
Heterologous Expression of CFL1 Confers Flocculating Ability to Cutaneotrichosporon oleaginosus Lipid-Rich Cells

{kind=link}
{kind=link}
{kind=link}
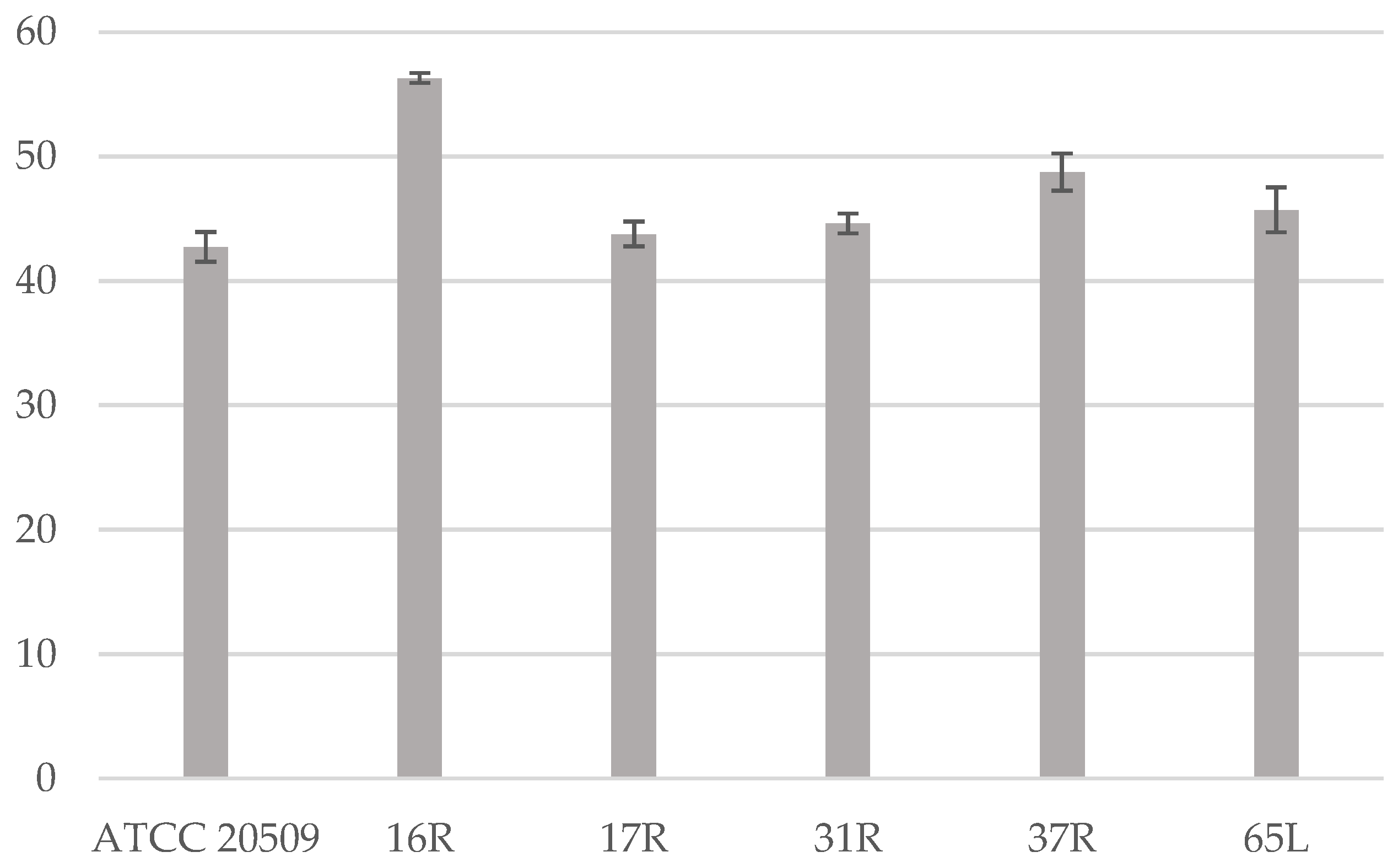{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
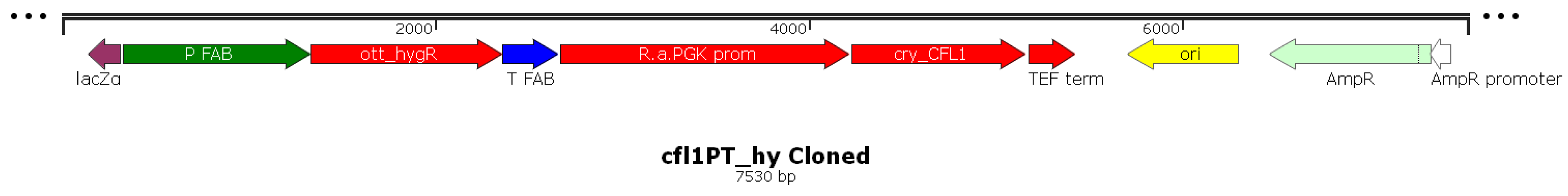
2.2. Assembly of cfl1PT_hy Vector
2.3. Transformation Protocol
2.4. DNA Extraction
2.5. RNA Extraction
2.6. Sugar and Nitrogen Determination
2.7. Dry Weight Determination and Lipid Quantification
2.8. Microscopy Imaging
2.9. Flocculation Test
2.10. Centrifugation Tests
3. Results and Discussion
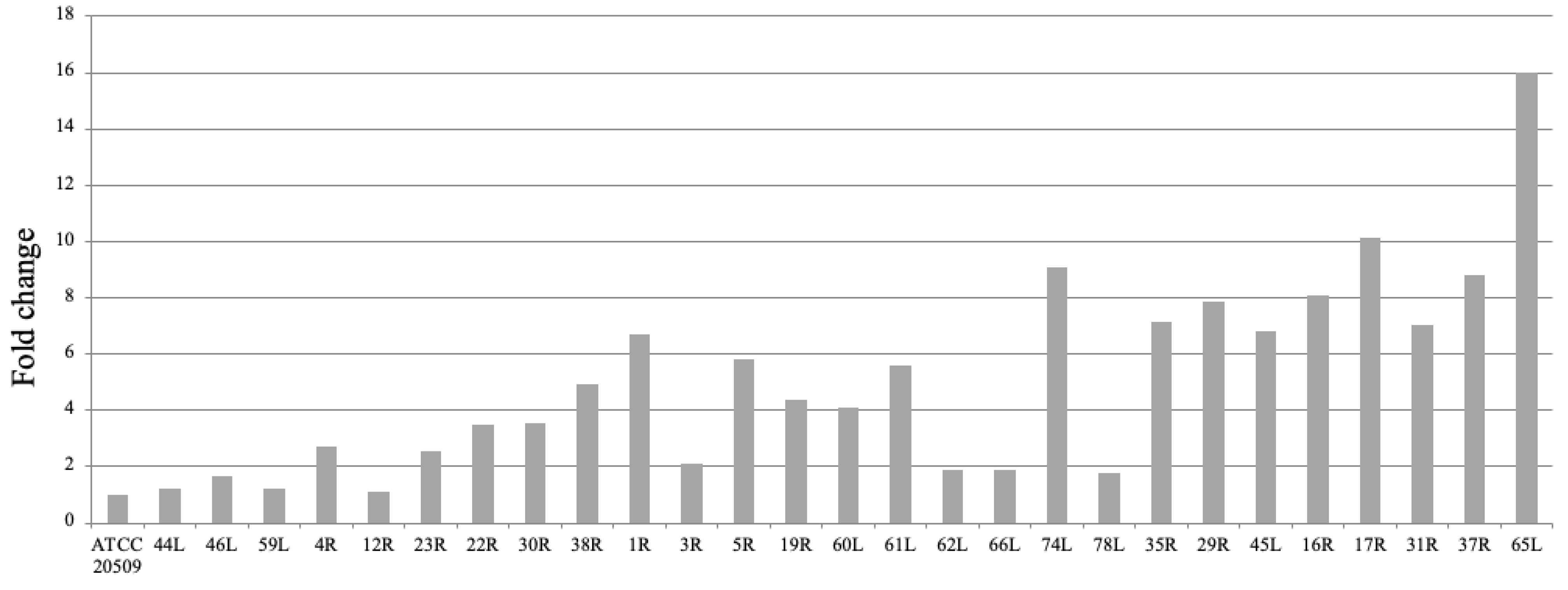
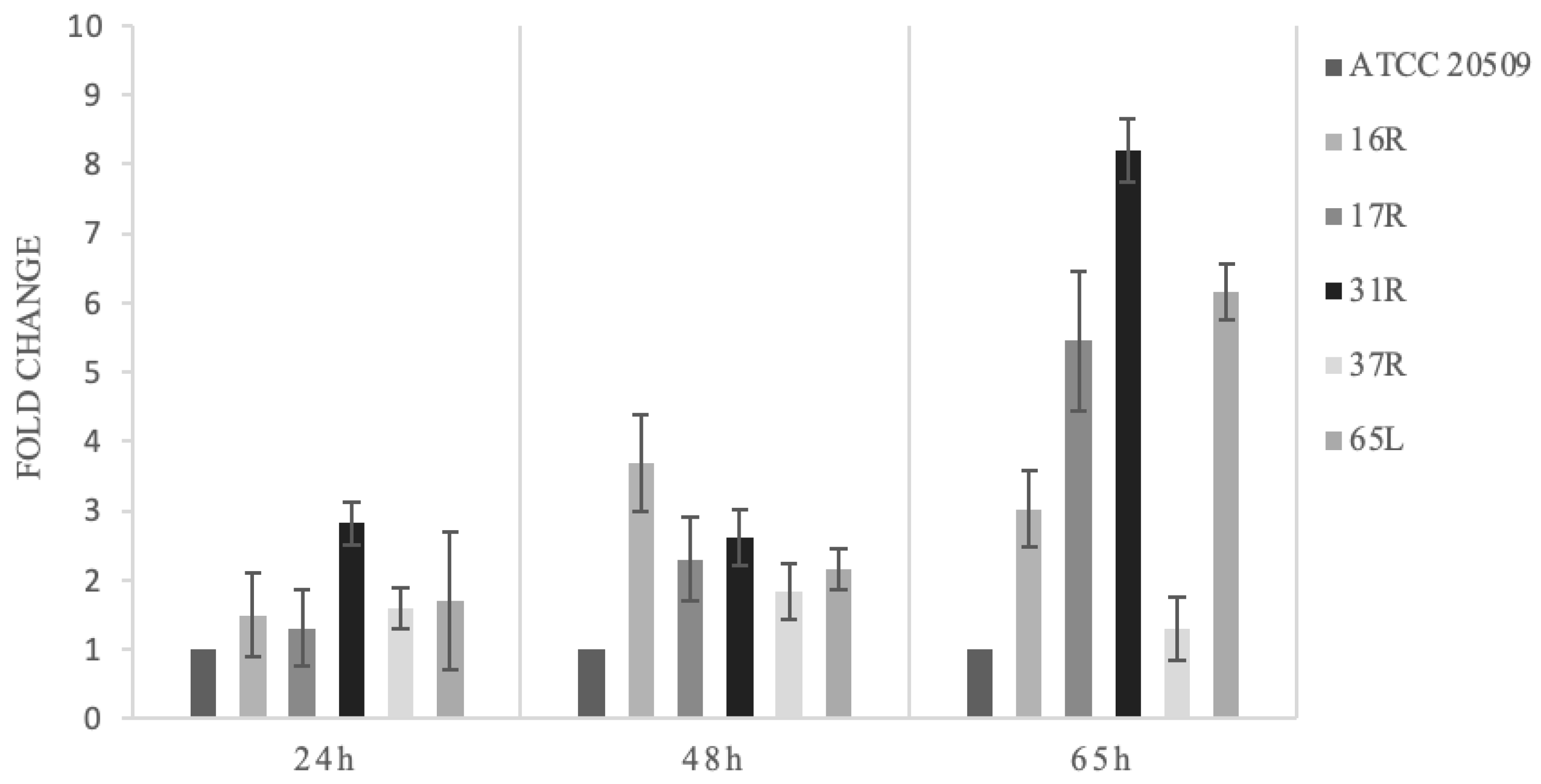
3.1. Engineering C. oleaginosus by Heterologous Expression of CFL1
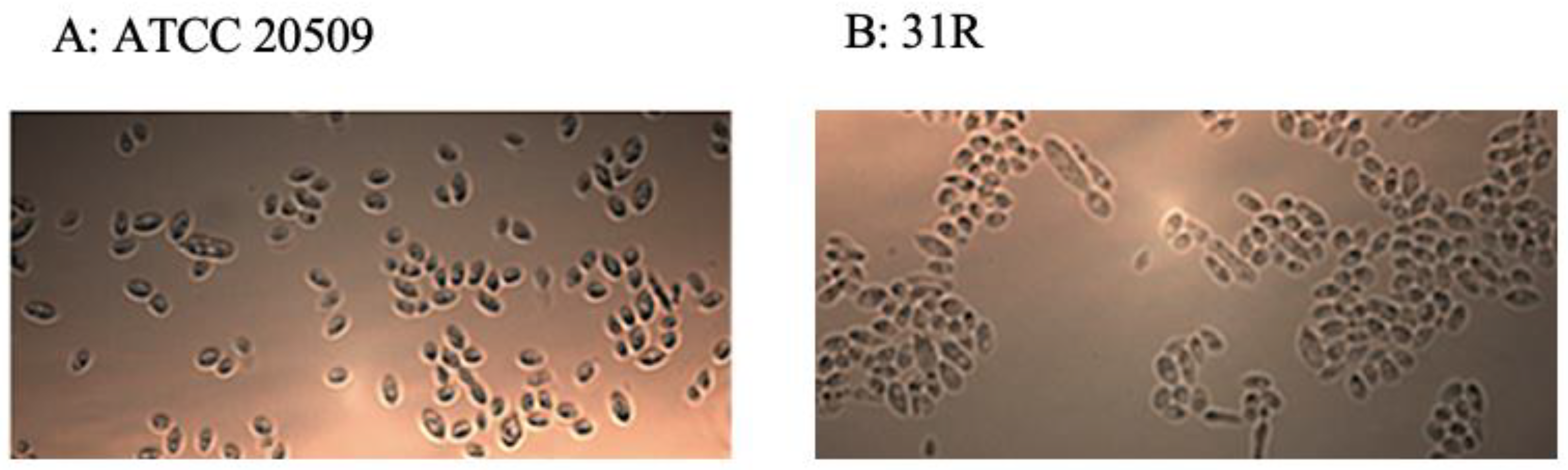
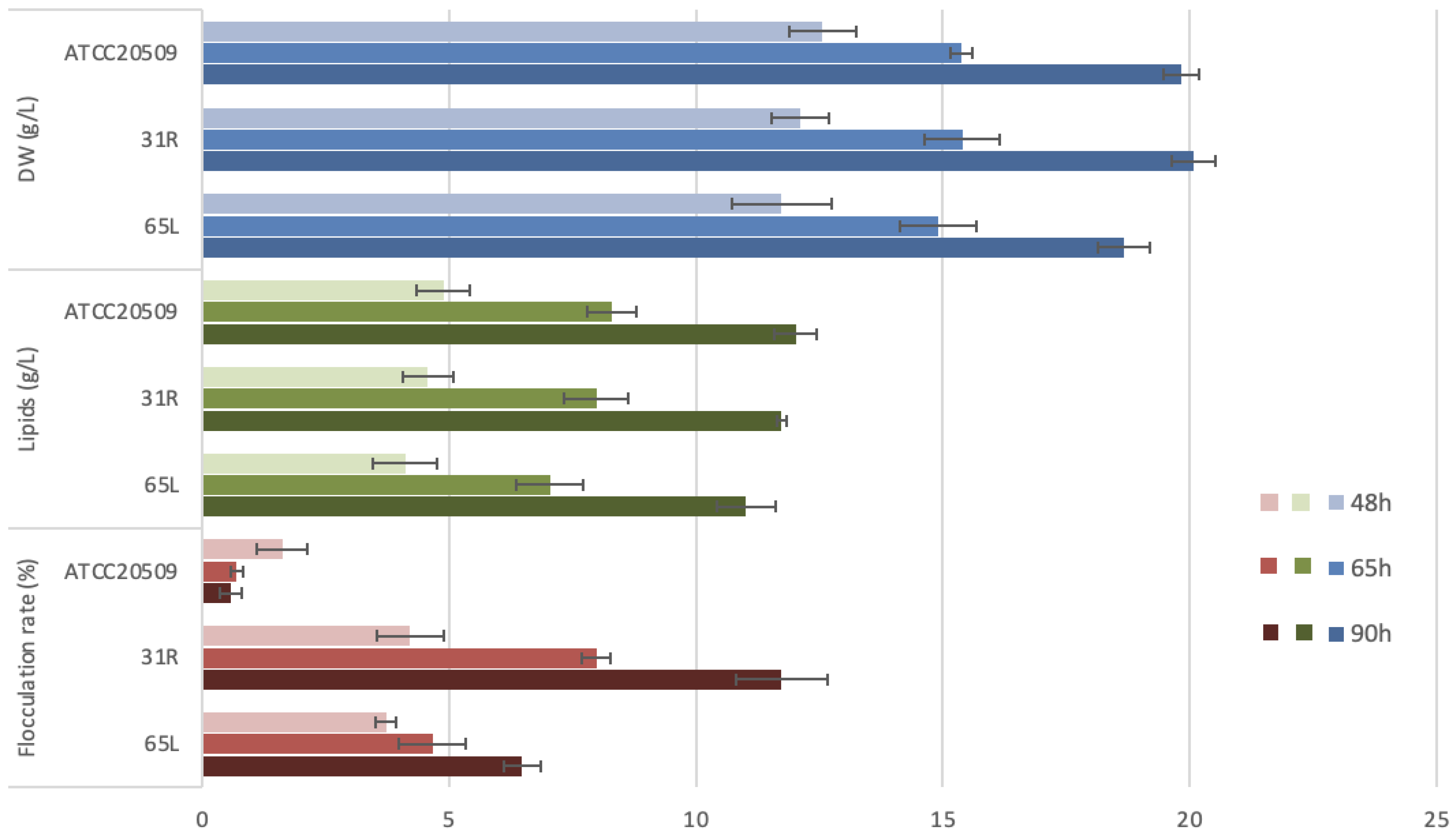
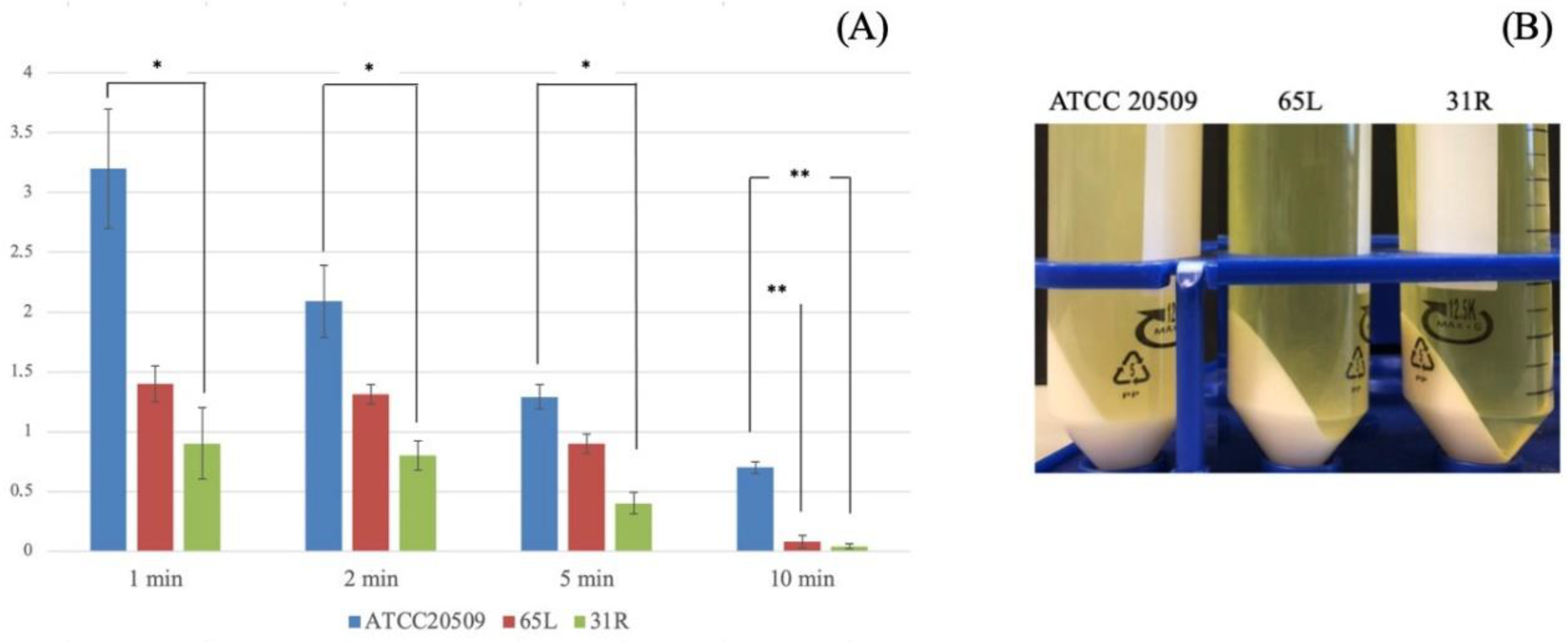
3.2. Flocculation of CFL1-Recombinant Strains Containing High Amount of Lipid
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Atabani, A.E.; Silitonga, A.S.; Badruddin, I.A.; Mahlia TM, I.; Masjuki, H.; Mekhilef, S. A comprehensive review on biodiesel as an alternative energy resource and its characteristics. Renew. Sust. Energ. Rev. 2012, 16, 2070–2093. [Google Scholar] [CrossRef]
- Abeln, F.; Chuck, C.J. The history, state of the art and future prospects for oleaginous yeast research. Microb. Cell Fact. 2021, 20, 221. [Google Scholar] [CrossRef] [PubMed]
- Ratledge, C. Yeasts, molds, algae and bacteria as sources of lipids. In Technological Advances in Improved and Alternative Sources of Lipids; Kamel, B.S., Kakuda, Y., Eds.; Springer: Boston, MA, USA, 1994; pp. 235–291. [Google Scholar] [CrossRef]
- Caporusso, A.; Capece, A.; De Bari, I. Oleaginous Yeasts as Cell Factories for the Sustainable Production of Microbial Lipids by the Valorization of Agri-Food Wastes. Fermentation 2021, 7, 50. [Google Scholar] [CrossRef]
- Christophe, G.; Kumar, V.; Nouaille, R.; Gaudet, G.; Fontanille, P.; Pandey, A.; Soccol, C.R.; Larroche, C. Recent developments in microbial oils production: A possible alternative to vegetable oils for biodiesel without competition with human food? Braz. Arch. Biol. Technol. 2012, 55, 29–46. [Google Scholar] [CrossRef] [Green Version]
