
First Record of Aspergillus fijiensis as an Entomopathogenic Fungus against Asian Citrus Psyllid, Diaphorina citri Kuwayama (Hemiptera: Liviidae)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. D. citri Cadaver Collection and Isolation of Aspergillus Species
2.2. Morphological Observation
2.3. DNA Extraction and Phylogenetic Analysis of the GDIZM-1 Strain
2.4. Detection of Aflatoxin B1
2.5. Plants and Insects for Testing
2.6. Pathogenicity Test
2.6.1. Laboratory Bioassays on D. citri
2.6.2. Greenhouse Bioassays on D. citri
2.7. Data Analysis
3. Results
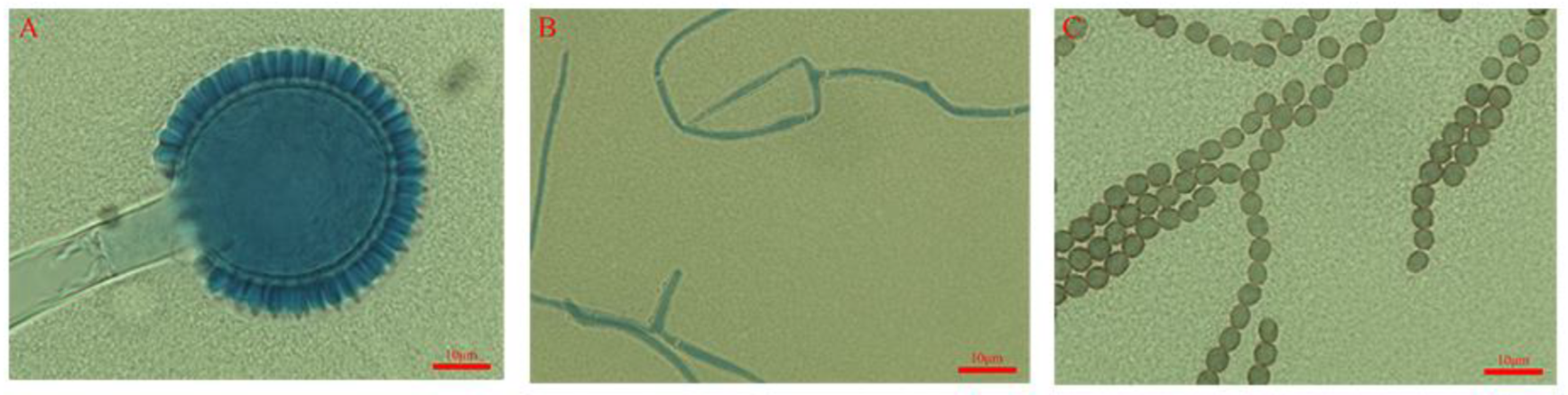
3.1. Morphological Identification of Infected D. citri
3.2. Sequencing and Phylogenetic Analysis
3.3. Aflatoxin B1 Detection
3.4. Pathogenicity Analysis of the GDIZM-1 Strain against D. citri
3.4.1. Pathogenicity Determination in the Laboratory
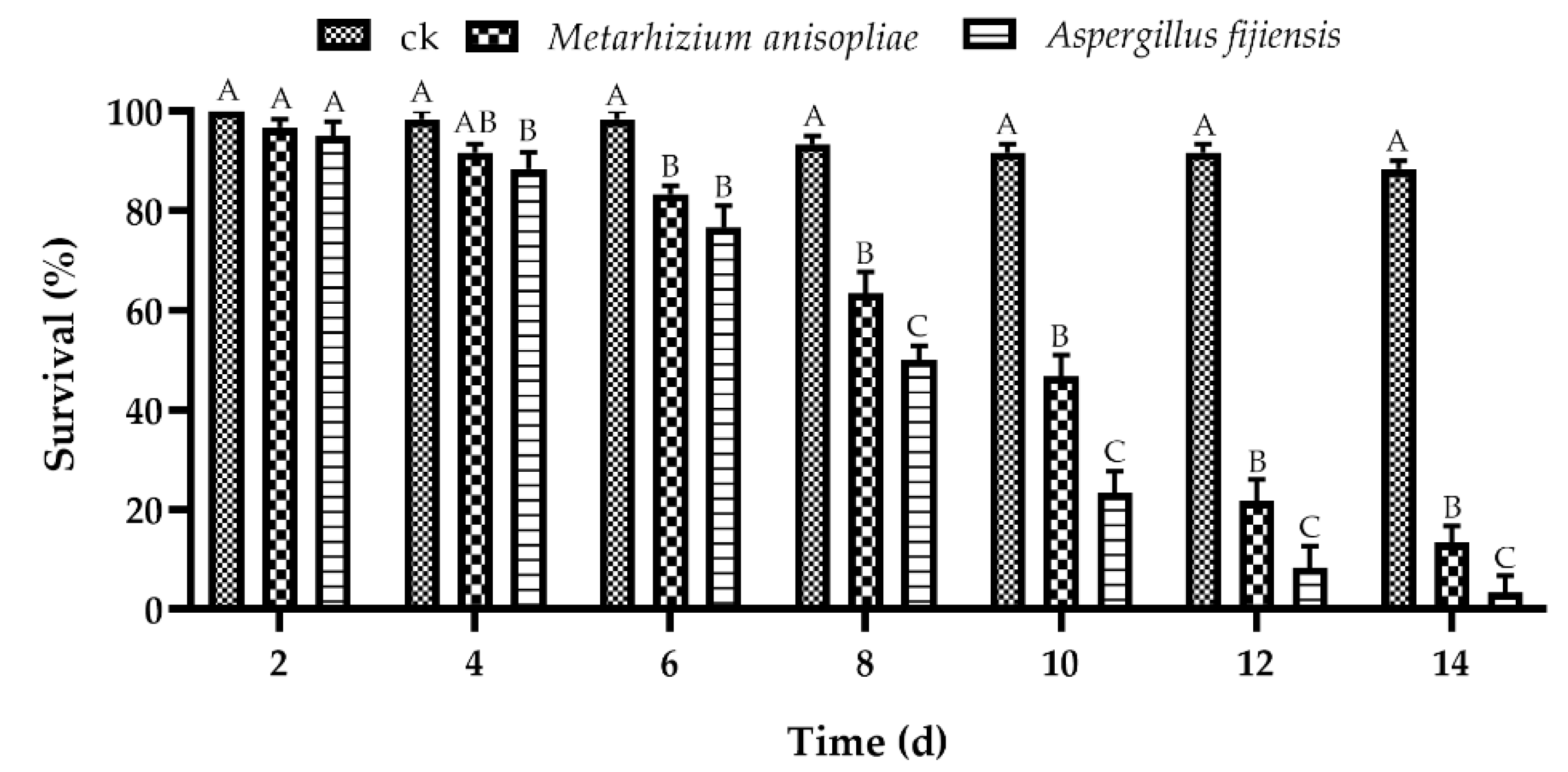
3.4.2. Efficacy of A. fijiensis and M. anisopliae against D. citri in Greenhouse Trials
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nava, D.E.; Torres, M.L.G.; Rodrigues, M.D.L.; Bento, J.M.S.; Parra, J.R.P. Biology of Diaphorina citri (Hem., Psyllidae) on different hosts and at different temperatures. J. Appl. Entomol. 2007, 131, 709–715. [Google Scholar] [CrossRef]
- Hall, D.G.; Richardson, M.L.; Ammar, E.D.; Halbert, S.E. Asian citrus psyllid, Diaphorina citri, vector of citrus huanglongbing disease. Entomol. Exp. Appl. 2013, 146, 207–223. [Google Scholar] [CrossRef]
- Ammar, E.D.; Hall, D.G.; Hosseinzadeh, S.; Heck, M. The quest for a non-vector psyllid: Natural variation in acquisition and transmission of the huanglongbing pathogen ‘Candidatus Liberibacter asiaticus’ by Asian citrus psyllid isofemale lines. PLoS ONE 2018, 13, e0195804. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.H.; Liu, Y.H. Biology of Diaphorina citri (Homoptera: Psyllidae) on four host plants. J. Econ. Entomol. 2000, 93, 1721–1725. [Google Scholar] [CrossRef] [PubMed]
- Boina, D.R.; Bloomquist, J.R. Chemical control of the Asian citrus psyllid and of huanglongbing disease in citrus. Pest Manag. Sci. 2015, 71, 808–823. [Google Scholar] [CrossRef]
- Chen, X.D.; Stelinski, L.L. Resistance management for Asian citrus psyllid, Diaphorina citri Kuwayama, in Florida. Insects 2017, 8, 103. [Google Scholar] [CrossRef] [Green Version]
- Naeem, A.; Afzal, M.B.S.; Freed, S.; Hafeez, F.; Zaka, S.M.; Ali, Q.; Anwar, H.M.Z.; Iftikhar, A.; Nawaz, M. First report of thiamethoxam resistance selection, cross resistance to various insecticides and realized heritability in Asian citrus psyllid Diaphorina citri from Pakistan. Crop Prot. 2019, 121, 11–17. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Zhao, Q.; Wang, C.; Cui, Y.; Li, J.; Chen, A.; Liang, G.; Jiao, B. Occurrence, temporal variation, quality and safety assessment of pesticide residues on citrus fruits in China. Chemosphere 2020, 258, 127381. [Google Scholar] [CrossRef]
