1. Introduction
Grapevine trunk diseases (GTDs) attack the wood of grapevines (
Vitis vinifera) and devastate vineyards worldwide. They have been recognized as a problem for well over a century, but their impact has increased significantly in recent decades [
1]. There are several reasons for this change, one being the sudden intensification of grapevine planting and large-scale production in the early 1990s, which increased the global movement of potentially contaminated material [
2]. Methodologies used in the wine industry modernized, and operations were often transferred from family-sized businesses to large-scale productions that were held to a higher standard of regulation. Such changes did not, however, take place at the nursery scale, leaving guidelines governing the quality of the planting material as they were [
3].
As a result, vine defects originating in nurseries have become a recurring problem. The issues are most commonly attributed to the cutting quality, nursery practices and cold storage conditions [
2,
4,
5]. These defects are then exacerbated in the field, especially as the increased demand for higher production puts additional stress on young vines by incorporating practices that favor fungal infection, such as training vines in high density spur-pruned trellises that are mechanically pruned, or leaving plants with more wounds [
1]. The phasing out of the chemical products effective against GTD fungus has also complicated control efforts. Sodium arsenite, benzimidasole fungicides and methyl bromide were deemed toxic to both humans and the environment in many countries in the early 2000s and the discovery of active ingredients or biocontrol agents that could serve as a replacement option has been ongoing [
6,
7,
8]. Consequently, in-depth study of the grapevine mycobiome and its interaction with the plant has been cited as necessary to fully understand the complexities of GTDs [
9], and for technological development in the control of GTDs [
10].
Grapevine trunk diseases are characterized as colonizing the wood of the perennial organs in the vines, causing wood necrosis and discoloration, vascular infections and/or white rot [
8,
11,
12]. The external symptoms include leaf stripe, apoplexy of some or all of the plant, black measles on berries, and stunted shoots, which can appear suddenly or over the course of several years, and may or may not overlap with the internal decay [
8]. Grapevine trunk disease pathogens are phylogenetically unrelated, belonging to different families, orders and even phyla, although infected plants may reveal similar symptomatology [
9]. Usually, exact symptoms and the timing of their appearance varies among the diseases, as some of the fungi can live for years in the wood without causing any symptoms at all. The fungi that cause GTD symptoms to appear sooner are Petri disease (
Phaeomoniella chlamydospora, Phaeoacremonium spp.,
Cadophora luteo-olivacea, Pleurostoma richardsiae, Cephalosporium spp. and
Acremonium spp.) [
2,
13] and Black-foot disease (
Campylocarpon, Cylindrocladiella, Dactylonectria, Ilyonectria and
Neonectria spp.) [
2]. Petri- and Black-foot diseases are more prone to attack young and replacement vineyards as symptoms will often appear several years after vines are planted in the ground when winter pruning begins (at three years), to when they reach maturity (at five years). In contrast, GTDs that affect mature vineyards are known as the Esca complex (
P. chlamydospora, Phaeoacremonium spp.,
Fomitiporia spp. and
Stereum hirsutum) [
14,
15], Botryosphaeria dieback (
Diplodia seriata, Botryosphaeria dothidea, Lasiodiplodia theobromae and Neofusicoccum parvum) [
16,
17], Phomopsis dieback (
Phomopsis viticola) [
18] and Eutypa dieback (
Eutypa lata and
Diatrypaceae spp.) [
19]. These GTDs are more likely to surface when the plants near 10 years of age, and any time after.
In Catalonia (NE Spain), the increased occurrence of sudden losses incurred by GTDs in 3–5-year-old vines poses a serious threat to the current economy and future outlook of the region. There are 10,000 wine growers and 853 companies with 11 denominations of origin (D.O.) [
20]. The regional industry generates a turnover of 1.18 billion euros annually and represents 19.9% of the wine sector in Spain, making it a growing international marketplace [
20]. A total of 55,118 hectares (ha) are currently utilized for vineyards, and more land has been approved for plantations in the coming years [
21]. Consequently, this region has become a desirable place for young growers to settle and invest. However, with the estimated cost of establishment at 14,860 euros per ha over the 3 years required to plant and install all infrastructure needed for a successful operation [
22], it is paramount that vineyards are equipped with healthy plant material, and the proper knowledge and tools to prevent GTD losses.