- Béligon, V.; Christophe, G.; Fontanille, P.; Larroche, C. Microbial lipids as potential source to food supplements. Curr. Opin. Food Sci. 2016, 7, 35–42. [Google Scholar] [CrossRef]
- Li, Q.; Du, W.; Liu, D. Perspectives of microbial oils for biodiesel production. Appl. Microbiol. Biotechnol. 2008, 80, 749–756. [Google Scholar] [CrossRef]
- Liu, Z.; Moradi, H.; Shi, S.; Darvishi, F. Yeasts as microbial cell factories for sustainable production of biofuels. Renew. Sustain. Energy Rev. 2021, 143, 110907. [Google Scholar] [CrossRef]
- Zhang, Y.; Nielsen, J.; Liu, Z. Yeast based biorefineries for oleochemical production. Curr. Opin. Biotechnol. 2021, 6, 26–34. [Google Scholar] [CrossRef]
- Amoah, J.; Kahar, P.; Ogino, C.; Kondo, A. Bioenergy and Biorefinery: Feedstock, Biotechnological Conversion, and Products. Biotechnol. J. 2019, 14, 1800494. [Google Scholar] [CrossRef]
- Donzella, S.; Serra, I.; Fumagalli, A.; Pellegrino, L.; Mosconi, G.; Lo Scalzo, R.; Compagno, C. Recycling industrial food wastes for lipid production by oleaginous yeasts Rhodosporidiobolus azoricus and Cutaneotrichosporon oleaginosus. Biotechnol. Biofuels Bioprod. 2022, 15, 51. [Google Scholar] [CrossRef]
- Masri, M.A.; Garbe, D.; Mehlmer, N.; Brück, T.B. A sustainable, high-performance process for the economic production of waste-free microbial oils that can replace plant-based equivalents. Energy Environ. Sci. 2019, 12, 2717–2732. [Google Scholar] [CrossRef]
- Koutinas, A.A.; Chatzifragkou, A.; Kopsahelis, N.; Papanikolaou, S.; Kookos, I.K. Design and techno-economic evaluation of microbial oil production as a renewable resource for biodiesel and oleochemical production. Fuel 2014, 116, 566–577. [Google Scholar] [CrossRef]
- Dong, T.; Knoshaug, E.P.; Pienkos, P.T.; Laurens, L.M.L. Lipid recovery from wet oleaginous microbial biomass for biofuel production: A critical review. Appl. Energy 2016, 177, 879–895. [Google Scholar] [CrossRef] [Green Version]