- Idrees, A.; Afzal, A.; Qadir, Z.A.; Li, J. Bioassays of Beauveria bassiana Isolates against the Fall Armyworm, Spodoptera frugiperda. J. Fungi 2022, 8, 717. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, S.; Bhandari, D.; Gautam, M.P. Entomopathogens, pathological symptoms and their role in present scenario of agriculture: A review. Int. J. Curr. Microbiol. App. Sci. 2020, 9, 2110–2124. [Google Scholar] [CrossRef]
- Deka, B.; Baruah, C.; Babu, A. Entomopathogenic microorganisms: Their role in insect pest management. Egypt. J. Biol. Pest Control 2021, 31, 121. [Google Scholar] [CrossRef]
- Idrees, A.; Qadir, Z.A.; Akutse, K.S.; Afzal, A.; Hussain, M.; Islam, W.; Waqas, M.S.; Bamisile, B.S.; Li, J. Effectiveness of Entomopathogenic Fungi on Immature Stages and Feeding Performance of Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) Larvae. Insects 2021, 12, 1044. [Google Scholar] [CrossRef] [PubMed]
- Wosten, H.A.B. Hydrophobins: Multipurpose proteins. Annu. Rev. Microbiol. 2001, 55, 625–646. [Google Scholar] [CrossRef] [Green Version]
- Holder, D.J.; Keyhani, N.O. Adhesion of the entomopathogenic fungus Beauveria (Cordyceps) bassiana to substrata. Appl. Environ. Microbiol. 2005, 71, 5260–5266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the surface: Entomopathogenic fungi versus the insect cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillespie, J.P.; Bailey, A.M.; Cobb, B.; Vilcinskas, A. Fungi as elicitors of insect immune responses. Arch. Insect Biochem. Physiol. 2000, 44, 49–68. [Google Scholar] [CrossRef]
- Kershaw, M.J.; Moorhouse, E.R.; Bateman, R.; Reynolds, S.E.; Charnley, A.K. The role of destruxins in the pathogenicity of Metarhizium anisopliae for three species of insect. J. Invertebr. Pathol. 1999, 74, 213–223. [Google Scholar] [CrossRef]
- Qu, S.; Wang, S. Interaction of entomopathogenic fungi with the host immune system. Dev. Comp. Immunol. 2018, 83, 96–103. [Google Scholar] [CrossRef]
- Mora, M.A.E.; Castilho, A.M.C.; Fraga, M.E. Classification and infection mechanism of entomopathogenic fungi. Arq. Inst. Biol. 2018, 84, e0552015. [Google Scholar] [CrossRef] [Green Version]
- Conceschi, M.R.; D’Alessandro, C.P.; Moral, R.d.A.; Demétrio, C.G.B.; Júnior, I.D. Transmission potential of the entomopathogenic fungi Isaria fumosorosea and Beauveria bassiana from sporulated cadavers of Diaphorina citri and Toxoptera citricida to uninfected D. citri adults. BioControl 2016, 61, 567–577. [Google Scholar] [CrossRef]
