
Population Diversity and Genetic Structure Reveal Patterns of Host Association and Anthropogenic Impact for the Globally Important Fungal Tree Pathogen Ceratocystis manginecans


Abstract
:1. Introduction
2. Materials and Methods
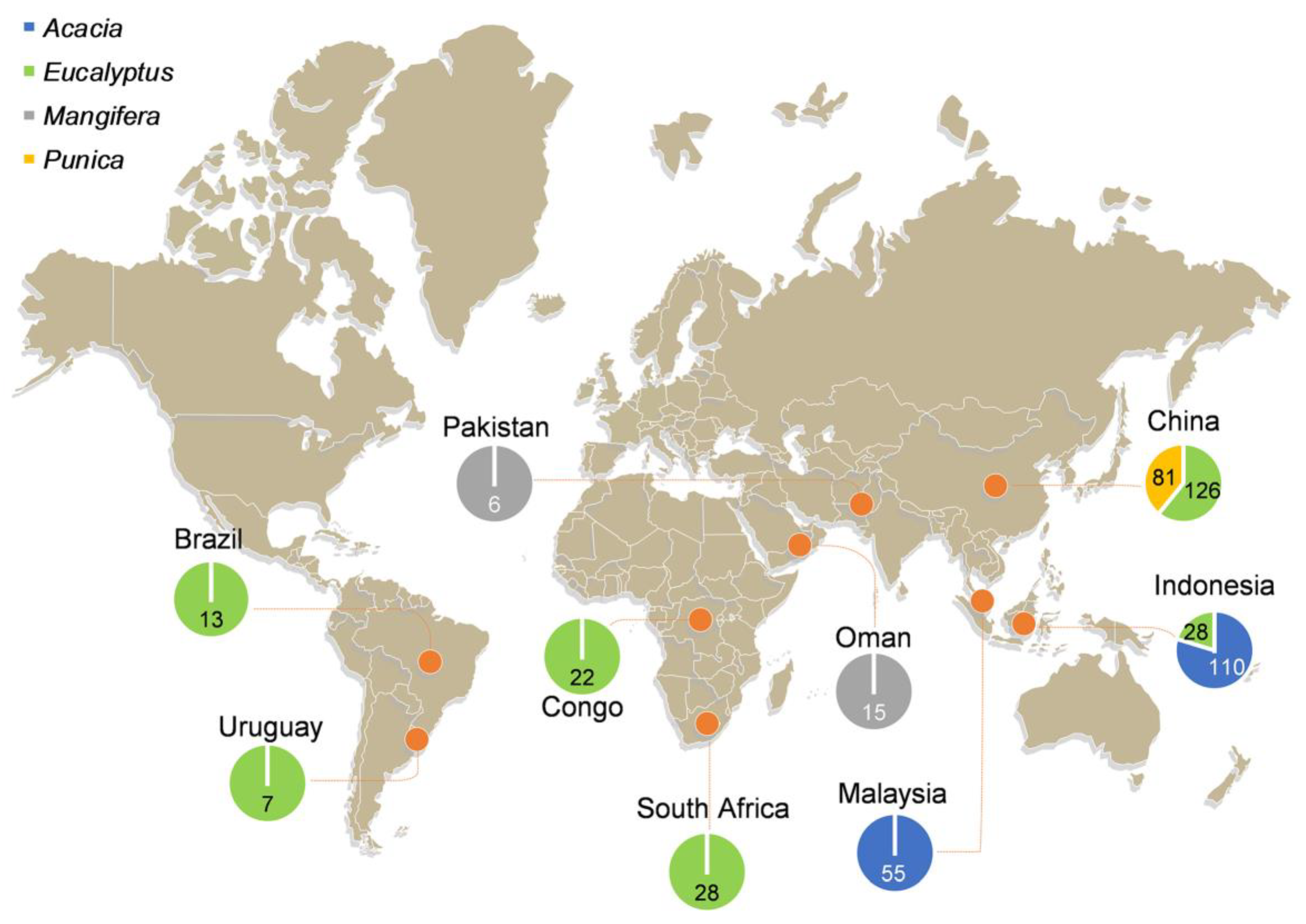
2.1. Fungal Isolates and DNA Extractions
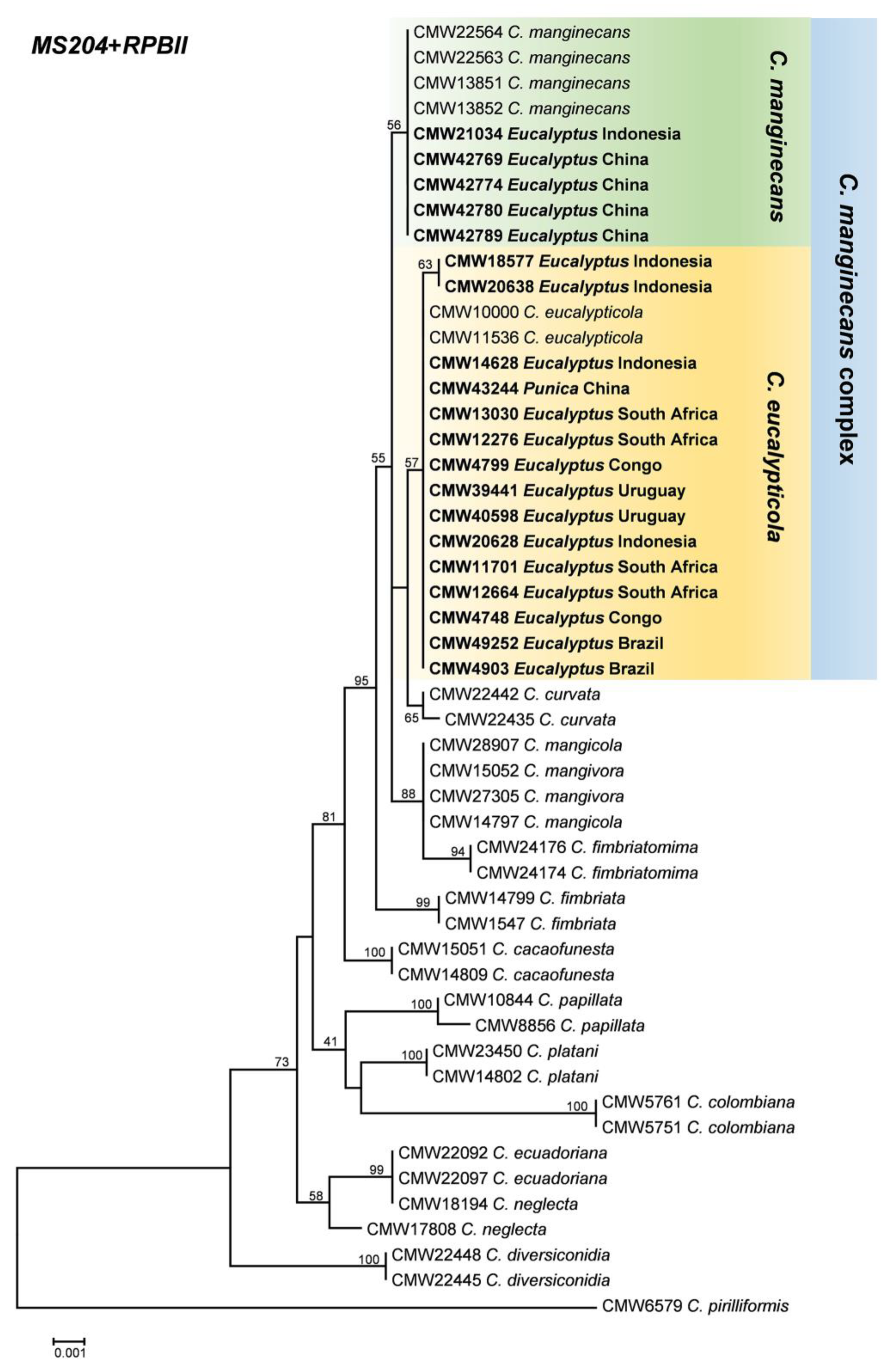
2.2. Confirmation of Isolate Identity
2.3. Microsatellite Marker Genotyping and Allele Scoring
2.4. Population Genetic Diversity Analyses
2.5. Population Molecular Variance, Population Differentiation, and Gene Flow
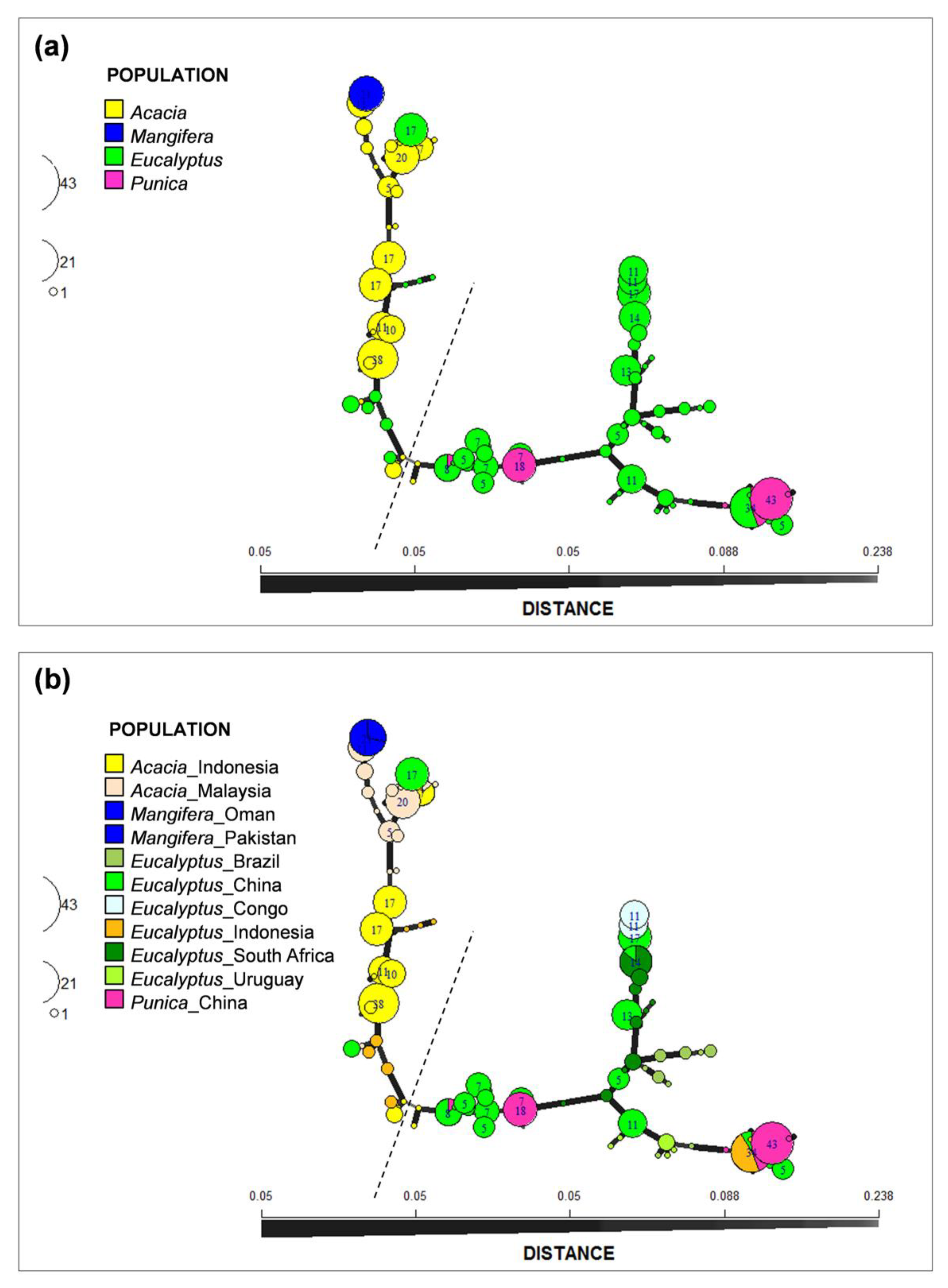
2.6. Minimum Spanning Network Analyses
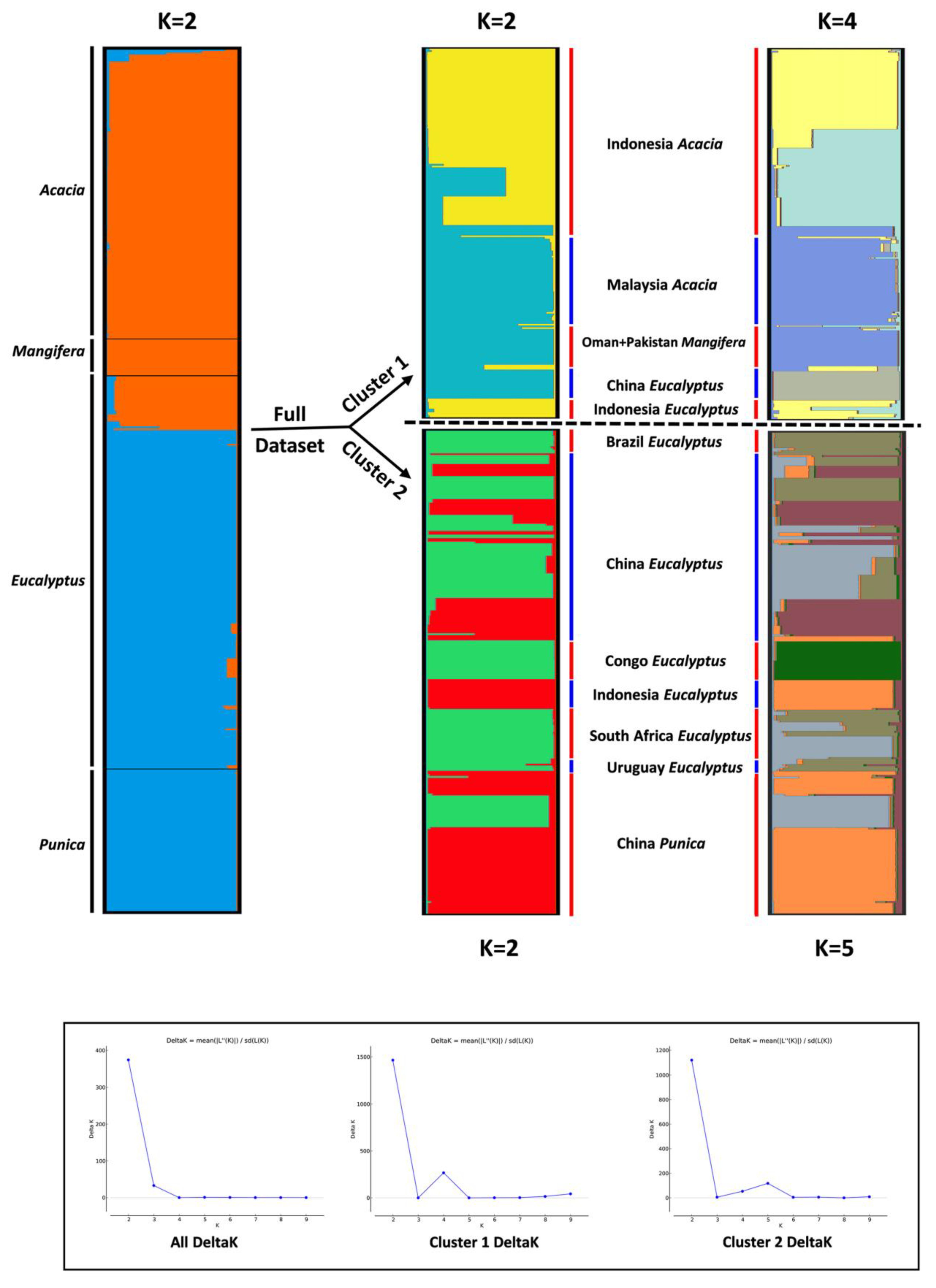
2.7. Analyses of Population Subdivision
2.8. Mode of Reproduction
3. Results
3.1. Fungal Isolates and Identification
3.2. Microsatellite Amplification
3.3. Population Genetic Diversity Analyses
3.4. Population Molecular Variance, Population Differentiation, and Gene Flow
3.5. Minimum Spanning Network Analyses
3.6. Analyses of Population Subdivision
3.7. Mode of Reproduction
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wingfield, M.J.; Slippers, B.; Wingfield, B.D. Novel associations between pathogens, insects and tree species threaten world forests. N. Z. J. For. Sci. 2010, 40, S95–S103. [Google Scholar]
- Cobb, R.C.; Filipe, J.A.; Meentemeyer, R.K.; Gilligan, C.A.; Rizzo, D.M. Ecosystem transformation by emerging infectious disease: Loss of large tanoak from California forests. J. Ecol. 2012, 100, 712–722. [Google Scholar] [CrossRef]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186. [Google Scholar] [CrossRef]
- Boyd, I.L.; Freer-Smith, P.H.; Gilligan, C.A.; Godfray, H.C. The consequence of tree pests and diseases for ecosystem services. Science 2013, 342, 1235773. [Google Scholar] [CrossRef] [PubMed]
- Lombaert, E.; Guillemaud, T.; Cornuet, J.M.; Malausa, T.; Facon, B.; Estoup, A. Bridgehead effect in the worldwide invasion of the biocontrol harlequin ladybird. PLoS ONE 2010, 5, e9743. [Google Scholar] [CrossRef]
- Santini, A.; Ghelardini, L.; De Pace, C.; Desprez-Loustau, M.L.; Capretti, P.; Chandelier, A.; Cech, T.; Chira, D.; Diamandis, S.; Gaitniekis, T.; et al. Biogeographical patterns and determinants of invasion by forest pathogens in Europe. New Phytol. 2013, 197, 238–250. [Google Scholar] [CrossRef] [Green Version]
