
The Effect of Mitochondria on Ganoderma lucidum Growth and Bioactive Components Based on Transcriptomics
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
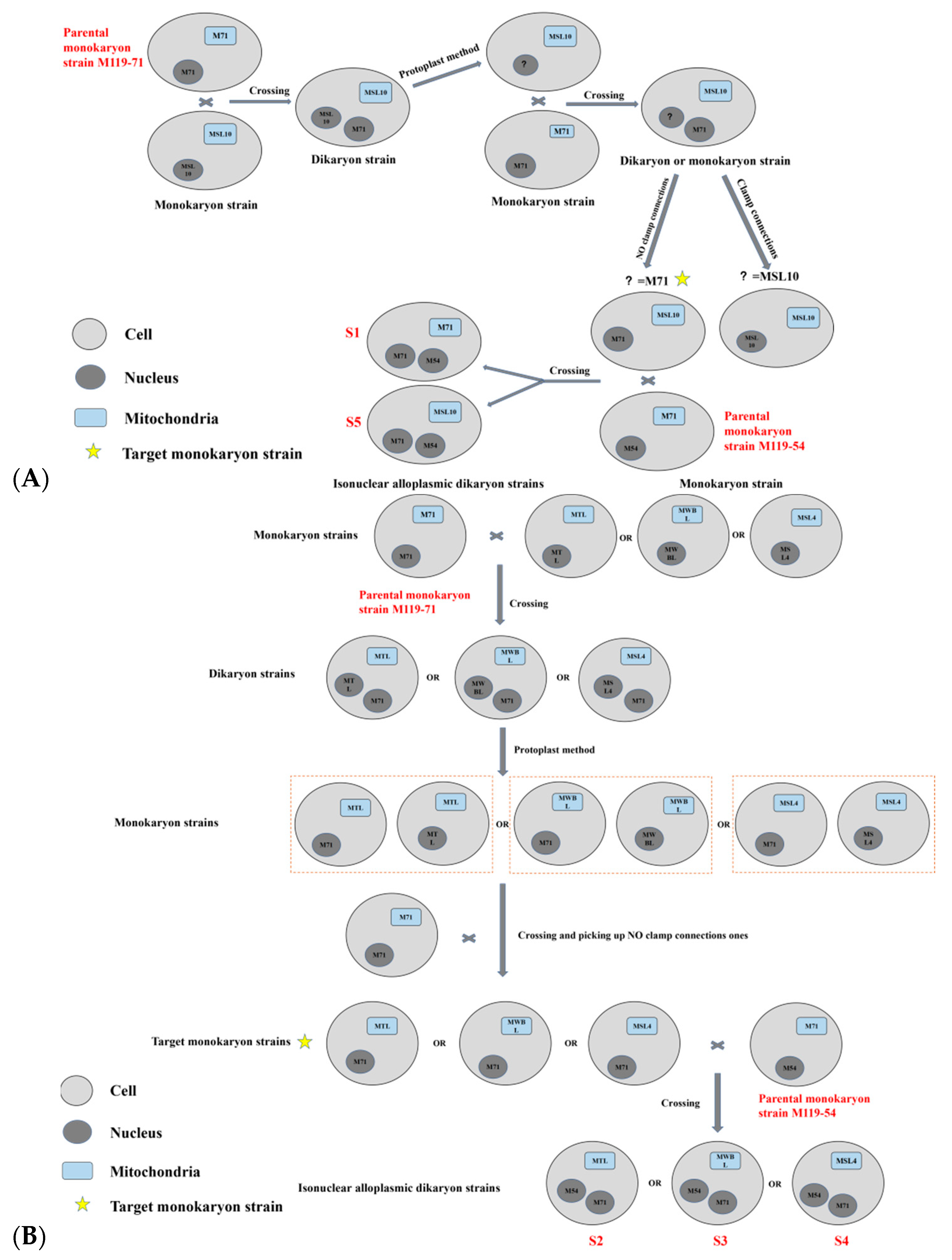
2.2. Isonuclear Alloplasmic Strains Obtention
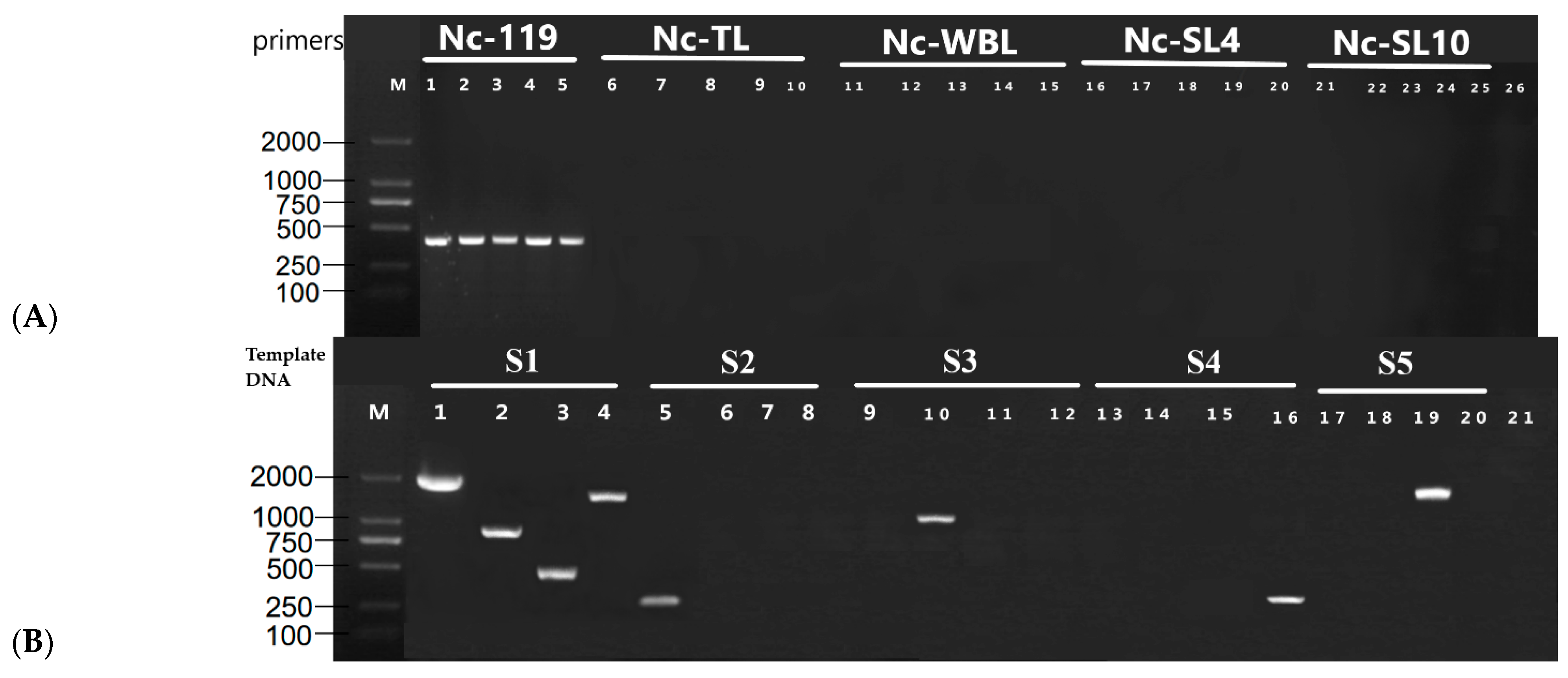
2.3. Isonuclear Alloplasmic Strains’ Nuclei DNA and mtDNA Proof
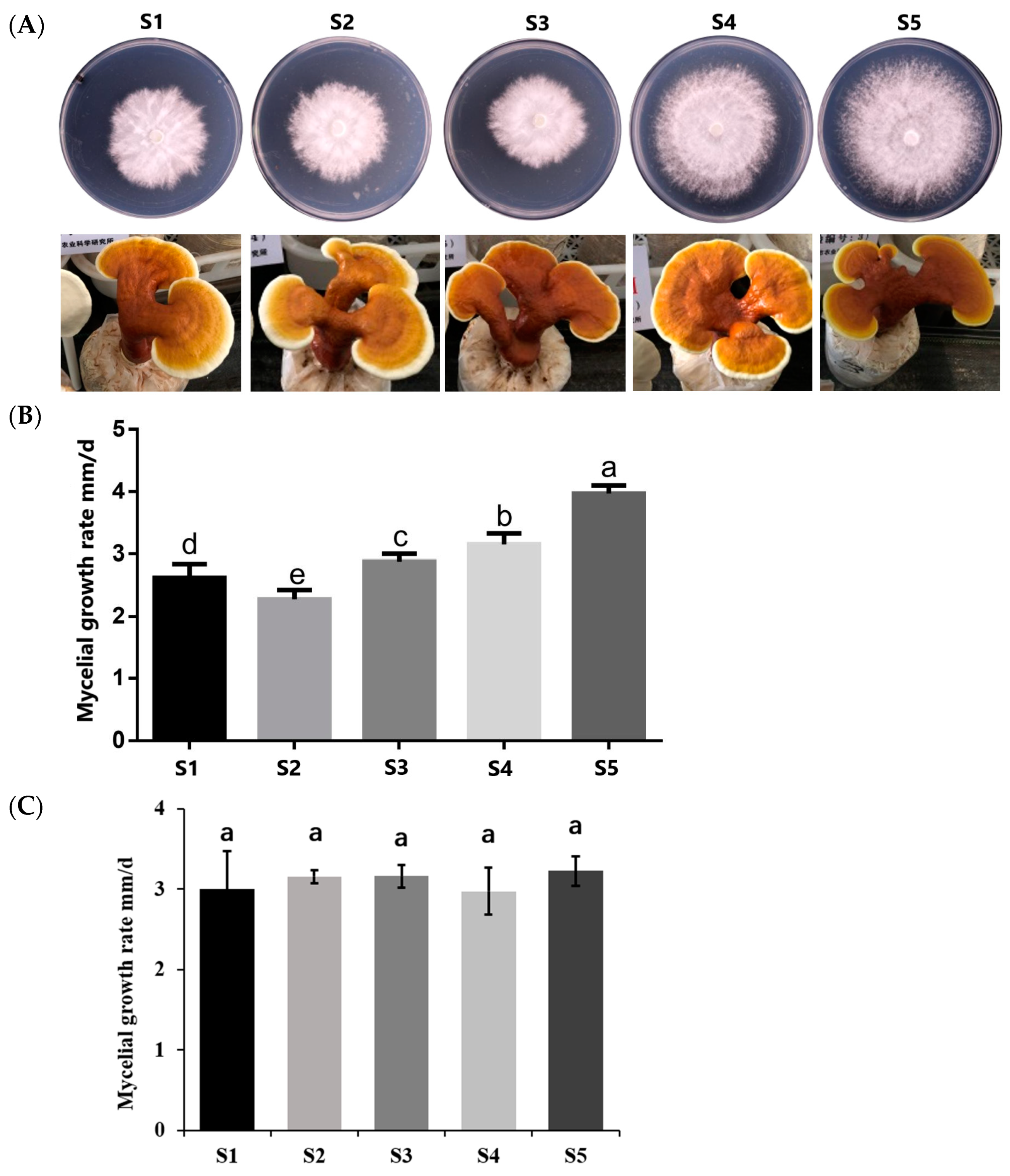
2.4. Biological Characteristics Observation
2.5. Total Polysaccharide and Triterpenoid Detection
2.6. Total RNA Isolation and Transcriptome Analysis
2.7. Statistical Analysis
3. Results
3.1. Isonuclear Alloplasmic Proof
3.2. Mycelia and Fruit Body Morphology Observation
3.3. Comparative Analysis of Yield and Polysaccharides and Triterpenoid Content
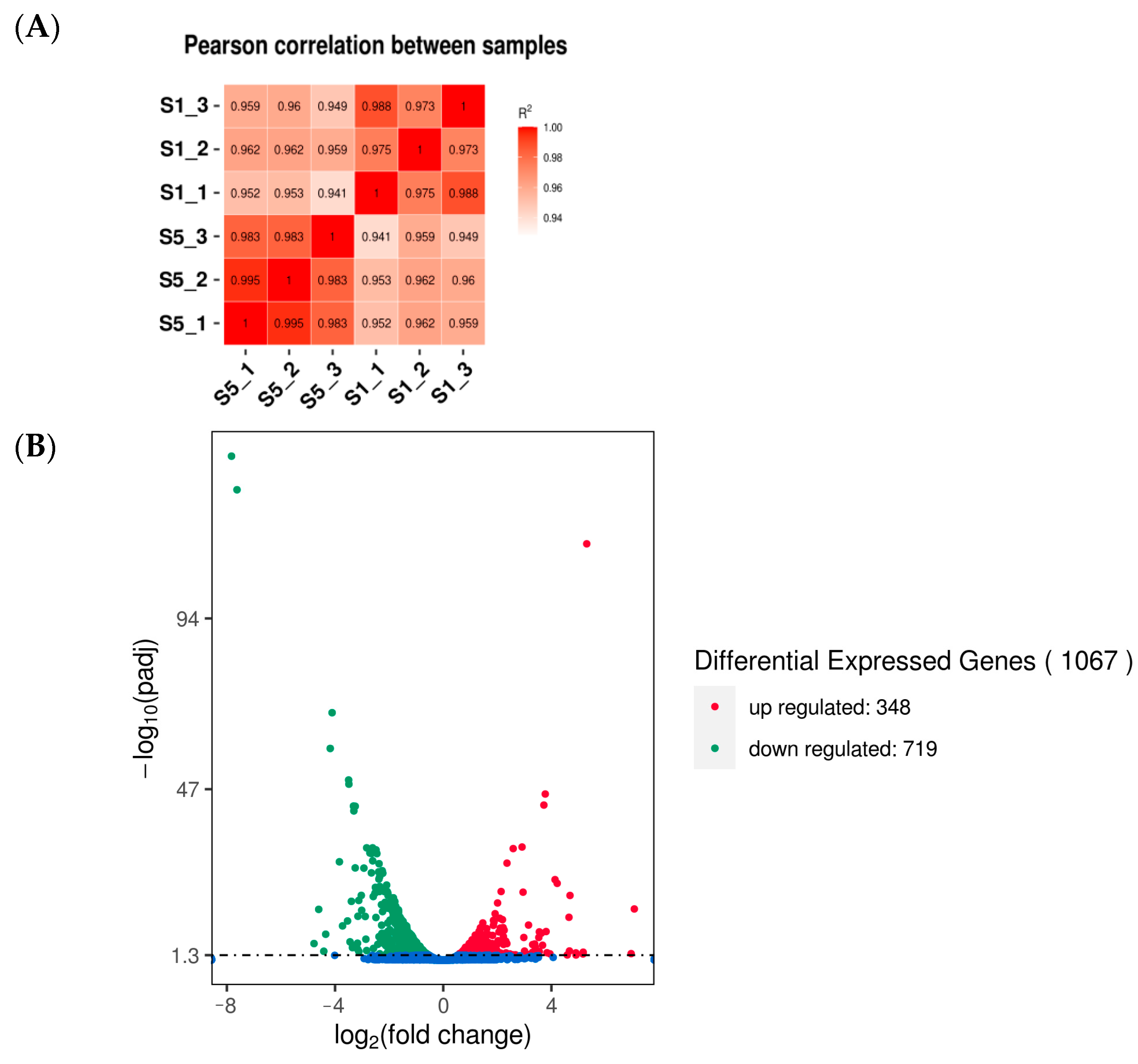
3.4. Assessment and Analysis of Transcriptome
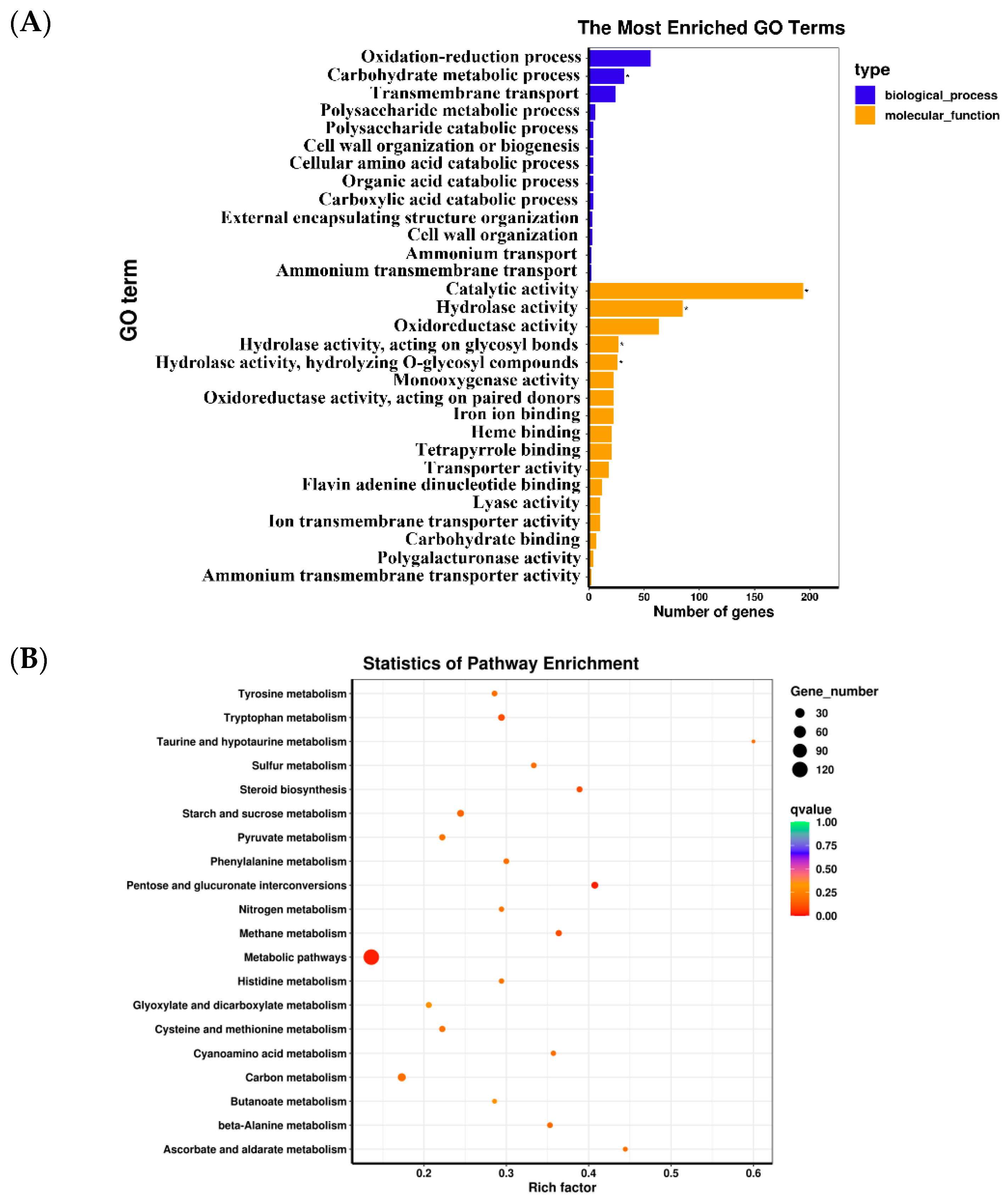
3.5. Enrichment Analysis of DEGs
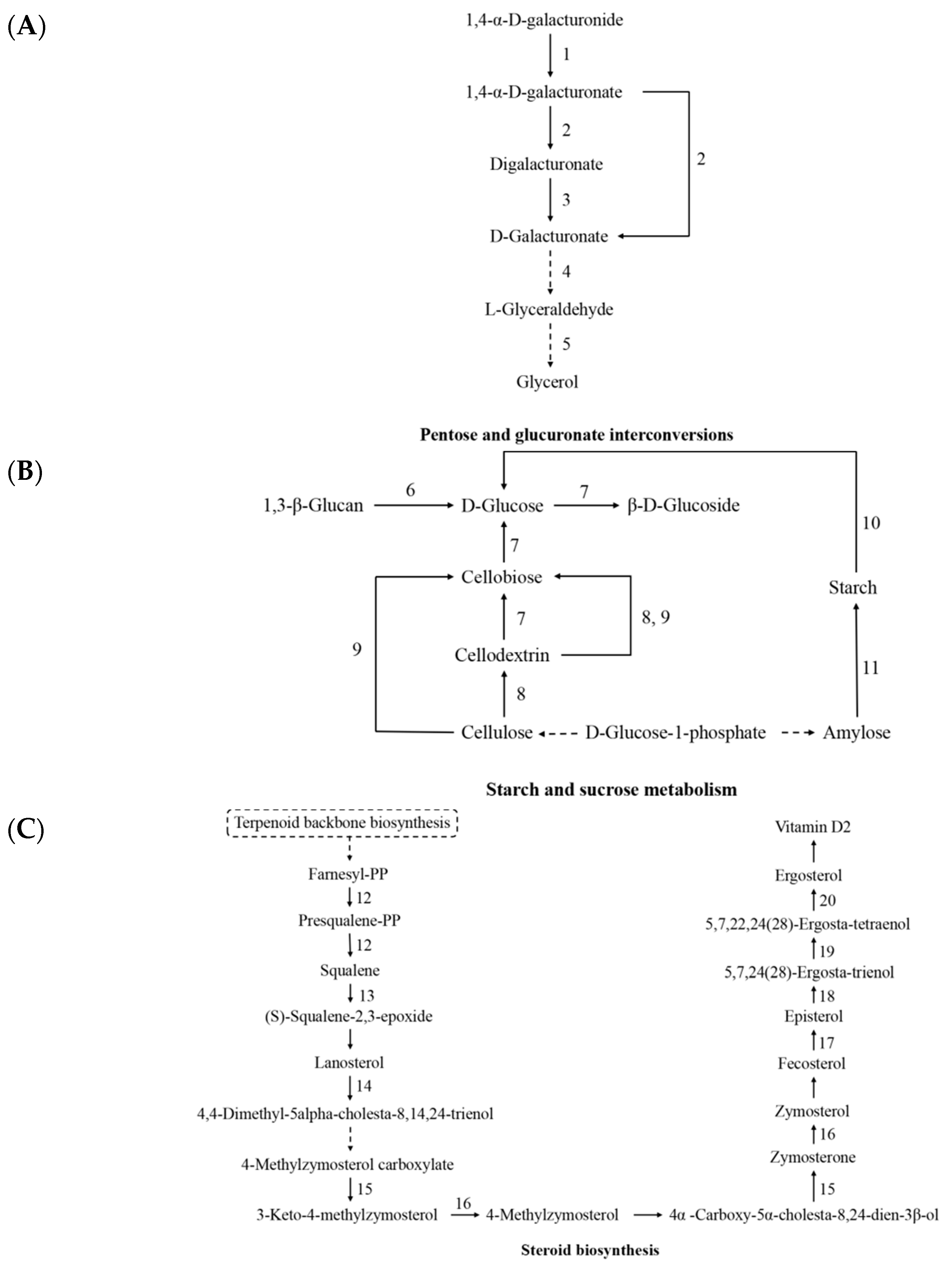
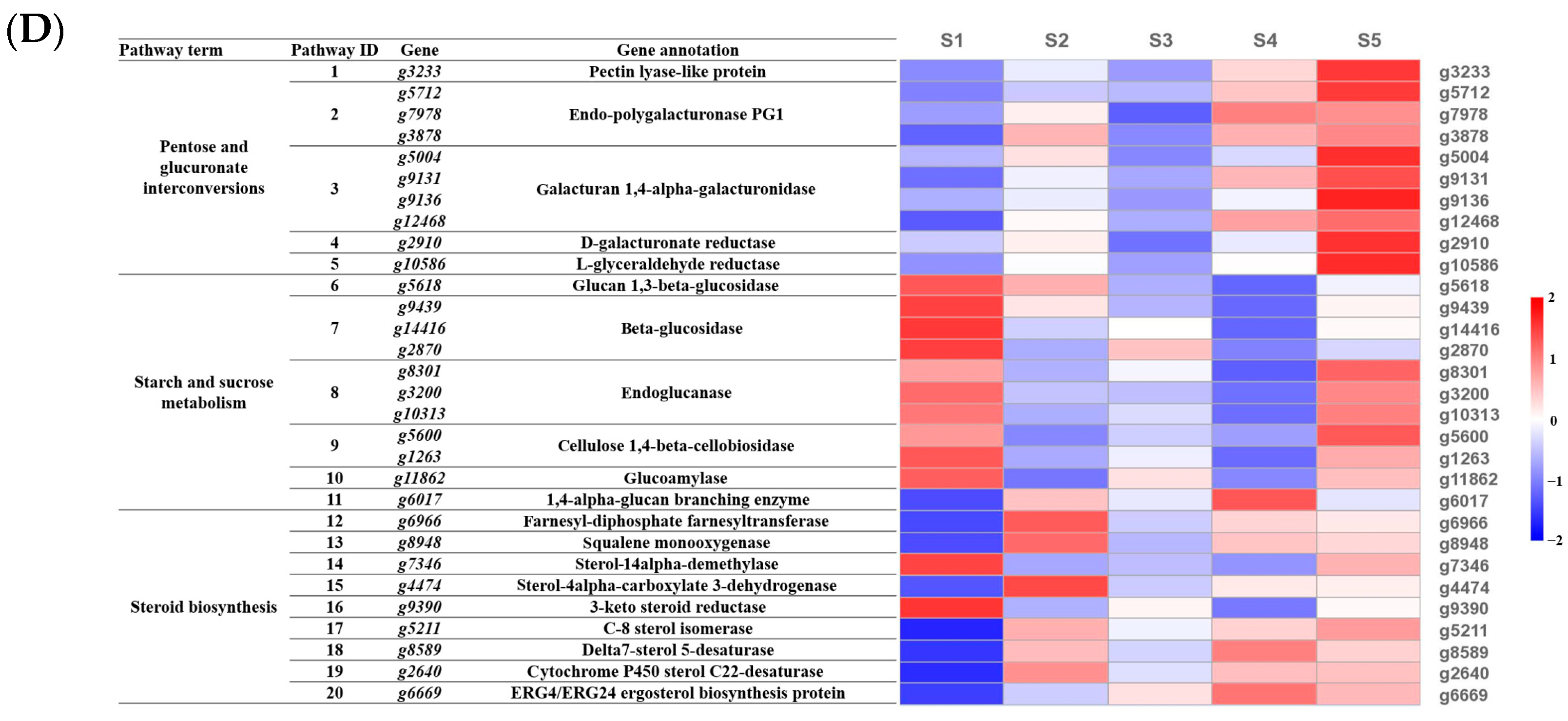
3.6. Analysis of Bioactive-Component-Related Biological Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Giordano, L.; Sillo, F.; Garbelotto, M.; Gonthier, P. Mitonuclear interactions may contribute to fitness of fungal hybrids. Sci. Rep. 2018, 8, 1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina, R.; Franco, M.E.E.; Bartel, L.C.; Alcántara, V.M.; Saparrat, M.C.N.; Balatti, P.A. Fungal mitogenomes: Relevant features to planning plant disease management. Front. Microbiol. 2020, 11, 978. [Google Scholar] [CrossRef] [PubMed]
