
Fusarium musae from Diseased Bananas and Human Patients: Susceptibility to Fungicides Used in Clinical and Agricultural Settings

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains Collection
2.2. DNA Isolation, PCR and Sequencing
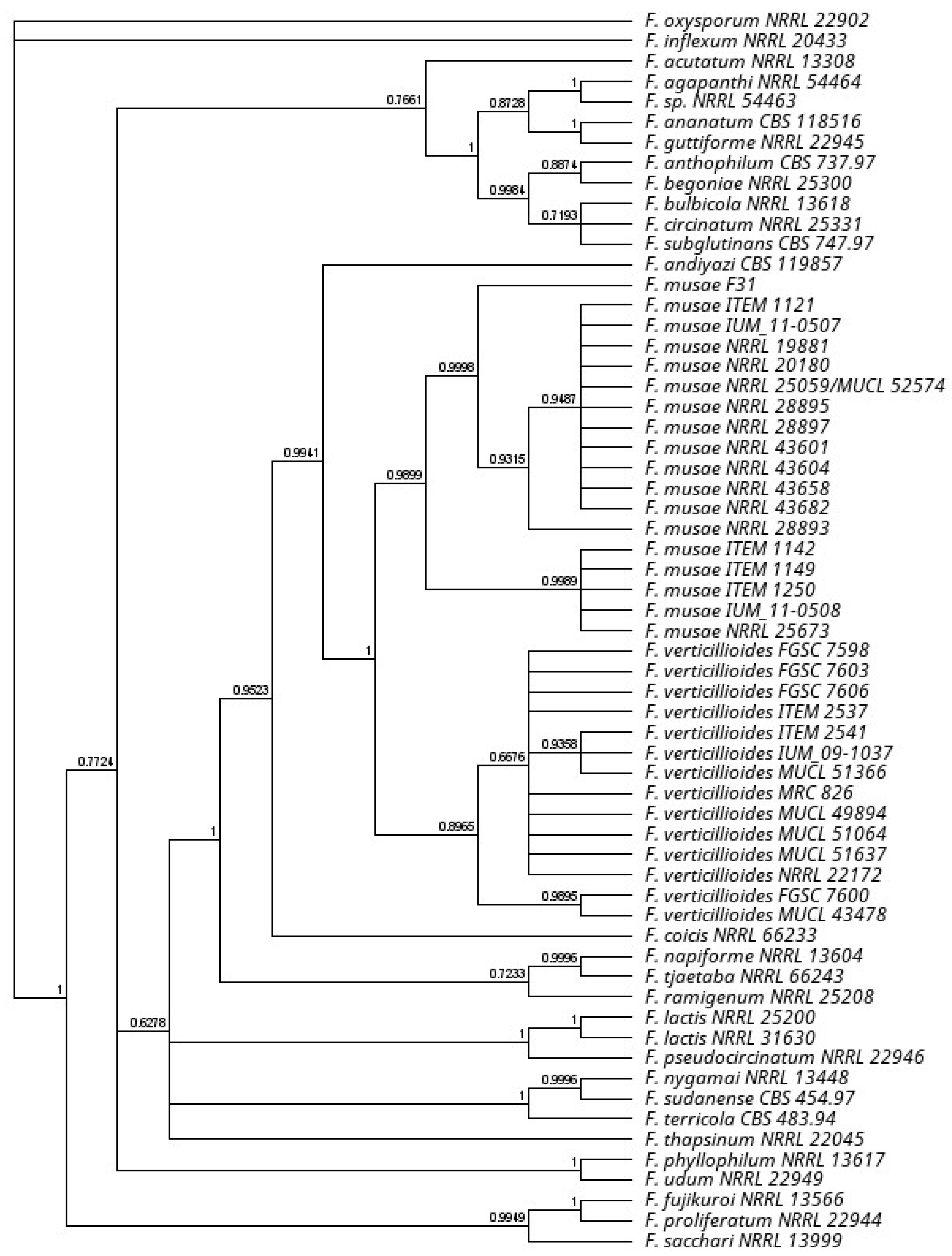
2.3. Phylogenetic Analysis
2.4. Antifungal Susceptibility
2.5. Statistical Analysis
3. Results
3.1. Strain Classification
3.2. Antifungal Susceptibility
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guarro, J. Fusariosis, a Complex Infection Caused by a High Diversity of Fungal Species Refractory to Treatment. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1491–1500. [Google Scholar] [CrossRef]
- Sáenz, V.; Alvarez-Moreno, C.; Pape, P.L.; Restrepo, S.; Guarro, J.; Ramírez, A.M.C. A One Health Perspective to Recognize Fusarium as Important in Clinical Practice. J. Fungi 2020, 6, 235. [Google Scholar] [CrossRef]
- Van Hove, F.; Waalwijk, C.; Logrieco, A.; Munaut, F.; Moretti, A. Gibberella Musae (Fusarium Musae) Sp. Nov., a Recently Discovered Species from Banana Is Sister to F. verticillioides. Mycologia 2011, 103, 570–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple Evolutionary Origins of the Fungus Causing Panama Disease of Banana: Concordant Evidence from Nuclear and Mitochondrial Gene Genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- Kamel, M.A.M.; Cortesi, P.; Saracchi, M. Etiological Agents of Crown Rot of Organic Bananas in Dominican Republic. Postharvest Biol. Technol. 2016, 120, 112–120. [Google Scholar] [CrossRef]
- Triest, D.; Hendrickx, M. Postharvest Disease of Banana Caused by Fusarium musae: A Public Health Concern? PLoS Pathog. 2016, 12, e1005940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triest, D. Banana Fruits Affected by Fusarium Post-Harvest Disease as Source of Human Fusariosis. Acta Microbiol. Immunol. Hung. 2016, 63, 359–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposto, M.C.; Prigitano, A.; Tortorano, A.M. Fusarium musae as Cause of Superficial and Deep-Seated Human Infections. J. Mycol. Med. 2016, 26, 403–405. [Google Scholar] [CrossRef] [PubMed]
- Triest, D.; Piérard, D.; De Cremer, K.; Hendrickx, M. Fusarium musae Infected Banana Fruits as Potential Source of Human Fusariosis: May Occur More Frequently than We Might Think and Hypotheses about Infection. Comm. Integr. Biol. 2016, 9, e1162934. [Google Scholar] [CrossRef] [PubMed]
