
Genetic Engineering of Talaromyces marneffei to Enhance Siderophore Production and Preliminary Testing for Medical Application Potential

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Fungal Strains and Culture Conditions
2.3. Generation of sreA Mutant Strain
2.4. Germination Assay
2.5. Chrome Azurol S (CAS) Assay
2.6. Purification of the Extracellular Siderophore
2.7. Characterization of the Purified Siderophore
2.8. Cytotoxicity
2.9. Labile Iron Pool (LIP) Assay
2.10. Antimicrobial Activity Testing by Broth Microdilution Assay
2.11. Statistical Analysis
3. Results
3.1. Characteristics of Talaromyces Marneffei ∆sreA Strain
3.2. Deletion of sreA Increased the Synthesis of Siderophores
3.3. Characterization of Secreted Siderophore of ΔsreA by RP-HPLC, LC-MS, FTIR, and NMR
3.4. Testing of Cytotoxic Effect and Iron Chelating Property of Coprogen B
3.4.1. Effect of Coprogen B on Cell Viability
3.4.2. Potential of Cellular Iron-Chelating Activity
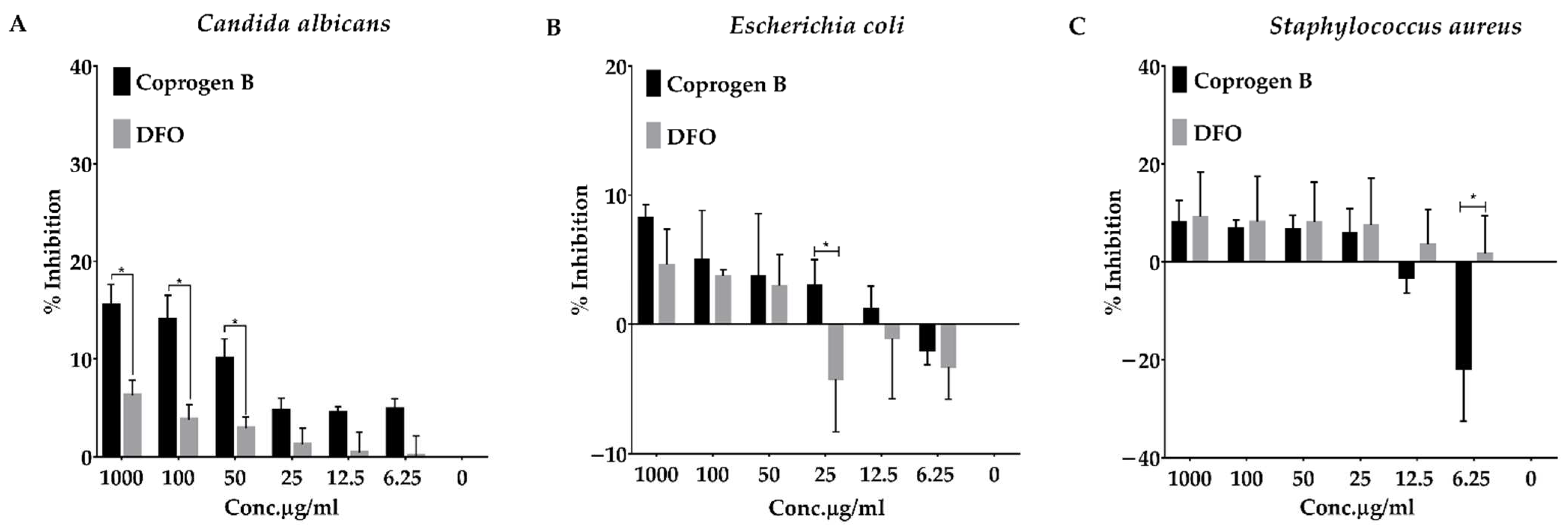
3.5. Evaluation of Ability of Coprogen B to Inhibit Growth of Escherichia coli, Staphylococcus aureus, and Candida albicans
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagoba, B.; Vedpathakv, D.V. Medical applications of siderophores–a review. Eur. J. Gen. Med. 2011, 8, 230–233. [Google Scholar]
- Diarra, M.; Lavoie, M.; Jacques, M.; Darwish, I.; Dolence, E.; Dolence, J.; Ghosh, A.; Ghosh, M.; Miller, M.; Malouin, F. Species selectivity of new siderophore-drug conjugates that use specific iron uptake for entry into bacteria. Antimicrob. Agents Chemother. 1996, 40, 2610–2617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ioppolo, J.A.; Caldwell, D.; Beiraghi, O.; Llano, L.; Blacker, M.; Valliant, J.F.; Berti, P.J. 67Ga-labeled deferoxamine derivatives for imaging bacterial infection: Preparation and screening of functionalized siderophore complexes. Nucl. Med. Biol. 2017, 52, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Bickel, H.; Gaeumann, E.; Keller-Schierlein, W.; Prelog, V.; Vischer, E.; Wettstein, A.; Zaehner, H. On iron-containing growth factors, sideramines, and their antagonists, the iron-containing antibiotics, sideromycins. Experientia 1960, 16, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Tsamesidis, I.; Fozza, C.; Vagdatli, E.; Kalpaka, A.; Cirotto, C.; Pau, M.C.; Pantaleo, A.; Turrini, F.; Grigoriou, E.; Lymperaki, E. Total antioxidant capacity in Mediterranean beta-thalassemic patients. Adv. Clin. Exp. Med. 2017, 26, 789–793. [Google Scholar] [CrossRef] [Green Version]
- Song, T.S.; Hsieh, Y.W.; Peng, C.T.; Chen, T.L.; Lee, H.Z.; Chung, J.G.; Hour, M.J. Combined versus monotherapy or concurrent therapy for treatment of thalassaemia. Vivo 2014, 28, 645–649. [Google Scholar]
- Britton, R.S.; Leicester, K.L.; Bacon, B.R. Iron toxicity and chelation therapy. Int. J. Hematol. 2002, 76, 219–228. [Google Scholar] [CrossRef]
- Orton, R.; De Veber, L.; Sulh, H. Ocular and auditory toxicity of long-term, high-dose subcutaneous deferoxamine therapy. Can. J. Ophthal. 1985, 20, 153–156. [Google Scholar]