- Kouvelis, V.N.; Hausner, G. Editorial: Mitochondrial genomes and mitochondrion related gene insights to fungal evolution. Front. Microbiol. 2022, 13, 897981. [Google Scholar] [CrossRef] [PubMed]
- Al-Reedy, R.M.; Malireddy, R.; Dillman, C.B.; Kennell, J.C. Comparative analysis of Fusarium mitochondrial genomes reveals a highly variable region that encodes an exceptionally large open reading frame. Fungal Genet. Biol. 2012, 49, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Freel, K.C.; Friedrich, A.; Schacherer, J. Mitochondrial genome evolution in yeasts: An all-encompassing view. FEMS Yeast Res. 2015, 15, fov023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Cai, Y.; Zhang, Q.; Shu, F.; Chen, L.; Ma, X.; Bian, Y.B. Subchromosome-scale nuclear and complete mitochondrial genome characteristics of Morchella crassipes. Int. J. Mol. Sci. 2020, 21, 483. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, E.; McGowan, J.; McCarthy, C.; Amini, A.; Grogan, H.; Fitzpatrick, D.A. Whole genome sequence of the commercially relevant mushroom strain Agaricus bisporus var. bisporus ARP23. G3 Genes Genomes Genet. 2019, 9, 3057–3066. [Google Scholar]
- Liu, X.R.; Wu, X.P.; Tan, H.; Xie, B.G.; Deng, Y.J. Large inverted repeats identified by intra-specific comparison of mitochondrial genomes provide insights into the evolution of Agrocybe aegerita. Comput. Struct. Biotechnol. J. 2020, 18, 2424–2437. [Google Scholar] [CrossRef]