- Jin, G.; Yang, F.; Hu, C.; Shen, H.; Zhao, Z.K. Enzyme-assisted extraction of lipids directly from the culture of the oleaginous yeast Rhodosporidium toruloides. Bioresour. Technol. 2012, 111, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Soccol, C.R.; Dalmas Neto, C.J.; Soccol, V.T.; Sydney, E.B.; da Costa, E.S.F.; Medeiros, A.B.P.; de SouzaVandenberghe, L.P. Pilot scale biodiesel production from microbial oil of Rhodosporidium toruloides DEBB5533 using sugarcane juice: Performance in diesel engine and preliminary economic study. Bioresour. Technol. 2017, 223, 259–268. [Google Scholar] [CrossRef]
- Lee, S.Y.; Cho, J.M.; Chang, Y.K.; Oh, Y.K. Cell disruption and lipid extraction for microalgal biorefineries: A review. Bioresour. Technol. 2017, 244, 1317–1328. [Google Scholar] [CrossRef]
- Harun, R.; Singh, M.; Forde, G.M.; Danquah, M.K. Bioprocess engineering of microalgae to produce a variety of consumer products. Renew. Sustain. Energy Rev. 2010, 14, 1037–1047. [Google Scholar] [CrossRef]
- Dassey, A.J.; Theegala, C.S. Harvesting economics and strategies using centrifugation for cost effective separation of microalgae cells for biodiesel applications. Bioresour. Technol. 2013, 128, 241–245. [Google Scholar] [CrossRef]
- Cerff, M.; Morweiser, M.; Dillschneider, R.; Michel, A.; Menzel, K.; Posten, C. Harvesting fresh water and marine algae by magnetic separation: Screening of separation parameters and high gradient magnetic filtration. Bioresour. Technol. 2012, 118, 289–295. [Google Scholar] [CrossRef]
- Xu, Y.; Purton, S.; Baganz, F. Chitosan flocculation to aid the harvesting of the microalga Chlorella sorokiniana. Bioresour. Technol. 2013, 129, 296–301. [Google Scholar] [CrossRef]
- Pérez, L.; Salgueiro, J.L.; Maceiras, R.; Cancela, Á.; Sánchez, Á. Study of influence of pH and salinity on combined flocculation of Chaetoceros gracilis microalgae. Chem. Eng. J. 2016, 286, 106–113. [Google Scholar] [CrossRef]
- Qiao, N.; Gao, M.; Zhang, X.; Du, Y.; Fan, X.; Wang, L.; Liu, N.; Yu, D. Trichosporon fermentans biomass flocculation from soybean oil refinery wastewater using bioflocculant produced from Paecilomyces sp. M2-1. Appl. Microbiol. Biotechnol. 2019, 103, 2821–2831. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Leong, Y.K.; Liow, J.L.; Wickramasinghe, S.R. Flocculation of yeast suspensions by a cationic flocculant. Powder Technol. 2013, 235, 426–430. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Klis, F.M. Flocculation, adhesion and biofilm formation in yeasts. Mol. Microbiol. 2006, 60, 5–15. [Google Scholar] [CrossRef]
