
Phylogenetic Analyses of Hydnobolites and New Species from China
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
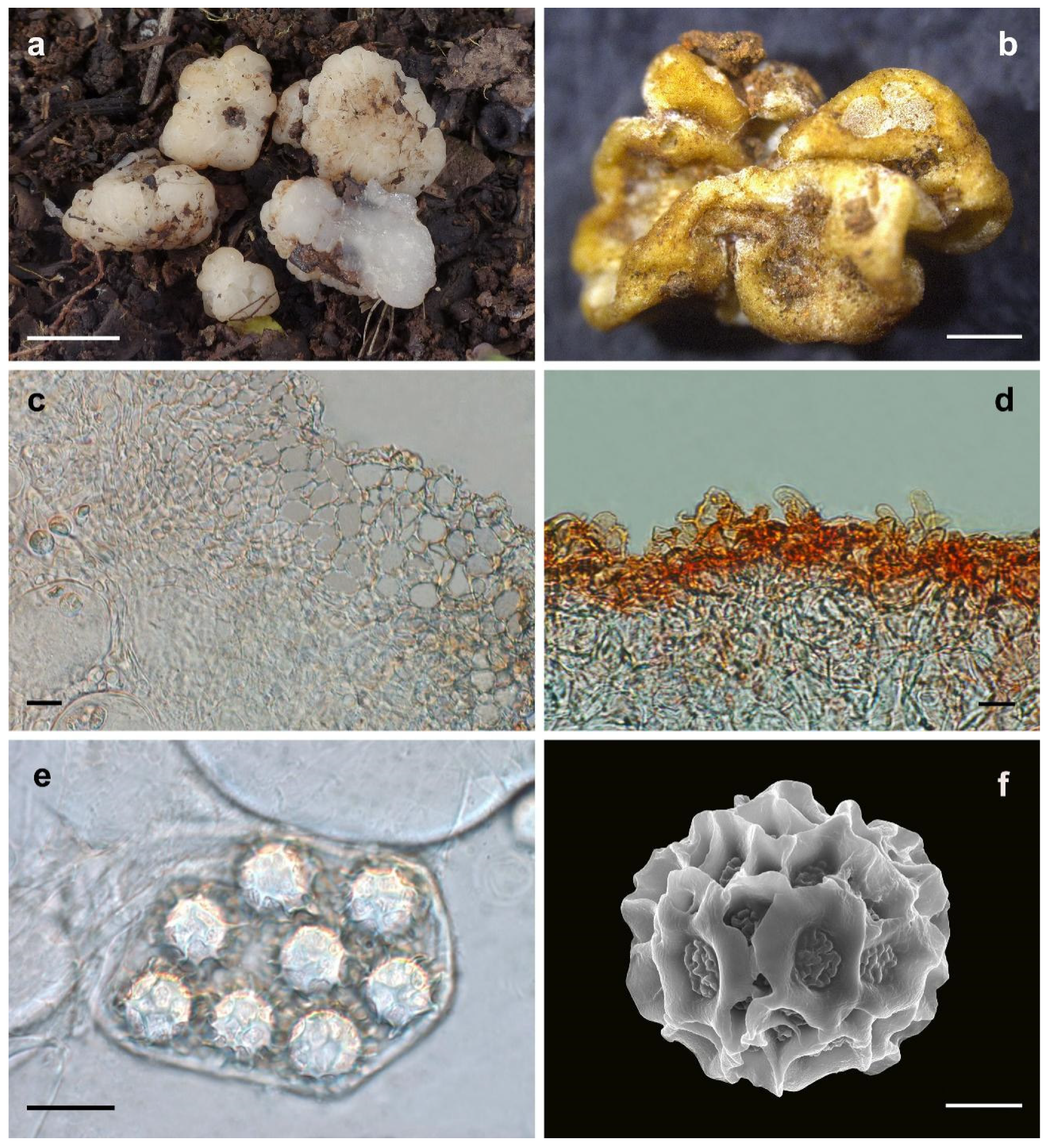
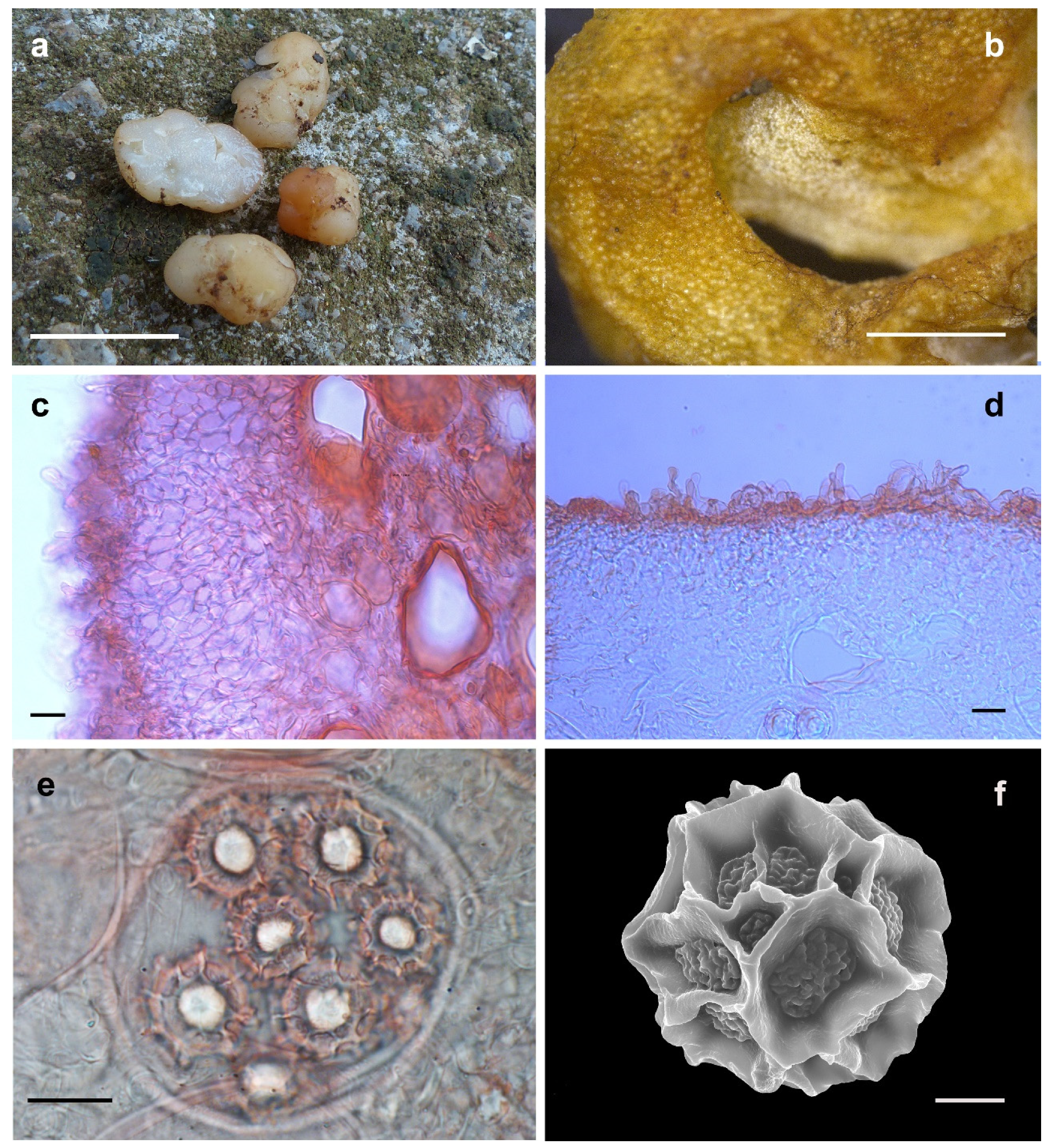