- Roy, B.A.; Alexander, H.M.; Davidson, J.; Campbell, F.T.; Burdon, J.J.; Sniezko, R.; Brasier, C. Increasing forest loss worldwide from invasive pests requires new trade regulations. Front. Ecol. Environ. 2014, 12, 457–465. [Google Scholar] [CrossRef] [Green Version]
- Wingfield, M.J.; Brockerhoff, E.G.; Wingfield, B.D.; Slippers, B. Planted forest health: The need for a global strategy. Science 2015, 349, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.K. Perspectives of disease threat in large-scale Pinus radiata monoculture-the New Zealand experience. Eur. J. For. Pathol. 1991, 21, 71–81. [Google Scholar] [CrossRef]
- Arnold, J.P.; Fonseca, C.R. Herbivory, pathogens, and epiphylls in Araucaria Forest and ecologically-managed tree monocultures. For. Ecol. Manag. 2011, 262, 1041–1046. [Google Scholar] [CrossRef]
- Felton, A.; Nilsson, U.; Sonesson, J.; Felton, A.M.; Roberge, J.M.; Ranius, T.; Ahlström, M.; Bergh, J.; Björkman, C.; Boberg, J.; et al. Replacing monocultures with mixed-species stands: Ecosystem service implications of two production forest alternatives in Sweden. Ambio 2016, 45, 124–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paine, T.D.; Steinbauer, M.J.; Lawson, S.A. Native and exotic pests of Eucalyptus: A worldwide perspective. Annu. Rev. Entomol. 2011, 56, 181–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, T.I.; Wingfield, M.J. Pathogens on the move: A 100-year global experiment with planted eucalypts. Bioscience 2016, 67, 14–25. [Google Scholar] [CrossRef]
- Campbell, F. The science of risk assessment for phytosanitary regulation and the impact of changing trade regulations. Bioscience 2001, 51, 148–153. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.K.M.; Hovmøller, M.S. Aerial dispersal of fungi on the global and continental scales and its consequences for plant disease. Science 2002, 297, 537–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifert, K.A.; De Beer, Z.W.; Wingfield, M.J. The Ophiostomatoid Fungi: Expanding Frontiers; CBS-KNAW Fungal Biodiversity Centre, CBS: Utrecht, The Netherlands, 2013. [Google Scholar]