- Ou, D.; Zhang, L.H.; Guo, C.F.; Chen, X.S.; Ali, S.; Qiu, B.L. Identification of a new Cordyceps javanica fungus isolate and its toxicity evaluation against Asian citrus psyllid. Microbiologyopen 2019, 8, e00760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, W.; Adnan, M.; Shabbir, A.; Naveed, H.; Abubakar, Y.S.; Qasim, M.; Tayyab, M.; Noman, A.; Nisar, M.S.; Khan, K.A.; et al. Insect-fungal-interactions: A detailed review on entomopathogenic fungi pathogenicity to combat insect pests. Microb. Pathog. 2021, 159, 105122. [Google Scholar] [CrossRef] [PubMed]
- Batta, Y.A.; Kavallieratos, N.G. The use of entomopathogenic fungi for the control of stored-grain insects. Int. J. Pest Manag. 2017, 64, 77–87. [Google Scholar] [CrossRef]
- Jiang, W.; Peng, Y.; Ye, J.; Wen, Y.; Liu, G.; Xie, J. Effects of the entomopathogenic fungus Metarhizium anisopliae on the mortality and immune response of Locusta migratoria. Insects 2019, 11, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, Y.; Park, S.E.; Lee, S.Y.; Kim, J.C.; Kim, J.S. Early-stage defense mechanism of the cotton aphid Aphis gossypii against infection with the insect-killing fungus Beauveria bassiana JEF-544. Front. Immunol. 2022, 13, 907088. [Google Scholar] [CrossRef]
- Kim, J.S.; Je, Y.H.; Skinner, M.; Parker, B.L. An oil-based formulation of Isaria fumosorosea blastospores for management of greenhouse whitefly Trialeurodes vaporariorum (Homoptera: Aleyrodidae). Pest Manag. Sci. 2013, 69, 576–581. [Google Scholar] [CrossRef]
- Cortez-Madrigal, H.; Sánchez-Saavedra, J.M.; Díaz-Godínez, G.; Mora-Aguilera, G. Enzymatic activity and pathogenicity of entomopathogenic fungi from central and southeastern Mexico to Diaphorina citri (Hemiptera: Psyllidae). Southwest. Entomol. 2014, 39, 491–502. [Google Scholar] [CrossRef]
- Gandarilla-Pacheco, F.L.; Galán-Wong, L.J.; López-Arroyo, J.I.; Rodríguez-Guerra, R.; Quintero-Zapata, I. Optimization of pathogenicity tests for selection of native isolates of entomopathogenic fungi isolated from citrusgrowing areas of México on adults of Diaphorina citri Kuwayama (Hemiptera: Liviidae). Fla. Entomol. 2013, 96, 187–195. [Google Scholar] [CrossRef]
- Hussain, M.; Akutse, K.S.; Lin, Y.; Chen, S.; Huang, W.; Zhang, J.; Idrees, A.; Qiu, D.; Wang, L. Susceptibilities of Candidatus Liberibacter asiaticus-infected and noninfected Diaphorina citri to entomopathogenic fungi and their detoxification enzyme activities under different temperatures. Microbiologyopen 2018, 7, e00607. [Google Scholar] [CrossRef]
- Naeem, A.; Freed, S.; Akmal, M. Biochemical analysis and pathogenicity of entomopathogenic fungi to Diaphorina citri Kuwayama (Hemiptera: Liviidae). Entomol. Res. 2020, 50, 245–254. [Google Scholar] [CrossRef]
- Ausique, S.J.J.; D’Alessandro, C.P.; Conceschi, M.R.; Mascarin, G.M.; Delalibera, I. Efficacy of entomopathogenic fungi against adult Diaphorina citri from laboratory to field applications. J. Pest Sci. 2017, 90, 947–960. [Google Scholar] [CrossRef]