- Guo, B.; Styles, C.A.; Feng, Q.; Fink, G.R. A Saccharomyces gene family involved in invasive growth, cell-cell adhesion, and mating. Proc. Natl. Acad. Sci. USA 2000, 97, 12158–12163. [Google Scholar] [CrossRef] [Green Version]
- Fichtner, L.; Schulze, F.; Braus, G.H. Differential Flo8p-dependent regulation of FLO1 and FLO11 for cell–cell and cell–substrate adherence of S. cerevisiae S288c. Mol. Microbiol. 2007, 66, 1276–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smukalla, S.; Caldara, M.; Pochet, N.; Beauvais, A.; Guadagnini, S.; Yan, C.; Vinces, M.D.; Jansen, A.; Prevost, M.C.; Latgé, J.P.; et al. FLO1 is a variable green beard gene that drives biofilm-like cooperation in budding yeast. Cell 2008, 135, 726–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goossens, K.V.Y.; Ielasi, F.S.; Nookaew, I.; Stals, I.; Alonso-Sarduy, L.; Daenen, L.; Van Mulders, S.E.; Stassen, C.; van Eijsden, R.G.E.; Siewers, V.; et al. Molecular mechanism of flocculation self-recognition in yeast and its role in mating and survival. mBio 2015, 6, e00427-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willaert, R.G. Adhesins of Yeasts: Protein Structure and Interactions. J. Fungi 2018, 4, 119. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Jansen, A.; Lewitter, F.; Fink, G.R. Intragenic tandem repeats generate functional variability. Nat. Genet. 2005, 37, 986–990. [Google Scholar] [CrossRef] [Green Version]
- Dranginis, A.M.; Rauceo, J.M.; Coronado, J.E.; Lipke, P.N. A biochemical guide to yeast adhesins: Glycoproteins for social and antisocial occasions. Microbiol. Mol. Biol. Rev. 2007, 71, 282–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, O.; Hayashi, N.; Kuroki, R.; Sone, H. Region of Flo1 proteins responsible for sugar recognition. J. Bacteriol. 1998, 180, 6503–6510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bony, M.; Thines-Sempoux, D.; Barre, P.; Blondin, B. Localization and cell surface anchoring of the Saccharomyces cerevisiae flocculation protein Flo1p. J. Bacteriol. 