- Bashir, K.M.I.; Rheu, K.M.; Kim, M.S.; Cho, M.G. The complete mitochondrial genome of an edible mushroom, Sparassis crispa. Mitochondrial DNA Part B Resour. 2020, 5, 862–863. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Wan, J.N.; Shang, J.J.; Feng, Z.; Jin, Y.; Li, H.; Guo, T.; Wu, Y.Y.; Bao, D.P.; Zhang, M.; et al. The complete mitochondrial genome of the edible mushroom Grifola frondosa. Mitochondrial DNA Part B Resour. 2022, 7, 286–288. [Google Scholar] [CrossRef]
- Li, J.Q.; Zhang, J.H.; Chen, H.M.; Chen, X.D.; Lan, J.; Liu, C. Complete mitochondrial genome of the medicinal mushroom Ganoderma lucidum. PLoS ONE 2013, 8, e72038. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Xiang, D.B.; Wan, Y.; Wu, Q.; Wu, X.Y.; Ma, C.R.; Song, Y.; Zhao, G.; Huang, W.L. The complete mitochondrial genomes of five important medicinal Ganoderma species: Features, evolution, and phylogeny. Int. J. Biol. Macromol. 2019, 139, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Steensels, J.; Gallone, B.; Verstrepen, K.J. Interspecific hybridization as a driver of fungal evolution and adaptation. Nat. Rev. Microbiol. 2021, 19, 485–500. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.K.; Thompson, L.J.; Madamanchi, N.; Ballinger, S.; Papaconstantinou, J.; Horaist, C.; Runge, M.S.; Patterson, C. Aging, oxidative responses, and proliferative capacity in cultured mouse aortic smooth muscle cells. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H2779–H2788. [Google Scholar] [CrossRef]