- Oliveira, L.S.; Harrington, T.C.; Ferreira, M.A.; Damacena, M.; Al-Sadi, A.M.; Al Mahmooli, I.; Alfenas, A. Species or genotypes? Reassessment of four recently described species of the Ceratocystis wilt pathogen, C. fimbriata, on Mangifera indica. Phytopathology 2015, 105, 1229–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, I.; Fourie, A.; Wingfield, M.J.; Harrington, T.C.; McNew, D.L.; Sugiyama, L.S.; Luiz, B.C.; Heller, W.P.; Keith, L.M. New Ceratocystis species associated with rapid death of Metrosideros polymorpha in Hawaii. Persoonia 2018, 40, 154–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Beer, Z.W.; Duong, T.A.; Barnes, I.; Wingfield, B.D.; Wingfield, M.J. Redefining Ceratocystis and allied genera. Stud. Mycol. 2014, 79, 187–219. [Google Scholar] [CrossRef] [Green Version]
- Morris, M.; Wingfield, M.J.; Beer, C.D. Gummosis and wilt of Acacia mearnsii in South Africa caused by Ceratocystis fimbriata. Plant Pathol. 1993, 42, 814–817. [Google Scholar] [CrossRef]
- Roux, J.; Heath, R.; Labuschagne, L.; Nkuekam, G.K.; Wingfield, M.J. Occurrence of the wattle wilt pathogen, Ceratocystis albifundus on native South African trees. For. Pathol. 2007, 37, 292–302. [Google Scholar] [CrossRef]
- Van Wyk, M.; Al Adawi, A.O.; Khan, I.A.; Deadman, M.L.; Al Jahwari, A.A.; Wingfield, B.D.; Ploetz, R.; Wingfield, M.J. Ceratocystis manginecans sp. nov., causal agent of a destructive mango wilt disease in Oman and Pakistan. Fungal Divers. 2007, 27, 213–230. [Google Scholar]
- Tsopelas, P.; Santini, A.; Wingfield, M.J.; De Beer, Z.W. Canker stain: A lethal disease destroying iconic plane trees. Plant Dis. 2017, 101, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Roux, J.; Wingfield, M.J. Ceratocystis species: Emerging pathogens of non–native plantation Eucalyptus and Acacia species. South. For. J. For. Sci. 2009, 71, 115–120. [Google Scholar] [CrossRef]
- Tarigan, M.; Roux, J.; Van Wyk, M.; Tjahjono, B.; Wingfield, M.J. A new wilt and die-back disease of Acacia mangium associated with Ceratocystis manginecans and C. acaciivora sp. nov. in Indonesia. S. Afr. J. Bot. 2011, 77, 292–304. [Google Scholar] [CrossRef] [Green Version]
- Brawner, J.; Japarudin, Y.; Lapammu, M.; Rauf, R.; Boden, D.; Wingfield, M.J. Evaluating the inheritance of Ceratocystis acaciivora symptom expression in a diverse Acacia mangium breeding population. South. For. 2015, 77, 83–90. [Google Scholar] [CrossRef]