2.1. Sampling and Morphological Observations
2.2. Molecular Identification
2.3. Phylogenetic Analyses
3. Results
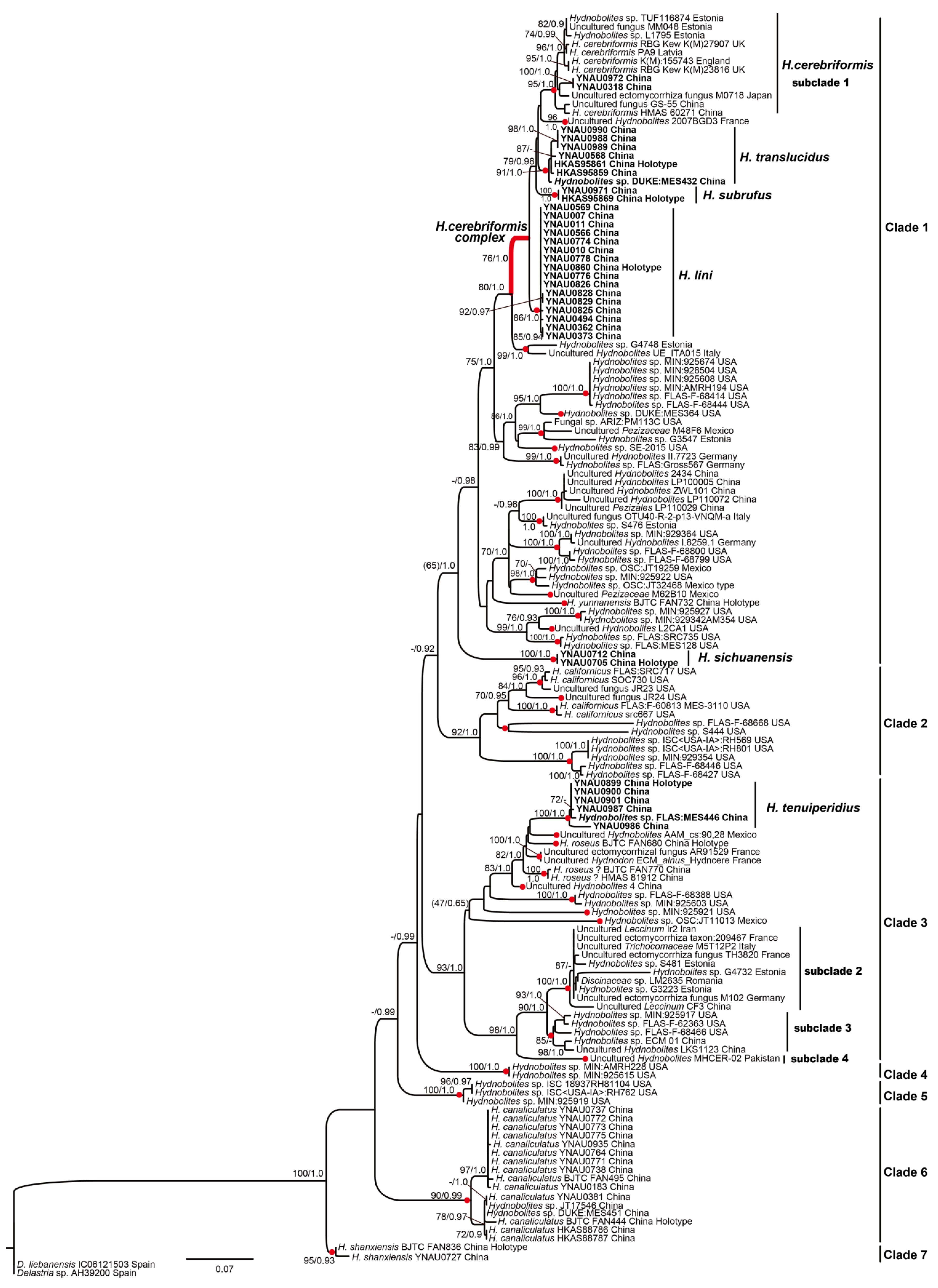
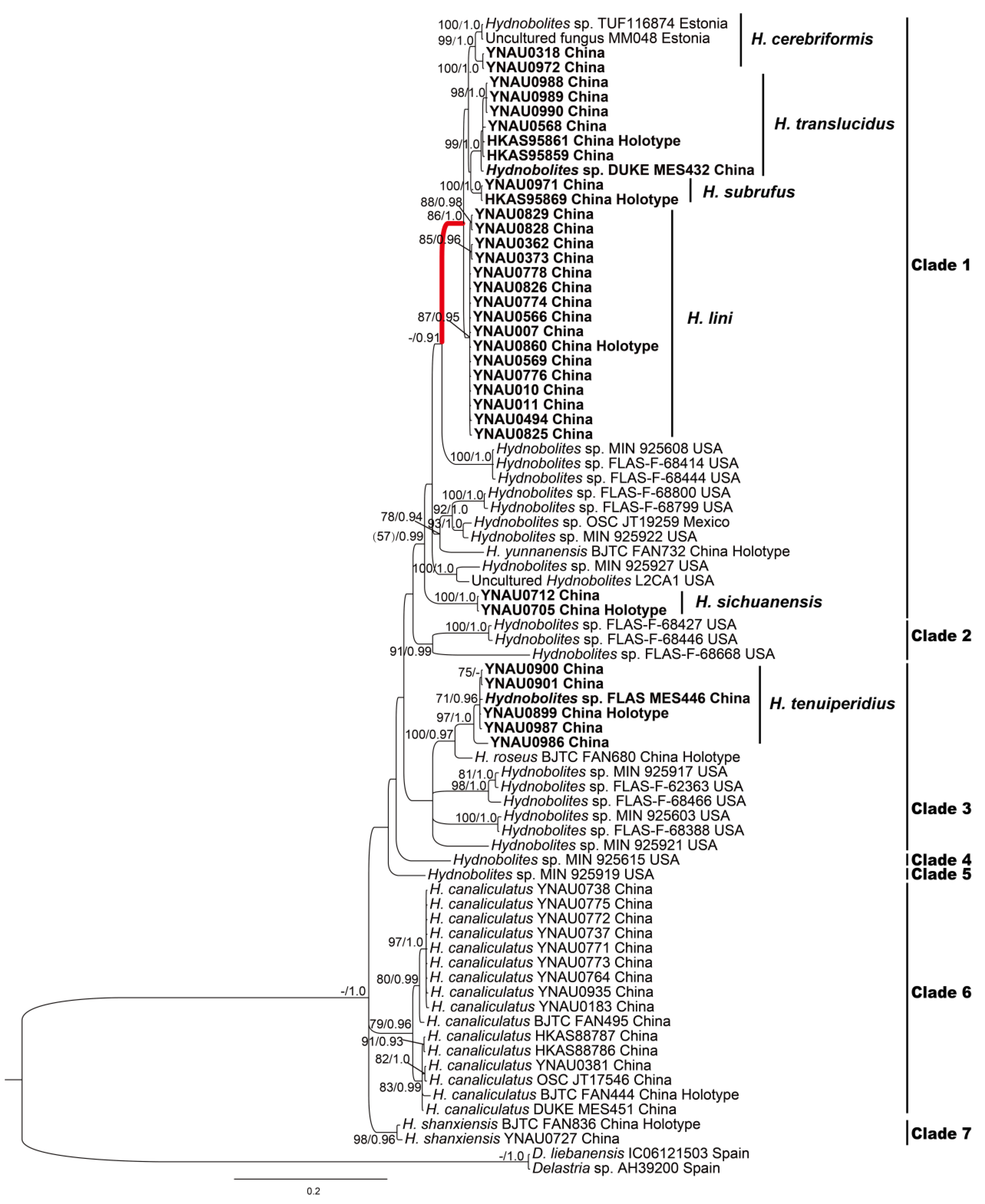
3.1. Molecular Phylogenetic Analyses
