
Evaluation of Resistance of Banana Genotypes with AAB Genome to Fusarium Wilt Tropical Race 4 in China
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Evaluation under Greenhouse Conditions
2.3. Evaluation under Field Conditions
2.4. Molecular Characterization of Foc TR4
2.5. Data Collection and Statistical Analysis
3. Results
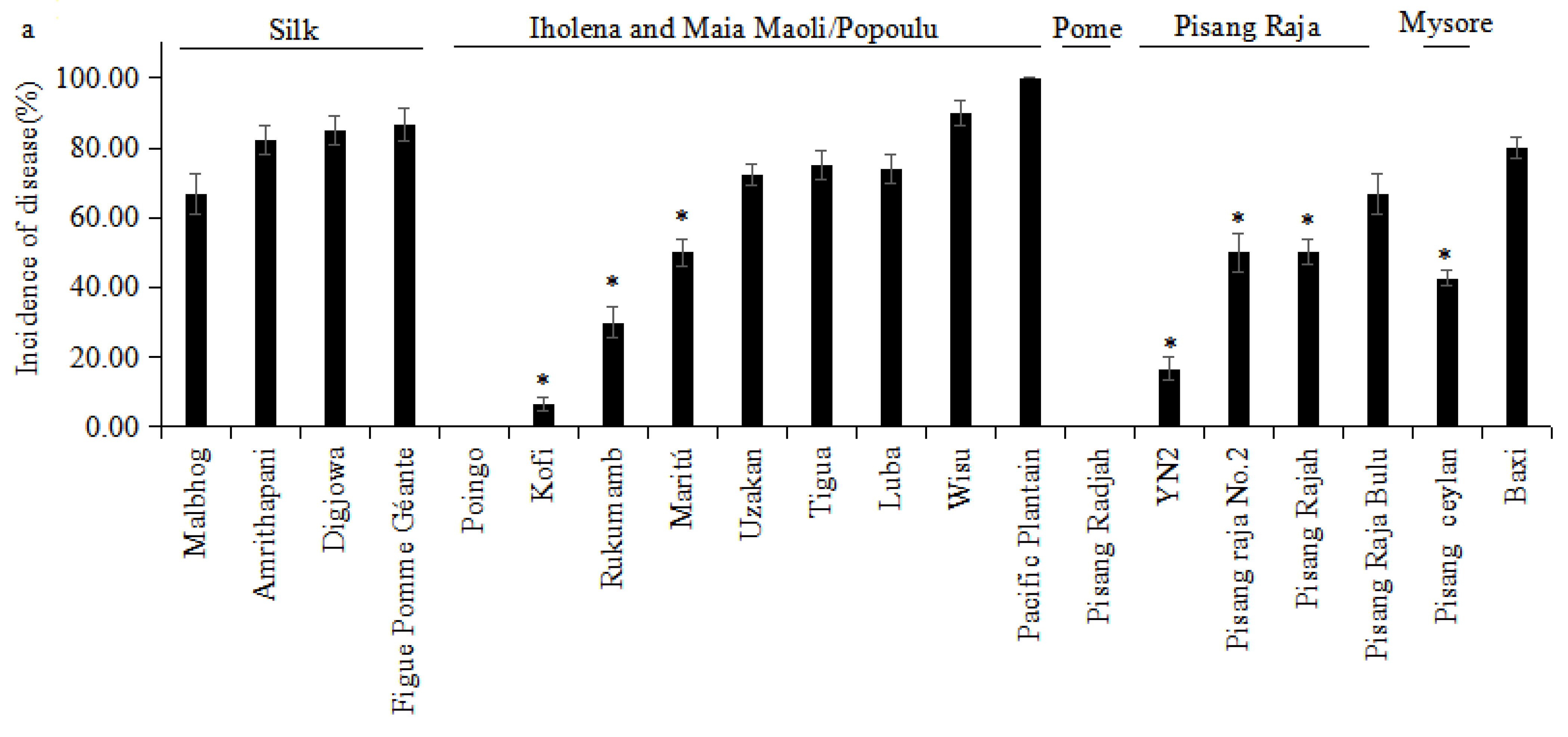
3.1. Evaluation under Greenhouse Conditions
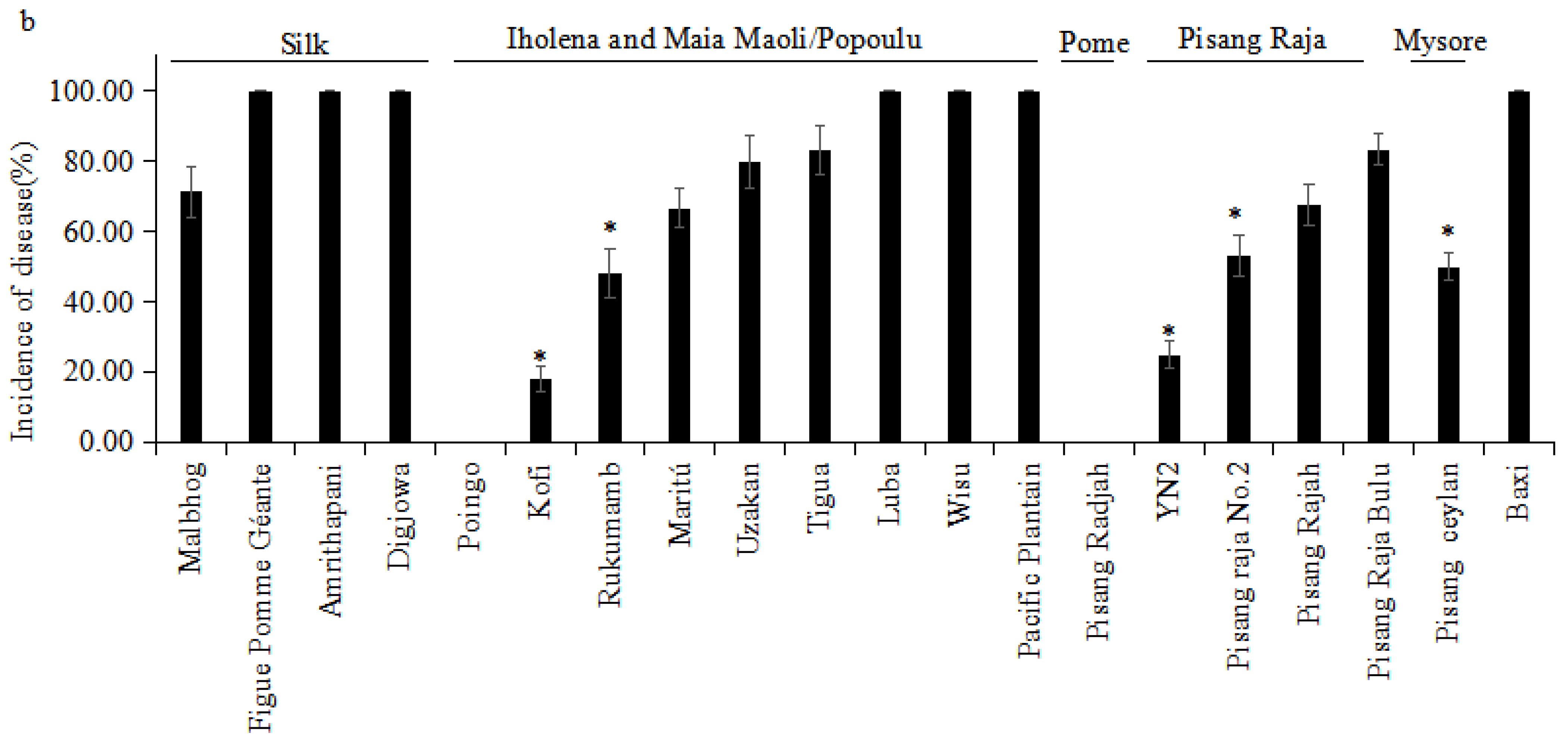
3.2. Evaluation under Field Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simmonds, N.W.; Shepherd, K. The taxonomy and origins of the cultivated bananas. Bot. J. Linn. Soc. 1995, 55, 302–312. [Google Scholar] [CrossRef]
- Robinson, J.C.; Sauco, V.G. Bananas and Plantains, 2nd ed.; CABI Publishing: Wallingford, UK, 2010. [Google Scholar]
- Bancroft, J. Report of the board appointed to inquire into to the cause of disease affecting livestock and plants. Votes Proc. 1876, 3, 1011–1038. [Google Scholar]
- Siamak, S.B.; Zheng, S. Banana Fusarium Wilt (Fusarium oxysporum f. sp. cubense) Control and Resistance, in the Context of Developing Wilt-resistant Bananas Within Sustainable Production Systems. Hortic. Plant J. 2018, 4, 208–218. [Google Scholar] [CrossRef]
- Stover, R.H. Fusarial wilt (Panama disease) of bananas and other Musa species. Commonwealth Mycological Institute. Phytopathology 1962, 4, 117. [Google Scholar]
- Tinzaara, W.; Mutambuka, M.; Oyesigye, E.; Blomme, G.; Dita, M.; Gold, C.S.; Rouard, M.; Tinzaara, E. Banana wilt diseases: Current status and future research strategies for their management. Int. J. Pest Manag. 2021, 1, 1–20. [Google Scholar] [CrossRef]
- Dita, M.; Barquero, M.; Heck, D.; Mizubuti, E.S.G.; Staver, C.P. Fusarium Wilt of Banana: Current Knowledge on Epidemiology and Research Needs Toward Sustainable Disease Management. Front. Plant Sci. 2018, 9, 1468. [Google Scholar] [CrossRef] [Green Version]