- Latorre-Pellicer, A.; Moreno-Loshuertos, R.; Lechuga-Vieco, A.V.; Sánchez-Cabo, F.; Torroja, C.; Acín-Pérez, R.; Calvo, E.; Aix, E.; González-Guerra, A.; Logan, A.; et al. Mitochondrial and nuclear DNA matching shapes metabolism and healthy ageing. Nature 2016, 535, 561–565. [Google Scholar] [CrossRef]
- Sun, X.M.; Wang, H.H.; Han, X.F.; Chen, S.W.; Zhu, S.; Dai, J. Fingerprint analysis of polysaccharides from different Ganoderma by HPLC combined with chemometrics methods. Carbohydr. Polym. 2014, 114, 432–439. [Google Scholar] [CrossRef]
- Qu, Z.W.; Zhou, S.Y.; Guan, S.X.; Gao, R.; Duan, Z.W.; Zhang, X.; Sun, W.Y.; Fan, W.L.; Chen, S.S.; Chen, L.J.; et al. Recombinant expression and bioactivity comparison of four typical fungal immunomodulatory proteins from three main Ganoderma Species. BMC Biotechnol. 2018, 18, 80. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.C.; Tang, X.C.; Hu, H.P.; Chen, D.L.; Xie, Y.Z.; Liang, X.W.; Li, X.M.; Xiao, C.; Huang, L.H.; Wu, Q.P. Genetic diversity and main functional composition of Lingzhi strains from main producing areas in China. AMB Express. 2021, 11, 119. [Google Scholar] [CrossRef]
- Xu, J.W.; Zhao, W.; Zhong, J.J. Biotechnological production and application of ganoderic acids. Appl. Microbiol. Biotechnol. 2010, 87, 457–466. [Google Scholar] [CrossRef]