- Fourie, A.; Wingfield, M.J.; Wingfield, B.D.; Thu, P.Q.; Barnes, I. A possible centre of diversity in South East Asia for the tree pathogen, Ceratocystis manginecans. Infect. Genet. Evol. 2016, 41, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.F.; Van Wyk, M.; Roux, J.; Wingfield, M.J.; Xie, Y.J.; Zhou, X.D. Taxonomy and pathogenicity of Ceratocystis species on Eucalyptus trees in South China, including C. chinaeucensis sp. nov. Fungal Divers. 2013, 58, 267–279. [Google Scholar] [CrossRef]
- Van Wyk, M.; Roux, J.; Nkuekam, G.K.; Wingfield, B.D.; Wingfield, M.J. Ceratocystis eucalypticola sp. nov. from Eucalyptus in South Africa and comparison to global isolates from this tree. IMA Fungus 2012, 3, 45–58. [Google Scholar] [CrossRef]
- Roux, J.; Wingfield, M.J.; Fourie, A.; Noeth, K.; Barnes, I. Ceratocystis wilt on Eucalyptus: First record from South Africa. South. For. J. For. Sci. 2020, 82, 24–31. [Google Scholar] [CrossRef]
- Masood, A.; Saeed, S. Bark beetle, Hypocryphalus mangiferae Stebbing (Coleoptera: Curculionidae: Scolytinae) is a vector of mango sudden death disease in Pakistan. Pak. J. Bot. 2012, 44, 813–820. [Google Scholar]
- Al Adawi, A.O.; Al Jabri, R.; Deadman, M.; Barnes, I.; Wingfield, B.D.; Wingfield, M.J. The mango sudden decline pathogen, Ceratocystis manginecans, is vectored by Hypocryphalus mangiferae (Coleoptera: Scolytinae) in Oman. Eur. J. Plant Pathol. 2013, 135, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Al Adawi, A.O.; Barnes, I.; Khan, I.A.; Deadman, M.L.; Wingfield, B.D.; Wingfield, M.J. Clonal structure of Ceratocystis manginecans populations from mango wilt disease in Oman and Pakistan. Australas. Plant Pathol. 2014, 43, 393–402. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.S.; Damacena, M.B.; Guimarães, L.M.; Siqueira, D.L.; Alfenas, A.C. Ceratocystis fimbriata isolates on Mangifera indica have different levels of aggressiveness. Eur. J. Plant Pathol. 2016, 1454, 847–856. [Google Scholar] [CrossRef]
- Al Adawi, A.O.; Barnes, I.; Khan, I.A.; Al Subhi, A.M.; Al Jahwari, A.A.; Deadman, M.L.; Wingfield, B.D.; Wingfield, M.J. Ceratocystis manginecans associated with a serious wilt disease of two native legume trees in Oman and Pakistan. Australas. Plant Pathol. 2013, 42, 179–193. [Google Scholar] [CrossRef] [Green Version]
- Harrington, T.C.; Kazmi, M.R.; Al-Sadi, A.M.; Ismail, S.I. Intraspecific and intragenomic variability of ITS rDNA sequences reveals taxonomic problems in Ceratocystis fimbriata sensu stricto. Mycologia 2014, 106, 224–242. [Google Scholar] [CrossRef] [PubMed]
