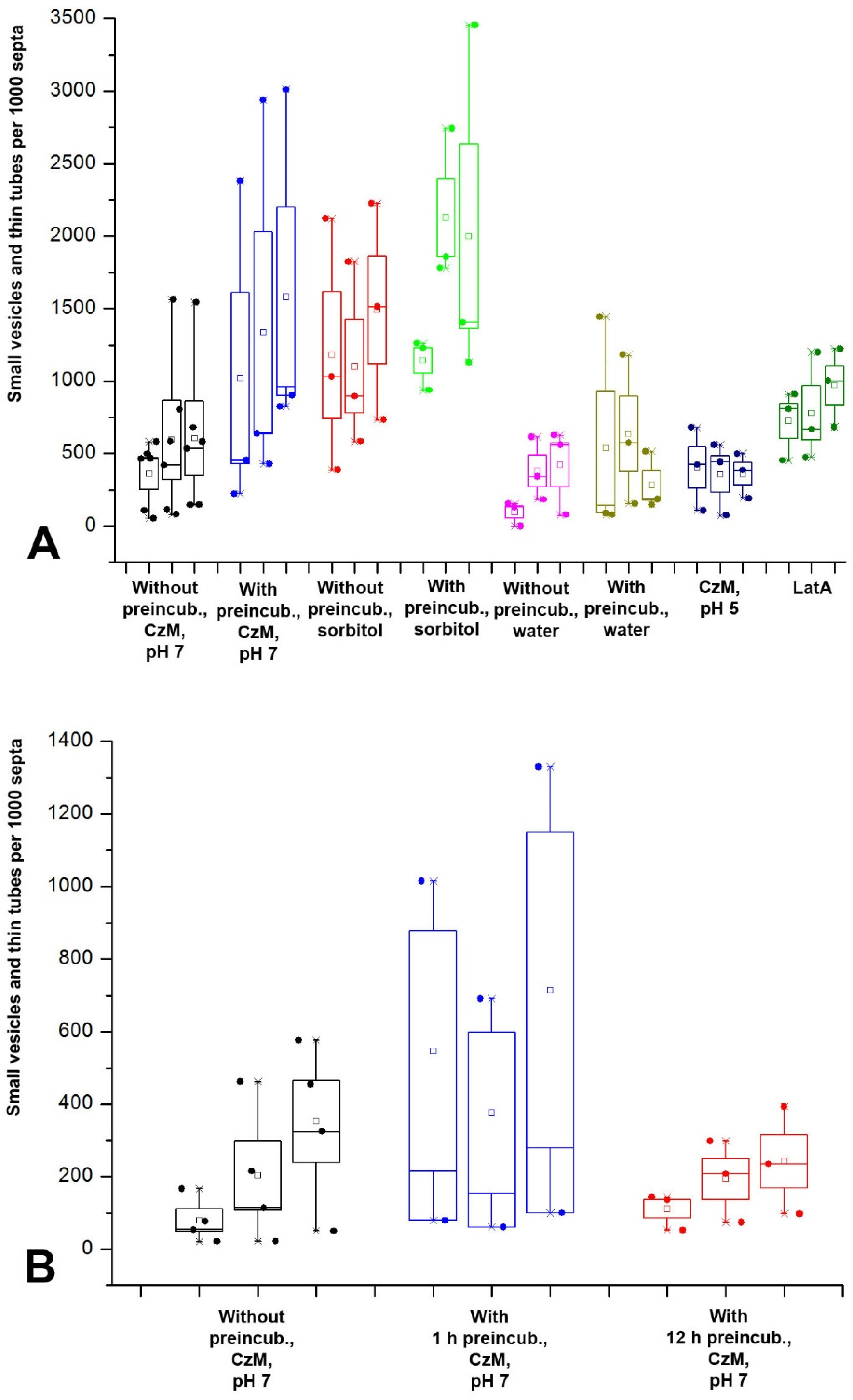
In the formation of fungal macroinvaginations of the PM, in addition to their tubular-lamellar nature, there are more regularities. The most important of them: in a microscopic slide, different types of macroinvaginations appear at different times after the preparation of the sample. There are two clear dependencies. First, the thicker the tubes from which the invagination is formed, the later such invagination is found in the microscopic slide. Therefore, the very first to appear in the slides are glomeruli and pendants, which are formed from very thin filamentous tubes and lamellae. Then, simultaneously or a little later, small vesicles appear, most often formed by transverse thin tubes. After 10–15 min, longitudinal thin tubes appear in the preparations. Finally, after 20–30 min, large vesicles and thick tubes can be found in individual hyphae cells. Moreover, the number of macroinvaginations per cell decreases along with an increase in the diameter of the forming tubes. Second, in the mycelium cells in freshly prepared slides, predominantly transverse tubules or lamellar rollers are formed. Longitudinal tubules, even thin ones, appear later.