Growers lack information on the likelihood of finding GTDs depending on commercial factors such as nursery, scion, or rootstock variety. There are not any cultivars or species in the genus
Vitis that have expressed complete resistance to GTDs [
11], though some have differed in their sensitivity to the development of GTD foliar symptoms [
23], suggesting that degrees of varietal resistance might be under genetic control [
24,
25]. Work has also been done to understand the degree of sensitivity to the disease of the rootstock, finding that some rootstocks or crosses are more resistant to certain GTDs [
26,
27]. For Esca disease in particular, higher sensitivity was seen for grapevine grafted onto rootstock with a higher tannin content [
28]. Regardless, the final disease or resistance results from an interaction among the virulence of the pathogen, the capabilities of the host-plant defense, and the environmental conditions [
29].
The mechanism by which GTD-related fungi shift from a latent to a pathogenic state and trigger the expression of disease is still a matter of debate [
1]. The main hypothesis is that young vines infected with the pioneer fungi
P. chlamydospora or species of
Phaeoacremonium can later develop symptoms following the colonization by other basidiomycetes, although Koch’s postulates have not yet been completed to support such claims [
15]. It has also been hypothesized that biotic or abiotic stresses act as an inciting factor in the shift of fungi to a pathogenic state [
30], but this hypothesis also lacks substantial supporting evidence. Therefore, thoroughly mapping grapevine fungal community structure and microbiome is the next logical step in clarifying community dynamics and understanding how GTDs evolve in various contexts. Thus far, DNA metabarcoding approaches have revealed a higher diversity of taxa and accurate relative abundances in samples coming from the vineyard [
31,
32,
33], though culture-independent studies that used the NGS approach to learn more about the grapevine endosphere are scarce [
33,
34]. Only one study investigated the mycobiome of GTD-affected plants to understand the spatial distribution of the communities present in each plant, linking microbial communities of the wood and the expression of leaf symptoms of Esca [
9]. Work has also been done to analyze the biogeographic characterization of the microbial community structure associated with grapevine via grape samples, demonstrating that both Chardonnay and Cabernet Sauvignon varieties, collected across the four major wine regions in California, USA, displayed a specific associated microbiome, suggesting a regional pattern [
35]. Furthermore, the grape cultivar and year of production influenced this microbial structure [
10].
In this study, our aim was to explore variation of the fungal community inhabiting grafted grapevine nursery-stock, with the objectives of investigating each possible source of this variation, and focus on those connected with higher rates of GTD-related fungi. We investigated (1) the role that nurseries were playing in the proliferation of GTD-related fungal communities prior to plant distribution, and tested if storage had a significant effect on the microbial communities; (2) if the presence of GTD-related fungi was correlated with specific varieties and rootstocks; and (3) whether there existed a link between trunk necrosis and the structure of GTD-related fungal communities.
2. Materials and Methods
2.1. Plant Material
The experiment was conducted on healthy-looking bench-grafted bare-root plants that had been propagated as one plant in the field for a year. Plants were collected from nurseries at two sampling times: once during the week of harvest from the field in late autumn/winter (“pre-storage”), and again in early spring (exactly three months later; “post-storage”), after plants had been bundled, bagged, and held in the nurseries’ own cold-storage facilities at 4 °C, subject to the conditions and practices of each individual nursery. In all cases, pre-storage plants received minimum handling and treatment by the nurseries, while post-storage specimens underwent typical preparation for distribution, in which roots were trimmed to 10 cm and graft unions were dipped in paraffin wax in the nursery workshop. Six nurseries from the Catalan regions of either Girona, Tarragona or Barcelona participated in the study.
