
Reassessment of Dyfrolomyces and Four New Species of Melomastia from Olive (Olea europaea) in Sichuan Province, China

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials & Methods
2.1. Isolation and Morphology
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Strain Numbers | GenBank Accession Numbers | References | ||
|---|---|---|---|---|---|
| LSU | SSU | tef1-α | |||
| Acrospermum adeanum | M 133 | EU940104 | EU940031 | N/A | [40] |
| A. compressum | M 151 | EU940084 | EU940012 | N/A | [40] |
| A. graminum | M 152 | EU940085 | EU940013 | N/A | [40] |
| Anisomeridium phaeospermum | MPN539 | JN887394 | JN887374 | JN887418 | [41] |
| A. ubianum | MPN94 | N/A | JN887379 | JN887421 | [41] |
| Melomastia chromolaenae | MFLUCC 17–1434 * | KY111905 | MT214413 | MT235800 | [42] |
| M. clematidis | MFLUCC 17–2092 * | MT214607 | MT226718 | MT394663 | [19] |
| M. distoseptata | NFCCI: 4377 * | MH971236 | N/A | N/A | [15] |
| M. fulvicomae | MFLUCC 17–2083 * | MT214608 | MT226719 | N/A | [19] |
| M. fusispora | CGMCC 3.20618 * | OK623464 | OK623494 | OL335189 | This study |
| M. fusispora | UESTCC 21.0001 | OK623465 | OK623495 | OL335190 | This study |
| M. italica | MFLUCC 15–0160 * | MG029458 | MG029459 | N/A | [18] |
| M. maolanensis | GZCC 16–0102 * | KY111905 | KY111906 | KY814762 | [28] |
| M. neothailandica | MFLU 17–2589 * | NG068294 | N/A | N/A | [43] |
| M. oleae | CGMCC 3.20619 * | OK623466 | OK623496 | OL335191 | This study |
| M. oleae | UESTCC 21.0003 | OK623467 | OK623497 | OL335192 | This study |
| M. oleae | UESTCC 21.0005 | OK623468 | OK623498 | OL335193 | This study |
| M. oleae | UESTCC 21.0006 | N/A | OK623499 | OL335194 | This study |