- Gong, X.; Ji, M.Y.; Xu, J.P.; Zhang, C.H.; Li, M.H. Hypoglycemic effects of bioactive ingredients from medicine food homology and medicinal health food species used in China. Crit. Rev. Food Sci. Nutr. 2020, 60, 2303–2326. [Google Scholar] [CrossRef]
- He, X.R.; Fang, J.C.; Guo, Q.; Wang, M.; Li, Y.S.; Meng, Y.B.; Huang, L.H. Advances in antiviral polysaccharides derived from edible and medicinal plants and mushrooms. Carbohydr. Polym. 2020, 229, 115548. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Wang, S.X.; Wu, F.Y.; Wu, K.J.; Shi, R.P.; Qin, L.H.; Lu, C.F.; Wang, S.Q.; Wang, F.F.; Zhou, S.B. Effects and mechanism of Ganoderma lucidum polysaccharides in the treatment of diabetic nephropathy in streptozotocin-induced diabetic rats. BioMed Res. Int. 2022, 2022, 4314415. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.D.; Guan, X.Y.; Yong, T.Q.; Gao, X.; Xiao, C.; Xie, Y.Z.; Chen, D.L.; Hu, H.P.; Wu, Q.P. Structural characterization and hepatoprotective activity of an acidic polysaccharide from Ganoderma lucidum. Food Chem. X 2022, 13, 100204. [Google Scholar] [CrossRef]
- Ye, L.Y.; Deng, Y.J.; Mukhtar, I.; Meng, G.L.; Song, Y.J.; Cheng, B.; Hao, J.B.; Wu, X.P. Mitochondrial genome and diverse inheritance patterns in Pleurotus pulmonarius. J. Microbiol. 2020, 58, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.Z.; Ke, B.R.; Ye, L.Y.; Jin, S.S.; Jie, F.; Zhao, L.L.; Wu, X.P. Isolation and varietal characterization of Ganoderma resinaceum from areas of Ganoderma lucidum production in China. Sci. Hortic. 2017, 224, 109–114. [Google Scholar] [CrossRef]