The plant material included three biological replicates of each scion-rootstock combination pre- and post-storage. We collected an array of red and white wine varieties (16 total), which depending upon the production of each nursery, grafted onto one the four most commonly-employed rootstocks in the region of Catalonia at this time: 110 Richter (R-110), 140 Ruggeri (RU-140), Selection Oppenheim 4 (SO4) and 41B (for white only). The 11 red varieties included: Autumn Royal, Cabernet Sauvignon, Caladoc, Carignan, Garnacha tinta, Merlot, Pinot Noir, Sumoll, Syrah, Tempranillo and Xarel.lo vermell; while the five white varieties collected were: Chardonnay, Macabeu, Malvasia, Parellada and Xarel.lo. The study was designed around color-rootstock combinations (with scion/varieties falling within the color category) because there were not enough replicates of each variety to cross them with each rootstock and nursery. Each color-rootstock combination was replicated at a minimum six times total, and from at least three nurseries (
Table 1). The more common use of certain rootstocks over others (R-110 vs. 41B, for example) is reflective of its frequency of use in Catalonian nursery-stock.
2.2. Necrosis Evaluation
At the time of analysis, which was immediately after leaving the nurseries and being transported to our labs, each plant stem was cut into transversal sections so we could observe the necrosis present. The length (cm) of necrosis starting in the root collar and graft unions was measured as it extended along the inside of the stem. Specifically, root collar necrosis was measured from the base of the stem after all roots had been trimmed, and upwards; carefully quantifying the necrosis present on either side of the cambium and finally taking an average of the two measurements. Graft union necrosis was measured in a similar way, starting the measurement from where the graft could be seen meeting the rootstock. The morphology of the necrosis was also noted (discoloration, black spots, triangular formations).
2.3. Fungal Isolates
For all samples, isolates were gathered from wood chips that had been cut from regions next to cambium of the root collar and graft union. Fungal isolation was carried out by plating surface-sterilized samples cut into 0.5 cm long pieces on 0.3% malt extract agar (MEA) media following Santamaría et al. [
36], which was amended with 0.10 g/L of chloramphenicol. Samples were incubated at 20 °C in the dark for ca. 2–3 days. A maximum of three morphologically distinct colonies per sample type (root collar or graft union) were transferred onto potato dextrose agar (PDA) media amended with 0.10 g/L chloramphenicol and stored at 20 °C in the dark for one week to provide active mycelial growth for DNA extractions. If morphological differences were not obvious, three isolates per shoot were taken at random. Mycelial growth was monitored to ensure the development of pure isolates, and further re-isolations were conducted as necessary.
Mycelium was scraped from the surface of the colony and placed in sterile Eppendorf tubes with 200 μL of NaOH (0.5 M). Samples were then vortexed briefly and centrifuged 12.0× g for 30 s. For the template, 5 μL of the supernatant was transferred to another tube containing 495 μL TrisHCl (0.1 M). PCRs were run by adding 4 μL template solution in 25 μL reactions with DreamTaq polymerase (Thermo Fisher Scientific, Waltham, Massachusetts), following manufacturer’s recommendations and using universal primers ITS1F (5′-3′: CTTGGTCATTTAGAGGAAGTAA) and ITS4 (5′-3′: TCCTCCGCTTATTGATATGC). Each sample was amplified using the following cycling conditions: an initial denaturation step at 95 °C for 3 min, 30 to 35 cycles at 95 °C for 30 s, 60 °C for 30 s, 72 °C for 1 min, and a final elongation at 72 °C for 6 min. Products were purified and sequenced by Macrogen Co. Ltd. (Seoul, South Korea). The sequences were cleaned and clustered using a 98.5% confidence threshold with DNASTAR software and blasted in GenBank, with a preference given to type material. The OTU identities were assigned according to match thresholds of >98% for species, >97% for genus, >95% for family, >92% for order, >90% for class and >80% for phylum. Sequences with <80% match that could not be identified as being fungal sequences were discarded from the dataset.
