
Hydrophobin CmHYD1 Is Involved in Conidiation, Infection and Primordium Formation, and Regulated by GATA Transcription Factor CmAreA in Edible Fungus, Cordyceps militaris



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Gene Deletion, Complementation and Overexpression
2.3. Determination of the Hydrophobicity of the Mycelia and Conidia
2.4. Observation of the Hyphae and Conidia by Light Microscope and Scanning Electron Microscope
2.5. Stress Adaptation Assays
2.6. Heterologous Expression of CmHYD1 in Pichia pastoris and Feeding
2.7. Assays for Fungal Virulence
2.8. Fruiting Body Production in Cordyceps militaris
2.9. Expression of CmAreA in E. coli and Electrophoretic Mobility Shift Assay (EMSA)
2.10. Transient Luciferase Expression in Tobacco
2.11. RNA Extraction and RT-qPCR Analysis
3. Results
3.1. Deletion, Complementation and Overexpression of Cmhyd1 in Cordyceps militaris
3.2. Deletion of Cmhyd1 Affects Cordyceps militaris Morphology and Hydrophobicity
3.3. Deletion of Cmhyd1 Affects Stress Responses in Cordyceps militaris
3.4. Cmhyd1 Is Associated with Infection in Silkworm Pupae
3.5. Deletion of Cmhyd1 Affects the Transcript Level of Conidiation and Infection-Related Genes during Conidium Production and Infection
3.6. Cmhyd1 Is Important in Fruiting Body Development in Cordyceps militaris
3.7. Transcript of Cmhyd1 Is Regulated by CmAreA Cooperating with CmOtam
3.8. CmHYD1 Positively Regulates the Expression of the Transcription Factor Cmarea
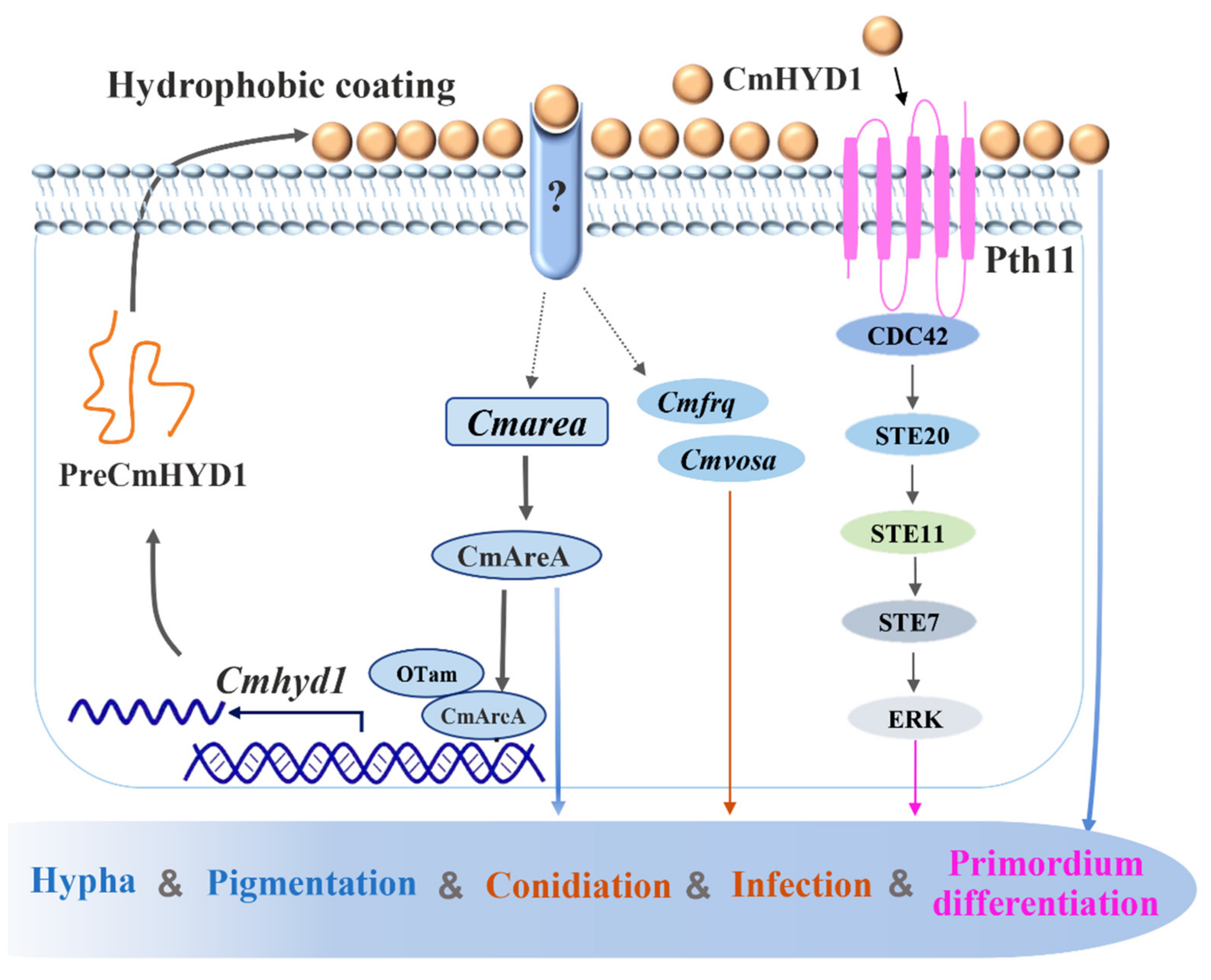
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sunde, M.; Kwan, A.H.; Templeton, M.D.; Beever, R.E.; Mackay, J.P. Structural analysis of hydrophobins. Micron 2008, 39, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Wessels, J.G.H. Developmental regulation of fungal cell-wall formation. Annu. Rev. Phytopathol. 1994, 32, 413–437. [Google Scholar] [CrossRef]
- Mgbeahuruike, A.C.; Kovalchuk, A.; Asiegbu, F.O. Comparative genomics and evolutionary analysis of hydrophobins from three species of wood-degrading fungi. Mycologia 2013, 105, 1471–1478. [Google Scholar] [CrossRef]
- Wösten, H.A. Hydrophobins: Multipurpose proteins. Annu. Rev. Microbiol. 2001, 55, 625–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckerman, J.L.; Ebbole, D.J. MPG1, a gene encoding a fungal hydrophobin of Magnaporthe grisea, is involved in surface recognition. Mol. Plant Microbe. Interact. 1996, 9, 450–456. [Google Scholar] [CrossRef]
- Talbot, N.J.; Kershaw, M.J.; Wakley, G.E.; De Vries, O.; Wessels, J.; Hamer, J.E. MPG1 encodes a fungal hydrophobin involved in surface interactions during infection-related development of Magnaporthe grisea. Plant Cell 1996, 8, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Ahn, I.P.; Rho, H.S.; Lee, Y.H. MHP1, a Magnaporthe grisea hydrophobin gene, is required for fungal development and plant colonization. Mol. Microbiol. 2005, 57, 1224–1237. [Google Scholar] [CrossRef]
- Zhang, S.; Xia, Y.X.; Kim, B.; Keyhani, N.O. Two hydrophobins are involved in fungal spore coat rodlet layer assembly and each play distinct roles in surface interactions, development and pathogenesis in the entomopathogenic fungus, Beauveria bassiana. Mol. Microbiol. 2011, 80, 811–826. [Google Scholar] [CrossRef] [PubMed]