| M. phetchaburiensis | MFLUCC 15-0951 * | MF615402 | MF615403 | N/A | [44] |
| M. rhizophorae | BCC 15481 | N/A | KF160009 | N/A | [20] |
| M. rhizophorae | JK 5439 A | GU479799 | GU479766 | GU479860 | [20] |
| M. sichuanensis | CGMCC 3.20620 * | OK623469 | OK623500 | OL335195 | This study |
| M. sichuanensis | UESTCC 21.0008 | OK623470 | OK623501 | OL335196 | This study |
| M. sinensis | MFLUCC 17–1344 * | MG836699 | MG836700 | N/A | [45] |
| M. thailandica | MFLUCC 15–0945 * | KX611366 | KX611367 | N/A | [46] |
| M. thamplaensis | MFLUCC 15–0635 * | KX925435 | KX925436 | KY814763 | [28] |
| M. tiomanensis | MFLUCC 13–0440 * | KC692156 | KC692155 | KC692157 | [20] |
| M. winteri | CGMCC 3.20621 * | OK623471 | OK623502 | OL335197 | This study |
| Muyocopron castanopsis | MFLUCC 14–1108 * | KU726965 | KU726968 | MT136753 | [47] |
| M. dipterocarpi | MFLU 17–2608 | KU726966 | KU726969 | MT136754 | [47] |
| M. garethjonesii | MFLUCC 16–2664 * | KY070274 | KY070275 | N/A | [48] |
| M. heveae | MFLUCC 17–0066 * | MH986832 | MH986828 | N/A | [29] |
| M. lithocarpi | MFLUCC 14–1106 * | KU726967 | KU726970 | MT136755 | [47] |
| Palawania thailandensis | MFLICC 14–1121 * | KY086494 | N/A | N/A | [49] |
| P. thailandensis | MFLU 16–1873 | KY086493 | KY086495 | N/A | [49] |
| Stigmatodiscus enigmaticus | CBS 132036 * | KU234108 | KU234130 | N/A | [50] |
| S. labiatus | CBS 144700 * | MH756065 | MH756065 | MH756083 | [51] |
| S. oculatus | CBS 144701 * | MH756069 | N/A | MH756086 | [51] |
| S. pruni | CBS 142598 * | KX611110 | KX611110 | KX611111 | [52] |
| Superstratomyces albomucosus | CBS 140270 * | KX950439 | KX950467 | KX950471 | [53] |
| S. atroviridis | CBS 140272 * | NG058271 | NG063075 | LR812724 | [53] |