2.4. Library Preparation for Metabarcoding
For all samples, specimens were gathered at the same time that isolate wood was gathered, from wood chips that had been cut from regions next to cambium of the root collar and graft union. The wood chips were immediately frozen for later analysis. Surface-sterilized graft union and root collar samples were defrosted and chipped with sterile hand shears. Fifty mg of each sample was manually ground using liquid nitrogen and ca. 100 mg of sand. The pestle and mortars used for extraction were cleaned using a 5% NaOCl solution to avoid cross-contamination of DNA between samples. DNA extraction was conducted with some adjustments to the NucleoSpin© Plant II protocol by Macherey-Nagel (2018) from [
37] to improve extractions from woody material. The ITS2 region was used as the metabarcoding marker, and amplifications used ITS4 and ITS7 tagged forward and reverse primers to enable de-multiplexing following [
38]. PCR reactions were conducted at 57 °C in triplicates, and 32 cycles were used to obtain faint bands corresponding to the linear phase of the amplification. PCR products were pooled and cleaned using bead suspension and magnetic separator according to the NucleoMag
® NGS protocol. DNA concentrations were assessed with Qubit™ (Waltham, MA, USA) and the eight equimolar mixtures (pools), totaling 740 samples, were sequenced with 300 bp paired-end read lengths using two lanes of Illumina MiSeq at the Centre for Genomic Regulation (CRG; Barcelona, Spain).
2.5. Quality Control and Bioinformatic Analysis of Metabarcoding Data
The application of quality control, screening and clustering of sequences was conducted using a similarity threshold of 98.5% with the bioinformatics SCATA pipeline (scata.mykopat.slu.se). Sequences with less than 150 base pairs per read were discarded using the amplicon quality option. In addition, primer mismatches and tag jumps were identified and removed from the dataset. In total, 3,889,132 sequences passed quality control, and they were clustered into operational taxonomic units (OTUs) with a 1.5% similarity threshold. The dataset was further trimmed to ensure all OTUs occurred at least in three samples with >10 sequence reads each for the community analysis. We taxonomically classified each OTU by using the Protax software [
39,
40] in PlutoF (26 June 2021,
https://plutof.ut.ee/), choosing a threshold value of 0.5 (plausible classification). This threshold is a stricter criterion than using a 97% sequence similarity threshold [
41]. To exclude OTUs belonging to plants and non-fungal organisms, we performed a Least Common Ancestor (LCA) analysis (minimum score of 300 and a minimum identity of 90%) in MEGAN [
42] with all OTUs that were not classified at phylum level by Protax. We kept the OTUs classified in MEGAN as ‘Fungi’, merged them with the OTUs previously classified by Protax, and classified them as ‘Fungi, unknown phylum’. Post-clustering curation of fungal OTUs was carried out with the ‘lulu’ package in R [
43]. We used the same taxonomic assignment rules as for isolated fungi. The OTU table, ITS sequence of each OTU, and corresponding metadata are deposited in Figshare.
2.6. Statistical Analysis
2.6.1. Necrosis
Statistical analyses and modeling were carried out using either R version 3.6.3 (64 bits) [
44], or JMP
®. The association between location of necrosis and plant characteristics was first examined by determining the significance of each characteristic in a one-way analysis of variance. The location of the necrotic tissue samples, either in the graft union (
n = 369) or root collar (
n = 370), was set as the response factor and two models were created. Sample (plant) characteristics included in the model as independent variables were: color, nursery, storage, rootstock (for root collar samples) and variety (for graft samples). In order to disentangle the interaction “variety x nursery” or “rootstock x nursery”, comparisons between varieties/rootstocks within nurseries and comparisons between nurseries within varieties/rootstocks were conducted. Post hoc comparisons were made using the Student’s t-test at the 0.05 level of significance; significant results were reported.