- Wardlaw, C.W. Banana Diseases Including Plantains and Abaca; Longmans, Green & Co., Ltd.: London, UK, 1961. [Google Scholar]
- Ploetz, R.C. Fusarium Wilt of Banana. Phytopathology 2015, 105, 1512–1521. [Google Scholar] [CrossRef] [Green Version]
- Fraser-Smith, S.; Czislowski, E.; Daly, A.; Meldrum, R.; Hamill, S.; Smith, M.; Aitken, E.A.B. Single gene resistance to Fusarium oxysporum f. sp. cubense Race 4 in the wild banana Musa acuminata subsp. Malaccensis. In Proceedings of the XXIX International Horticultural Congress on Horticulture, Brisbane, Australia, 17–22 August 2014. [Google Scholar]
- Ploetz, R.C. Fusarium wilt. In Handbook of Diseases of Banana, Abacá and Enset; Jones, D., Ed.; CAB International: Wallingford, UK, 2019; pp. 207–229. [Google Scholar]
- Ploetz, R.C. Fusarium Wilt of Banana Is Caused by Several Pathogens Referred to as Fusarium oxysporum f. sp. cubense. Phytopathology 2006, 96, 653–656. [Google Scholar] [CrossRef] [Green Version]
- Czislowski, E.; Fraser-Smith, S.; Zander, M.; O’Neill, W.T.; Meldrum, R.A.; Tran-Nguyen, L.T.; Batley, J.; Aitken, E.A. Investigation of the diversity of effector genes in the banana pathogen, Fusarium oxysporum f. sp. cubense, reveals evidence of horizontal gene transfer. Mol. Plant Pathol. 2018, 19, 1155–1171. [Google Scholar] [CrossRef] [Green Version]
- Carvalhais, L.C.; Henderson, J.; Rincon-Florez, V.A.; O’Dwyer, C.; Czislowski, E.; Aitken, E.A.B.; Drenth, A. Molecular Diagnostics of Banana Fusarium Wilt Targeting Secreted-in-Xylem Genes. Front. Plant Sci. 2019, 10, 547. [Google Scholar] [CrossRef]
- Molina, A.B.; Fabregar, E.; Sinohin, V.; Yi, G.; Viljoen, A. Recent occurrence of Fusarium oxysporum f. sp. cubense tropical race 4 in Asia. In Proceedings of the III International Symposium on Banana, ISHS-ProMusa Symposium on Recent Advances in Banana Crop Protection for Sustainable 828, White River, South Africa, 10–14 September 2007. [Google Scholar]