| S. flavomucosus | CBS 353.84 * | KX950438 | KX950462 | KX950470 | [53] |
3. Results
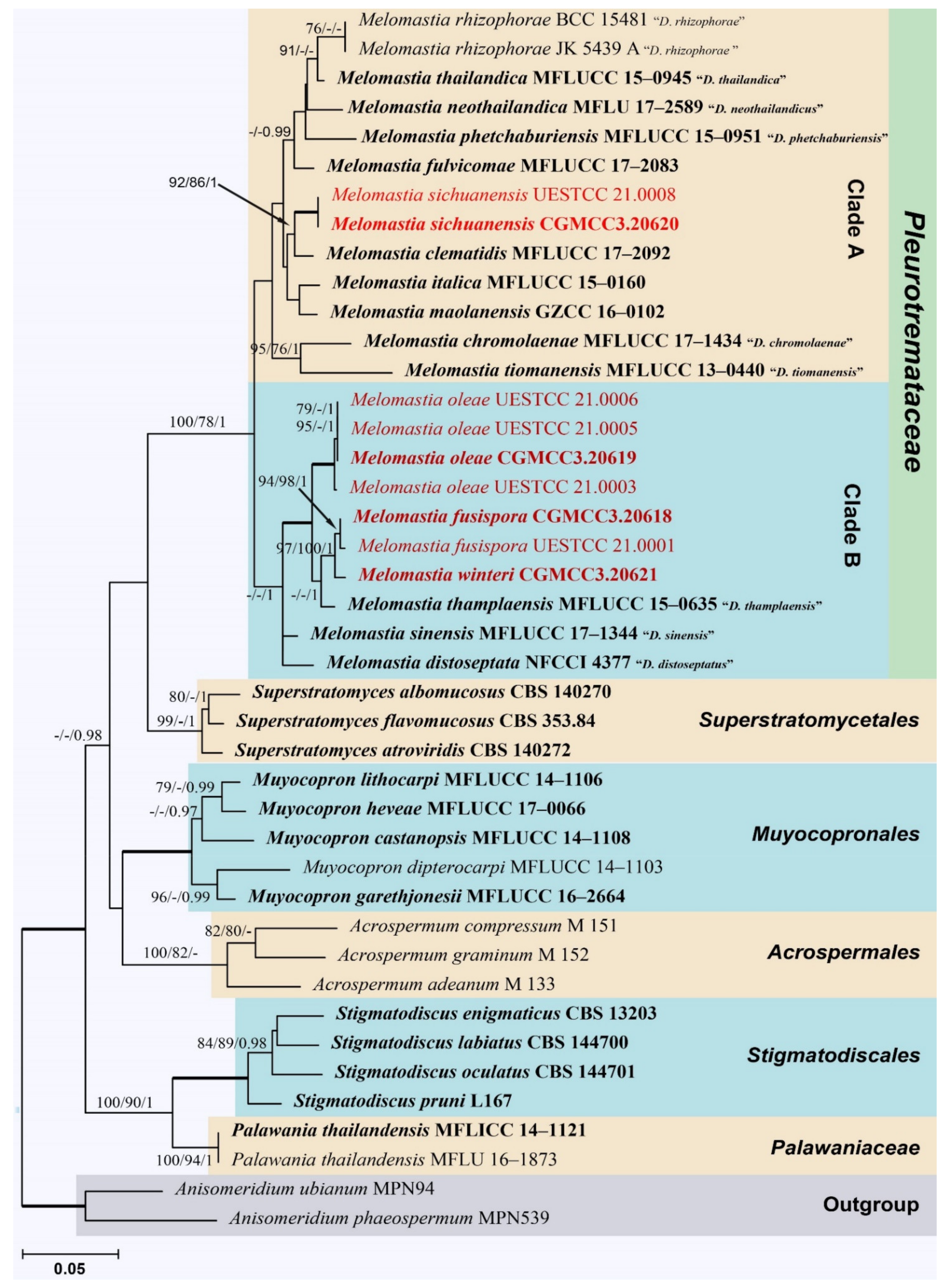
3.1. Phylogenetic Analyses
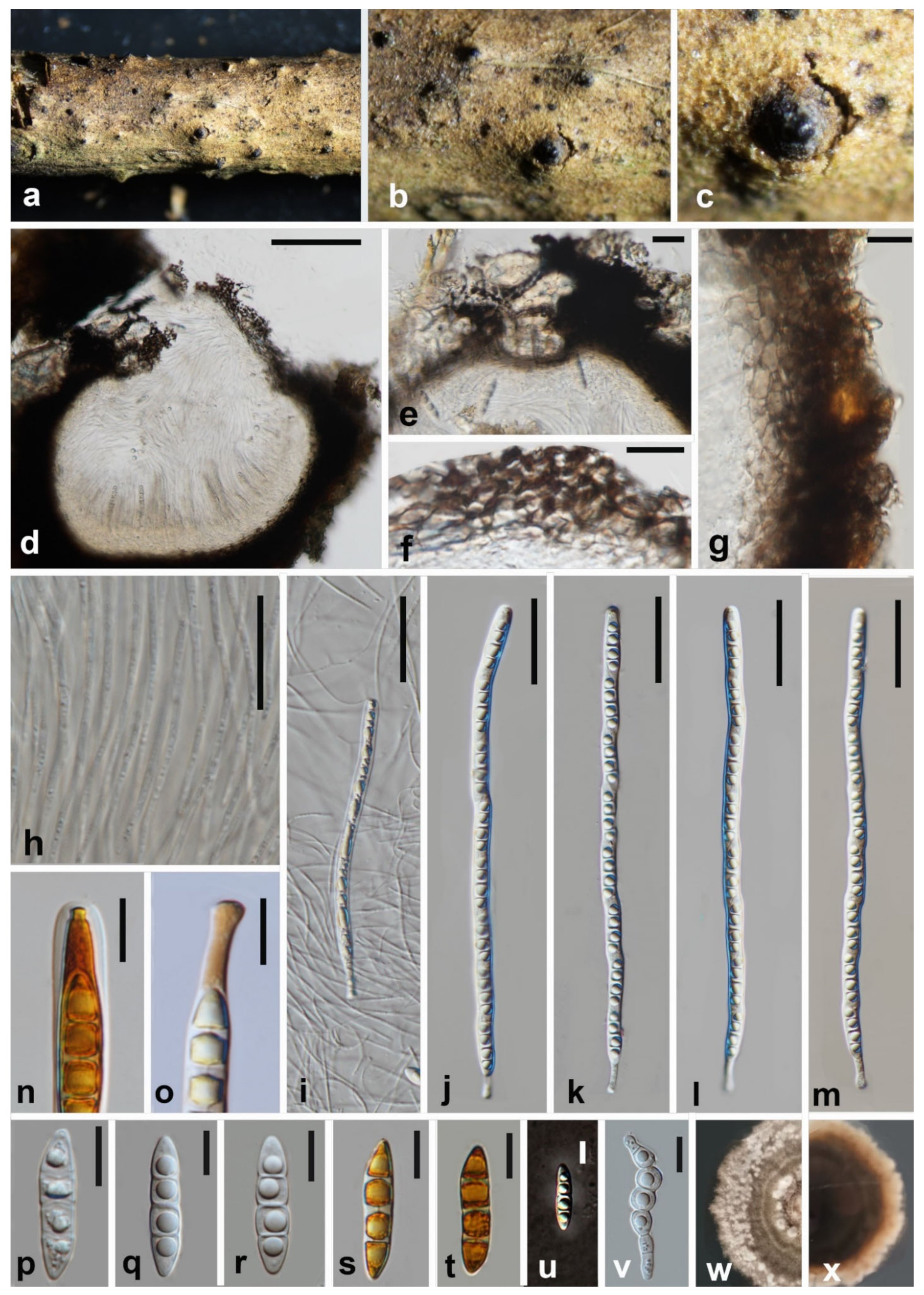
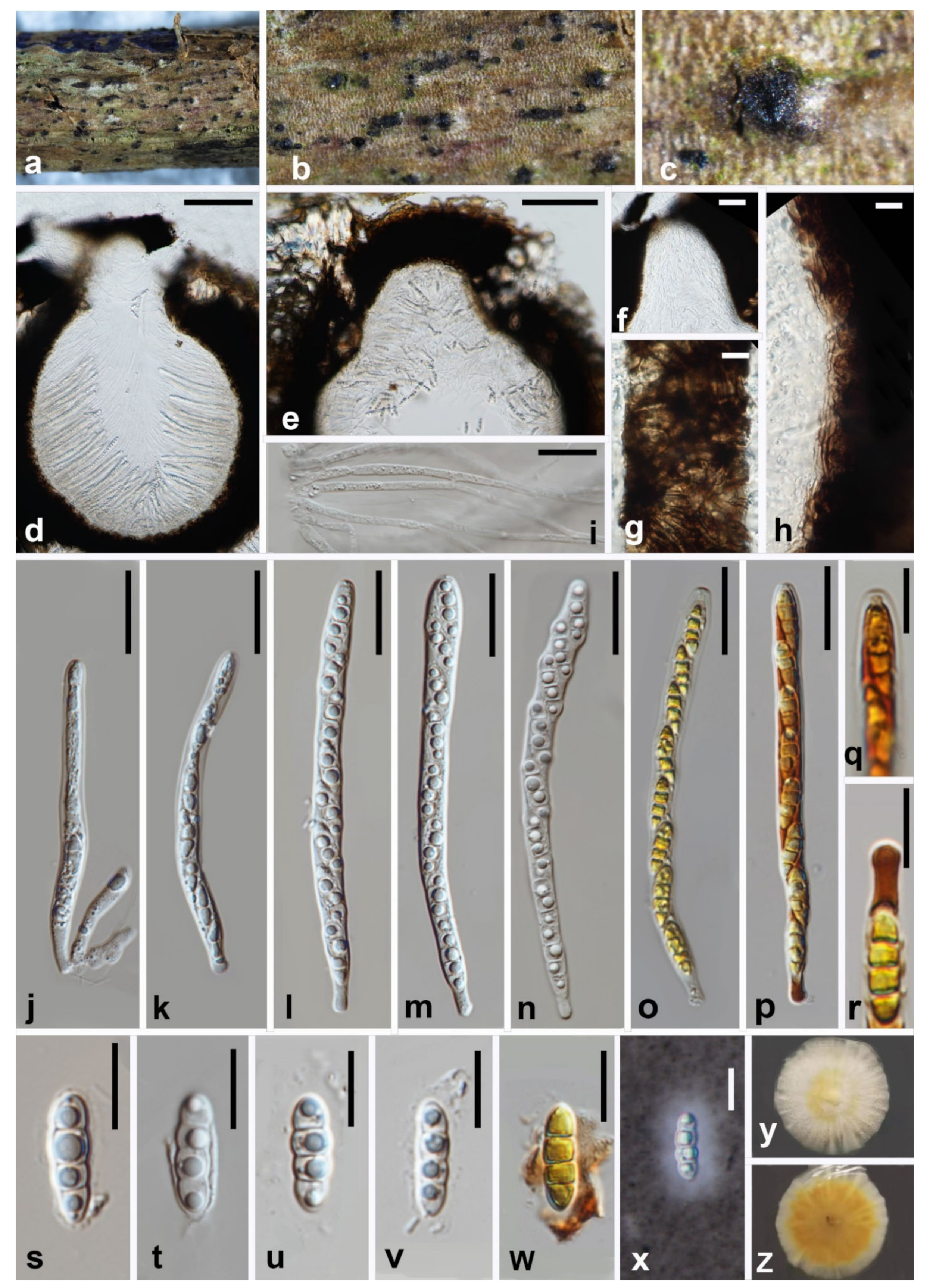
3.2. Taxonomy
3.2.1. Melomastia




3.2.2. New Combination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Watson, W. The classification of lichens. New Phytol. 1929, 28, 85–116. [Google Scholar] [CrossRef]
- Barr, M.E. Notes on the Amphisphaeriaceae and related families. Mycotaxon 1994, 51, 191–224. [Google Scholar]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of the Fungi, 10th ed.; CABI: Oxford, UK, 2008. [Google Scholar]
- Senanayake, I.; Al-Sadi, A.; Bhat, J.; Camporesi, E.; Dissanayake, A.; Lumyong, S.; Maharachchikumbura, S.; Hyde, K. Phomatosporales ord. nov. and Phomatosporaceae fam. nov., to accommodate Lanspora, Phomatospora and Tenuimurus, gen. nov. Mycosphere 2016, 7, 628–641. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; David, J.C.; Stalpers, J.A. Ainsworth and Bisby’s Dictionary of the Fungi, 8th ed.; CABI: London, UK, 2001. [Google Scholar]
- Lumbsch, H.T.; Huhndorf, S.M. Myconet Volume 14. Part One. Outline of Ascomycota—2009. Part Two. Notes on Ascomycete Systematics. Nos. 4751–5113. Fieldiana Life Earth Sci. 2010, 2010, 1–64. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Kirk, P.M.; Sutton, B.C.; Pegler, D.N. Dictionary of the Fungi, 8th ed.; CABI: Oxford, UK, 1995. [Google Scholar]
- Mathiassen, G. Some Corticolous and Lignicolous Pyrenomycetes s. lat. (Ascomycetes) on Salix in Troms, N Norway; Botanical Garden and Museum, University of Oslo: Oslo, Norway, 1989. [Google Scholar]