- Ibarra-Cortes, K.H.; Guzman-Franco, A.W.; Gonzalez-Hernandez, H.; Ortega-Arenas, L.D.; Villanueva-Jimenez, J.A.; Robles-Bermudez, A. Susceptibility of Diaphorina citri (Hemiptera: Liviidae) and its parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) to entomopathogenic fungi under laboratory conditions. Neotrop. Entomol. 2018, 47, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; You, Y.; Song, Y.; Wang, Y.; Zhang, L. First record of Aspergillus oryzae (Eurotiales: Trichocomaceae) as an entomopathogenic fungus of the locust, Locusta migratoria (Orthoptera: Acrididae). Biocontrol Sci. Technol. 2015, 25, 1285–1298. [Google Scholar] [CrossRef]
- Lin, W.J.; Chiu, M.C.; Lin, C.C.; Chung, Y.K.; Chou, J.Y. Efficacy of Entomopathogenic fungus Aspergillus nomius against Dolichoderus thoracicus. BioControl 2021, 66, 463–473. [Google Scholar] [CrossRef]
- Karthi, S.; Vaideki, K.; Shivakumar, M.S.; Ponsankar, A.; Thanigaivel, A.; Chellappandian, M.; Vasantha-Srinivasan, P.; Muthu-Pandian, C.K.; Hunter, W.B.; Senthil-Nathan, S. Effect of Aspergillus flavus on the mortality and activity of antioxidant enzymes of Spodoptera litura Fab.(Lepidoptera: Noctuidae) larvae. Pestic. Biochem. Physiol. 2018, 149, 54–60. [Google Scholar] [CrossRef]
- Ali, S.; Zhang, C.; Wang, Z.Q.; Wang, X.M.; Wu, J.H.; Cuthbertson, A.G.S.; Shao, Z.F.; Qiu, B.L. Toxicological and biochemical basis of synergism between the entomopathogenic fungus Lecanicillium muscarium and the insecticide matrine against Bemisia tabaci (Gennadius). Sci. Rep. 2017, 7, 46558. [Google Scholar] [CrossRef]
- Barra, P.; Rosso, L.; Nesci, A.; Etcheverry, M. Isolation and identification of entomopathogenic fungi and their evaluation against Tribolium confusum, Sitophilus zeamais, and Rhyzopertha dominica in stored maize. J. Pest Sci. 2012, 86, 217–226. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, L.; Dong, G.; Chen, F. Isolation and identification of entomopathogenic fungi and an evaluation of their actions against the larvae of the fall webworm, Hyphantria cunea (Drury)(Lepidoptera: Arctiidae). BioControl 2019, 65, 101–111. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Rehner, S.A.; Minnis, A.M.; Sung, G.H.; Luangsa-ard, J.J.; Devotto, L.; Humber, R.A. Phylogeny and systematics of the anamorphic, entomopathogenic genus Beauveria. Mycologia 2011, 103, 1055–1073. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Tamura, K.; Nei, M. MEGA: Molecular Evolutionary Genetics Analysis software for microcomputers. Bioinformatics 1994, 10, 189–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Sun, Z. Random local neighbor joining: A new method for reconstructing phylogenetic trees. Mol. Phylogenet. Evol. 2008, 47, 117–128. [Google Scholar] [CrossRef]
- Zharkikh, A.; Li, W.H. Estimation of confidence in phylogeny: The complete-and-partial bootstrap technique. Mol. Phylogenet. Evol. 1995, 4, 44–63. [Google Scholar] [CrossRef]