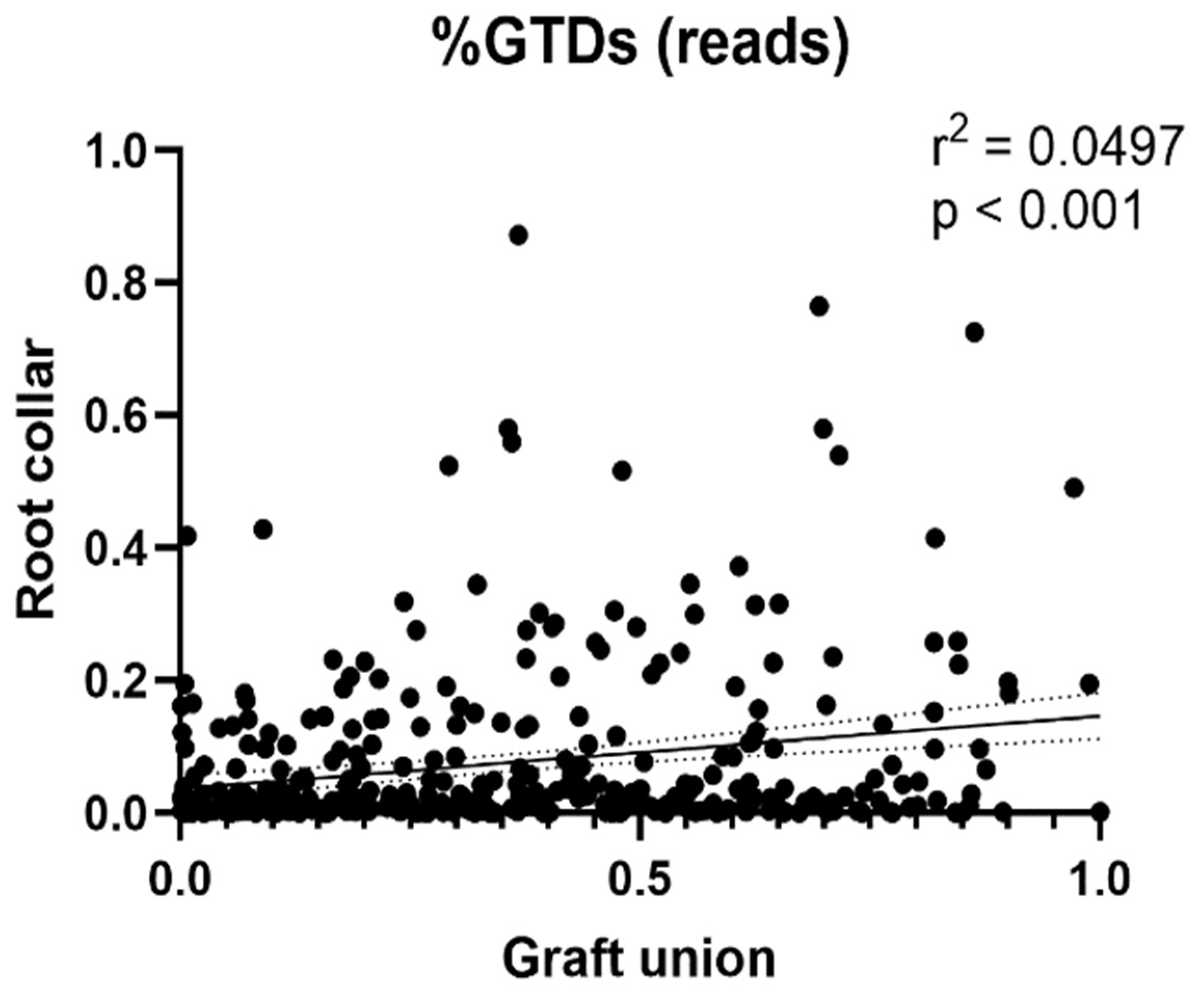
We correlated the relative abundance of GTD reads in each sample (calculated as a sum of GTD reads for each sample, over the total in each tissue type) with the length of necrosis present in the same sample.
To compare if particular GTD-related OTUs were associated with various degrees of necrosis in the two tissue types, we analyzed the first 100 most abundant operational taxonomic units (OTUs), comparing them as a percentage of the total number of reads for each tissue type via stepwise analysis. The top 100 most abundant OTUs were used for this analysis so as to remove rare species and have robust comparison with the more common species. Other parameters (plant characteristics; color, nursery, storage, rootstock, and variety) were also considered in the model, but locked as current estimates in JMP®.