- Sevim, A.; Donzelli, B.G.; Wu, D.; Demirbag, Z.; Gibson, D.M.; Turgeon, B.G. Hydrophobin genes of the entomopathogenic fungus, Metarhizium brunneum, are differentially expressed and corresponding mutants are decreased in virulence. Curr. Genet. 2012, 58, 79–92. [Google Scholar] [CrossRef]
- Jiang, Z.Y.; Ligoxygakis, P.; Xia, Y.X. HYD3, a conidial hydrophobin of the fungal entomopathogen Metarhizium acridum induces the immunity of its specialist host locust. Int. J. Biol. Macromol. 2020, 165, 1303–1311. [Google Scholar] [CrossRef]
- Dubey, M.K.; Jensen, D.F.; Karlsson, M. Hydrophobins are required for conidial hydrophobicity and plant root colonization in the fungal biocontrol agent Clonostachys rosea. BMC Microbiol. 2014, 14, 18. [Google Scholar] [CrossRef] [Green Version]
- Ruocco, M.; Lanzuise, S.; Lombardi, N.; Woo, S.L.; Vinale, F.; Marra, R.; Varlese, R.; Manganiello, G.; Pascale, A.; Scala, V.; et al. Multiple roles and effects of a novel Trichoderma hydrophobin. Mol. Plant Microbe. Interact. 2015, 28, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, U.; Czymmek, K.J.; Sweigard, J.A. Five hydrophobin genes in Fusarium verticillioides include two required for microconidial chain formation. Fungal Genet. Biol. 2004, 41, 852–864. [Google Scholar] [CrossRef] [PubMed]
- Mosbach, A.; Leroch, M.; Mendgen, K.W.; Hahn, M. Lack of evidence for a role of hydrophobins in conferring surface hydrophobicity to conidia and hyphae of Botrytis cinerea. BMC Microbiol. 2011, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- van Wetter, M.; Schuren, F.; Schuurs, T.; Wessels, J. Targeted mutation of the sc3 hydrophobin gene of affects formation of aerial hyphae. FEMS Microbiol. Lett. 1996, 140, 265–269. [Google Scholar] [CrossRef]
- Tao, Y.X.; Chen, R.L.; Yan, J.J.; Long, Y.; Tong, Z.J.; Song, H.B.; Xie, B.G. A hydrophobin gene, Hyd9, plays an important role in the formation of aerial hyphae and primordia in Flammulina filiformis. Gene 2019, 706, 84–90. [Google Scholar] [CrossRef]
- Xu, D.; Wang, Y.; Keerio, A.A.; Ma, A. Identification of hydrophobin genes and their physiological functions related to growth and development in Pleurotus ostreatus. Microbiol. Res. 2021, 247, 126723. [Google Scholar] [CrossRef]
- Banerjee, G.; Robertson, D.L.; Leonard, T.J. Hydrophobins Sc3 and Sc4 gene expression in mounds, fruiting bodies and vegetative hyphae of Schizophyllum commune. Fungal Genet. Biol. 2008, 45, 171–179. [Google Scholar] [CrossRef]
- Yamada, M.; Sakuraba, S.; Shibata, K.; Inatomi, S.; Okazaki, M.; Shimosaka, M. Cloning and characterization of a gene coding for a hydrophobin, Fv-hyd1, specifically expressed during fruiting body development in the basidiomycete Flammulina velutipes. Appl. Microbiol. Biot. 2005, 67, 240–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sammer, D.; Krause, K.; Gube, M.; Wagner, K.; Kothe, E. Hydrophobins in the life cycle of the ectomycorrhizal Basidiomycete Tricholoma vaccinum. PLoS ONE 2016, 11, e0167773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrestha, B.; Zhang, W.; Zhang, Y.; Liu, X. The medicinal fungus Cordyceps militaris: Research and development. Mycol. Prog. 2012, 11, 599–614. [Google Scholar] [CrossRef]