- Triest, D.; Stubbe, D.; De Cremer, K.; Piérard, D.; Detandt, M.; Hendrickx, M. Banana Infecting Fungus, Fusarium musae, is Also an Opportunistic Human Pathogen: Are Bananas Potential Carriers and Source of Fusariosis? Mycologia 2015, 107, 46–53. [Google Scholar] [CrossRef]
- Tortorano, A.M.; Prigitano, A.; Esposto, M.C.; Arsenijevic, V.A.; Kolarovic, J.; Ivanovic, D.; Paripovic, L.; Klingspor, L.; Nordøy, I.; Hamal, P.; et al. European Confederation of Medical Mycology (ECMM) Epidemiological Survey on Invasive Infections Due to Fusarium Species in Europe. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1623–1630. [Google Scholar] [CrossRef]
- Molnár, O.; Bartók, T.; Szécsi, Á. Occurrence of Fusarium verticillioides and Fusarium musae on Banana Fruits Marketed in Hungary. Acta Microbiol. Immunol. Hung. 2015, 62, 109–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, W.; Tan, Y.; Wang, S.; Gardiner, D.M.; Saeger, S.D.; Liao, Y.; Wang, C.; Fan, Y.; Wang, Z.; Wu, A. Mycotoxigenic Potentials of Fusarium Species in Various Culture Matrices Revealed by Mycotoxin Profiling. Toxins 2017, 9, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirata, T.; Kimishima, E.; Aoki, T.; Nirenberg, H.I.; O’Donnell, K. Morphological and Molecular Characterization of Fusarium verticillioides from Rotten Banana Imported into Japan. Mycoscience 2001, 42, 155–166. [Google Scholar] [CrossRef]
- O’Donnell, K.; Sarver, B.A.J.; Brandt, M.; Chang, D.C.; Noble-Wang, J.; Park, B.J.; Sutton, D.A.; Benjamin, L.; Lindsley, M.; Padhye, A.; et al. Phylogenetic Diversity and Microsphere Array-Based Genotyping of Human Pathogenic Fusaria, Including Isolates from the Multistate Contact Lens-Associated U.S. Keratitis Outbreaks of 2005 and 2006. J. Clin. Microbiol. 2007, 45, 2235–2248. [Google Scholar] [CrossRef] [Green Version]
- Batista, B.G.; de Chaves, M.A.; Reginatto, P.; Saraiva, O.J.; Fuentefria, A.M. Human Fusariosis: An Emerging Infection That Is Difficult to Treat. Rev. Soc. Bras. Med. Trop. 2020, 53, e20200013. [Google Scholar] [CrossRef]
- Al-Hatmi, A.M.S.; Curfs-Breuker, I.; De Hoog, G.S.; Meis, J.F.; Verweij, P.E. Antifungal Susceptibility Testing of Fusarium: A Practical Approach. J. Fungi 2017, 3, 19. [Google Scholar] [CrossRef] [Green Version]