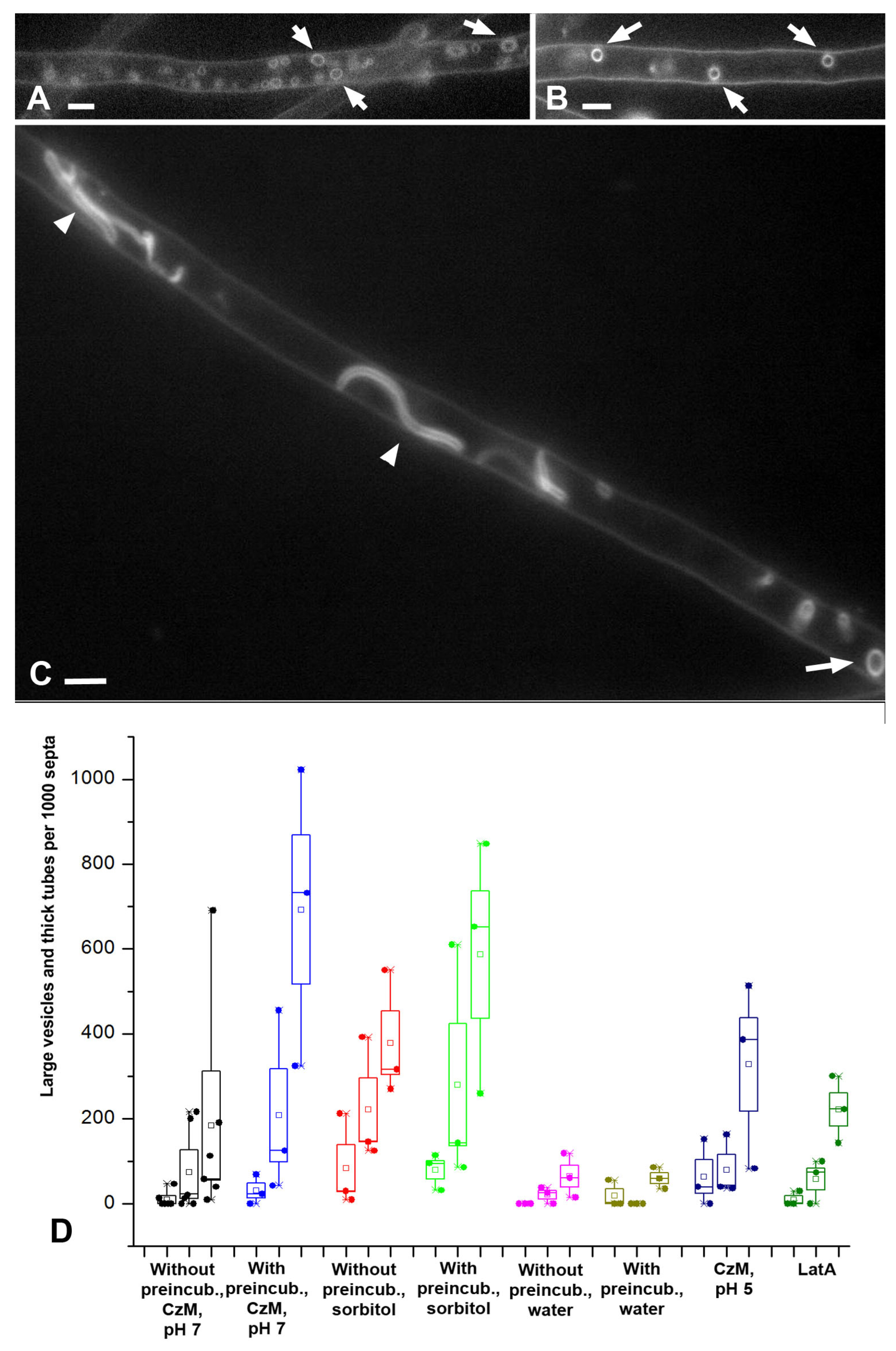
The successive appearance of different types of macroinvaginations in the samples is due to the specific conditions in the microscopic slides and the peculiarities of the control of the tension of the PM by the fungal cell. The preparation of a microscopic slide is accompanied by a strong mechanical effect on the fungal mycelium, and then mechanical stress continues—the pressure of the cover glass, the tension forces of a thin layer of liquid between the cover glass and slide, the movement of the liquid due to drying. Among other things, due to the drying of the slide, the osmotic pressure in the mounting fluid changes. However, here mechanical stress plays a greater role than osmotic changes; in preparations sealed with glue, evaporation in which is minimal, the dynamics of the formation of macroinvaginations do not change. Conversely, tapping the slides on the cover glass accelerates the formation of thick tubes and large vesicles (these observations of the authors are not presented in the Results). An important result should be added here: mounting the slides with a hyperosmotic medium enhances the formation of all types of macroinvaginations from glomeruli to thick tubes, but partial destruction of F-actin by LatA increases the number of only glomeruli/pendants and small vesicles, without affecting the longitudinal thin tubes, large vesicles, and thick tubes. All this can be explained as follows. In freshly prepared microscopic preparations under the conditions of this study, the control over the tension of the PM is performed mainly by the curtain actin subsystem (for the curtain model, see Introduction and [
7]). The fungus, using the system of actin driver cables, creates a certain tension of the PM, at which only tubular invaginations of small diameter are formed (most likely with the participation of BAR proteins). This probably roughly corresponds to the native state of the mycelium growing under stable conditions without significant stress. In this case, true glomeruli and pendants are formed, and lamellar rollers or tubes dominate, located across the cell and responsible for the appearance of elongated or pseudo-glomeruli, as well as small vesicles. The transverse arrangement of the rollers and tubes, apparently, is associated with the peculiarities of the tension of the PM (for example, it is not the same along and across the cell). Then, along with an increase in the time elapsed after the preparation of the slides due to mechanical stress, the curtain actin subsystem begins to reorganize (possibly, there is a partial disassembly of actin cables) and loses control over the tension of the PM. The tension of the PM in the cells decreases and its character changes—it becomes possible to form tubular invaginations of a larger diameter and location, including along the cells. It is not clear how the neck of such thick tubular invaginations is formed—it is unlikely that BAR proteins, dynamin, and other neck-forming proteins can control invagination of such a diameter (greater than 1 µm in the case of thick tubes). Perhaps the thick tubes start out as thin tubes and then expand.