- Sheth, S. Iron chelation: An update. Curr. Opin. Hematol. 2014, 21, 179–185. [Google Scholar] [CrossRef]
- Pasricha, S.; Schafferer, L.; Lindner, H.; Joanne Boyce, K.; Haas, H.; Andrianopoulos, A. Differentially regulated high-affinity iron assimilation systems support growth of the various cell types in the dimorphic pathogen Talaromyces Marneffei. Mol. Microbiol. 2016, 102, 715–737. [Google Scholar] [CrossRef]
- Misslinger, M.; Hortschansky, P.; Brakhage, A.A.; Haas, H. Fungal iron homeostasis with a focus on Aspergillus fumigatus. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118885. [Google Scholar] [CrossRef]
- Schrettl, M.; Bignell, E.; Kragl, C.; Sabiha, Y.; Loss, O.; Eisendle, M.; Wallner, A.; Arst Jr, H.N.; Haynes, K.; Haas, H. Distinct roles for intra-and extracellular siderophores during Aspergillus fumigatus infection. PLoS Pathog. 2007, 3, e128. [Google Scholar] [CrossRef]
- Schrettl, M.; Kim, H.S.; Eisendle, M.; Kragl, C.; Nierman, W.C.; Heinekamp, T.; Werner, E.R.; Jacobsen, I.; Illmer, P.; Yi, H.; et al. SreA-mediated iron regulation in Aspergillus fumigatus. Mol. Microbiol. 2008, 70, 27–43. [Google Scholar] [CrossRef]
- Chi, Z.; Wang, X.X.; Geng, Q.; Chi, Z.M. Role of a GATA-type transcriptional repressor Sre1 in regulation of siderophore biosynthesis in the marine-derived Aureobasidium pullulans HN6. 2. Biometals 2013, 26, 955–967. [Google Scholar] [CrossRef]
- Chung, K.R.; Wu, P.C.; Chen, Y.K.; Yago, J.I. The siderophore repressor SreA maintains growth, hydrogen peroxide resistance, and cell wall integrity in the phytopathogenic fungus Alternaria alternata. Fungal Genet. Biol. 2020, 139, 103384. [Google Scholar] [CrossRef]
- Bugeja, H.E.; Boyce, K.J.; Weerasinghe, H.; Beard, S.; Jeziorowski, A.; Pasricha, S.; Payne, M.; Schreider, L.; Andrianopoulos, A. Tools for high efficiency genetic manipulation of the human pathogen Penicillium marneffei. Fungal Genet. Biol. 2012, 49, 772–778. [Google Scholar] [CrossRef]
- Borneman, A.R.; Hynes, M.J.; Andrianopoulos, A. An STE12 homolog from the asexual, dimorphic fungus Penicillium marneffei complements the defect in sexual development of an Aspergillus nidulans steA mutant. Genetics 2001, 157, 1003–1014. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Paradee, N.; Howes, M.R.; Utama-Ang, N.; Chaikitwattna, A.; Hider, R.C.; Srichairatanakool, S. A chemically characterized ethanolic extract of Thai Perilla frutescens (L.) Britton fruits (nutlets) reduces oxidative stress and lipid peroxidation in human hepatoma (HuH7) cells. Phytother. Res. 2019, 33, 2064–2074. [Google Scholar] [CrossRef]
- Carroll, C.S.; Amankwa, L.N.; Pinto, L.J.; Fuller, J.D.; Moore, M.M. Detection of a serum siderophore by LC-MS/MS as a potential biomarker of invasive aspergillosis. PLoS ONE 2016, 11, e0151260. [Google Scholar] [CrossRef]