- Wu, J.; Yang, B.; Xu, J.; Cuthbertson, A.G.S.; Ali, S. Characterization and toxicity of crude toxins produced by cordyceps fumosorosea against Bemisia tabaci (Gennadius) and Aphis craccivora (Koch). Toxins 2021, 13, 220. [Google Scholar] [CrossRef]
- Wang, C.; Huang, Y.; Zhao, J.; Ma, Y.; Xu, X.; Wan, Q.; Li, H.; Yu, H.; Pan, B. First record of Aspergillus oryzae as an entomopathogenic fungus against the poultry red mite Dermanyssus gallinae. Vet. Parasitol. 2019, 271, 57–63. [Google Scholar] [CrossRef]
- Liu, B.H.; Hsu, Y.T.; Lu, C.C.; Yu, F.Y. Detecting aflatoxin B1 in foods and feeds by using sensitive rapid enzyme-linked immunosorbent assay and gold nanoparticle immunochromatographic strip. Food Control 2013, 30(1), 184–189. [Google Scholar] [CrossRef]
- Liang, G.; Fu, W.; Wang, K. Analysis of t-test misuses and spss operations in medical research papers. Burn. Trauma 2019, 7, 31. [Google Scholar] [CrossRef] [Green Version]
- Carmo-Sousa, M.; Cortés, M.T.B.; Lopes, J.R.S. Understanding psyllid transmission of Candidatus Liberibacter as a basis for managing huanglongbing. Trop. Plant Pathol. 2020, 45, 572–585. [Google Scholar] [CrossRef]
- Graham, J.; Gottwald, T.; Setamou, M. Status of huanglongbing (HLB) outbreaks in Florida, California and Texas. Trop. Plant Pathol. 2020, 45, 265–278. [Google Scholar] [CrossRef]
- Idrees, A.; Qadir, Z.A.; Afzal, A.; Ranran, Q.; Li, J. Laboratory efficacy of selected synthetic insecticides against second instar invasive fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae. PLoS ONE 2022, 17, e0265265. [Google Scholar] [CrossRef] [PubMed]
- Wendel, J.; Cisneros, J.; Jaronski, S.; Vitek, C.; Ciomperlik, M.; Flores, D. Screening commercial entomopathogenic fungi for the management of Diaphorina citri populations in the Lower Rio Grande Valley, Texas, USA. BioControl 2022, 67, 225–235. [Google Scholar] [CrossRef]
- Orduño-Cruz, N.; Guzmán-Franco, A.W.; Rodríguez-Leyva, E.; Alatorre-Rosas, R.; González-Hernández, H.; Mora-Aguilera, G. In vivo selection of entomopathogenic fungal isolates for control of Diaphorina citri (Hemiptera: Liviidae). Biol. Control 2015, 90, 1–5. [Google Scholar] [CrossRef]
- Ojeda-Lopez, M.; Chen, W.; Eagle, C.E.; Gutierrez, G.; Jia, W.L.; Swilaiman, S.S.; Huang, Z.; Park, H.S.; Yu, J.H.; Canovas, D.; et al. Evolution of asexual and sexual reproduction in the Aspergilli. Stud. Mycol. 2018, 91, 37–59. [Google Scholar] [CrossRef]
- Hazen, K.C. Methods for fungal identification in the clinical mycology laboratory. Clin. Microbiol. Newsl. 1996, 18, 137–141. [Google Scholar] [CrossRef]
- Aharwar, A.; Parihar, D.K. Talaromyces verruculosus tannase production, characterization and application in fruit juices detannification. Biocatal. Agric. Biotechnol. 2019, 18, 101014. [Google Scholar] [CrossRef]