- Thangavelu, R.; Edwin, R.E.; Pushpakanth, P.; Loganathan, M.; Uma, S. Draft Genome of Fusarium oxysporum f. sp. cubense Strain Tropical Race-4 Infecting Cavendish (AAA) Group of Banana in India. Plant Dis. 2020, 105, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Scheerer, L.; Pemsl, D.; Dita, M.; Pérez Vicente, L.; Staver, C. A quantified approach to project losses caused by Fusarium wilt tropical race 4. Proc. Int. Symp. Banana, 10th Acta Hortic. 2018, 1196, 2112–2218. [Google Scholar]
- Zuo, C.; Deng, G.; Li, B.; Huo, H.; Li, C.; Hu, C.; Kuang, R.; Yang, Q.; Dong, T.; Sheng, O.; et al. Germplasm screening of Musa spp. for resistance to Fusarium oxysporum f. sp. cubense tropical race 4 (Foc TR4). Eur. J. Plant Pathol. 2018, 151, 723–734. [Google Scholar] [CrossRef]
- Molina, A.B.; Sinohin, V.O.; Fabregar, E.; Ramillete, E.B.; Yi, G.; Sheng, O.; Karamura, D.; Van den Bergh, I.; Viljoen, A. Resistance to Fusarium oxysporum f. sp. cubense tropical race 4 in African Bananas. Acta Hortic. 2016, 1114, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Mintoff, S.J.; Nguyen, T.V.; Kelly, C.; Cullen, S.; Hearnden, M.; Williams, R.; Daniells, J.W.; Tran-Nguyen, L.T. Banana Cultivar Field Screening for Resistance to Fusarium oxysporum f.sp. cubense Tropical Race 4 in the Northern Territory. J. Fungi 2021, 7, 627. [Google Scholar] [CrossRef] [PubMed]
- Li, W.M.; Dita, M.; Wu, W.; Hu, G.B.; Xie, J.H.; Ge, X.J. Resistance sources to Fusarium oxysporum f. sp. cubense tropical race 4 in banana wild relatives. Plant Pathol. 2015, 64, 1061–1067. [Google Scholar] [CrossRef]
- Chen, A.; Sun, J.; Matthews, A.; Armas-Egas, L.; Chen, N.; Hamill, S.; Mintoff, S.; Tran-Nguyen, L.T.; Batley, J.; Aitken, E.A. Assessing Variations in Host Resistance to Fusarium oxysporum f sp. cubense Race 4 in Musa Species, With a Focus on the Subtropical Race 4. Front. Microbiol. 2019, 10, 1062. [Google Scholar] [CrossRef] [Green Version]