- Ye, L.Y.; Liu, S.R.; Xie, F.; Zhao, L.L.; Wu, X.P. Enhanced production of polysaccharides and triterpenoids in Ganoderma lucidum fruit bodies on induction with signal transduction during the fruiting stage. PLoS ONE 2018, 13, e0196287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.B.; Ye, L.Y.; Meng, G.L.; Song, Y.J.; Fu, J.S.; Wu, X.P. The protective effect and crucial biological pathways analysis of Trametes lactinea mycelium polysaccharides on acute alcoholic liver injury in mice based on transcriptomics and metabolomics. Food Sci. Hum. Well. 2021, 10, 480–489. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.H.; Liu, L.L.; Guo, X.F.; Li, Y.J.; Hou, B.C.; Fan, Q.L.; Wang, K.X.; Luo, Y.D.; Zhong, J.J. Sucrose fed-batch strategy enhanced biomass, polysaccharide, and ganoderic acids production in fermentation of Ganoderma lucidum 5.26. Bioprocess Biosyst. Eng. 2016, 39, 37–44. [Google Scholar] [CrossRef]
- Tang, Y.J.; Zhu, L.W. Improvement of ganoderic acid and Ganoderma polysaccharide biosynthesis by Ganoderma lucidum fermentation under the inducement of Cu2+. Biotechnol. Prog. 2010, 26, 417–423. [Google Scholar] [PubMed]