- Prus, E.; Fibach, E. Flow cytometry measurement of the labile iron pool in human hematopoietic cells. Cytom. A 2008, 73, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Sarker, S.D.; Nahar, L.; Kumarasamy, Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods 2007, 42, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Konetschny-Rapp, S.; Huschka, H.-G.; Winkelmanne, G.; Jung, G. High-performance liquid chromatography of siderophores from fungi. Biol. Met. 1988, 1, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.K.; Deshattiwar, M.K.; Chaudhari, B.L.; Chincholkar, S.B. Production, purification and chemical characterization of the catecholate siderophore from potent probiotic strains of Bacillus spp. Bioresour. Technol. 2009, 100, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Murugappan, R.; Aravinth, A.; Karthikeyan, M. Chemical and structural characterization of hydroxamate siderophore produced by marine Vibrio harveyi. J. Ind. Microbiol. Biotechnol. 2011, 38, 265–273. [Google Scholar] [CrossRef]
- Antelo, L.; Hof, C.; Welzel, K.; Eisfeld, K.; Sterner, O.; Anke, H. Siderophores produced by Magnaporthe grisea in the presence and absence of iron. Z. Für Nat. C 2006, 61, 461–464. [Google Scholar] [CrossRef]
- Jalal, M.A.; Van der Helm, D. Siderophores of highly phytopathogenic Alternaria longipes. Biol. Met. 1989, 2, 11–17. [Google Scholar] [CrossRef]
- Zhang, J.; Qi, L.; Chen, G.; Yin, W.-B. Discovery and genetic identification of amphiphilic coprogen siderophores from Trichoderm hypoxylon. Appl. Microbiol. Biotech. 2021, 105, 2831–2839. [Google Scholar] [CrossRef]
- Huang, Z.-H.; Liang, X.; Shu-Hua, Q. A new iron (III) chelator of coprogen-type siderophore from the deep-sea-derived fungus Mycosphaerella sp. SCSIO z059. Chin. J. Nat. Med. 2020, 18, 243–249. [Google Scholar] [CrossRef]
- Chuljerm, H.; Deeudom, M.; Fucharoen, S.; Mazzacuva, F.; Hider, R.C.; Srichairatanakool, S.; Cilibrizzi, A. Characterization of two siderophores produced by Bacillus megaterium: A preliminary investigation into their potential as therapeutic agents. Biochim. Biophys. Acta 2020, 1864, 129670. [Google Scholar] [CrossRef]
- Lesic, B.; Foulon, J.; Carniel, E. Comparison of the effects of deferiprone versus deferoxamine on growth and virulence of Yersinia enterocolitica. Antimicrob. Agents Chemother. 2002, 46, 1741–1745. [Google Scholar] [CrossRef] [Green Version]
- Kalinowski, D.S.; Richardson, D.R. The evolution of iron chelators for the treatment of iron overload disease and cancer. Pharmacol. Rev. 2005, 57, 547–583. [Google Scholar] [CrossRef]
- Boda, S.K.; Pandit, S.; Garai, A.; Pal, D.; Basu, B. Bacterial siderophore mimicking iron complexes as DNA targeting antimicrobials. RSC Adv. 2016, 6, 39245–39260. [Google Scholar] [CrossRef]
- Oberegger, H.; Schoeser, M.; Zadra, I.; Abt, B.; Haas, H. SREA is involved in regulation of siderophore biosynthesis, utilization and uptake in Aspergillus nidulans. Mole. Microbiol. 2001, 41, 1077–1089. [Google Scholar] [CrossRef]
- An, Z.; Mei, B.; Yuan, W.M.; Leong, S.A. The distal GATA sequences of the sid1 promoter of Ustilago maydis mediate iron repression of siderophore production and interact directly with Urbs1, a GATA family transcription factor. EMBO J. 1997, 16, 1742–1750. [Google Scholar] [CrossRef] [Green Version]