2.6.2. Mapping Fungal Communities
To know which factor(s) had caused significant shifts in fungal taxa, we conducted analysis of our metabarcoding data using R version 3.6.3 (64 bits) [
44]. First, relative abundance of each OTU was standardized with a Hellinger transformation. The entire fungal community and then just GTD-related fungal community were tested by tissue types (graft union and root collar) using Permutational Multivariate Analysis of Variance (PERMANOVA; Adonis2 function) in the Vegan package (v. 2.5–5) [
45]. The data set was then split by tissue type and analyzed for each factor (nursery, storage, variety, and rootstock). Variety was only analyzed for graft unions and rootstock only for root collars. Interactions were also investigated by running separate analysis for each of the levels of the factors. We used ‘vegdist’ function to calculate Bray-Curtis dissimilarities of the community matrices and tested for homogeneity of multivariate dispersion ‘betadisper’ (i.e., multivariate dispersion or beta diversity) using ‘permutest’ function. Changes in community composition were visualized with a principal coordinate analysis (PCoA) based on Bray distances amongst samples. More in-depth analysis was run on representative examples of the degree of variation between nurseries within varieties, and of the degree of variation of storage between nurseries influencing total fungal community and GTD-related fungal community composition. For this we examined variation in Macabeu plantings across nurseries, then the effect of storage in nurseries II and IV pre- and post-storage. The R
2 and
p-values shown in results are extracted from the PERMANOVA analyses performed with the adonis2 function.
The functions ‘diversity’ and ‘specnumber’ from the same vegan package were used to calculate alpha diversity indices, namely species richness, evenness, and the Simpson diversity index. To determine the efficacy of GTD occurrence rates based on reads vs. isolate data, we calculated the rates for each data set and compared them by various factors.
2.7. Indicator Species
An indicator species provides information on the overall condition of the ecosystem and of other species in that ecosystem. They reflect the quality and changes in environmental conditions as well as aspects of community composition. In this work, indicator species tests were used to compare frequency shifts of fungal taxa between tissue types for NGS data (OTU abundance). Indicator species tests were done with a multi-level pattern analysis in R with the ‘multipatt’ function of the ‘indicspecies’ package [
46]. The ‘multipatt’ command results in lists of species that are associated with a particular group of samples and identify species that are statistically more abundant in combinations of categories. We searched for indicators by variety in graft unions and rootstock in root collar.
4. Discussion
Grapevine trunk diseases (GTDs) are caused by cryptic combinations of wood-inhabiting fungi. The intricacies of the relationships between these particular fungi, the plant hosts and environment are still not understood, so an in-depth study of the grapevine mycobiome has been cited as necessary to fully understand the complexities of GTD infections [