- Hyde, K.D. The genus Saccardoella from intertidal mangrove wood. Mycologia 1992, 84, 803–810. [Google Scholar] [CrossRef]
- Barr, M. The genus Dothidotthia (Botryosphaeriaceae) in North America. Mycotaxon 1989, 34, 517–526. [Google Scholar]
- Aptroot, A. A monograph of the Pyrenulaceae (excluding Anthracothecium and Pyrenula) and the Requinellaceae, with notes on the Pleomassariaceae, the Trypetheliaceae and Mycomicrothelia (lichenized and non-lichenized Ascomycetes). Bibl. Lichenol. 1991, 44, 1–178. [Google Scholar]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Huang, S.K.; Abdel-Wahab, M.A.; Daranagama, D.A.; Dayarathne, M.; D’souza, M.J.; Goonasekara, I.D.; et al. Towards a natural classification and backbone tree for Sordariomycetes. Fungal Divers. 2015, 72, 199–301. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Bhat, J.D.; Dayarathne, M.C.; Huang, S.K.; Norphanphoun, C.; Senanayake, I.C.; Perera, R.H.; et al. Families of Sordariomycetes. Fungal Divers. 2016, 79, 1–317. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Rajeshkumar, K.C.; Hawksworth, D.L.; Madrid, H.; Kirk, P.M.; Braun, U.; Singh, R.V.; Crous, P.W.; Kukwa, M.; et al. Notes for genera: Ascomycota. Fungal Divers. 2017, 86, 1–594. [Google Scholar] [CrossRef] [Green Version]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Bhat, D.J.; Liu, N.G.; et al. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Saccardo, P. Conspectus generum pyrenomycetum italicorum additis speciebus fungorum Venetorum novis vel criticis, systemate carpologico dispositorum. Atti Soc. Veneziana-Trent.-Istriana Sci. Nat. 1875, 4, 77–100. [Google Scholar]
- Kang, J.C.; Hyde, K.D.; Kong, R.Y.C. Studies on Amphisphaeriales: The genera excluded from the Amphisphaeriaceae, Cainiaceae and Clypeosphaeriaceae. Fungal Divers. 1999, 2, 135–151. [Google Scholar]