- Al-Hatmi, A.M.S.; Bonifaz, A.; Ranque, S.; Sybren de Hoog, G.; Verweij, P.E.; Meis, J.F. Current Antifungal Treatment of Fusariosis. Int. J. Antimicrob. Agents 2018, 51, 326–332. [Google Scholar] [CrossRef] [Green Version]
- Lamb, D.; Kelly, D.; Kelly, S. Molecular Aspects of Azole Antifungal Action and Resistance. Drug Resist. Updates 1999, 2, 390–402. [Google Scholar] [CrossRef]
- Pasquali, M.; Pallez-Barthel, M.; Beyer, M. Searching molecular determinants of sensitivity differences towards four demethylase inhibitors in Fusarium graminearum field strains. Pest. Biochem. Physiol. 2020, 164, 209–220. [Google Scholar] [CrossRef]
- Hof, H. A New, Broad-Spectrum Azole Antifungal: Posaconazole? Mechanisms of Action and Resistance, Spectrum of Activity. Mycoses 2006, 49, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Al-Hatmi, A.M.S.; Meis, J.F.; de Hoog, G.S. Fusarium: Molecular Diversity and Intrinsic Drug Resistance. PLoS Pathog. 2016, 12, e1005464. [Google Scholar] [CrossRef] [PubMed]
- Degradi, L.; Tava, V.; Kunova, A.; Cortesi, P.; Saracchi, M.; Pasquali, M. Telomere to telomere genome assembly of Fusarium musae F31, causal agent of crown rot disease of banana. Mol. Plant-Microbe Interact. 2021. [Google Scholar] [CrossRef]
- Colombo, E.M.; Kunova, A.; Gardana, C.; Pizzatti, C.; Simonetti, P.; Cortesi, P.; Saracchi, M.; Pasquali, M. Investigating Useful Properties of Four Streptomyces Strains Active against Fusarium graminearum Growth and Deoxynivalenol Production on Wheat Grains by qPCR. Toxins 2020, 12, 560. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic Relationships among Ascomycetes: Evidence from an RNA Polymerse II Subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Reeb, V.; Lutzoni, F.; Roux, C. Contribution of RPB2 to Multilocus Phylogenetic Studies of the Euascomycetes (Pezizomycotina, Fungi) with Special Emphasis on the Lichen-Forming Acarosporaceae and Evolution of Polyspory. Mol. Phylogenet. Evol. 2004, 32, 1036–1060. [Google Scholar] [CrossRef]
- Al-Hatmi, A.M.S.; Sandoval-Denis, M.; Nabet, C.; Ahmed, S.A.; Demar, M.; Normand, A.-C.; de Hoog, G.S. Fusarium volatile, a New Potential Pathogen from a Human Respiratory Sample. Fungal Syst. Evol. 2019, 4, 171–181. [Google Scholar] [CrossRef]