9]. In this experiment, we used NGS technology to examine fungal communities in Spanish nursery-stock, and compare results with degrees of internal trunk necrosis present in above- and below-ground plant tissues.
The outcome of mapping the mycobiomes of two tissue locations in the stems of young plants highlighted the influence of nurseries in fungal community composition. Total and GTD-specific fungal communities were significantly different between graft and root tissues, however in both tissues, nursery was the factor explaining the largest amount of variation in both in terms of fungal amplicons or isolates. Our results suggest that either nursery practices or inherent biogeographic fungal community profiles unique to each nursery, play a crucial role in the development of GTDs in nursery stock. With only six nurseries it is not possible to disentangle such effects given the number of steps that could affect GTD variation before plants are sold to the grower. Previous studies have also cited nurseries and their practices as having an impact on vine health, referencing the variation in the many steps and practices involved in creating nursery-stock as a detriment to the quality of planting material [
139]. Other research has directly demonstrated how infection by GTD pathogens occurs during the grafting processes in nurseries [
2,
140]. Certain grafting practices have proven to be more detrimental than others, for example the use of mechanical (omega) grafting machines, which allow plants to be grafted faster and with little training, but which have also been associated with a 50-fold increase in the incidence of esca in the field [
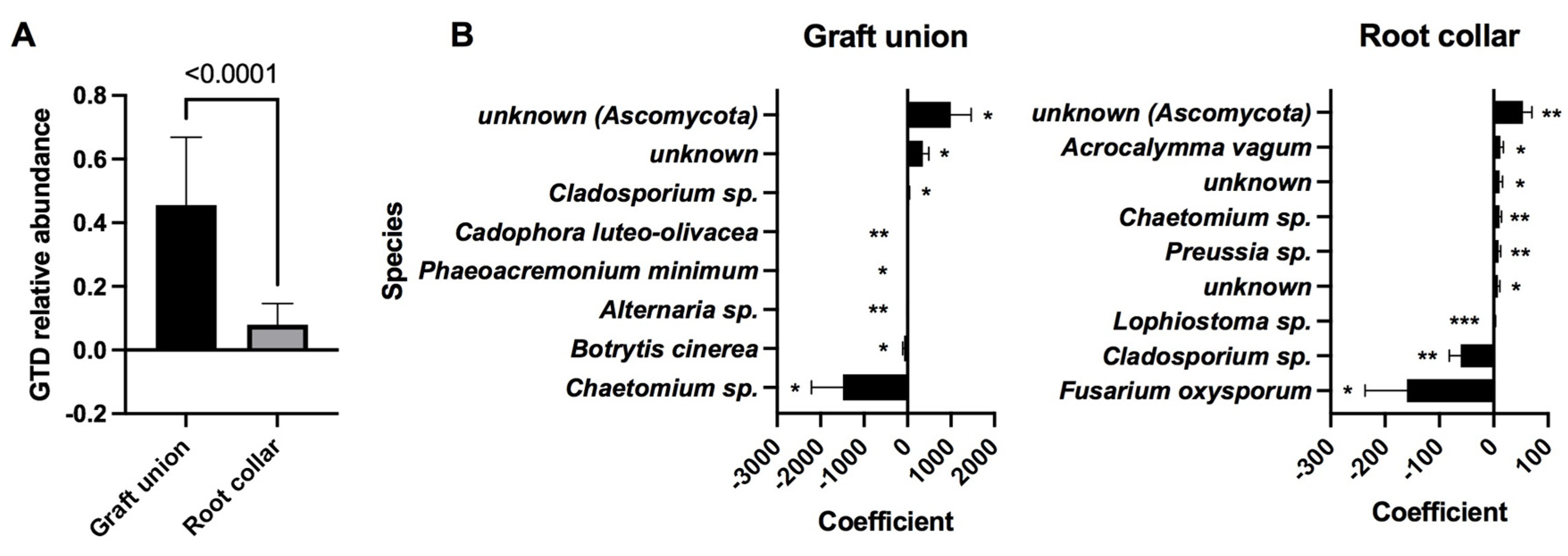
141]. Regardless of the cause, our results show that nurseries explain 24% of the GTD variation in the grafts and 20% of the variation in the rootstock. Buying plants in nurseries with the best practices seems to be the best strategy to minimize the pathogen load and prevent future losses.
The abundance of GTD-related fungus decreased as necrosis increased. This trend was consistent for data from both analyses (isolate and NGS). Previous work has cited similarities in the community structure of symptomatic and non-symptomatic wood [
9], but little explanation has been given as to why some communities induce necrosis, and others do not. There are a number of studies examining the principle factors involved in the proliferation and pathogenicity of GTDs, such as the different fungi present, how these fungi interact with the grapevine cultivar [
11], physiological factors in the plant [
142], and the environment [
8]. The patterns observed at community levels were mirrored by correlations at species level. Both
Cadophora luteo-olivaceae, and
Phaeoacremonium minimum were negatively correlated with graft necrosis.
Cadophora luteo-olivacea is a major pathogen in Petri disease, which is a form of Esca that effects young grapevines, and which in our case was associated with shorter necrosis. The lack of correlation between necrosis and GTD abundance does not downplay the fact that some plants more than others harbored a significant amount of GTDs which could trigger a disease later on.