- Guo, M.M.; Guo, S.P.; Yan, H.J.; Bu, N.; Dong, C.H. Comparison of major bioactive compounds of the caterpillar medicinal mushroom, Cordyceps militaris (Ascomycetes), fruiting bodies cultured on wheat substrate and pupae. Int. J. Med. Mushrooms 2016, 18, 327–336. [Google Scholar] [CrossRef]
- Li, X.; Wang, F.; Xu, Y.Y.; Liu, G.J.; Dong, C.H. Cysteine-rich hydrophobin gene family: Genome wide analysis, phylogeny and transcript profiling in Cordyceps militaris. Int. J. Mol. Sci. 2021, 22, 643. [Google Scholar] [CrossRef]
- Lau, G.; Hamer, J.E. Regulatory genes controlling MPG1 expression and pathogenicity in the rice blast fungus Magnaporthe grisea. Plant Cell 1996, 8, 771–781. [Google Scholar] [CrossRef] [Green Version]
- Soanes, D.M.; Kershaw, M.J.; Cooley, R.N.; Talbot, N.J. Regulation of the MPG1 hydrophobin gene in the rice blast fungus Magnaporthe grisea. Mol. Plant Microbe Interact. 2002, 15, 1253–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, W.; Pei, Y.; Bidochka, M.J. A regulator of a G protein signalling (RGS) gene, cag8, from the insect-pathogenic fungus Metarhizium anisopliae is involved in conidiation, virulence and hydrophobin synthesis. Microbiology 2007, 153, 1017–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ries, L.N.; Beattie, S.R.; Espeso, E.A.; Cramer, R.A.; Goldman, G.H. Diverse regulation of the CreA carbon catabolite repressor in Aspergillus nidulans. Genetics 2016, 203, 335–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, Y.L.; Cao, X.; Chen, J.J.; Wang, L.L.; Wei, G.; Wang, S.B. Coordinated regulation of infection-related morphogenesis by the KMT2-Cre1-Hyd4 regulatory pathway to facilitate fungal infection. Sci. Adv. 2020, 6, eaaz1659. [Google Scholar] [CrossRef] [Green Version]
- Zheng, P.; Xia, Y.L.; Xiao, G.H.; Xiong, C.H.; Hu, X.; Zhang, S.W.; Zheng, H.J.; Huang, Y.; Zhou, Y.; Wang, S.Y.; et al. Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional Chinese medicine. Genome Biol. 2011, 12, R116. [Google Scholar] [CrossRef] [Green Version]
- Whiteford, J.R.; Spanu, P.D. The hydrophobin HCf-1 of Cladosporium fulvum is required for efficient water-mediated dispersal of conidia. Fungal Genet. Biol. 2001, 32, 159–168. [Google Scholar] [CrossRef]
- Nebesářová, J. Essential methods of plant sample preparation for high-resolution scanning electron microscopy at room temperature. Methods Mol. Biol. 2019, 1992, 63–76. [Google Scholar] [CrossRef]
- Stübner, M.; Lutterschmid, G.; Vogel, R.F.; Niessen, L. Heterologous expression of the hydrophobin FcHyd5p from Fusarium culmorum in Pichia pastoris and evaluation of its surface activity and contribution to gushing of carbonated beverages. Int. J. Food Microbiol. 2010, 141, 110–115. [Google Scholar] [CrossRef]
- Harith, N.; Abdullah, N.; Sabaratnam, V. Cultivation of Flammulina velutipes mushroom using various agro-residues as a fruiting substrate. Pesqui. Agropecu. Bras. 2014, 49, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.L.; Gong, W.B.; Yan, L.; Zhu, Z.H.; Hu, Z.X.; Peng, Y.D. Biodegradation of ramie stalk by Flammulina velutipes: Mushroom production and substrate utilization. AMB Express 2017, 7, 171. [Google Scholar] [CrossRef]