3.2. Taxonomy
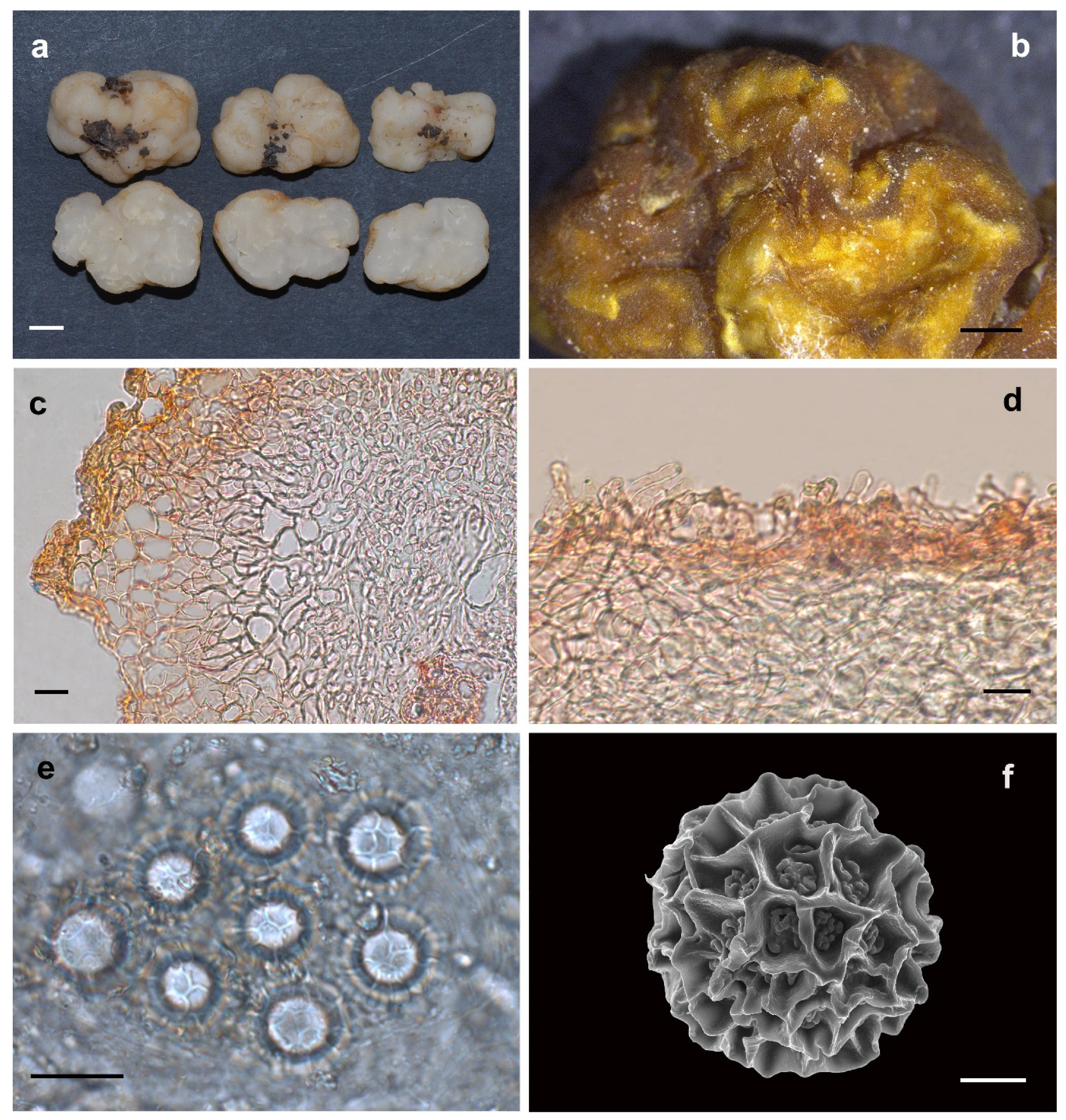
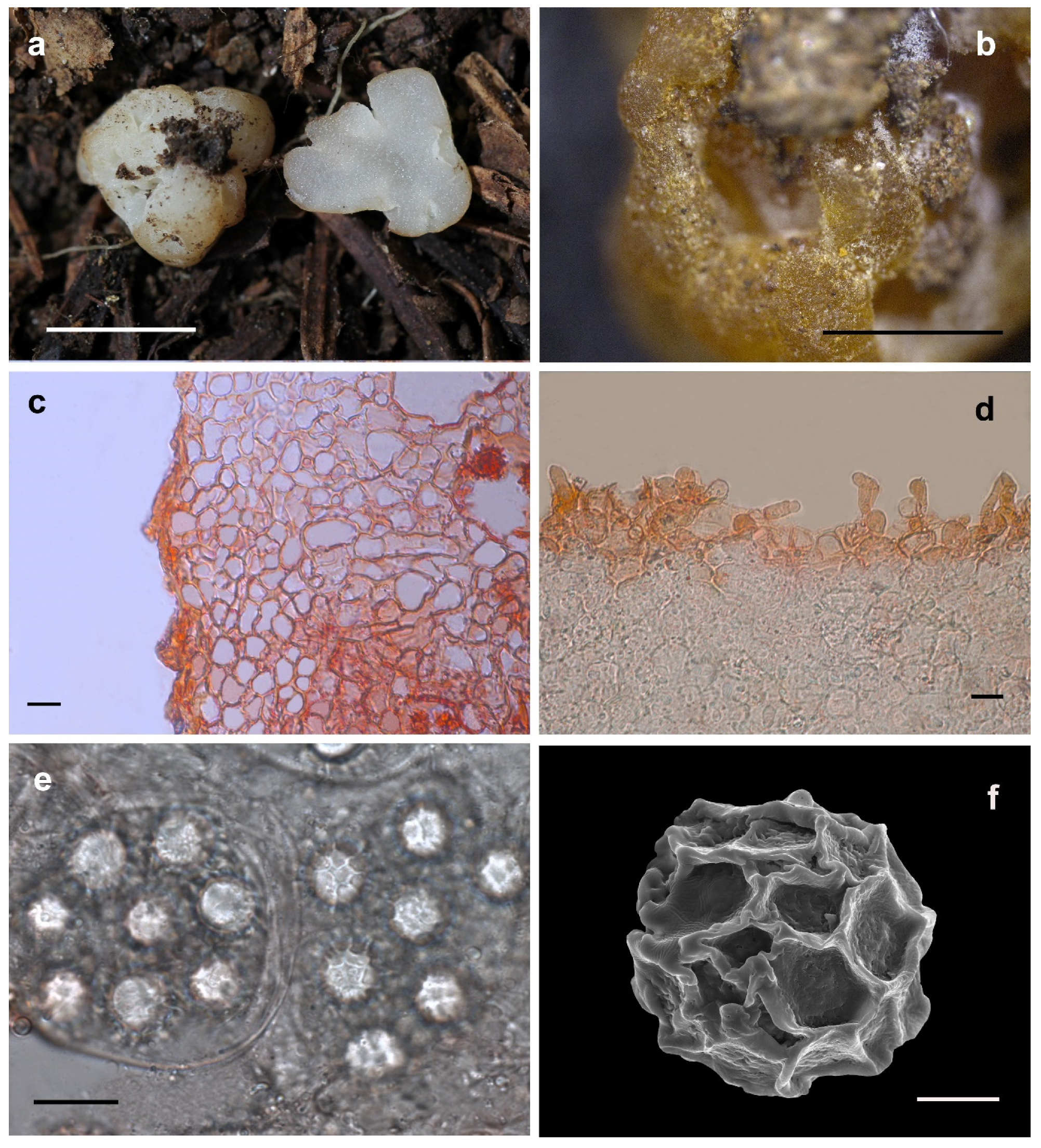
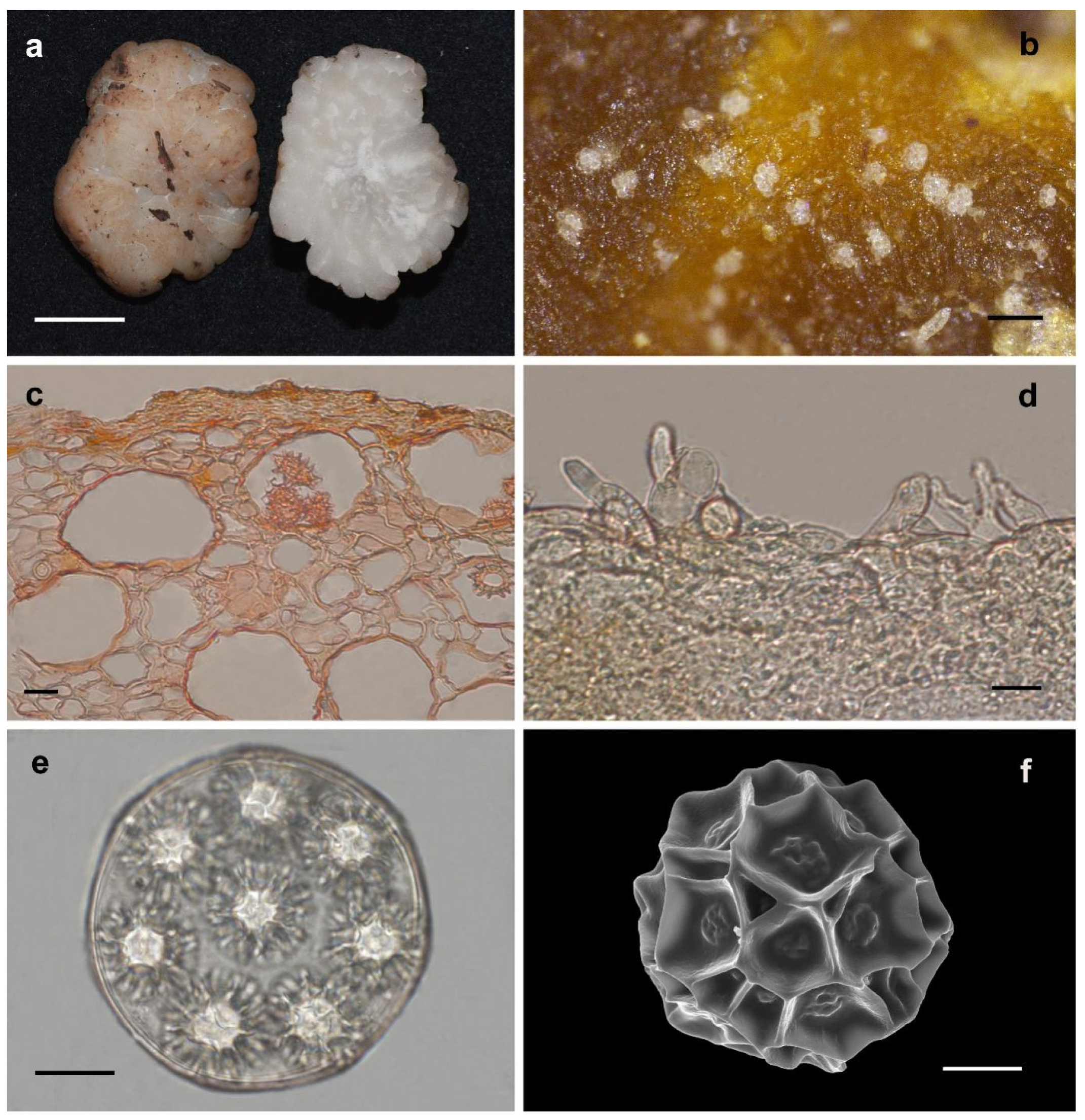
3.2.1. Hydnobolites Translucidus S.P. Wan and F.Q. Yu, sp. nov.