Our study highlighted the importance of the methodology used to identify fungi: isolation and metabarcoding. These techniques rendered two distinct sets of GTD-related fungi in young nursery plants with little crossover: those that were determined from sequencing isolates collected from symptomatic tissues and those that could only be detected with NGS technology. From isolates,
Neofusicoccum sp.,
Botryosphaeria sp.,
Ilyonectria sp.,
Dactylonectria sp. and
Diplodia sp. were the most common GTD-related species, while
Acremonium alternatum,
C. luteo-olivaceae, and
P. minimum were the most abundant NGS-detected species. Differences between isolation could stem from methodological biases due to growth on PDA media, or due to primer affinity. They could also reflect the viability of the organisms themselves. Before NGS-technology could be used to understand the entire mycobiome of symptomatic and asymptomatic plants, researchers like Hofstetter, Buyck [
143] had found that when comparing asymptomatic and Esca-symptomatic plants, the incidence and abundance of Esca-related fungi were independent of the plant attributes, as all seemingly carried the same fungal parasitic load. These conclusions were drawn with full knowledge that “the term fungal community or mycota that were isolated represented only part of the culturable fungi and missed uncultivable fungal species”. Since there was no significant difference in the systematic structure of the mycota associated with asymptomatic and Esca-symptomatic plants, it was suggested that they are part of the normal mycota associated with adult grapevine plants [
144,
145].
Both variety and rootstock had an influence on necrosis and fungal community. In general, rootstock had a larger influence than variety, which may be explained by biological events tied to the plants’ capacity to store moisture from the soil or an earlier harvest time prior to grafting. More difficult to interpret are the mechanisms behind a higher prevalence of GTDs associated with one variety versus another. The fact that both nursery and variety have effects on necrosis and community structure may reflect the fact that nurseries take grafts from different mother plants, which can in turn harbor a different community of GTDs. We evaluated our results in context with recent findings by Chacón-Vozmediano et al. [
146], to see if there were any varieties that were evidently more susceptible to developing necrosis across conditions. While there were not any direct correlations, our findings coincided in that varieties such as Malvasia, Tempranillo and Macabeu were more susceptible to developing GTD-related symptoms in the field tended to harbor more GTD-related fungi, while Pinot Noir and Parellada were less so (
Table S1). Our results showed Cabernet Sauvignon to be outwardly the least susceptible variety, while observations in the field showed it to rank quite high. The lack of correlation between field and nursery observations may also be explained by the large role played by the nursery producing the plants that were outplanted.
We detected the presence of several biocontrol agents as indicator species in rootstocks SO4 and 41B.
Alternaria sp. was detected in SO4 rootstocks, and has been identified as a promising biocontrol agent against the
Vitis vinifera pathogens
Plasmopara viticola, the causal agent of downy mildew [
147], and
Botrytis cinerea, which causes Botrytis bunch rot [
148]. The mode of action for
Alternaria sp. is the production of resveratrol, an antioxidant compound known to increase resistance to stress and prolong the life of the organisms which it inhabits [
149]. While there were not any GTD-related indicator species in SO4, this rootstock did have the most necrosis, so
Alternaria sp. may be interacting with the rootstock to increase its tolerance to the stress of the necrosis, or may play a role in preventing pathogen colonization.
Lophiostoma sp. is another potential biocontrol agent that was detected as as indicator species in 41B rootstocks. Specifically,
Lophiostoma sp. was reported as producing antifungal compounds (in addition to metabolites that are effective against pathogenic bacteria), so has been cited as a potential biocontrol agent for vine-related pathogens, such as
B. cinerea [
56]. In our work,
Lophiostoma sp. appeared accompanied by the GTD-related fungus
Truncatella sp., a species that has repeatedly been associated with esca complex fungi [
150], as well as
Acremonium sp., which is a non-pathogenic fungus that has recently been found to be an indicator of symptoms (necrosis) in non-GTD infected vines [
151]. Considering the co-occurrence of these indicators in the context of previous work emphasizes the plasticity of their presence and relationships to other indicator species, which can differ depending on the situation in which they are detected (variety, rootstock, environment, etc.).
Our study highlighted how drastically GTD composition can change depending on nursery provenance, even if considering the same rootstocks or varieties. Likewise, variation of GTDs can be acquired if plants belong to one variety or rootstock, or another. In general, we found that Carignan was the variety with the highest amount of GTD-related fungi, while Cabernet sauvignon was the variety with the lowest amount of GTDs. We do not know the implications that higher or lower abundance of GTD-related fungi may have after being planted. However, from this study, it is clear that nursery practices can have an impact on pathogen load and a potential influence on the future health of the plantation.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}