- Shang, Y.; Yan, L.; Liu, Z.Q.; Cao, Z.; Mei, C.; Xin, Q.; Wu, F.Q.; Wang, X.F.; Du, S.Y.; Jiang, T.; et al. The Mg-chelatase H subunit of Arabidopsis antagonizes a group of WRKY transcription repressors to relieve ABA-responsive genes of inhibition. Plant Cell 2010, 22, 1909–1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lian, T.T.; Yang, T.; Liu, G.J.; Sun, J.D.; Dong, C.H. Reliable reference gene selection for Cordyceps militaris gene expression studies under different developmental stages and media. FEMS Microbiol. Lett. 2014, 356, 97–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arkles, B. Hydrophobicity, Hydrophilicity and Silanes. Paint. Coat. Ind. 2006, 22, 114. [Google Scholar]
- Tong, S.M.; Gao, B.J.; Peng, H.; Feng, M.G. Essential roles of two FRQ Proteins (Frq1 and Frq2) in Beauveria bassiana’s virulence, infection cycle, and calcofluor-specific signaling. Appl. Environ. Microbiol. 2021, 87, e02545-20. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Shi, H.Q.; Ying, S.H.; Feng, M.G. WetA and VosA are distinct regulators of conidiation capacity, conidial quality, and biological control potential of a fungal insect pathogen. Appl. Microbiol. Biotechnol. 2015, 99, 10069–10081. [Google Scholar] [CrossRef]
- Guo, C.T.; Peng, H.; Tong, S.M.; Ying, S.H.; Feng, M.G. Distinctive role of fluG in the adaptation of Beauveria bassiana to insect-pathogenic lifecycle and environmental stresses. Environ. Microbiol. 2021, 05. [Google Scholar] [CrossRef]
- Yang, T.; Dong, C.H. Photo morphogenesis and photo response of the blue-light receptor gene Cmwc-1 in different strains of Cordyceps militaris. FEMS Microbiol. Lett. 2014, 352, 190–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scazzocchio, C. The fungal GATA factors. Curr. Opin. Microbiol. 2000, 3, 126–131. [Google Scholar] [CrossRef]
- Wong, K.H.; Hynes, M.J.; Davis, M.A. Recent advances in nitrogen regulation: A comparison between Saccharomyces cerevisiae and filamentous fungi. Eukaryot. Cell 2008, 7, 917–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giese, H.; Sondergaard, T.E.; Sørensen, J.L. The AreA transcription factor in Fusarium graminearum regulates the use of some nonpreferred nitrogen sources and secondary metabolite production. Fungal Biol. 2013, 117, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Small, A.J.; Hynes, M.J.; Davis, M.A. The TamA protein fused to a DNA-binding domain can recruit AreA, the major nitrogen regulatory protein, to activate gene expression in Aspergillus nidulans. Genetics 1999, 153, 95–105. [Google Scholar] [CrossRef]
- He, R.; Li, C.; Feng, J.; Zhang, D. A class II hydrophobin gene, Trhfb3, participates in fungal asexual development of Trichoderma reesei. FEMS Microbiol. Lett. 2017, 364, fnw297. [Google Scholar] [CrossRef] [Green Version]
- Ball, S.R.; Kwan, A.H.; Sunde, M. Hydrophobin rodlets on the fungal cell wall. Curr. Top. Microbiol. Immunol. 2020, 425, 29–51, ISBN: 978-3-030-49927-3. [Google Scholar]
- Kubicek, C.P.; Baker, S.; Gamauf, C.; Kenerley, C.M.; Druzhinina, I.S. Purifying selection and birth-and-death evolution in the class II hydrophobin gene families of the ascomycete Trichoderma/Hypocrea. BMC Evol. Biol. 2008, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Guo, M.M.; Yang, H.J.; Guo, S.P.; Dong, C.H. The blue-light receptor CmWC-1 mediates fruit body development and secondary metabolism in Cordyceps militaris. Appl. Microbiol. Biotechnol. 2016, 100, 743–755. [Google Scholar] [CrossRef]