- Fourie, A.; Wingfield, M.J.; Wingfield, B.D.; Barnes, I. Molecular markers delimit cryptic species in Ceratocystis sensu stricto. Mycol. Prog. 2015, 14, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Valdetaro, D.C.; Oliveira, L.S.; Guimarães, L.M.; Harrington, T.C.; Ferreira, M.A.; Freitas, R.G.; Alfenas, A.C. Genetic variation, morphology and pathogenicity of Ceratocystis fimbriata on Hevea brasiliensis in Brazil. Trop. Plant Pathol. 2015, 40, 184–192. [Google Scholar] [CrossRef]
- Li, Q.; Harrington, T.C.; McNew, D.; Li, J.; Huang, Q.; Somasekhara, Y.M.; Alfenas, A.C. Genetic bottlenecks for two populations of Ceratocystis fimbriata on sweet potato and pomegranate in China. Plant Dis. 2016, 100, 2266–2274. [Google Scholar] [CrossRef]
- Möller, E.M.; Bahnweg, G.; Sandermann, H.; Geiger, H.H. A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tissues. Nucleic Acids Res. 1992, 20, 6115. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 230–257. [Google Scholar]
- Liu, F.F.; Barnes, I.; Roux, J.; Wingfield, M.J.; Chen, S.F. Molecular phylogenetics and microsatellite analysis reveal a new pathogenic Ceratocystis species in the Asian-Australian Clade. Plant Pathol. 2018, 67, 1097–1113. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, R.; Tarpy, D.R.; Reeve, H.K. Estimating effective paternity number in social insects and the effective number of alleles in a population. Mol. Ecol. 2003, 12, 3157–3164. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoddart, J.A.; Taylor, J.F. Genotypic diversity: Estimation and prediction in samples. Genetics 1988, 118, 705–711. [Google Scholar] [CrossRef]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.; von Holdt, B. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, J.K.; Wen, W. Documentation for Structure Software: Version 2; University of Chicago Press: Chicago, IL, USA, 2004. [Google Scholar]
- Agapow, P.M.; Burt, A. Indices of multilocus linkage disequilibrium. Mol. Ecol. Notes 2001, 1, 101102. [Google Scholar] [CrossRef]
- Liu, F.F.; Mbenoun, M.; Barnes, I.; Roux, J.; Wingfield, M.J.; Li, G.Q.; Li, J.Q.; Chen, S.F. New Ceratocystis species from Eucalyptus and Cunninghamia in South China. Antonie Van Leeuwenhoek 2015, 107, 1451–1473. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.D.; Wingfield, M.J. Eucalypt diseases and their management in China. Australas. Plant Pathol. 2011, 40, 339–345. [Google Scholar] [CrossRef]