3.2.2. Hydnobolites subrufus S.P. Wan and F.Q. Yu, sp. nov.
3.2.3. Hydnobolites lini S.P. Wan and F.Q. Yu, sp. nov.
3.2.4. Hydnobolites sichuanensis S.P. Wan and F.Q. Yu, sp. nov.
3.2.5. Hydnobolites tenuiperidius S.P. Wan and F.Q. Yu, sp. nov.
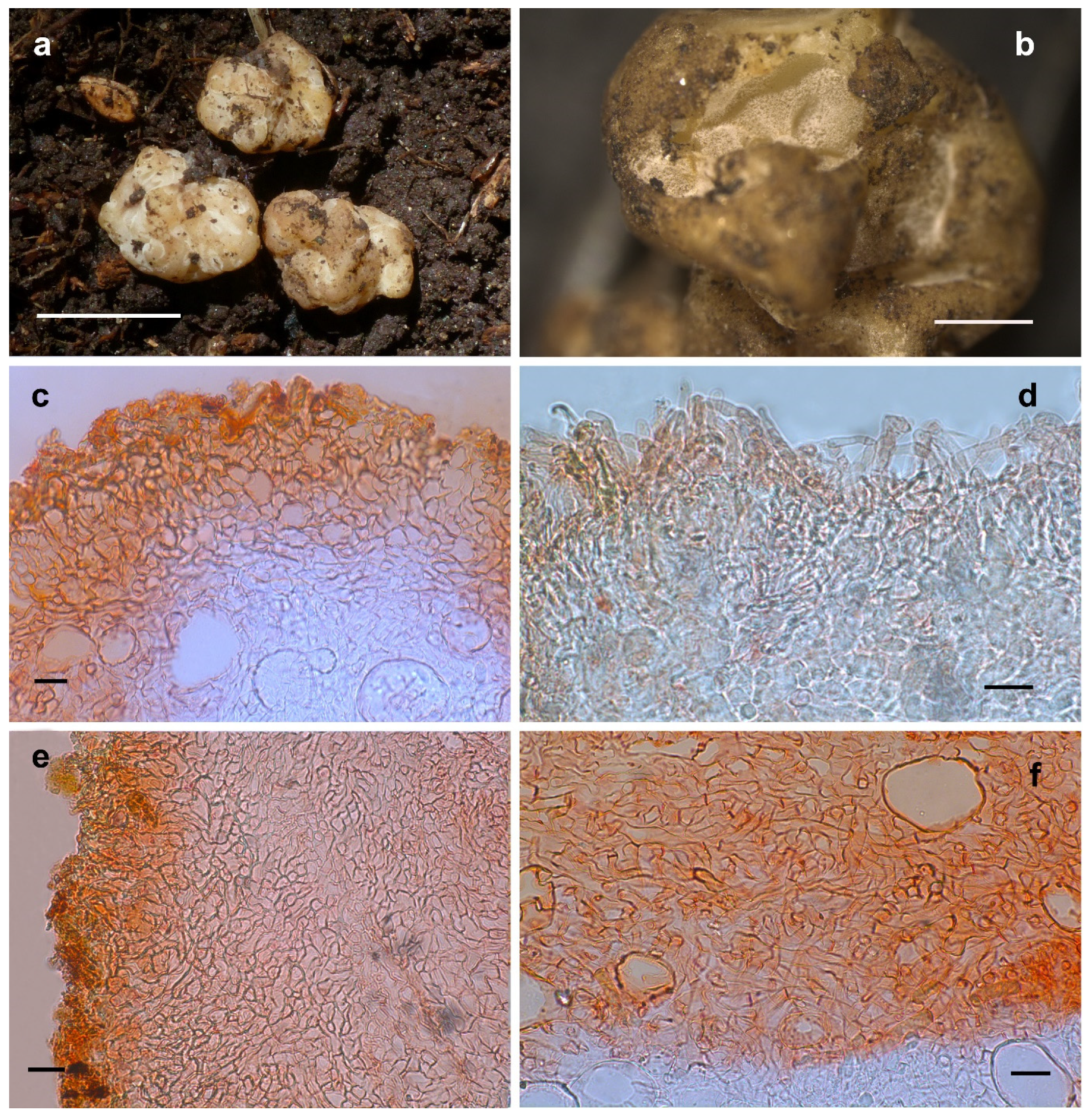
3.2.6. Hydnobolites cerebriformis Tul., Tul., Annales des Sciences Naturelles Botanique 19: 378 (1843) [1] (Figure 8)
4. Discussion
- 1.