- Haas, H.; Zadra, I.; Stoffler, G.; Angermayr, K. The Aspergillus nidulans GATA factor SREA is involved in regulation of siderophore biosynthesis and control of iron uptake. J. Biol. Chem. 1999, 274, 4613–4619. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, B.; Beaudoin, J.; Mukai, Y.; Labbé, S. Fep1, an iron sensor regulating iron transporter gene expression in Schizosaccharomyces pombe. J. Biol. Chem. 2002, 277, 22950–22958. [Google Scholar] [CrossRef] [Green Version]
- Hai, Y.; Jenner, M.; Tang, Y. Fungal siderophore biosynthesis catalysed by an iterative nonribosomal peptide synthetase. Chem. Sci. 2020, 11, 11525–11530. [Google Scholar] [CrossRef]
- Hider, R.C.; Kong, X. Chemistry and biology of siderophores. Nat. Prod. Rep. 2010, 27, 637–657. [Google Scholar] [CrossRef]
- Haas, H. Fungal siderophore metabolism with a focus on Aspergillus fumigatus. Nat. Prod. Rep. 2014, 31, 1266–1276. [Google Scholar] [CrossRef] [Green Version]
- Le Govic, Y.; Papon, N.; Le Gal, S.; Bouchara, J.-P.; Vandeputte, P. Non-ribosomal peptide synthetase gene clusters in the human pathogenic fungus Scedosporium apiospermum. Front. Microbiol. 2019, 10, 2062. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.; Jalal, M.; Benson, B.; Barnes, C.; Van der Helm, D. Structure and conformation of two coprogen-type siderophores. Neocoprogen I and neocoprogen II. J. Am. Chem. 1987, 109, 4948–4954. [Google Scholar] [CrossRef]
- Charlang, G.; Ng, B.; Horowitz, N.H.; Horowitz, R.M. Cellular and extracellular siderophores of Aspergillus nidulans and Penicillium chrysogenum. Mol. Cell. Biol. 1981, 1, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Leiter, E.; Emri, T.; Gyémánt, G.; Nagy, I.; Pócsi, I.; Winkelmann, G.; Pócsi, I. Penicillin V production by Penicillium chrysogenum in the presence of Fe3+ and in low-iron culture medium. Folia Microbiol. 2001, 46, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.H.; Lin, C.H.; Chung, K.R. A nonribosomal peptide synthetase mediates siderophore production and virulence in the citrus fungal pathogen Alternaria alternata. Mol. Plant Pathol. 2013, 14, 497–505. [Google Scholar] [CrossRef]
- Voß, B.; Kirschhöfer, F.; Brenner-Weiß, G.; Fischer, R. Alternaria alternata uses two siderophore systems for iron acquisition. Sci. Rep. 2020, 10, 3587. [Google Scholar] [CrossRef] [Green Version]
- Franken, A.C.; Lechner, B.E.; Werner, E.R.; Haas, H.; Lokman, B.C.; Ram, A.F.; van den Hondel, C.A.; de Weert, S.; Punt, P.J. Genome mining and functional genomics for siderophore production in Aspergillus niger. Brief. Funct. Genom. 2014, 13, 482–492. [Google Scholar] [CrossRef] [Green Version]
- Burt, W.R. Identification of coprogen B and its breakdown products from Histoplasma capsulatum. Infect. Immun. 1982, 35, 990–996. [Google Scholar] [CrossRef] [Green Version]