- Laraba, I.; Kim, H.-S.; Proctor, R.H.; Busman, M.; O’Donnell, K.; Felker, F.C. Fusarium xyrophilum, sp. nov., a member of the Fusarium fujikuroi species complex recovered from pseudoflowers on yellow-eyed grass (Xyris spp.) from Guyana. Mycologia 2020, 112, 39–51. [Google Scholar] [CrossRef]
- Moussa, T.A.A.; Al-Zahrani, H.S.; Kadasa, N.M.S.; Ahmed, S.A.; de Hoog, G.S.; Al-Hatmi, A.M.S. Two New Species of the Fusarium fujikuroi Species Complex Isolated from the Natural Environment. Antonie Van Leeuwenhoek 2017, 110, 819–832. [Google Scholar] [CrossRef] [Green Version]
- Tortorano, A.M.; Prigitano, A.; Dho, G.; Esposto, M.C.; Gianni, C.; Grancini, A.; Ossi, C.; Viviani, M.A. Species Distribution and In Vitro Antifungal Susceptibility Patterns of 75 Clinical Isolates of Fusarium spp. from Northern Italy. Antimicrob. Agents Chemother. 2008, 52, 2683–2685. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Ward, T.J.; Robert, V.A.R.G.; Crous, P.W.; Geiser, D.M.; Kang, S. DNA sequence-based identification of Fusarium: Current status and future directions. Phytoparasitica 2015, 43, 583–595. [Google Scholar] [CrossRef] [Green Version]
- Al-Hatmi, A.M.; Hagen, F.; Menken, S.B.; Meis, J.F.; de Hoog, G.S. Global Molecular Epidemiology and Genetic Diversity of Fusarium, a Significant Emerging Group of Human Opportunists from 1958 to 2015. Emerg. Microbes Infect. 2016, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Herkert, P.F.; Al-Hatmi, A.M.S.; de Oliveira Salvador, G.L.; Muro, M.D.; Pinheiro, R.L.; Nucci, M.; Queiroz-Telles, F.; de Hoog, G.S.; Meis, J.F. Molecular Characterization and Antifungal Susceptibility of Clinical Fusarium Species From Brazil. Front. Microbiol. 2019, 10, 737. [Google Scholar] [CrossRef] [Green Version]
- Espinel-Ingroff, A.; Colombo, A.L.; Cordoba, S.; Dufresne, P.J.; Fuller, J.; Ghannoum, M.; Gonzalez, G.M.; Guarro, J.; Kidd, S.E.; Meis, J.F.; et al. International Evaluation of MIC Distributions and Epidemiological Cutoff Value (ECV) Definitions for Fusarium Species Identified by Molecular Methods for the CLSI Broth Microdilution Method. Antimicrob. Agents Chemother. 2016, 60, 1079–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Status—Eurostat. Available online: https://ec.europa.eu/eurostat/portal/page/portal/product_details/publication?p_product_code=KS-76-06-669 (accessed on 22 April 2021).