- Norphanphoun, C.; Jeewon, R.; Mckenzie, E.H.C.; Wen, T.C.; Camporesi, E.; Hyde, K.D. Taxonomic Position of Melomastia italica sp. nov. and Phylogenetic Reappraisal of Dyfrolomycetales. Cryptogam. Mycol. 2017, 38, 507–525. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; McKenzie, E.H.C.; Phillips, A.J.L.; Gareth Jones, E.B.; Jayarama Bhat, D.; Stadler, M.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Pang, K.L.; Hyde, K.D.; Alias, S.A.; Suetrong, S.; Guo, S.Y.; Idid, R.; Jones, E.G. Dyfrolomycetaceae, a new family in the Dothideomycetes, Ascomycota. Cryptogam. Mycol. 2013, 34, 223–232. [Google Scholar] [CrossRef]
- Hyde, K.D.; Jones, E.B.G.; Liu, J.K.; Ariyawansa, H.; Boehm, E.; Boonmee, S.; Braun, U.; Chomnunti, P.; Crous, P.W.; Dai, D.Q.; et al. Families of Dothideomycetes. Fungal Divers. 2013, 63, 1–313. [Google Scholar] [CrossRef]
- Samarakoon, M.C.; Maharachchikumbura, S.S.N.; Liu, J.J.; Hyde, K.D.; Promputtha, I.; Stadler, M. Molecular Phylogeny and Morphology of Amphisphaeria (= Lepteutypa) (Amphisphaeriaceae). J. Fungi 2020, 6, 174. [Google Scholar] [CrossRef]
- Liu, J.K.; Chomnunti, P.; Cai, L.; Phookamsak, R. Phylogeny and morphology of Neodeightonia palmicola sp. nov. from palms. Sydowia 2010, 62, 261–276. [Google Scholar]
- Chomnunti, P.; Hongsanan, S.; Aguirre-Hudson, B.; Tian, Q.; Peršoh, D.; Dhami, M.K.; Alias, A.S.; Xu, J.; Liu, X.; Stadler, M.; et al. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990. [Google Scholar]
- Rehner, S. Primers for Elongation Factor 1-a (EF1-a). Available online: http://ocid.nacse.org/research/deephyphae/EF1primer.pdf (accessed on 25 November 2021).