- Pianalto, K.M.; Billmyre, R.B.; Telzrow, C.L.; Alspaugh, J.A. Roles for stress response and cell wall biosynthesis pathways in caspofungin tolerance in Cryptococcus neoformans. Genetics 2019, 213, 213–227. [Google Scholar] [CrossRef] [Green Version]
- Moonjely, S.; Keyhani, N.O.; Bidochka, M.J. Hydrophobins contribute to root colonization and stress responses in the rhizosphere-competent insect pathogenic fungus Beauveria bassiana. Microbiology 2018, 164, 517–528. [Google Scholar] [CrossRef]
- Deng, S.; Sun, W.; Dong, L.; Cui, G.; Deng, Y.Z. MoGT2 is essential for morphogenesis and pathogenicity of Magnaporthe oryzae. MSphere 2019, 4, e00309-19. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Llano, Y.; Rodríguez-Pupo, E.C.; Druzhinina, I.S.; Chenthamara, K.; Cai, F.; Gunde-Cimerman, N.; Zalar, P.; Gostinčar, C.; Kostanjšek, R.; Folch-Mallol, J.L.; et al. Stress reshapes the physiological response of halophile fungi to salinity. Cells 2020, 9, 525. [Google Scholar] [CrossRef] [Green Version]
- Khalesi, M.; Jahanbani, R.; Riveros-Galan, D.; Sheikh-Hassani, V.; Sheikh-Zeinoddin, M.; Sahihi, M.; Winterburn, J.; Derdelinckx, G.; Moosavi-Movahedi, A.A. Antioxidant activity and ACE-inhibitory of class II hydrophobin from wild strain Trichoderma reesei. Int. J. Biol. Macromol. 2016, 91, 174–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, G.H.; Ying, S.H.; Zheng, P.; Wang, Z.L.; Zhang, S.W.; Xie, X.Q.; Shang, Y.F.; St Leger, R.J.; Zhao, G.P.; Wang, C.S.; et al. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Sci. Rep. 2012, 2, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wessels, J.G.; de Vries, O.M.; Asgeirsdóttir, S.A.; Springer, J. The thn mutation of Schizophyllum commune, which suppresses formation of aerial hyphae, affects expression of the Sc3 hydrophobin gene. Microbiology 1991, 137, 2439–2445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, R.; Sanz, A.B.; Rodríguez-Peña, J.M.; Nombela, C.; Arroyo, J. Rlm1 mediates positive autoregulatory transcriptional feedback that is essential for Slt2-dependent gene expression. J. Cell Sci. 2016, 129, 1649–1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Wang, F.; Liu, M.; Dong, C. Hydrophobin CmHYD1 Is Involved in Conidiation, Infection and Primordium Formation, and Regulated by GATA Transcription Factor CmAreA in Edible Fungus, Cordyceps militaris. J. Fungi 2021, 7, 674. https://doi.org/10.3390/jof7080674
Li X, Wang F, Liu M, Dong C. Hydrophobin CmHYD1 Is Involved in Conidiation, Infection and Primordium Formation, and Regulated by GATA Transcription Factor CmAreA in Edible Fungus, Cordyceps militaris. Journal of Fungi. 2021; 7(8):674. https://doi.org/10.3390/jof7080674
Chicago/Turabian StyleLi, Xiao, Fen Wang, Mengqian Liu, and Caihong Dong. 2021. "Hydrophobin CmHYD1 Is Involved in Conidiation, Infection and Primordium Formation, and Regulated by GATA Transcription Factor CmAreA in Edible Fungus, Cordyceps militaris" Journal of Fungi 7, no. 8: 674. https://doi.org/10.3390/jof7080674