- Jones, R.K. Seedling Blight Development and Control in Spring Wheat Damaged by Fusarium graminearum Group 2. Plant Dis. 1999, 83, 1013–1018. [Google Scholar] [CrossRef] [Green Version]
- Azor, M.; Gené, J.; Cano, J.; Sutton, D.A.; Fothergill, A.W.; Rinaldi, M.G.; Guarro, J. In Vitro Antifungal Susceptibility and Molecular Characterization of Clinical Isolates of Fusarium verticillioides (F. moniliforme) and Fusarium thapsinum. Antimicrob. Agents Chemother. 2008, 52, 2228–2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snelders, E.; Camps, S.M.T.; Karawajczyk, A.; Schaftenaar, G.; Kema, G.H.J.; van der Lee, H.A.; Klaassen, C.H.; Melchers, W.J.G.; Verweij, P.E. Triazole Fungicides Can Induce Cross-Resistance to Medical Triazoles in Aspergillus fumigatus. PLoS ONE 2012, 7, e31801. [Google Scholar] [CrossRef] [Green Version]
- Chowdhary, A.; Sharma, C.; van den Boom, M.; Yntema, J.B.; Hagen, F.; Verweij, P.E.; Meis, J.F. Multi-Azole-Resistant Aspergillus Fumigatus in the Environment in Tanzania. J. Antimicrob. Chem. 2014, 69, 2979–2983. [Google Scholar] [CrossRef] [PubMed]
- Pujol, I.; Guarro, J.; Gene, J.; Sala, J. In-Vitro Antifungal Susceptibility of Clinical and Environmental Fusarium spp. Strains. J. Antimicrob. Chem. 1997, 39, 163–167. [Google Scholar] [CrossRef]
- Hartmann, F.E.; Vonlanthen, T.; Singh, N.K.; McDonald, M.C.; Milgate, A.; Croll, D. The complex genomic basis of rapid convergent adaptation to pesticides across continents in a fungal plant pathogen. Mol. Ecol. 2021. [Google Scholar] [CrossRef]


{kind=link}
| Strain | Species | Country | Host (Tissue) | Reference | NCBI Accession Number TEF-1α | NCBI Accession Number RPB2 |
|---|---|---|---|---|---|---|
| F31 | F. musae | Dominican Republic | Banana (fruit) | [23] | MW916961 a | MW916958 a |
| IUM 11-0507 | F. musae | Greece | Human (blood) | [8] | MW916959 a | MW916956 a |
| IUM 11-0508 | F. musae | Greece | Human (cornea) | [8] | MW916960 a | MW916957 a |
| NRRL 28893 | F. musae | Mexico | Banana (fruit) | [3] | FN552092 | FN552114 |
| NRRL 28895 | F. musae | Mexico | Banana | [3] | AF273314.1 | MZ346032 a |
| NRRL 28897 | F. musae | Mexico | Banana | [14] | AF273316.1 | MZ346033 a |
| NRRL 43601 | F. musae | Maryland, USA | Human (skin) | [15] | MZ346030 a | EF470191 |
| NRRL 43604 | F. musae | Ohio, USA | Human (nasal sinus) | [15] | MZ346031 a | EF470194 |
| NRRL 43658 | F. musae | Minnesota, USA | Human (contact lens) | [15] | EF452989 | EF470028 |
| NRRL 43682 | F. musae | Minnesota, USA | Human (cornea) | [15] | EF453009 | EF470048 |
| NRRL 25673 | F. musae | Guatemala | Banana (fruit) | [3] | FN552091 | FN552113 |
| NRRL 25059 (MUCL 52574) * | F. musae | Honduras | Banana (fruit) | [3] | FN552086 | FN552108 |