- Hunter, G.C.; Crous, P.W.; Carnegie, A.J.; Wingfield, M.J. Teratosphaeria nubilosa, a serious leaf disease pathogen of Eucalyptus spp. in native and introduced areas. Mol. Plant Pathol. 2009, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Morin, L.; Aveyard, R.; Lidbetter, J.R.; Wilson, P.G. Investigating the host-range of the rust fungus Puccinia psidii sensu lato across tribes of the family Myrtaceae present in Australia. PLoS ONE 2012, 7, e35434. [Google Scholar] [CrossRef]
- Slippers, B.; Burgess, T.; Pavlic, D.; Ahumada, R.; Maleme, H.; Mohali, S.; Rodas, C.; Wingfield, M.J. A diverse assemblage of Botryosphaeriaceae infect Eucalyptus in native and non-native environments. South. For. 2009, 71, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Wingfield, M.J.; Slippers, B.; Roux, J.; Wingfield, B.D. Worldwide movement of exotic forest fungi, especially in the Tropics and the Southern hemisphere. Bioscience 2001, 51, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Gryzenhout, M.; Wingfield, B.D.; Wingfield, M.J. Taxonomy, Phylogeny, and Ecology of Bark-Inhabiting and Tree-Pathogenic Fungi in the Cryphonectriaceae; American Phytopathological Society Press: Saint Paul, MN, USA, 2009. [Google Scholar]
- Chen, S.F.; Gryzenhout, M.; Roux, J.; Xie, Y.J.; Wingfield, M.J.; Zhou, X.D. Identification and pathogenicity of Chrysoporthe cubensis on Eucalyptus and Syzygium spp. in South China. Plant Dis. 2010, 94, 1143–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahlgren, R.; Thorne, R.F. The order Myrtales: Circumscription, variation, and relationships. Ann. Mo. Bot. Gard. 1984, 1, 633–699. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Seifert, K.A.; Webber, J.F. Ceratocystis and Ophiostoma: Taxonomy, Ecology and Pathogenicity; American Phytopathological Society Press: Saint Paul, MN, USA, 1993. [Google Scholar]
- Lee, D.H.; Roux, J.; Wingfield, B.D.; Barnes, I.; Mostert, L.; Wingfield, M.J. The genetic landscape of Ceratocystis albifundus populations in South Africa reveals a recent fungal introduction event. Fungal Biol. 2016, 120, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Roux, J.; Wingfield, M.; Bouillet, J.P.; Wingfield, B.D.; Alfenas, A. A serious new wilt disease of Eucalyptus caused by Ceratocystis fimbriata in Central Africa. For. Pathol. 2000, 30, 175–184. [Google Scholar] [CrossRef]