- Dita, M.; Teixeira, L.; Li, C.; Zheng, S.; O’Neill, W.; Daniels, J. Phenotyping Musa spp. for host reaction to Fusarium oxysporum f. sp. cubense, under greenhouse and field conditions. In Practical Guidelines for Early Screening and Field Evaluation of Banana against Fusarium Wilt, Pseudocercospora Leaf Spots and Drought; Dita, M., Ed.; Bioversity International: Montpellier, France, 2021; pp. 5–18. [Google Scholar]
- Swamy, R.D.; Rao, N.K.S.; Chacko, E.K. Tissue-culture propagation of banana. Sci. Hortic. 1983, 18, 247–252. [Google Scholar] [CrossRef]
- Li, C.Y.; Mostert, G.; Zuo, C.W.; Beukes, I.; Yang, Q.S.; Sheng, O.; Kuang, R.B.; Wei, Y.R.; Hu, C.H.; Rose, L.; et al. Diversity and distribution of the banana wilt pathogen Fusarium oxysporum f. sp. cubense in China. Fungal Genom. Biol. 2013, 3, 111. [Google Scholar]
- Mostert, D.; Molina, A.B.; Daniells, J.; Fourie, G.; Hermanto, C.; Chao, C.P.; Fabregar, E.; Sinohin, V.G.; Masdek, N.; Thangavelu, R.; et al. The distribution and host range of the banana Fusarium wilt fungus, Fusarium oxysporum f. sp. cubense, in Asia. PLoS ONE 2017, 12, e0181630. [Google Scholar] [CrossRef] [Green Version]
- Dita, M.A.; Waalwijk, C.; Buddenhagen, I.W.; Souza, M.T., Jr.; Kema, G.H.J. A molecular diagnostic for tropical race 4 of the banana fusarium wilt pathogen. Plant Pathol. 2010, 59, 348–357. [Google Scholar] [CrossRef]
- Viljoen, A.; Mahuku, G.; Massawe, C.; Ssali, R.T.; Kimunye, J.; Mostert, G.; Ndayihanzamaso, P.; Coyne, D.L. Banana Pests and Diseases: Field Guide for Disease Diagnostics and Data Collection; International Institute of Tropical Agriculture: Ibadan, Nigeria, 2017. [Google Scholar]
- Shen, Z.; Zhong, S.; Wang, Y.; Wang, B.; Mei, X.; Li, R.; Ruan, Y.; Shen, Q. Induced soil microbial suppression of banana fusarium wilt disease using compost and biofertilizers to improve yield and quality. Eur. J. Soil Biol. 2013, 57, 1–8. [Google Scholar] [CrossRef]