- Zhang, J.F.; Liu, J.K.; Hyde, K.D.; Chen, Y.Y.; Liu, X.Z.; Liu, Z.Y. Two new species of Dyfrolomyces (Dyfrolomycetaceae, Dothideomycetes) from karst landforms. Phytotaxa 2017, 313, 267–277. [Google Scholar] [CrossRef]
- Senwanna, C.; Hongsanan, S.; Phookamsak, R.; Tibpromma, S.; Cheewangkoon, R.; Hyde, K.D. Muyocopron heveae sp. nov. and M. dipterocarpi appears to have host-jumped to rubber. Mycol. Prog. 2019, 18, 741–752. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A. MrModeltest v2, Program Distributed by the Author, Evolutionary Biology Centre; Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Dissanayake, A.J.; Bhunjun, C.S.; Maharachchikumbura, S.S.N.; Liu, J.K. Applied aspects of methods to infer phylogenetic relationships amongst fungi. Mycosphere 2020, 11, 2653–2677. [Google Scholar] [CrossRef]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (And Other Methods). Version 4; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- Rambaut, A. FigTree v1. 4.0. A Graphical Viewer of Phylogenetic Trees. 2012. Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 1 December 2021).
- Stenroosa, S.; Laukka, T.; Huhtinen, S.; Döbbeler, P.; Myllys, L.; Syrjänen, K.; Hyvönen, J. Multiple origins of symbioses between ascomycetes and bryophytes suggested by a five-gene phylogeny. Cladistics 2010, 26, 281–300. [Google Scholar] [CrossRef] [PubMed]
- Nelsen, M.P.; Lücking, R.; Mbatchou, J.S.; Andrew, C.J.; Spielmann, A.A.; Lumbsch, H.T. New insights into relationships of lichen-forming Dothideomycetes. Fungal Divers. 2011, 51, 155–162. [Google Scholar] [CrossRef]
- Mapook, A.; Hyde, K.D.; McKenzie, E.H.C.; Jones, E.B.G.; Bhat, D.J.; Jeewon, R.; Stadler, M.; Samarakoon, M.C.; Malaithong, M.; Tanunchai, B.; et al. Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 2020, 101, 1–175. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Devadatha, B.; Sarma, V.V.; Khongphinitbunjong, K.; Chomnunti, P.; Hyde, K.D. Morpho-molecular characterization of microfungi associated with marine based habitats. Mycosphere 2020, 11, 1–188. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Abreu, V.P.; Bazzicalupo, A.; Thilini Chethana, K.W.; Clericuzio, M.; Dayarathne, M.C.; Dissanayake, A.J.; Ekanayaka, A.H.; He, M.Q.; et al. Fungal diversity notes 603–708: Taxonomic and phylogenetic notes on genera and species. Fungal Divers. 2017, 87, 1–235. [Google Scholar] [CrossRef]
- Hyde, K.D.; Chaiwan, N.; Norphanphoun, C.; Boonmee, S.; Camporesi, E.; Chethana, K.W.T.; Dayarathne, M.C.; de Silva, N.I.; Dissanayake, A.J.; Ekanayaka, A.H.; et al. Mycosphere notes 169–224. Mycosphere 2018, 9, 271–430. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Mapook, A.; Hyde, K.D.; Dai, D.Q.; Li, J.; Jones, E.B.G.; Bahkali, A.H.; Boonmee, S. Muyocopronales, ord. nov., (Dothideomycetes, Ascomycota) and a reappraisal of Muyocopron species from northern Thailand. Phytotaxa 2016, 265, 225–237. [Google Scholar] [CrossRef]