- Ascomata whitish to yellowish or light brown.................................................................................................................................................2
- 1.
- Ascomata whitish to reddish...............................................................................................................................................................................3
- 2.
- Ascomata downy...................................................................................................................................................................................................4
- 2.
- Ascomata glabrous................................................................................................................................................................................................5
- 3.
- Asci or ascospores can be observed from the surface of ascomata................................................................................................................6
- 3.
- Asci or ascospores can not be observed from the surface of ascomata.........................................................................................................7
- 4.
- Ascospores have larger sizes (14.5–)16.0–23.0(–25.5) × (14.0–)15.5–23.0(–25.0)..................................................................H. translucidus
- 4.
- Ascospores have medium sizes (19–20–22 μm)..................................................................................................................H. cerebriformis [3]
- 4.
- Ascosporeshave smaller sizes (14–18 μm).............................................................................................................................H. californicus [2]
- 5.
- Ascospores have larger sizes (17.5–32.5 μm)....................................................................................................................H. canaliculatus [25]
- 5.
- Ascospores have medium sizes (18.75–26.25 μm)..............................................................................................................H. shanxiensis [25]
- 5.
- Ascospores have smaller sizes (17.5–25 μm).....................................................................................................................H. yunnanensis [25]
- 6.
- Ascospores have smaller sizes (13.3–22.0(–23.0) × (12.5–)–22.0(–22.6) μm).......................................................................H. tenuiperidius
- 6.
- Ascospores have larger sizes...............................................................................................................................................................................8
- 7.
- Ascomata downy, ascospores smaller (up to 21.0 × 20.5 μm)................................................................................................H. sichuanensis
- 7.
- Ascomata glabrous, ascospores larger (up to 30 μm)..................................................................................................................H. roseus [25]
- 8.
- Ascomata have obvious red color.....................................................................................................................................................H. subrufus
- 8.
- Ascomata white to slightly reddish...........................................................................................................................................................H. lini
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tulasne, L.R.; Tulasne, C. Champignons hypogés de la famille des Lycoperdacées, observés dans les environs de Paris et les départements de la Vienne et d’lndre-et-Loire. Ann. Des Sci. Nat. Série 1843, 2, 373–381. [Google Scholar]
- Gilkey, H.M. A Revision of the Tuberales of California; University of California Publications in Botany: Botany, CA, USA, 1916; Volume 6, pp. 275–356. [Google Scholar]
- Hawker, L.E. British hypogeous fungi. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1954, 237, 429–546. [Google Scholar] [CrossRef]
- Castellano, M.A.; Trappe, J.M.; Maser, Z.; Maser, S. Key to Spores of the Genera of Hypogeous Fungi of North America, with Reference to Animal Mycophagy; Mad River Press: Eureka, CA, USA, 1986; p. 185. [Google Scholar]
- Trappe, J.M. The orders, families, and genera of hypogeous Ascomycotina (truffles and their relatives). Mycotaxon 1979, 9, 297–340. [Google Scholar]
- Pegler, D.N.; Spooner, B.M.; Young, T.W.K. British truffles: A Revision of British Hypogeous Fungi; Royal Botanic Gardens: Kew, UK, 1993; p. 216. [Google Scholar]
- Tulasne, L.R.; Tulasne, C. Fungi Hypogæi. Histoire et Monographie des Champignon Hypogés; Apud Friedrich Klincksieck: Paris, France, 1851; pp. 123–125. [Google Scholar]
- Gilkey, H.M. Taxonomic notes on Tuberales. Mycologia 1955, 46, 783–793. [Google Scholar] [CrossRef]
- Korf, R.P. Discomycetes and Tuberales. In The Fungi: An Advanced Treatise; Ainsworth, G.C., Sparrow, F.K., Sussman, A.S., Eds.; Academic Press: New York, NY, USA, 1973; Volume IVA, pp. 249–319. [Google Scholar]
- Korf, R.P. Sparassoid ascomata in Pezizales and Tuberales. Repert. Tottori Mycol. Inst. 1973, 10, 389–403. [Google Scholar]
- Castellano, M.A.; Trappe, J.M.; Luoma, D.L. Sequestrate fungi. In Biodiversity of Fungi: Inventory and Monitoring Methods; Mueller, G.M., Bills, G.F., Foster, M.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; pp. 197–213. [Google Scholar]