- Zorrilla, F.Y.; Pauwels, L.; Panis, B.; Signorelli, S.; Vanderschuren, H.; Swennen, R. Strategies to revise agrosystems and breeding to control Fusarium wilt of banana. Nat. Food 2020, 1, 599–604. [Google Scholar] [CrossRef]
- Molina, A.B.; Fabregar, E.; Sinohin, V.G.; Fourie, A.G.; Viljoen, A. Tropical Race 4 of Fusarium oxysporum f. sp. cubense causing new Panama wilt epidemics in Cavendish varieties in the Philippines. Phytopathology 2008, 98, 108. [Google Scholar]
- Molina, A.B.; Fabregar, E.; Sinohin, G.; Yi, G.; Fourie, G.; Viljoen, A. Recent occurrence of Fusarium oxysporum f.sp. cubense tropical race 4 in Asia. Acta Hortic. 2009, 828, 109–116. [Google Scholar] [CrossRef]
- Bakry, F.; Horry, J.P. Tetraploid hybrids from interploid 3x/2x crosses in cooking bananas. Fruits 1993, 47, 641. [Google Scholar]
- Kepler, A.K.; Rust, F.G. The World of Bananas in Hawai’i: Then and Now; Pali-O-Waipi’o Press: Honolulu, HI, USA, 2011. [Google Scholar]
- Ploetz, R.C.; Kepler, A.K.; Daniells, J.; Nelson, S.C. Banana and plantain: An overview with emphasis on Pacific island cultivars Musaceae (banana family). In Species Profiles for Pacific Island Agroforestry; Elevitch, R., Ed.; Permanent Agriculture Resources: Holualoa, HI, USA, 2007. [Google Scholar]
- Probojati, R.T.; Wahyudi, D.; Hapsari, L. Clustering analysis and genome inference of pisang raja local cultivars (Musa Spp.) from Java Island by random amplified polymorphic DNA (RAPD) marker. J. Trop. Biotechnol. Biodivers. 2019, 4, 42–53. [Google Scholar] [CrossRef] [Green Version]
- Fontes, S.D.M.; Cavalcanti, M.T.; Candeia, R.A.; Almeida, E.L. Characterization and study of functional properties of banana starch green variety of Mysore (Musa AAB-Mysore). Food Sci. Technol. 2017, 37, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.K.; Langdon, P.W.; Pegg, K.G.; Daniells, J.W. Growth, yield and Fusarium wilt resistance of six FHIA tetraploid bananas (Musa spp.) grown in the Australian subtropics. Sci. Hortic. 