- Naidoo, K.; Steenkamp, E.T.; Coetzee, M.P.; Wingfield, M.J.; Wingfield, B.D. Concerted evolution in the ribosomal RNA cistron. PLoS ONE 2013, 8, e59355. [Google Scholar]
- Kanzi, A.M.; Trollip, C.; Wingfield, M.J.; Barnes, I.; Van der Nest, M.A.; Wingfield, B.D. Phylogenomic incongruence in Ceratocystis: A clue to speciation? BMC Genom. 2020, 21, 1–11. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | N a | Na b | Nef c | Hexp d | E e | MLG f | Emlg g | G h | |
|---|---|---|---|---|---|---|---|---|---|
| Population based on country | Indonesia (Eucalyptus + Acacia) | 138 | 26 | 1.578 | 0.335 | 0.705 | 23 | 6.290 | 7.399 |
| Malaysia (Acacia) | 55 | 23 | 1.240 | 0.147 | 0.513 | 17 | 5.835 | 5.243 | |
| Oman (Mangifera) | 15 | 10 | 1 | 0 | 0 | 1 | 1 | 1 | |
| Pakistan (Mangifera) | 6 | 10 | 1 | 0 | 0 | 1 | 1 | 1 | |
| Congo (Eucalyptus) | 22 | 12 | 1.2 | 0.1 | 1 | 2 | 2 | 2 | |
| China (Eucalyptus + Punica) | 207 | 30 | 1.671 | 0.351 | 0.680 | 28 | 7.301 | 11.528 | |
| Uruguay (Eucalyptus) | 7 | 14 | 1.271 | 0.160 | 0.840 | 5 | 5 | 3.769 | |
| Brazil (Eucalyptus) | 13 | 20 | 1.639 | 0.280 | 0.770 | 9 | 7.692 | 8.048 | |
| South Africa (Eucalyptus) | 28 | 15 | 1.301 | 0.176 | 0.830 | 10 | 5.716 | 4.404 | |
| Sub-population based on country and plant host i | Indonesia (Acacia) | 110 | 23 | 1.421 | 0.250 | 0.684 | 14 | 8.313 | 5.265 |
| Indonesia (Eucalyptus) | 28 | 21 | 1.555 | 0.318 | 0.496 | 9 | 9 | 2.841 | |
| China (Eucalyptus) | 126 | 27 | 1.760 | 0.398 | 0.795 | 22 | 13.977 | 13.121 | |
| China (Punica) | 81 | 17 | 1.273 | 0.165 | 0.691 | 8 | 4.727 | 2.730 |
| Populations | Source of Variation | Degrees of Freedom | Sum of Squares | Mean Squares | Estimate of Variance | Percentage of Total Variation (%) | PhiPT Value | p-Value a |
|---|---|---|---|---|---|---|---|---|
| Geographical origin (Countries) | Among Populations | 8 | 650.64 | 81.83 | 1.80 | 56 | 0.558 | 0.001 |
| Within Populations | 482 | 686.72 | 1.43 | 1.43 | 44 | – | 0.001 | |
| Total | 490 | 1337.36 | – b | 3.22 | 100 | – | – | |
| Host plant | Among Populations | 3 | 548.79 | 182.93 | 1.71 | 51 | 0.513 | 0.001 |
| Within Populations | 487 | 788.57 | 1.62 | 1.62 | 49 | – | 0.001 | |
| Total | 490 | 1337.36 | – | 3.32 | 100 | – | – |