- Wong, G.B.; Kappel, M.J.; Raymond, K.N.; Matzanke, B.; Winkelmann, G. Coordination chemistry of microbial iron transport compounds. 24. Characterization of coprogen and ferricrocin, two ferric hydroxamate siderophores. J. Am. Chem. 1983, 105, 810–815. [Google Scholar] [CrossRef]
- Al Shaer, D.; Al Musaimi, O.; Beatriz, G.; Albericio, F. Hydroxamate siderophores: Natural occurrence, chemical synthesis, iron binding affinity and use as trojan horses against pathogens. Eur. J. Med. Chem. 2020, 208, 112791. [Google Scholar] [CrossRef]
- Emri, T.; Tóth, V.; Nagy, C.T.; Nagy, G.; Pócsi, I.; Gyémánt, G.; Antal, K.; Balla, J.; Balla, G.; Román, G. Towards high-siderophore-content foods: Optimisation of coprogen production in submerged cultures of Penicillium nalgiovense. J. Sci. Food Agric. 2013, 93, 2221–2228. [Google Scholar] [CrossRef]
- De Serrano, L.O. Biotechnology of siderophores in high-impact scientific fields. Biomol. Concepts 2017, 8, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Pócsi, I.; Jeney, V.; Kertai, P.; Pócsi, I.; Emri, T.; Gyémánt, G.; Fésüs, L.; Balla, J.; Balla, G. Fungal siderophores function as protective agents of LDL oxidation and are promising anti-atherosclerotic metabolites in functional food. Mol. Nutr. Food Res. 2008, 52, 1434–1447. [Google Scholar] [CrossRef]
- Mobarra, N.; Shanaki, M.; Ehteram, H.; Nasiri, H.; Sahmani, M.; Saeidi, M.; Goudarzi, M.; Pourkarim, H.; Azad, M. A review on iron chelators in treatment of iron overload syndromes. Int. J. Hematol. Oncol. Stem Cell Res. 2016, 10, 239. [Google Scholar]
- Cabantchik, Z.; Link, G.; Glickstein, H.; El, R.B.; Hershko, C.; Konijn, A.; Nick, H. Deferasirox (Exjade®, ICL670): A journey into labile iron centers of living cardiomyocytes. Blood 2005, 106, 824. [Google Scholar] [CrossRef]
- Glickstein, H.; Ben El, R.; Shvartsman, M.; Cabantchik, Z.I. Intracellular labile iron pools as direct targets of iron chelators: A fluorescence study of chelator action in living cells. Blood 2005, 106, 3242–3250. [Google Scholar] [CrossRef]
- Jomova, K.; Valko, M. Advances in metal-induced oxidative stress and human disease. Toxicology 2011, 283, 65–87. [Google Scholar] [CrossRef]
- Adjimani, J.P.; Asare, P. Antioxidant and free radical scavenging activity of iron chelators. Toxicol. Rep. 2015, 2, 721–728. [Google Scholar] [CrossRef] [Green Version]
- Schubert, S.R.; Autenrieth, I.B. Conjugation of hydroxyethyl starch to desferrioxamine (DFO) modulates the dual role of DFO in Yersinia enterocolitica infection. Clin. Diag. Lab. Immunol. 2000, 7, 457–462. [Google Scholar] [CrossRef] [Green Version]
- Thompson, M.G.; Corey, B.W.; Si, Y.; Craft, D.W.; Zurawski, D.V. Antibacterial activities of iron chelators against common nosocomial pathogens. Antimicrob. Agents Chemother. 2012, 56, 5419–5421. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.-W.; Campbell, L.T.; Wilkins, M.R.; Pang, C.N.I.; Chen, S.; Carter, D.A. Synergy and antagonism between iron chelators and antifungal drugs in Cryptococcus. Int. J. Antimicrob. Agents 2016, 48, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Gokarn, K.; Pal, R.B. Activity of siderophores against drug-resistant gram-positive and gram-negative bacteria. Infect. Drug Resist. 