- Tibpromma, S.; McKenzie, E.H.C.; Karunarathna, S.C.; Mortimer, P.E.; Xu, J.; Hyde, K.D.; Hu, D.M. Muyocopron garethjonesii sp. nov. (Muyocopronales, Dothideomycetes) on Pandanus sp. Mycosphere 2016, 7, 1480–1489. [Google Scholar] [CrossRef]
- Mapook, A.; Hyde, K.D.; Hongsanan, S.; Phukhamsakda, C.; Li, J.F.; Boonmee, S. Palawaniaceae fam. nov., a new family (Dothideomycetes, Ascomycota) to accommodate Palawania species and their evolutionary time estimates. Mycosphere 2016, 7, 1732–1745. [Google Scholar] [CrossRef]
- Voglmayr, H.; Gardiennet, A.; Jaklitsch, W.M. Asterodiscus and Stigmatodiscus, two new apothecial dothideomycete genera and the new order Stigmatodiscales. Fungal Divers. 2016, 80, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Voglmayr, H.; Amengual, A.P. Three new species of Stigmatodiscus from Mallorca (Spain). Mycol. Prog. 2018, 17, 1189–1201. [Google Scholar] [CrossRef]
- Voglmayr, H.; Fournier, J.; Jaklitsch, W.M. Stigmatodiscus pruni, a new dothideomycete with hysteriform ascomata. Sydowia 2017, 69, 29–35. [Google Scholar] [CrossRef]
- Van Nieuwenhuijzen, E.J.; Miadlikowska, J.M.; Houbraken, J.A.; Adan, O.C.; Lutzoni, F.M.; Samson, R.A. Wood staining fungi revealed taxonomic novelties in Pezizomycotina: New order Superstratomycetales and new species Cyanodermella oleoligni. Stud. Mycol. 2016, 85, 107–124. [Google Scholar] [CrossRef]
- Schröter, J. Die Pilze Schlesiens. J. U. Kern. 1894, 2, 1889–1908. [Google Scholar]
- Chen, C.Y.; Hsieh, W.H. Two new species and some new records of ascomycetes from Taiwan. Bot. Bull. Acad. Sin. 1996, 37, 219–227. [Google Scholar]
- Thambugala, K.M.; Hyde, K.D.; Tanaka, K.; Tian, Q.; Wanasinghe, D.N.; Ariyawansa, H.A.; Jayasiri, S.C.; Boonmee, S.; Camporesi, E.; Hashimoto, A.; et al. Towards a natural classification and backbone tree for Lophiostomataceae, Floricolaceae, and Amorosiaceae fam. nov. Fungal Divers. 2015, 74, 199–266. [Google Scholar] [CrossRef]
- Hashimoto, A.; Hirayama, K.; Takahashi, H.; Matsumura, M.; Okada, G.; Chen, C.Y.; Huang, J.W.; Kakishima, M.; Ono, T.; Tanaka, K. Resolving the Lophiostoma bipolare complex: Generic delimitations within Lophiostomataceae. Stud. Mycol. 2018, 90, 161–189. [Google Scholar] [CrossRef]

| Taxa | Asci Size (μm) | Ascospores | Septa in Ascospores | Sheath | Habitats & Host Records | Location | References | |
|---|---|---|---|---|---|---|---|---|
| Morphology | Size (μm) | |||||||
| Melomastia aquatica | 185–230 × 7–9 | Fusiform | 26–34 × 6–8 | 3 | Yes | Freshwater/Unknown | China | [9] |
| M. chromolaenae | 135–160 × 7–8 | Fusiform | 29–35 × 4.5–6 | 9–11 | No | Terrestrial/Chromolaena odorata | Thailand | [42] |
| M. clematidis | 115–160 × 4–7 | Broad fusiform with acute ends | 13–20 × 3.8–5 | 3 | Yes | Terrestrial/Clematis sikkimensis | Thailand | [19] |
| M. distoseptata | 126.7–146.2 × 4.7–6.3 | Fusoid, obtuse ends | 19.7–24.9 × 4.3–5 | 3 | No | Terrestrial/Unknown | Andaman | [15] |