1997, 179, 4929–4936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domingues, L.; Vicente, A.A.; Lima, N.; Teixeira, J.A. Applications of yeast flocculation in biotechnological processes. Biotechnol. Bioprocess Eng. 2000, 5, 288–305. [Google Scholar] [CrossRef] [Green Version]
- Verstrepen, K.J.; Derdelinckx, G.; Verachtert, H.; Delvaux, F.R. Yeast flocculation: What brewers should know. Appl. Microbiol. Biotechnol. 2003, 61, 197–205. [Google Scholar] [CrossRef]
- Sivakumar, G.; Vail, D.R.; Xu, J.; Burner, D.M.; Lay, J.O.; Ge, X.; Weathers, P.J. Bioethanol and biodiesel: Alternative liquid fuels for future generations. Eng. Life Sci. 2010, 10, 8–18. [Google Scholar] [CrossRef]
- Westman, J.O.; Mapelli, V.; Taherzadeh, M.J.; Franzén, C.J. Flocculation causes inhibitor tolerance in Saccharomyces cerevisiae for second-generation bioethanol production. Appl. Environ. Microbiol. 2014, 80, 6908–6918. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Lin, X. Matricellular protein Cfl1 regulates cell differentiation. Commun. Integr. Biol. 2013, 6, e26444. [Google Scholar] [CrossRef]
- Wang, L.; Tian, X.; Gyawali, R.; Lin, X. Fungal adhesion protein guides community behaviors and autoinduction in a paracrine manner. Proc. Natl. Acad. Sci. USA 2013, 110, 11571–11576. [Google Scholar] [CrossRef]
- Gyawali, R.; Upadhyay, S.; Way, J.; Lin, X. A family of secretory proteins is associated with different morphotypes in Cryptococcus neoformans. Appl. Environ. Microbiol. 2017, 83, e02967-16. [Google Scholar] [CrossRef] [Green Version]
- Hoyer, L.L. The ALS gene family of Candida albicans. Trends Microbiol. 2001, 9, 176–180. [Google Scholar] [CrossRef]
- Wang, L.; Zhai, B.; Lin, X. The link between morphotype transition and virulence in Cryptococcus neoformans. PLoS Pathog. 2012, 8, e1002765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, N.; Reijnders, M.; Suarez-Diez, M.; Nijsse, B.; Springer, J.; Eggink, G.; Schaap, P.J. Genome-scale metabolic modeling underscores the potential of Cutaneotrichosporon oleaginosus ATCC 20509 as a cell factory for biofuel production. Biotechnol. Biofuels 2021, 14, 2. [Google Scholar] [CrossRef] [PubMed]