2018, 11, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, H. Iron—A Key Nexus in the Virulence of Aspergillus fumigatus. Front. Microbiol. 2012, 3, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfister, J.; Petrik, M.; Bendova, K.; Matuszczak, B.; Binder, U.; Misslinger, M.; Kühbacher, A.; Gsaller, F.; Haas, H.; Decristoforo, C. Antifungal siderophore conjugates for theranostic applications in invasive pulmonary aspergillosis using low-molecular TAFC scaffolds. J. Fungi 2021, 7, 558. [Google Scholar] [CrossRef]
- Petrik, M.; Haas, H.; Dobrozemsky, G.; Lass-Flörl, C.; Helbok, A.; Blatzer, M.; Dietrich, H.; Decristoforo, C. 68Ga-siderophores for PET imaging of invasive pulmonary aspergillosis: Proof of principle. J. Nucl. Med. 2010, 51, 639–645. [Google Scholar] [CrossRef] [Green Version]
- Petrik, M.; Haas, H.; Laverman, P.; Schrettl, M.; Franssen, G.M.; Blatzer, M.; Decristoforo, C. 68Ga-triacetylfusarinine C and 68Ga-ferrioxamine E for Aspergillus infection imaging: Uptake specificity in various microorganisms. Mol. Imaging Biol. 2014, 16, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Petrik, M.; Umlaufova, E.; Raclavsky, V.; Palyzova, A.; Havlicek, V.; Pfister, J.; Mair, C.; Novy, Z.; Popper, M.; Hajduch, M. 68Ga-labelled desferrioxamine-B for bacterial infection imaging. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 372–382. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1H Chemical Shift δ in D2O (ppm) | 13C Chemical Shift δ in D2O (ppm) | Structural Group |
|---|---|---|---|
| 1 | 168.4 | >C=O | |
| 1′/1” | 169.9 | >C=O | |
| 2/2′/2” | 3.7, m | 52.4 | >CH- |
| 3/3′/3” | 1.6–1.7, m | 27.4 | -CH2-CH2- |
| 4/4′/4” | 1.6–1.7, m | 23.7 | -CH2-CH2- |
| 5/5′/5” | 4.3–4.4, m | 46.8 | -CH2-CH2- |
| 6/6′/6” | 168.4 | >C=O | |
| 7/7′/7” | 6.25, brs | 117.4 | -CH= |
| 8/8′/8” | 151.3 | >C= | |
| 9/9′/9” | 2.8, m | 43.3 | -CH2- |
| 10/10′/10” | 4.08, t (6.28) | 64.8 | -CH2- |
| 11/11′/11” | 1.86, s | 21.9 | -CH3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amsri, A.; Srichairatanakool, S.; Teerawutgulrag, A.; Youngchim, S.; Pongpom, M. Genetic Engineering of Talaromyces marneffei to Enhance Siderophore Production and Preliminary Testing for Medical Application Potential. J. Fungi 2022, 8, 1183. https://doi.org/10.3390/jof8111183
Amsri A, Srichairatanakool S, Teerawutgulrag A, Youngchim S, Pongpom M. Genetic Engineering of Talaromyces marneffei to Enhance Siderophore Production and Preliminary Testing for Medical Application Potential. Journal of Fungi. 2022; 8(11):1183. https://doi.org/10.3390/jof8111183
Chicago/Turabian StyleAmsri, Artid, Somdet Srichairatanakool, Aphiwat Teerawutgulrag, Sirida Youngchim, and Monsicha Pongpom. 2022. "Genetic Engineering of Talaromyces marneffei to Enhance Siderophore Production and Preliminary Testing for Medical Application Potential" Journal of Fungi 8, no. 11: 1183. https://doi.org/10.3390/jof8111183