- Sun, B.; You, H.; Xu, J.W. Enhancement of ganoderic acid production by promoting sporulation in a liquid static culture of Ganoderma species. J. Biotechnol. 2021, 328, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.H.; Zhang, Q.Q.; Zhang, Q.F.; He, H.Q.; Chen, Z.; Zhao, Y.; Wei, D.; Kong, M.G.; Huang, Q. Improved production of polysaccharides in Ganoderma lingzhi mycelia by plasma mutagenesis and rapid screening of mutated strains through infrared spectroscopy. PLoS ONE 2018, 13, e0204266. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.K.; Zhu, J.; Sun, Z.H.; Zhang, G.; Liu, R.; Zhang, T.J.; Wang, S.L.; Ren, A.; Zhao, M.W. Alternative oxidase impacts ganoderic acid biosynthesis by regulating intracellular ROS levels in Ganoderma lucidum. Microbiology 2017, 163, 1466–1476. [Google Scholar] [CrossRef] [PubMed]
- Mezhnina, V.; Ebeigbe, O.P.; Poe, A.; Kondratov, R.V. Circadian control of mitochondria in reactive oxygen species homeostasis. Antioxid. Redox Signal. 2022, 37, 647–663. [Google Scholar] [CrossRef]
- Suski, J.; Lebiedzinska, M.; Bonora, M.; Pinton, P.; Duszynski, J.; Wieckowski, M.R. Relation between mitochondrial membrane potential and ROS formation. Methods Mol. Biol. 2018, 1782, 357–381. [Google Scholar]
- Miyazaki, T.; Nishijima, M. Studies on fungal polysaccharides. XXVII. Structural examination of a water-soluble, antitumor polysaccharide of Ganoderma lucidum. Chem. Pharm. Bull. 1981, 29, 3611–3616. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Zhang, J.; Kan, Q.; Song, M.; Hou, T.; An, S.; Lin, H.; Chen, H.; Hu, L.; Xiao, J.; et al. Extraction, structural characterization, and immunomodulatory activity of a high molecular weight polysaccharide from Ganoderma lucidum. Front. Nutr. 2022, 9, 846080. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Mushroom Yield g/bag | Basidiospores Yield g/bag | Polysaccharide Content (mg/g) | Triterpenoid Content (mg/g) |
|---|---|---|---|---|
| S1 | 70.67 ± 3.42 a | 19.67 ± 1.34 a | 31.16 ± 0.97 a | 19.34 ± 0.38 d |
| S2 | 70.67 ± 5.41 a | 15.94 ± 1.86 b | 29.19 ± 1.57 b | 20.04 ± 0.19 bc |
| S3 | 70.83 ± 3.75 a | 16.89 ± 1.24 ab | 25.93 ± 0.85 d | 20.87 ± 0.50 b |
| S4 | 73.17 ± 4.91 a | 15.33 ± 2.17 b | 26.58 ± 1.05 cd | 22.51 ± 0.76 a |
| S5 | 66.67 ± 4.13 a | 5.55 ± 0.99 c | 28.00 ± 0.43 bc | 19.94 ± 0.26 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, L.; He, X.; Su, C.; Feng, H.; Meng, G.; Chen, B.; Wu, X. The Effect of Mitochondria on Ganoderma lucidum Growth and Bioactive Components Based on Transcriptomics. J. Fungi 2022, 8, 1182. https://doi.org/10.3390/jof8111182
Ye L, He X, Su C, Feng H, Meng G, Chen B, Wu X. The Effect of Mitochondria on Ganoderma lucidum Growth and Bioactive Components Based on Transcriptomics. Journal of Fungi. 2022; 8(11):1182. https://doi.org/10.3390/jof8111182
Chicago/Turabian StyleYe, Liyun, Xiaofang He, Congbao Su, Haiying Feng, Guoliang Meng, Bingzhi Chen, and Xiaoping Wu. 2022. "The Effect of Mitochondria on Ganoderma lucidum Growth and Bioactive Components Based on Transcriptomics" Journal of Fungi 8, no. 11: 1182. https://doi.org/10.3390/jof8111182