2014, 170, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Thangavelu, R.; Saraswathi, M.S.; Uma, S.; Loganathan, M.; Backiyarani, S.; Durai, P.; Raj, E.E.; Marimuthu, N.; Kannan, G.; Swennen, R. Identification of sources resistant to a virulent Fusarium wilt strain (VCG 0124) infecting Cavendish bananas. Sci. Rep. 2021, 11, 3183. [Google Scholar] [CrossRef]
- Ndayihanzamaso, P.; Mostert, D.; Matthews, M.C.; Mahuku, G.; Jomanga, K.; Mpanda, H.J.; Mduma, H.; Brown, A.; Uwimana, B.; Swennen, R.; et al. Evaluation of Mchare and Matooke Bananas for Resistance to Fusarium oxysporum f. sp. cubense Race 1. Plants 2020, 9, 1082. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | ITC Code | Genome | Subgroup/Type |
|---|---|---|---|
| Obubit Ntanga green mutant | ITC0519 | AAB | Plantain/French |
| Kakira | NA | AAB | Plantain/French |
| Uganda Plantain | NA | AAB | Plantain/French |
| French Sombre | ITC1668 | AAB | Plantain/French |
| French P | NA | AAB | Plantain/French |
| Nakatansese | NA | AAB | Plantain/French |
| Ntanga 4 | ITC0226 | AAB | Plantain/French |
| Njombe N°2 | ITC1124 | AAB | Plantain/French |
| Batard | NA | AAB | Plantain/French Horn |
| Batard 2 | NA | AAB | Plantain/French Horn |
| Plantain no.3 | ITC0498 | AAB | Plantain/French Horn |
| Orishele | ITC1325 | AAB | Plantain/False Horn |
| Curare | ITC1165 | AAB | Plantain/False Horn |
| Big Ebanga | NA | AAB | Plantain/False Horn |
| CB5 | NA | AAB | Plantain/False Horn |
| CEMSA3/4 | NA | AAB | Plantain/False Horn |
| Ihitisim | ITC0121 | AAB | Plantain/Horn |
| FHIA-21 | NA | AAAB | Hybrid/Plantain |
| Figue Pomme Géante | ITC0769 | AAB | Silk |
| Amrithapani | ITC1612 | AAB | Silk |
| Malbhog | ITC1631 | AAB | Silk |
| Digjowa | ITC1633 | AAB | Silk |
| Maritú | ITC0639 | AAB | Iholena |
| Luba | ITC0802 | AAB | Iholena |
| Wisu | ITC0880 | AAB | Iholena |
| Kofi | ITC0912 | AAB | Iholena |
| Tigua | ITC0875 | AAB | Iholena |