| M. fulvicomae | 70–90 × 4–6 | Broad fusiform with rounded ends, ends acute | 9–15 × 3.5–5.5 | 2–3 | Yes | Terrestrial/Clematis fulvicoma | Thailand | [19] |
| M. fusispora | 200–231 × 7.6–9.2 | Fusiform | 27.5–32 × 6.5–7.5 | 3 | Yes | Terrestrial/Olea europaea | China | this study |
| M. italica | 120–190 × 5.1–8.9 | Ellipsoidal | 8.8–10.5 × 2.8–4.11 | 2 | Yes | Terrestrial/Vitis vinifera | Italy | [18] |
| M. mangrovei | 154–216 × 8.5–14 | Fusiform | 26–33 × 6–8 | 7–9 | Yes | Intertidal/Rhizophora sp. | Thailand | [21] |
| M. maolanensis | (95–) 103–118 (–122) × 4–5.5 | Fusiform with round ends | 13.5–18 × 3.5–4.5 | 3 | No | Terrestrial/Unknown | China | [28] |
| M. marinospora | 190–240 × 10–12 | Cylindrical with acute poles | 25–31 × 7.5–10 | 3 | Yes | Intertidal/Kandelia candel | Brunei | [21] |
| M. neothailandica | 165–190 × 10–12 | Ellipsoidal | 26–28 × 7.2–8 | 5 | Yes | Marine/Rhizophora sp. | Thailand | [43] |
| M. oleae | 209–237 × 7.5–9 | Fusiform, obtuse ends | 28–34 × 6–7 | 3 | No | Terrestrial/Olea europaea | China | this study |
| M. phetchaburiensis | 190–300 × 8–12 | Ellipsoidal | 35–40 × 5– 10 | 1–10 | No | Marine/Rhizophora apiculata | Thailand | [44] |
| M. rhizophorae | 135–160 × 8–10 | Ellipsoidal | 19–26 × 6– 8 | 4–6 | Yes | Intertidal/Rhizophora apiculata | Thailand | [9] |
| M. sichuanensis | 101–112.5 × 6.5–7.6 | Broad fusiform with rounded ends | 15–17.5 × 4.7–5.1 | 3 | Yes | Terrestrial/Olea europaea | China | this study |
| M. sinensis | 160–220 × 8–10 | Cylindrical | 18–30 × 5– 8 | 6–7 | No | Terrestrial/Camellia sinensis | Thailand | [45] |
| M. thailandica | 146– 158 × 7–9 | Ellipsoidal | 24–32 × 6–8 | 3–5 | Yes | Marine/Avicennia marina | Thailand | [46] |
| M. thamplaensis | 114–160 × 6–8.5 | Fusiform with acute angular ends | 19.5–23.5 × 5–6.5 | 3 | No | Terrestrial/Unknown | China | [28] |
| M. tiomanensis | 316–333 × 12–17 | Spindle-shaped | 69–82 × 9–11 | 6–7 | No | Terrestrial/Rhizophora sp. | Malaysia | [20] |
| M. winteri | 165–189 × 7–8.5 | Fusiform with acute ends | 25–30 × 5–6.5 | 3 | No | Terrestrial/Olea europaea | China | this study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.-L.; Maharachchikumbura, S.S.N.; Cheewangkoon, R.; Liu, J.-K. Reassessment of Dyfrolomyces and Four New Species of Melomastia from Olive (Olea europaea) in Sichuan Province, China. J. Fungi 2022, 8, 76. https://doi.org/10.3390/jof8010076
Li W-L, Maharachchikumbura SSN, Cheewangkoon R, Liu J-K. Reassessment of Dyfrolomyces and Four New Species of Melomastia from Olive (Olea europaea) in Sichuan Province, China. Journal of Fungi. 2022; 8(1):76. https://doi.org/10.3390/jof8010076
Chicago/Turabian StyleLi, Wen-Li, Sajeewa S. N. Maharachchikumbura, Ratchadawan Cheewangkoon, and Jian-Kui Liu. 2022. "Reassessment of Dyfrolomyces and Four New Species of Melomastia from Olive (Olea europaea) in Sichuan Province, China" Journal of Fungi 8, no. 1: 76. https://doi.org/10.3390/jof8010076