4.2. AM4-64 Only Mimics the Endocytic Pathway: What about Classical Endocytosis?
In our previous works [
1,
20], studying static photographs of AM4-64 labeling taken in a single focal section in xylotrophs, we took glomeruli for primary endocytic invaginations/macrovesicles, pendants and small vesicles for endosomes, and large vesicles for vacuoles-lysosomes. The consistent appearance of the macroinvaginations in microscopic slides: glomeruli—small vesicles—and finally large vesicles—additionally confirmed that we were dealing with endocytic absorption of the styryl label and was in good agreement with the endocytic pathways described in the literature in yeast and some filamentous fungi [
16,
17,
18]. However, the change in methodical approaches in this study completely changed the interpretation of the previous and present results and made it possible to draw the second important conclusion of the study: AM4-64 in xylotrophic basidiomycetes marks mainly PM macroinvaginations of various types, only imitating the endocytic pathway due to the similarity of membrane structures and the non-simultaneity of the appearance of macroinvaginations in microscopic slides. This is confirmed by direct observation of the formation of invaginations using time-lapse video, tracking the connection of tubular invaginations with PM using 3D reconstructions of Z-stacks, and the absence of joint labeling of AM4-64 structures and the vacuolar system, etc. In this case, a natural question arises about classical (microvesicular) endocytosis in xylotrophic fungi. If everything that AM4-64 obviously labels in fungal cells has nothing to do with endocytosis, is there endocytic uptake of AM4-64, does it reach vacuole-lysosome tonoplasts by endocytosis?
The answer to this question requires further research. However, the presence of VLCs in xylotrophic cells indicates that classical endocytosis still occurs, not only in the subapical ring but also in non-apical hyphal cells. On the other hand, it is probably not as intense and fast as we previously thought. If the VLC tonoplast is not contaminated with the label from the PM or the diffuse label from the cytoplasm, then AM4-64 is transferred to it through a chain of primary endocytic vesicles and endosomes, which are difficult to trace against the background of a strong signal from macroinvaginations. Active transport of vesicles and endosomes along the hyphae is not ruled out: it is possible that the marginal hyphae of the colony, which were used for microscopy, transport endomembranes with the AM4-64 label to the basal part of the colony, where they already merge with vacuole-lysosomes.
4.4. Biological Functions of Fungal PM Macroinvaginations
It is most likely that fungal macroinvaginations of the PM are not an artifact. This also applies to large vesicles and thick tubes, despite the fact that they are presumably the response of fungus to the specific conditions of the microscope slide. Similar conditions can also occur in the natural environment, causing the formation of especially large PM invaginations.