| Uzakan | ITC0825 | AAB | Iholena |
| Rukumamb | ITC0831 | AAB | Iholena |
| Pacific Plantain | ITC0210 | AAB | Maia Maoli/Popoulu |
| Poingo | ITC1327 | AAB | Maia Maoli/Popoulu |
| Pisang Rajah | ITC0587 | AAB | Pisang Raja |
| Pisang Raja Bulu | ITC0843 | AAB | Pisang Raja |
| YN2 | NA | AAB | Pisang Raja |
| Pisang Raja No.2 | NA | AAB | Pisang Raja |
| Pisang Ceylan | ITC1441 | AAB | Mysore |
| Pisang Radjah | ITC0243 | AAB | Pome |
| Baxi | NA | AAA | Cavendish |
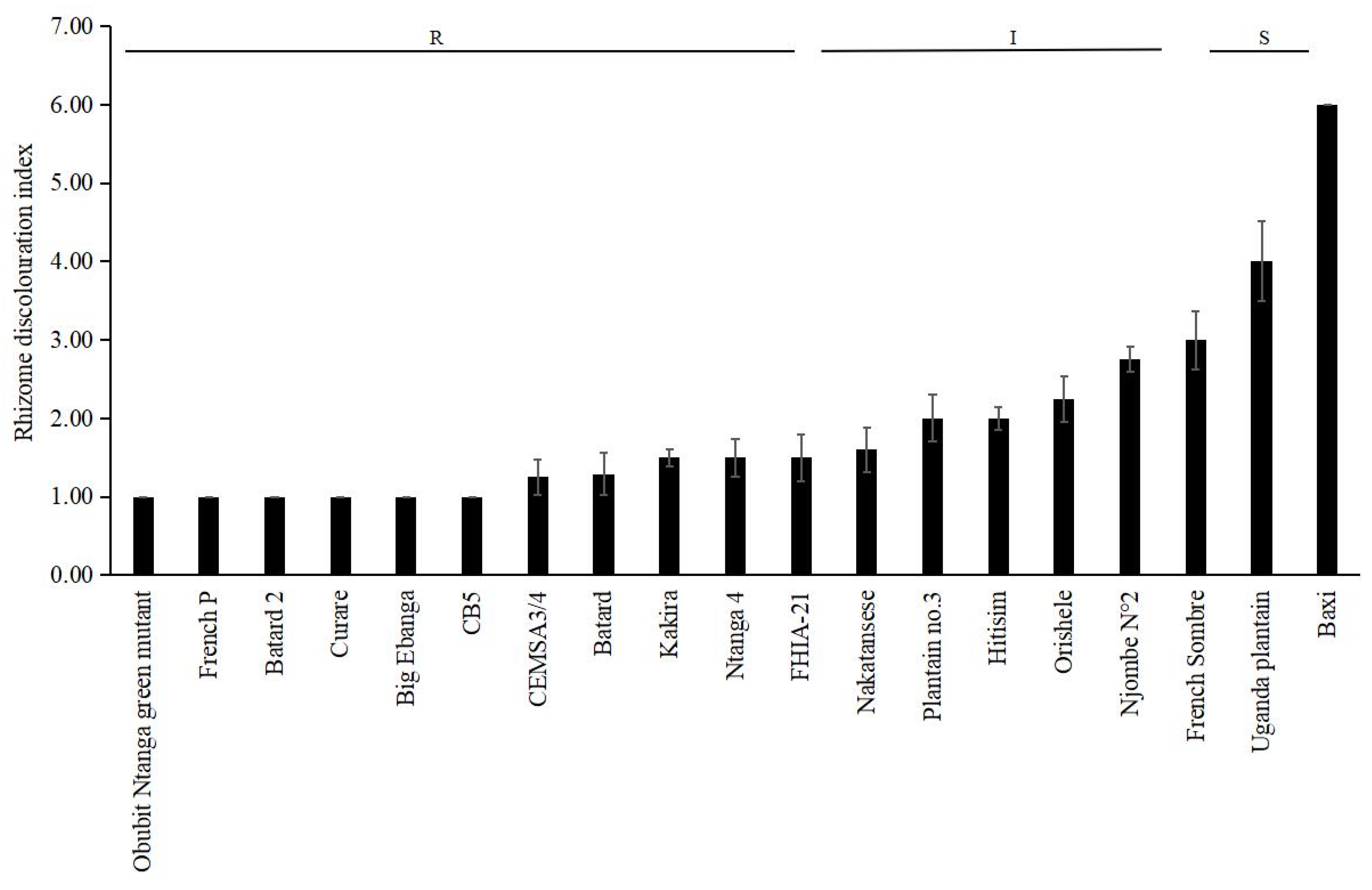
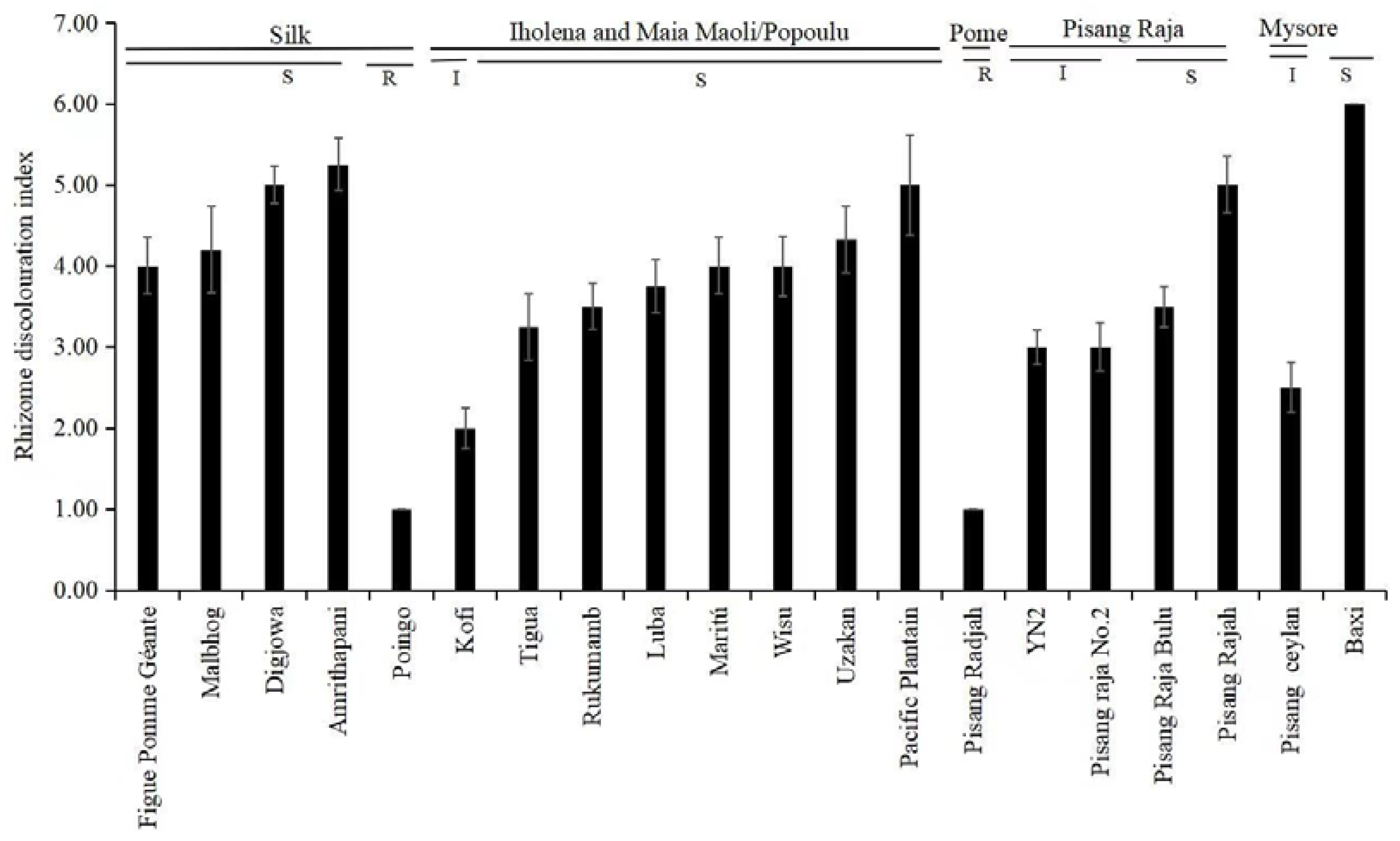
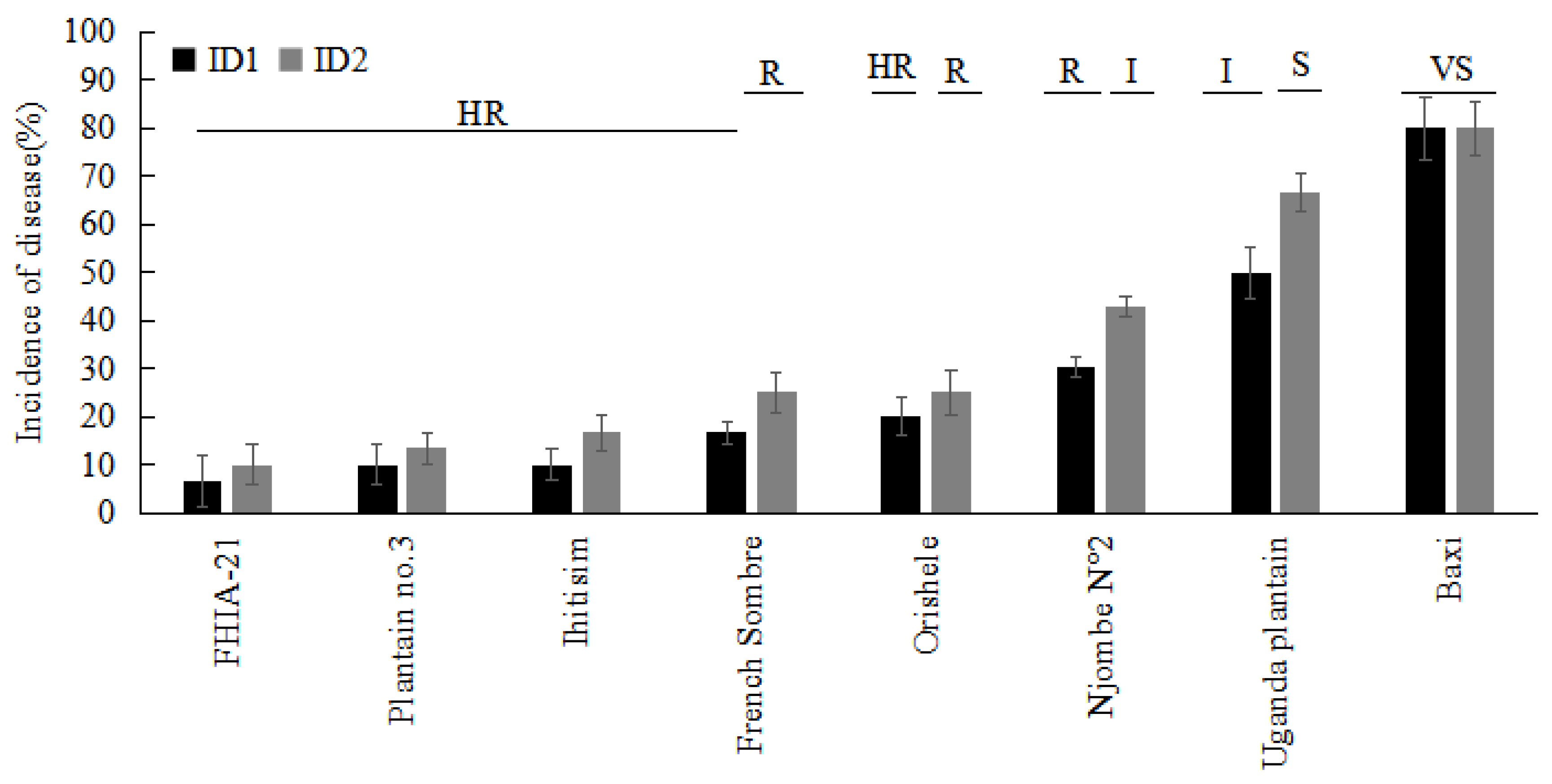
| Cultivar | Subgroup/Type | Greenhouse | Plant Crop | First Ratoon |
|---|---|---|---|---|
| RDI—Rating | ID (%)—Rating | ID (%)—Rating | ||
| Baxi | Cavendish | 6.00—S | 80.00—VS | 80.00—VS |
| Big Ebanga | Plantain/False Horn | 1.00—R | 0.00—HR | 0.00—HR |
| CB5 | Plantain/False Horn | 1.00—R | 0.00—HR | 0.00—HR |
| CEMSA3/4 | Plantain/False Horn | 1.25—R | 0.00—HR | 0.00—HR |
| Curare | Plantain/False Horn | 1.00—R | 0.00—HR | 0.00—HR |
| Orishele | Plantain/False Horn | 2.25—I | 20.00—HR | 25.00—R |
| Batard | Plantain/ French Horn | 1.29—R | 0.00—HR | 0.00—HR |
| Batard 2 | Plantain/ French Horn | 1.00—R | 0.00—HR | 0.00—HR |
| Plantain no.3 | Plantain/ French Horn | 2.00—I | 10.00—HR | 13.33—HR |
| French P | Plantain/French | 1.00—R | 0.00—HR | 0.00—HR |
| French Sombre | Plantain/French | 3.00—I | 16.67—HR | 25.00—R |
| Kakira | Plantain/French | 1.50—R | 0.00—HR | 0.00—HR |
| Nakatansese | Plantain/French | 1.60—R | 0.00—HR | 0.00—HR |
| Njombe N°2 | Plantain/French | 2.75—I | 30.33—R | 42.86—I |
| Ntanga 4 | Plantain/French | 1.50—R | 0.00—HR | 0.00—HR |
| Obubit Ntanga green mutant | Plantain/French | 1.00—R | 0.00—HR | 0.00—HR |
| Uganda Plantain | Plantain/French | 4.00—S | 50.00—I | 66.67—S |
| Ihitisim | Plantain/Horn | 2.00—I | 10.00—HR | 16.67—HR |
| FHIA-21 | Hybrid/Plantain | 1.50—R | 6.67—HR | 10.00—HR |