- Læssøe, T.; Hansen, K. Truffle trouble: What happened to the Tuberales? Mycol. Res. 2007, 111, 1075–1099. [Google Scholar] [CrossRef]
- Fischer, E. Tuberineae, Plectascineae. In Die natürlichen Pflanzenfamilien; Engler, A., Prantl, K., Eds.; Verlag von Wilhelm Engelmann: Leipzig, Germany, 1897; pp. 278–320. [Google Scholar]
- Fischer, E. Tuberaceen und Hemiasceen. In Kryptogamen Flora Deutschlands; Österreich und der Schweiz; Rabenhorst, L., Ed.; Verlag-Kummer: Leipzig, Germany, 1897; pp. 3–81. [Google Scholar]
- Trappe, J.M. A synoptis of the Carbomycetaceae and Terfeziaceae (Tuberales). Trans. Br. Mycol. Soc. 1971, 57, 85–92. [Google Scholar] [CrossRef]
- Pfister, D.H. Genea-Jafneadelphus- a Tuberalean-Pezizalean connection. Mycologia 1984, 76, 170–172. [Google Scholar] [CrossRef]
- Kimbrough, J.W.; Wu, C.G.; Gibson, J.L. Ultrastructural evidence for a phylogenetic linkage of the truffle genus Hydnobolites to the Pezizaceae (Pezizales, Ascomycetes). Bot. Gaz. 1991, 152, 408–420. [Google Scholar] [CrossRef]
- Kimbrough, J.W. Septal ultrastructure and ascomycete systematics. In Ascomycete Systematics: Problems and Perspectives in the Nineties; Hawksworth, D.L., Sutton, B.C., Eds.; Plenum Press: New York, NY, USA, 1994; pp. 127–141. [Google Scholar]
- Tedersoo, L.; May, T.W.; Smith, M.E. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 2010, 20, 217–263. [Google Scholar] [CrossRef]
- Kovács, G.M.; Trappe, J.M.; Alsheikh, A.M.; Hansen, K.; Healy, R.A.; Vági, P. Terfezia disappears from the American truffle mycota as two new genera and Mattirolomyces species emerge. Mycologia 2011, 103, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Healy, R.A.; Smith, M.E.; Bonito, G.M.; Pfister, D.H.; Ge, Z.W.; Guevara, G.G.; Williams, G.; Stafford, K.; Kumar, L.; Lee, T.; et al. High diversity and widespread occurrence of mitotic spore mats in ectomycorrhizal Pezizales. Mol. Ecol. 2013, 22, 1717–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfister, D.H.; Healy, R. Pezizomycetes. Encycl. Mycol. 2021, 1, 295–309. [Google Scholar]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth and Bisby’s Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008; p. 325. [Google Scholar]
- Wu, E.; Lan, Z.; Xiang, Y.; Han, Z. A new species of Tuberales from China. Mycol. Res. 1993, 97, 1021–1022. [Google Scholar] [CrossRef]
- Li, T.; Cheng, M.; Xu, Y.Y.; Fan, L. Four new species of Hydnobolites (sequestrate Pezizaceae, Ascomycota) from China. Mycol. Prog. 2019, 18, 405–414. [Google Scholar] [CrossRef]
- Smith, M.E.; Schmull, M. Tropical truffles: English translation and critical review of F. von Hhnel’s truffles from Java. Mycol. Prog. 2011, 10, 249–260. [Google Scholar] [CrossRef]
- Xu, A.S. Additional hypogeous fungi in Xizang. In Proceedings of the AnnMSA Conference, Beijing, China, 1 April 2003; p. 445. [Google Scholar]
- Trappe, J.M.; Molina, R.; Luoma, D.L.; Cázares, E.; Pilz, D.; Smith, J.E.; Castellano, M.A.; Miller, S.L.; Trappe, M.J. Diversity, Ecology and Conservation of the Truffle Fungi in Forests of the Pacific Northwest; Forest Service General Technical Report, PNW-GTR-772; US Department of Agriculture: Portland, OR, USA, 2009; p. 194. [Google Scholar]
- Smith, M.E.; Douhan, G.W.; Rizzo, D.M. Ectomycorrhizal Community Structure in a Xeric Quercus Woodland Based on rDNA Sequence Analysis of Sporocarps and Pooled Roots. New Phytol. 2007, 174, 847–863. [Google Scholar] [CrossRef]
- Morris, M.H.; Smith, M.E.; Rizzo, D.M.; Rejmanek, M.; Bledsoe, C.S. Contrasting ectomycorrhizal fungal communites on the roots of co-occuring oaks (Quercus spp.) in a California woodland. New Phytol. 2008, 178, 167–176. [Google Scholar] [CrossRef]
- Tedersoo, L.; Suvi, T.; Jairus, T.; Kõljalg, U. Forest microsite effects on community composition of ectomycorrhizal fungi on seedlings of Picea abies and Betula pendula. Environ. Microbiol. 2008, 10, 1189–1201. [Google Scholar] [CrossRef]
- Lang, C.; Seven, J.; Polle, A. Host preferences and differential contributions of deciduous tree species shape mycorrhizal species richness in a mixed central European forest. Mycorrhiza 2011, 21, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Medel, R.; Castillo, R.; Marmolejo, J.; Baeza, Y. Análisis de la familia Pezizaceae (Pezizales: Ascomycota) en México. Rev. Mex. Biodivers. 2013, 84, 21–38. [Google Scholar] [CrossRef] [Green Version]
- Long, D.; Liu, J.; Han, Q.; Wang, X.; Huang, J. Ectomycorrhizal fungal communities associated with Populus simonii and Pinus tabuliformis in the hilly-gully region of the loess plateau, China. Sci. Rep. 2016, 6, 24336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klavina, D.; Pennanen, T.; Gaitnieks, T.; Velmala, S.; Lazdins, A.; Lazdina, D.; Menkis, A. The ectomycorrhizal community of conifer stands on peat soils 12 years after fertilization with wood ash. Mycorrhiza 2016, 26, 153–160. [Google Scholar] [CrossRef]
- Obase, K.; Yamanaka, S.; Yamanaka, T.; Ozaki, K. Short-Term Effects of Retention Forestry on the Diversity of Root-Associated Ectomycorrhizal Fungi in Sakhalin Fir Plantations, Hokkaido, Japan. For. Ecol. Manag. 2022, 523, 120501. [Google Scholar] [CrossRef]