- Klich, M.; Mendoza, C.; Mullaney, E.; Keller, N.; Bennett, J.W. A new sterigmatocystin-producing emericella variant from agricultural desert soils. Syst. Appl. Microbiol. 2001, 24, 131–138. [Google Scholar] [CrossRef]
- Balajee, S.A.; Houbraken, J.; Verweij, P.E.; Hong, S.B.; Yaghuchi, T.; Varga, J.; Samson, R.A. Aspergillus species identification in the clinical setting. Stud. Mycol. 2007, 59, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Varga, J.; Frisvad, J.C.; Kocsube, S.; Brankovics, B.; Toth, B.; Szigeti, G.; Samson, R.A. New and revisited species in Aspergillus section Nigri. Stud. Mycol. 2011, 69, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Coetzee, G.; Smith, J.J.; Görgens, J.F. Influence of codon optimization, promoter, and strain selection on the heterologous production of a β-fructofuranosidase from Aspergillus fijiensis ATCC 20611 in Pichia pastoris. Folia Microbiol. 2022, 67, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, C.N.; Maurya, P.; Sharma, P.; Mohan, L. Prospective role of insecticides of fungal origin: Review. Entomol. Res. 2009, 39, 341–355. [Google Scholar] [CrossRef]
- Mensah, R.K.; Young, A. Microbial control of cotton pests: Use of the naturally occurring entomopathogenic fungus Aspergillus sp. (bc 639) in the management of Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae) and beneficial insects on transgenic cotton crops. Biocontrol Sci. Technol. 2017, 27, 844–866. [Google Scholar] [CrossRef]
- Kaur, T.; Kaur, J.; Kaur, A.; Kaur, S. Larvicidal and growth inhibitory effects of endophytic Aspergillus niger on a polyphagous pest, Spodoptera litura. Phytoparasitica 2016, 44, 465–476. [Google Scholar] [CrossRef]
- Cruz-Juárez, G.; Maldonado-Blanco, M.G.; Rodríguez-Guerra, R.; de la Torre-Zavala, S.; Avilés-Arnaut, H.; Flores-González, M.d.S. Mutation to increase sporulation of a strain of Hirsutella citriformis from Mexico and evaluation against Diaphorina citri. Southwest. Entomol. 2018, 43, 891–904. [Google Scholar] [CrossRef]
- Santos, M.S.; Zanardi, O.Z.; Pauli, K.S.; Forim, M.R.; Yamamoto, P.T.; Vendramim, J.D. Toxicity of an azadirachtin-based biopesticide on Diaphorina citri Kuwayama (Hemiptera: Liviidae) and its ectoparasitoid Tamarixia radiata (Waterston) (Hymenoptera: Eulophidae). Crop Prot. 2015, 74, 116–123. [Google Scholar] [CrossRef]
- Cafarchia, C.; Immediato, D.; Iatta, R.; Ramos, R.A.; Lia, R.P.; Porretta, D.; Figueredo, L.A.; Dantas-Torres, F.; Otranto, D. Native strains of Beauveria bassiana for the control of Rhipicephalus sanguineus sensu lato. Parasites Vectors 2015, 8, 80. [Google Scholar] [CrossRef] [Green Version]
- Meyer, J.M.; Hoy, M.A.; Boucias, D.G. Isolation and characterization of an Isaria fumosorosea isolate infecting the Asian citrus psyllid in Florida. J. Invertebr. Pathol. 2008, 99, 96–102. [Google Scholar] [CrossRef]