| Kofi | Iholena | 2.00—I | 6.67—HR | 18.18—HR |
| Luba | Iholena | 3.75—S | 75.00—S | 100.00—VS |
| Maritú | Iholena | 4.00—S | 50.00—I | 66.67—S |
| Rukumamb | Iholena | 3.50—S | 30.00—R | 48.25—I |
| Tigua | Iholena | 3.25—S | 75.00—S | 83.33—VS |
| Uzakan | Iholena | 4.33—S | 72.25—S | 80.00—VS |
| Wisu | Iholena | 4.00—S | 90.00—VS | 100.00—VS |
| Pacific Plantain | Maia Maoli/Popoulu | 5.00—S | 100.00—VS | 100.00—VS |
| Poingo | Maia Maoli/Popoulu | 1.00—R | 0.00—HR | 0.00—HR |
| Pisang Raja Bulu | Pisang Raja | 3.50—S | 66.67—S | 83.33—VS |
| Pisang Raja No.2 | Pisang Raja | 3.00—I | 50.00—I | 53.00—I |
| Pisang Rajah | Pisang Raja | 5.00—S | 50.00—I | 67.66—S |
| YN2 | Pisang Raja | 3.00—I | 16.67—HR | 25.00—R |
| Pisang Raja | Pome | 1.00—R | 0.00—HR | 0.00—HR |
| Pisang Ceylan | Mysore | 2.50—I | 42.75—I | 50.00—I |
| Amrithapani | Silk | 5.25—S | 82.25—VS | 100.00—VS |
| Digjowa | Silk | 5.00—S | 85.00—VS | 100.00—VS |
| Figue Pomme Géante | Silk | 4.00—S | 86.67—VS | 100.00—VS |
| Malbhog | Silk | 4.20—S | 66.67—S | 71.43—S |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhan, N.; Kuang, M.; He, W.; Deng, G.; Liu, S.; Li, C.; Roux, N.; Dita, M.; Yi, G.; Sheng, O. Evaluation of Resistance of Banana Genotypes with AAB Genome to Fusarium Wilt Tropical Race 4 in China. J. Fungi 2022, 8, 1274. https://doi.org/10.3390/jof8121274
Zhan N, Kuang M, He W, Deng G, Liu S, Li C, Roux N, Dita M, Yi G, Sheng O. Evaluation of Resistance of Banana Genotypes with AAB Genome to Fusarium Wilt Tropical Race 4 in China. Journal of Fungi. 2022; 8(12):1274. https://doi.org/10.3390/jof8121274
Chicago/Turabian StyleZhan, Ni, Mengyu Kuang, Weidi He, Guiming Deng, Siwen Liu, Chunyu Li, Nicolas Roux, Miguel Dita, Ganjun Yi, and Ou Sheng. 2022. "Evaluation of Resistance of Banana Genotypes with AAB Genome to Fusarium Wilt Tropical Race 4 in China" Journal of Fungi 8, no. 12: 1274. https://doi.org/10.3390/jof8121274