- Yang, Z.L.; Zhang, L.F. Type studies on Clitocybe macrospora and Xerula furfuracea var. bispora. Mycotaxon 2003, 88, 447–454. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user–friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Automat. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest 2.3. Program Distributed by the Author; Evolutionary biology Center, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Bidartondo, M.I.; Read, D.J. Fungal specificity bottlenecks during orchid germination and development. Mol. Ecol. 2008, 17, 3707–3716. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Mett, M.; Ishida, T.A.; Bahram, M. Phylogenetic relationships among host plants explain differences in fungal species richness and community composition in ectomycorrhizal symbiosis. New Phytol. 2013, 199, 822–831. [Google Scholar] [CrossRef] [PubMed]
- Brock, P.M.; Döring, H.; Bidartondo, M.I. How to know unknown fungi: The role of a herbarium. New Phytol. 2009, 181, 719–724. [Google Scholar] [CrossRef] [Green Version]
- Alvarado, P.; Moreno, G.; Manjón, J.L.; Gelpi, C.; Kaounas, V.; Konstantinidis, G.; Barseghyan, G.S.; Venturella, G. First molecular data on Delastria rosea, Fischerula macrospora and Hydnocystis piligera. Boln. Soc. Micol. Madrid. 2011, 35, 31–37. [Google Scholar]
- Leonardi, M.; Iotti, M.; Oddis, M.; Lalli, G.; Pacioni, G.; Leonardi, P.; Maccherini, S.; Perini, C.; Salerni, E.; Zambonelli, A. Assessment of ectomycorrhizal fungal communities in the natural habitats of Tuber magnatum (Ascomycota, Pezizales). Mycorrhiza 2013, 23, 349–358. [Google Scholar] [CrossRef]
- Roy, M.; Rochet, J.; Manzi, S.; Gardes, M. What determines Alnus-associated ectomycorrhizal community diversity and specificity? A comparison of host and habitat effects at a regional scale. New Phytol. 2013, 198, 1228–1238. [Google Scholar] [CrossRef]
- Zhang, J.; Taniguchi, T.; Xu, M.; Du, S.; Liu, G.B.; Yamanaka, N. Ectomycorrhizal fungal communities of Quercus liaotungensis along different successional stands on the Loess Plateau, China. J. For. Res. Jpn. 2014, 19, 395–403. [Google Scholar] [CrossRef]
- Benucci, G.M.N.; Raggi, L.; Albertini, E.; Csorbai, A.G.; Donnini, D. Assessment of ectomycorrhizal biodiversity in Tuber macrosporum productive sites. Mycorrhiza 2014, 24, 281–292. [Google Scholar] [CrossRef]
- Erlandson, S.R.; Savage, J.A.; Cavender-Bares, J.M.; Peay, K.G. Soil moisture and chemistry influence diversity of ectomycorrhizal fungal communities associating with willow along an hydrologic gradient. FEMS Microbiol. Ecol. 2015, 92, fiv148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argüelles-Moyao, A.; Garibay-Orijel, R.; Arellano-Torres, E. Clavulina-Membranomyces is the most important lineage within the highly diverse ectomycorrhizal fungal community of Abies religiosa. Mycorrhiza 2016, 27, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.B.; Liu, J.J.; Long, D.F.; Han, Q.S.; Huang, J. The ectomycorrhizal fungal communities associated with Quercus liaotungensis in different habitats across northern China. Mycorrhiza 2017, 27, 441–449. [Google Scholar] [CrossRef]
- Morris, M.H.; Perez-Perez, M.A.; Smith, M.E.; Bledsoe, C.S. Influence of host species on ectomycorrhizal communities associated with two co-occurring oaks (Quercus spp.) in a tropical cloud forest. FEMS Microbiol. Ecol. 2009, 69, 274–287. [Google Scholar] [CrossRef] [PubMed]
- Belfiori, B.; Riccioni, C.; Tempesta, S.; Pasqualetti, M.; Paolocci, F.; Rubini, A. Comparison of ectomycorrhizal communities in natural and cultivated Tuber melanosporum truffle grounds. FEMS Microbiol. Ecol. 2012, 81, 547–561. [Google Scholar] [CrossRef]
- Bowman, E.A.; Arnold, A.E. Distributions of ectomycorrhizal and foliar endophytic fungal communities associated with Pinus ponderosa along a spatially constrained elevation gradient. Am. J. Bot. 2018, 105, 687–699. [Google Scholar] [CrossRef] [Green Version]
- Suz, L.M.; Barsoum, N.; Benham, S.; Dietrich, H.P.; Fetzer, K.D.; Fischer, R.; García, P.; Gehrman, J.; Kristöfel, F.; Manninger, M.; et al. Environmental drivers of ectomycorrhizal communities in Europe’s temperate oak forests. Mol. Ecol. 2014, 23, 628–5644. [Google Scholar] [CrossRef]
- Paz, A.; Lavoise, C.; Chautrand, P.; Moreau, P.A.; Bellanger, J.M. The genus Delastria (Pezizaceae), a worldwide revision. Ascomytete. Org. 2018, 10, 229–243. [Google Scholar] [CrossRef]
- Richard, F.; Roy, M.; Shahin, O.; Sthultz, C.; Duchemin, M.; Joffre, R.; Selosse, M.A. Ectomycorrhizal communities in a Mediterranean forest ecosystem dominated by Quercus ilex: Seasonal dynamics and response to drought in the surface organic horizon. Ann. For. Sci. 2011, 68, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Bahram, M.; Põlme, S.; Kõljalg, U.; Zarre, S.; Tedersoo, L. Regional and local patterns of ectomycorrhizal fungal diversity and community structure along an altitudinal gradient in the Hyrcanian forests of northern Iran. New Phytol. 2012, 193, 465–473. [Google Scholar] [CrossRef]
- Cázares, E.; García, J.; Castillo, J.; Trappe, J.M. Hypogeous fungi from northern Mexico. Mycologia 1992, 84, 341–359. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, S.-P.; Huang, L.-L.; Cui, M.-J.; Yu, C.-J.; Liu, W.; Wang, R.; Shi, X.-F.; Yu, F.-Q. Phylogenetic Analyses of Hydnobolites and New Species from China. J. Fungi 2022, 8, 1302. https://doi.org/10.3390/jof8121302
Wan S-P, Huang L-L, Cui M-J, Yu C-J, Liu W, Wang R, Shi X-F, Yu F-Q. Phylogenetic Analyses of Hydnobolites and New Species from China. Journal of Fungi. 2022; 8(12):1302. https://doi.org/10.3390/jof8121302
Chicago/Turabian StyleWan, Shan-Ping, Lan-Lan Huang, Meng-Jin Cui, Cheng-Jin Yu, Wei Liu, Rui Wang, Xiao-Fei Shi, and Fu-Qiang Yu. 2022. "Phylogenetic Analyses of Hydnobolites and New Species from China" Journal of Fungi 8, no. 12: 1302. https://doi.org/10.3390/jof8121302