| Congo (Eucalyptus) | South Africa (Eucalyptus) | Uruguay (Eucalyptus) | Brazil (Eucalyptus) | China (Eucalyptus) | China (Punica) | Indonesia (Eucalyptus) | Indonesia (Acacia) | Malaysia (Acacia) | Oman (Mangifera) | Pakistan (Mangifera) | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Congo (Eucalyptus) | - | 0.286 | 0.237 | 0.284 | 0.884 | 0.213 | 0.335 | 0.205 | 0.115 | 0.045 | 0.061 |
| South Africa (Eucalyptus) | 0.637 | - | 0.449 | 0.754 | 3.209 | 0.421 | 0.452 | 0.232 | 0.113 | 0.072 | 0.093 |
| Uruguay (Eucalyptus) | 0.679 | 0.527 | - | 0.788 | 2.305 | 0.526 | 0.720 | 0.228 | 0.110 | 0.030 | 0.056 |
| Brazil (Eucalyptus) | 0.637 | 0.399 | 0.388 | - | 1.416 | 0.454 | 0.614 | 0.265 | 0.132 | 0.084 | 0.131 |
| China (Eucalyptus) | 0.361 | 0.135 | 0.178 | 0.261 | - | 1.671 | 1.631 | 0.455 | 0.294 | 0.272 | 0.300 |
| China (Punica) | 0.701 | 0.543 | 0.487 | 0.524 | 0.230 | - | 1.360 | 0.238 | 0.101 | 0.079 | 0.088 |
| Indonesia (Eucalyptus) | 0.599 | 0.525 | 0.410 | 0.449 | 0.235 | 0.269 | - | 0.601 | 0.190 | 0.159 | 0.210 |
| Indonesia (Acacia) | 0.709 | 0.683 | 0.687 | 0.654 | 0.524 | 0.677 | 0.454 | - | 0.478 | 0.328 | 0.368 |
| Malaysia (Acacia) | 0.813 | 0.815 | 0.820 | 0.791 | 0.630 | 0.831 | 0.725 | 0.511 | - | 0.737 | 0.937 |
| Oman (Mangifera) | 0.917 | 0.875 | 0.943 | 0.856 | 0.648 | 0.864 | 0.758 | 0.604 | 0.404 | - | - |
| Pakistan (Mangifera) | 0.891 | 0.843 | 0.899 | 0.792 | 0.625 | 0.850 | 0.705 | 0.576 | 0.348 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.; Duong, T.A.; Barnes, I.; Wingfield, M.J.; Chen, S. Population Diversity and Genetic Structure Reveal Patterns of Host Association and Anthropogenic Impact for the Globally Important Fungal Tree Pathogen Ceratocystis manginecans. J. Fungi 2021, 7, 759. https://doi.org/10.3390/jof7090759
Liu F, Duong TA, Barnes I, Wingfield MJ, Chen S. Population Diversity and Genetic Structure Reveal Patterns of Host Association and Anthropogenic Impact for the Globally Important Fungal Tree Pathogen Ceratocystis manginecans. Journal of Fungi. 2021; 7(9):759. https://doi.org/10.3390/jof7090759
Chicago/Turabian StyleLiu, Feifei, Tuan A. Duong, Irene Barnes, Michael J. Wingfield, and Shuaifei Chen. 2021. "Population Diversity and Genetic Structure Reveal Patterns of Host Association and Anthropogenic Impact for the Globally Important Fungal Tree Pathogen Ceratocystis manginecans" Journal of Fungi 7, no. 9: 759. https://doi.org/10.3390/jof7090759