| IHEM 20180 | F. musae | Brussels, Belgium | Human (sinus biopsy) | [10] | KJ865533 | KM582792 |
| IHEM 19881 | F. musae | Brest, France | Human (shoulder biopsy) | [10] | KJ865532 | KM582791 |
| ITEM 1121 | F. musae | Panama | Banana (fruit) | [3] | FN552093 | FN552115 |
| ITEM 1142 | F. musae | Ecuador | Banana (fruit) | [3] | FN552094 | FN552116 |
| ITEM 1149 | F. musae | Panama | Banana (fruit) | [3] | FN552095 | FN552117 |
| ITEM 1250 | F. musae | Canary Islands | Banana (fruit) | [3] | FN552090 | FN552112 |
| IUM 09-1037 | F. verticillioides | Italy | Human (blood) | [8] | MW915565 a | MW915564 a |
| MUCL 43478 | F. verticillioides | Kansas, USA | Corn | [3] | FN552074 | FN552096 |
| Fusarium (No. of Tested Isolates) | Antifungal | Isolates Origin | No. of Isolates with MIC (mg/L) of: | a G-MIC | b MIC50 | c MIC90 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.03 | 0.06 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | >16 | ||||||
| F. musae (19) | Isavuconazole | total | 2 | 10 | 7 | 2.4 | 2 | 4 | ||||||||
| C (8) | 2 | 4 | 2 | 2 | 2 | 4 | ||||||||||
| V (11) | 6 | 5 | 2.74 | 2 | 4 | |||||||||||
| Itraconazole | total | 5 | 14 | 16 | >16 | >16 | ||||||||||
| C (8) | 8 | 16 | >16 | >16 | ||||||||||||
| V (11) | 5 | 6 | 16 | >16 | >16 | |||||||||||
| Posaconazole | total | 3 | 11 | 5 | 0.54 | 0.5 | 1 | |||||||||
| C (8) | 4 | 4 | 0.71 | 0.5 | 1 | |||||||||||
| V (11) | 3 | 7 | 1 | 0.44 | 0.5 | 1 | ||||||||||
| Voriconazole | total | 14 | 5 | 1.2 | 1 | 2 | ||||||||||
| C (8) | 6 | 2 | 1.19 | 1 | 2 | |||||||||||
| V (11) | 8 | 3 | 1.21 | 1 | 2 | |||||||||||
| Amphotericin B | total | 1 | 18 | 1.93 | 2 | 2 | ||||||||||
| C (8) | 1 | 7 | 1.83 | 2 | 2 | |||||||||||
| V (11) | 11 | 2 | 2 | 2 | ||||||||||||
| Difenoconazole | total | 5 | 12 | 2 | 3.58 | 4 | 4 | |||||||||
| C (8) | 2 | 4 | 2 | 4 | 4 | 8 | ||||||||||
| V (11) | 3 | 8 | 3.31 | 4 | 4 | |||||||||||
| Epoxiconazole | total | 16 | 3 | 1.11 | 1 | 2 | ||||||||||
| C (8) | 5 | 3 | 1.29 | 1 | 2 | |||||||||||
| V (11) | 11 | 1 | 1 | 1 | ||||||||||||
| Fenbuconazole | total | 3 | 16 | >16 | >16 | >16 | ||||||||||
| C (8) | 8 | >16 | >16 | >16 | ||||||||||||
| V (11) | 3 | 8 | >16 | >16 | >16 | |||||||||||
| Flusilazole | total | 7 | 12 | 1.55 | 2 | 2 | ||||||||||
| C (8) | 1 | 7 | 1.83 | 2 | 2 | |||||||||||
| V (11) | 6 | 5 | 1.37 | 1 | 2 | |||||||||||
| Propiconazole | total | 3 | 16 | 3.58 | 4 | 4 | ||||||||||
| C (8) | 1 | 7 | 3.67 | 4 | 4 | |||||||||||
| V (11) | 2 | 9 | 3.53 | 4 | 4 | |||||||||||
| Tebuconazole | total | 5 | 13 | 1 | 0.83 | 1 | 1 | |||||||||
| C (8) | 7 | 1 | 1.09 | 1 | 1 | |||||||||||
| V (11) | 5 | 6 | 0.73 | 1 | 1 | |||||||||||
| Tetraconazole | total | 19 | >16 | >16 | >16 | |||||||||||