- Freimoser, F.M.; Hu, G.; St. Leger, R.J. Variation in gene expression patterns as the insect pathogen Metarhizium anisopliae adapts to different host cuticles or nutrient deprivation in vitro. Microbiology 2005, 151, 361–371. [Google Scholar] [CrossRef] [Green Version]
- Bidochka, M.J.; Small, C.L.N.; Spironello, M. Recombination within sympatric cryptic species of the insect pathogenic fungus Metarhizium anisopliae. Environ. Microbiol. 2005, 7, 1361–1368. [Google Scholar] [CrossRef]
- Lord, J.C. Low humidity, moderate temperature, and desiccant dust favor efficacy of Beauveria bassiana (Hyphomycetes: Moniliales) for the lesser grain borer, Rhyzopertha dominica (Coleoptera: Bruchidae). Biol. Control. 2005, 34, 180–186. [Google Scholar] [CrossRef]
- Erler, F.; Ates, A.O. Potential of two entomopathogenic fungi, Beauveria bassiana and Metarhizium anisopliae (Coleoptera: Scarabaeidae), as biological control agents against the June beetle. J. Insect Sci. 2015, 15, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Oh, J.; Yoon, D.H.; Sung, G.H. Suppression of a methionine synthase by calmodulin under environmental stress in the entomopathogenic fungus Beauveria bassiana. Environ. Microbiol. Rep. 2017, 9, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yang, B.; Zhang, X.; Cuthbertson, A.G.S.; Ali, S. Synergistic interaction between the entomopathogenic fungus Akanthomyces attenuatus (Zare & Gams) and the botanical insecticide Matrine against Megalurothrips usitatus (Bagrall). J. Fungi 2021, 7, 536. [Google Scholar] [CrossRef]
- Ghallab, A.; Hassan, R.; Myllys, M.; Albrecht, W.; Friebel, A.; Hoehme, S.; Hofmann, U.; Seddek, A.l.; Braeuning, A.; Kuepfer, L.; et al. Subcellular spatio-temporal intravital kinetics of aflatoxin B1 and ochratoxin A in liver and kidney. Arch. Toxicol. 2021, 95, 2163–2177. [Google Scholar] [CrossRef]
- Fouad, A.M.; Ruan, D.; El-Senousey, H.A.K.; Chen, W.; Jiang, S.Q.; Zheng, C.T. Harmful effects and control strategies of aflatoxin b1 produced by Aspergillus flavus and Aspergillus parasiticus strains on poultry: Review. Toxins 2019, 11, 176. [Google Scholar] [CrossRef] [Green Version]
- Rustom, I.Y.S. Aflatoxin in food and feed: Occurrence, legislation and inactivation by physical methods. Food Chem. 1997, 59, 57–67. [Google Scholar] [CrossRef]
- Wood, G.E. Mycotoxins in foods and feeds in the United States. J. Anim. Sci. 1992, 70, 3941–3949. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Yoshizawa, T.; Kawamura, O.; Luo, X.; Li, Y. Aflatoxins and fumonisins in corn from the high-incidence area for human hepatocellular carcinoma in Guangxi, China. J. Agric. Food Chem. 2001, 49, 4122–4126. [Google Scholar] [CrossRef]
- Wild, C.P.; Gong, Y.Y. Mycotoxins and human disease: A largely ignored global health issue. Carcinogenesis 2010, 31, 71–82. [Google Scholar] [CrossRef]