- Bracharz, F.; Beukhout, T.; Mehlmer, N.; Bru¨ck, T. Opportunities and challenges in the development of Cutaneotrichosporon oleaginosus ATCC 20509 as a new cell factory for custom tailored microbial oils. Microb. Cell Fact. 2017, 16, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Fidio, N.; Minonne, F.; Antonetti, C.; Raspolli Galletti, A.M. Cutaneotrichosporon oleaginosus: A Versatile Whole-Cell Biocatalyst for the Production of Single-Cell Oil from Agro-Industrial Wastes. Catalysts 2021, 11, 1291. [Google Scholar] [CrossRef]
- Donzella, S.; Cucchetti, D.; Capusoni, C.; Rizzi, A.; Galafassi, S.; Chiara, G.; Compagno, C. Engineering cytoplasmic acetyl-CoA synthesis decouples lipid production from nitrogen starvation in the oleaginous yeast Rhodosporidium azoricum. Microb. Cell Fact. 2019, 18, 199. [Google Scholar] [CrossRef]
- Querol, A.; Barrio, E.; Huerta, T.; Ramón, R. Molecular monitoring of wine fermentations conducted by active dry yeast strains. Appl. Environ. Microbiol. 1992, 58, 2948–2953. [Google Scholar] [CrossRef] [Green Version]
- Helm, E.; Nohr, B.; Thorne, R.S.W. The measurement of yeast flocculation and its significance in brewing. Wall Lab. Commun. 1953, 16, 315–326. [Google Scholar]
- Dahiya, P.; Bhat, D.; Thakur, J. Expression of AtMed15 of Arabidopsis in yeast causes flocculation and increases ethanol production in yeast culture. Sci. Rep. 2016, 6, 27967. [Google Scholar] [CrossRef] [Green Version]
- Görner, C.; Redai, V.; Bracharz, F.; Schrepfer, P.; Garbe, D.; Brück, T. Genetic engineering and production of modified fatty acids by the non-conventional oleaginous yeast Trichosporon oleaginosus ATCC 20509. Green Chem. 2016, 18, 2037–2046. [Google Scholar] [CrossRef]
- Koivuranta, K.; Castillo, S.; Jouhten, P.; Ruohonen, L.; Penttila, M.; Wiebe, M.G. Enhanced triacylglycerol production with genetically modified Trichosporon oleaginosus. Front. Microbiol. 2018, 9, 1337. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Wang, Y.; Zhang, S.; Zhu, Z.W.; Zhou, Y.J.J.; Yang, F.; Sun, W.Y.; Wang, X.Y.; Zhao, Z.B.K. Functional integration of multiple genes into the genome of the oleaginous yeast Rhodosporidium toruloides. FEMS Yeast Res. 2014, 14, 547–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonturi, N.; Matsakas, L.; Nilsson, R.; Christakopoulos, P.; Miranda, E.A.; Berglund, K.A.; Rova, U. Single cell oil producing yeasts Lipomyces starkeyi and Rhodosporidium toruloides: Selection of extraction strategies and biodiesel property prediction. Energies 2015, 8, 5040–5052. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.; Mikes, F.; Matsakas, L. An Overview of Current Pretreatment Methods Used to Improve Lipid Extraction from Oleaginous Micro-Organisms. Molecules 2018, 23, 1562. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, B.; Teixeira, J.C.; Dragone, G.; Teixeira, J.A. Optimization of lipid extraction from the oleaginous yeasts Rhodotorula glutinis and Lipomyces kononenkoae. AMB Exp. 2018, 8, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalmas Neto, C.J.; Sydney, E.B.; Candeo, E.S.; de Souza, E.B.S.; Camargo, D.; Sydney, A.C.N.; de Carvalho, C.J.; Letti, L.A.J.; Pandey, A.; Soccol, C.R. New Method for the Extraction of Single-Cell Oils from Wet Oleaginous Microbial Biomass: Efficiency, Oil Characterization and Energy Assessment. Waste Biomass Valorization 2020, 11, 3443–3452. [Google Scholar] [CrossRef]
- Govender, P.; Domingo, J.L.; Bester, M.C.; Pretorius, I.S.; Bauer, F.F. Controlled expression of the dominant flocculation genes FLO1, FLO5 and FLO11 in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2008, 74, 6041–6052. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.-Q.; Li, Q.; He, L.-Y.; Li, F.; Que, W.-W.; Bai, F.-W. Exploration of a natural reservoir of flocculating genes from various Saccharomyces cerevisiae strains and improved ethanol fermentation using stable genetically engineered flocculating yeast strains. Process Biochem. 2011, 47, 1612–1619. [Google Scholar] [CrossRef]
- Varela, C.; Bartel, C.; Nandorfy, D.E.; Borneman, A.; Schmidt, S.; Curtin, C. Identification of flocculant wine yeast strains with improved filtration-related phenotypes through application of high-throughput sedimentation rate assays. Sci. Rep. 2020, 10, 2738. [Google Scholar] [CrossRef] [Green Version]