| C (8) | 8 | >16 | >16 | >16 | ||||||||||||
| V (11) | 11 | >16 | >16 | >16 | ||||||||||||
| Prochloraz | total | 15 | 4 | 0.14 | 0.12 | 0.25 | ||||||||||
| C (8) | 5 | 3 | 0.16 | 0.12 | 0.25 | |||||||||||
| V (11) | 10 | 1 | 0.13 | 0.12 | 0.12 | |||||||||||
| F. verticillioides (2) | Isavuconazole | total | 1 | 1 | 1.5 | 1 | 2 | |||||||||
| C (1) | 1 | |||||||||||||||
| V (1) | 1 | |||||||||||||||
| Itraconazole | total | 1 | 1 | 8.25 | 0.5 | >16 | ||||||||||
| C (1) | 1 | |||||||||||||||
| V (1) | 1 | |||||||||||||||
| Posaconazole | total | 2 | 0.5 | 0.5 | 0.5 | |||||||||||
| C (1) | 1 | |||||||||||||||
| V (1) | 1 | |||||||||||||||
| Voriconazole | total | 2 | 1 | 1 | 1 | |||||||||||
| C (1) | 1 | |||||||||||||||
| V (1) | 1 | |||||||||||||||
| Amphotericin B | total | 1 | 1 | 1.5 | 1 | 2 | ||||||||||
| C (1) | 1 | |||||||||||||||
| V (1) | 1 | |||||||||||||||
| Difenoconazole | total | 2 | 2 | 2 | 2 | |||||||||||
| C (1) | 1 | |||||||||||||||
| V (1) | 1 | |||||||||||||||
| Epoxiconazole | total | 1 | 1 | 0.31 | 0.12 | 0.5 | ||||||||||
| C (1) | 1 | |||||||||||||||
| V (1) | 1 | |||||||||||||||
| Fenbuconazole | total | 1 | 1 | 1 | 0.5 | 2 | ||||||||||
| C (1) | 1 | |||||||||||||||
| V (1) | 1 | |||||||||||||||
| Flusilazole | total | 1 | 1 | 0.37 | 0.25 | 0.5 | ||||||||||
| C (1) | 1 | |||||||||||||||
| V (1) | 1 | |||||||||||||||
| Propiconazole | total | 1 | 1 | 0.37 | 0.25 | 0.5 | ||||||||||
| C (1) | 1 | |||||||||||||||
| V (1) | 1 | |||||||||||||||
| Tebuconazole | total | 1 | 1 | 0.75 | 0.5 | 1 | ||||||||||
| C (1) | 1 | |||||||||||||||
| V (1) | 1 | |||||||||||||||
| Tetraconazole | total | 1 | 1 | 8.5 | 1 | >16 | ||||||||||
| C (1) | 1 | |||||||||||||||
| V (1) | 1 | |||||||||||||||
| Prochloraz | total | 1 | 1 | 0.18 | 0.12 | 0.25 | ||||||||||
| C (1) | 1 | |||||||||||||||
| V (1) | 1 | |||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tava, V.; Prigitano, A.; Cortesi, P.; Esposto, M.C.; Pasquali, M. Fusarium musae from Diseased Bananas and Human Patients: Susceptibility to Fungicides Used in Clinical and Agricultural Settings. J. Fungi 2021, 7, 784. https://doi.org/10.3390/jof7090784
Tava V, Prigitano A, Cortesi P, Esposto MC, Pasquali M. Fusarium musae from Diseased Bananas and Human Patients: Susceptibility to Fungicides Used in Clinical and Agricultural Settings. Journal of Fungi. 2021; 7(9):784. https://doi.org/10.3390/jof7090784
Chicago/Turabian StyleTava, Valeria, Anna Prigitano, Paolo Cortesi, Maria Carmela Esposto, and Matias Pasquali. 2021. "Fusarium musae from Diseased Bananas and Human Patients: Susceptibility to Fungicides Used in Clinical and Agricultural Settings" Journal of Fungi 7, no. 9: 784. https://doi.org/10.3390/jof7090784