- Wu, H.C.; Wang, Q.; Yang, H.I.; Ahsan, H.; Tsai, W.Y.; Wang, L.Y.; Chen, S.Y.; Chen, C.J.; Santella, R.M. Aflatoxin B1 exposure, hepatitis B virus infection, and hepatocellular carcinoma in Taiwan. Cancer Epidemiol. Biomarkers Prev. 2009, 18, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Creppy, E.E. Update of survey, regulation and toxic effects of mycotoxins in Europe. Toxicol. Lett. 2002, 127, 19–28. [Google Scholar] [CrossRef]
- Anttila, A.; Bhat, R.V.; Bond, J.A.; Borghoff, S.J.; Bosch, F.X.; Carlson, G.P.; Castegnaro, M.; Cruzan, G.; Gelderblom, W.C.A.; Hass, U.; et al. Some traditional herbal medicines, some mycotoxins, naphthalene and styrene. IARC Monogr. Eval. Carcinog. Risks Hum. 2002, 82, 1–556. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | GeneBank Accession Number | ||
|---|---|---|---|
| ITS | TEF1-α | RPB2 | |
| Aspergillus brunneoviolaceus | MT102843 | HE984384 | KX650010 |
| Aspergillus aculeatus | KY320594 | HE984398 | MK340898 |
| Aspergillus japonicus | KX621981 | HE984394 | MN969079 |
| Aspergillus fijiensis | MH856458 | HE984402 | HE984375 |
| Aspergillus fijiensis | OM925539 * | ON000912 * | ON000911 * |
| Aspergillus uvarum | MZ541955 | HE984397 | HE984364 |
| Emericella acristata | EF652446 | KM882998 | KU867032 |
| Strain | Colony Diameter (mm) | Growth Rate (mm/d) | Sporulation (×108 Conidia/mL) |
|---|---|---|---|
| GDIZM-1 | 83.70 ± 0.47 | 16.74 ± 0.10 | 3.75 ± 0.30 |
| Insect Stages | Toxicity Regression Equation | LC50 (Conidia/mL) | 95% Confidence Interval (Conidia/mL) | χ2 | p |
|---|---|---|---|---|---|
| 1st~2nd instar nymph | 6.40 × 103 | 5.08 × 10−1~7.40 × 104 | 0.15 | 0.99 | |
| 3rd~4th instar nymph | 1.15 × 104 | 1.81 × 10−1~1.44 × 105 | 0.22 | 0.98 | |
| 5th instar nymph | 1.20 × 105 | 0.86 × 103~9.76 × 105 | 0.08 | 0.99 | |
| Adult | 2.77 × 105 | 5.45 × 103~2.48 × 106 | 0.48 | 0.92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, J.; Liu, H.; Idrees, A.; Chen, F.; Lu, H.; Ouyang, G.; Meng, X. First Record of Aspergillus fijiensis as an Entomopathogenic Fungus against Asian Citrus Psyllid, Diaphorina citri Kuwayama (Hemiptera: Liviidae). J. Fungi 2022, 8, 1222. https://doi.org/10.3390/jof8111222
Yan J, Liu H, Idrees A, Chen F, Lu H, Ouyang G, Meng X. First Record of Aspergillus fijiensis as an Entomopathogenic Fungus against Asian Citrus Psyllid, Diaphorina citri Kuwayama (Hemiptera: Liviidae). Journal of Fungi. 2022; 8(11):1222. https://doi.org/10.3390/jof8111222
Chicago/Turabian StyleYan, Jianquan, Hao Liu, Atif Idrees, Fenghao Chen, Huilin Lu, Gecheng Ouyang, and Xiang Meng. 2022. "First Record of Aspergillus fijiensis as an Entomopathogenic Fungus against Asian Citrus Psyllid, Diaphorina citri Kuwayama (Hemiptera: Liviidae)" Journal of Fungi 8, no. 11: 1222. https://doi.org/10.3390/jof8111222