Most of all, macroinvaginations of xylotrophs are similar to yeast eisosomes (especially lamellar invaginations), only they are significantly larger. Yeast eisosomes have been assigned different functions. Currently, the most popular concept is that yeast eisosomes are compartments of complexes of regulatory proteins involved in the regulation of yeast metabolism. Previously, it was assumed that eisosomes could take part in the regulation of PM tension, in particular, take in excess membrane during cell shrinkage. However, this hypothesis is currently not popular [
25]. However, it is the hypothesis of a reserve membrane pool and instantaneous packing of the PM membrane excess that is most adequate in the case of basidial xylotrophs. Consider the evidence supporting this hypothesis.
Our previous publication showed that hyphae of
Rhizoctonia solani under hypertonic conditions could shrink in cross-section with a loss of up to 15% in hyphae diameter [
7]. The cell wall of fungi is elastic, and the protoplast is attached to it from the inside through multiple sites of focal adhesion; therefore, when cells lose water, hyphae, like yeast cells, shrink entirely. Plasmolysis usually does not occur; the hyphae retain their shape and functionality. If to consider a fungal cell as a cylinder, then simple calculation shows that when the cylinder diameter decreases by 15%, the area of its lateral surface will also decrease by 15%. For example, the PM area of a fungal cell 100 µm long and 5 µm in diameter will be 1570 µm
2. When the cell is shrunk by 15% in diameter, the lateral area will be 1334.5 µm
2, i.e., it will decrease by 235.5 µm
2 (excluding possible longitudinal compression of the hyphae). To instantly remove the excess membrane of such an area, it is needed, for example, approximately 38 invagination tubes 5 µm long and 400 nm thick. These numbers are in good agreement with the results of this study. In other words, the most likely function of macroinvaginations in xylotrophs, and most likely in other filamentous fungi, is to provide a rapid change in the surface area of the PM without damaging the membranes. Under hypotonic conditions (in water), macroinvaginations are also formed, which indicates the existence of a reserve membrane pool that prevents cell destruction with a sharp increase in hyphae diameter.
Figure 3B demonstrates that in some cases, when the mycelium is placed in the hypophase, there may be outliers—a sharp increase in the number of glomeruli and pendants. Perhaps this is due to the delay in the establishment of osmotic equilibrium during the transfer of mycelium to the liquid due to the hydrophobic surface of the aerial mycelium growing in the air or water-air phase on cellophane in a Petri dish.
Fungal macroinvaginations may have other functions. For example, some time-lapse videos show the contact of thin tubes with large vacuoles (not shown). Such contact of the vacuole with the external environment can be an additional regulator of the osmotic pressure on the membranes. Previously, we assumed that lomasomes are actively involved in macrovesicular endocytosis and provide the fungus with intracellular digestion (receptor-independent bulk capture of nutrients by macroinvagination from the external environment and their lysis inside the detached invagination or after fusion with the vacuole-lysosome [
20]). This type of nutrition would give the basidiomycete an advantage in the rate of absorption of nutrients over the microorganisms of the hyphosphere. However, according to the present study, macroendocytosis in xylotrophs is irregular and is unlikely to be seriously involved in nutrition. On the other hand, under certain conditions in nature, macroendocytosis can become more active and contribute to nutrition, membrane metabolism, etc.
Thus, in the present study, the veil of one of the oldest mycological enigmas, lomasomes, has been lifted. With the help of fluorescent microscopic analysis, due to the capabilities of modern motorized epifluorescent microscopes, it was shown that basidial xylotrophs form many macroinvaginations of the PM, which are formed from tubes and lamellae of different thicknesses. The thinnest—filamentous tubes (lamellae) are folded into glomeruli and rollers (or otherwise transformed), forming lomasomes. Thicker invaginations do not fold but may kink, swell, etc. The activity of formation of macroinvaginations depends on the tension of the PM, which in turn is regulated by the actin cytoskeleton, and the thickness of the tubes that form macroinvaginations depends on the actin system and its integrity. Most likely, the macroinvaginations perform the function of conserving an excess of PM or a reserve membrane pool during changes in the thickness of hyphae (which probably occurs constantly and dynamically).
A new foreshortening of consideration of the membrane structures of the fungal cell has been established, in which large vesicular formations inside the hyphae represent two major groups: either large PM invaginations or vacuoles. Both groups are little involved in the classical endocytic pathway (only a small proportion of vacuoles are VLCs), but some lomasomes may be involved in specific macrovesicular endocytosis. The new perspective explains the plasticity inherent in fungal mycelium and its ability to quickly adapt to changing osmotic conditions, water-air phases, humidity, and so on. However, it also raises new biological questions: it is necessary to establish the intracellular localization of the membrane components of classical endocytosis in non-apical hyphal cells, functions of macrovesicular endocytosis, etc. An efficient solution to the problems associated with these biological issues requires complex methodological approaches that combine genetic labeling of membrane structures with fluorescent proteins and other labels, cytochemical labeling, and modern methods of light and electron microscopy.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