- Westman, J.O.; Nyman, J.; Manara, R.M.A.; Mapelli, V.; Franzén, C.J. A novel chimaeric flocculation protein enhances flocculation in Saccharomyces cerevisiae. Metab. Eng. Commun. 2018, 6, 49–55. [Google Scholar] [CrossRef]
- Yang, L.; Zheng, C.; Chen, Y.; Ying, H. FLO genes family and transcription factor MIG1 regulate Saccharomyces cerevisiae biofilm formation during immobilized fermentation. Front. Microbiol. 2018, 9, 1860. [Google Scholar] [CrossRef] [PubMed]
- Moreno-García, J.; Martín-García, F.J.; Ogawa, M.; García-Martínez, T.; Moreno, J.; Mauricio, J.C.; Bisson, L.F. FLO1, FLO5 and FLO11 flocculation gene expression impacts Saccharomyces cerevisiae attachment to Penicillium chrysogenum in a co-immobilization technique. Front. Microbiol. 2018, 9, 2586. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.-L.; Speers, R.A. Effect of environmental conditions on the flocculation of Saccharomyces cerevisiae. J. Am. Soc. Brew. Chem. 2000, 58, 108–116. [Google Scholar] [CrossRef]
- Wilcocks, K.L.; Smart, K.A. The importance of surface charge and hydrophobicity for the flocculation of chain-forming. brewing yeast strains and resistance of these parameters to acid washing. FEMS Microbiol. Lett. 1995, 134, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Van Hamersveld, E.H.; van der Lans, R.G.J.M.; Luyben, K.C.A.M. Quantification of brewer’s yeast flocculation in a stirred tank: Effect of physical parameters on flocculation. Biotechnol. Bioeng. 1997, 56, 190–200. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donzella, S.; Compagno, C. Heterologous Expression of CFL1 Confers Flocculating Ability to Cutaneotrichosporon oleaginosus Lipid-Rich Cells. J. Fungi 2022, 8, 1293. https://doi.org/10.3390/jof8121293
Donzella S, Compagno C. Heterologous Expression of CFL1 Confers Flocculating Ability to Cutaneotrichosporon oleaginosus Lipid-Rich Cells. Journal of Fungi. 2022; 8(12):1293. https://doi.org/10.3390/jof8121293
Chicago/Turabian StyleDonzella, Silvia, and Concetta Compagno. 2022. "Heterologous Expression of CFL1 Confers Flocculating Ability to Cutaneotrichosporon oleaginosus Lipid-Rich Cells" Journal of Fungi 8, no. 12: 1293. https://doi.org/10.3390/jof8121293