
Fungal Pathogen Emergence: Investigations with an Ustilago maydis × Sporisorium reilianum Hybrid


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Mating Assays and Filamentous Growth
2.3. Seedling Pathogenesis Assays: Virulence Assessment, Quantification of Anthocyanin Coverage, and Time-Course Harvesting
2.4. Microscopy
2.5. Accession Numbers, Orthology Assessment, and Sequence Analyses
2.6. Total RNA Isolation, DNaseI Treatment, Reverse Transcription, and Species-Specific Transcript Analysis
2.7. Statistical Analyses
2.8. Creation of U. maydis Expression Constructs and Strains
3. Results
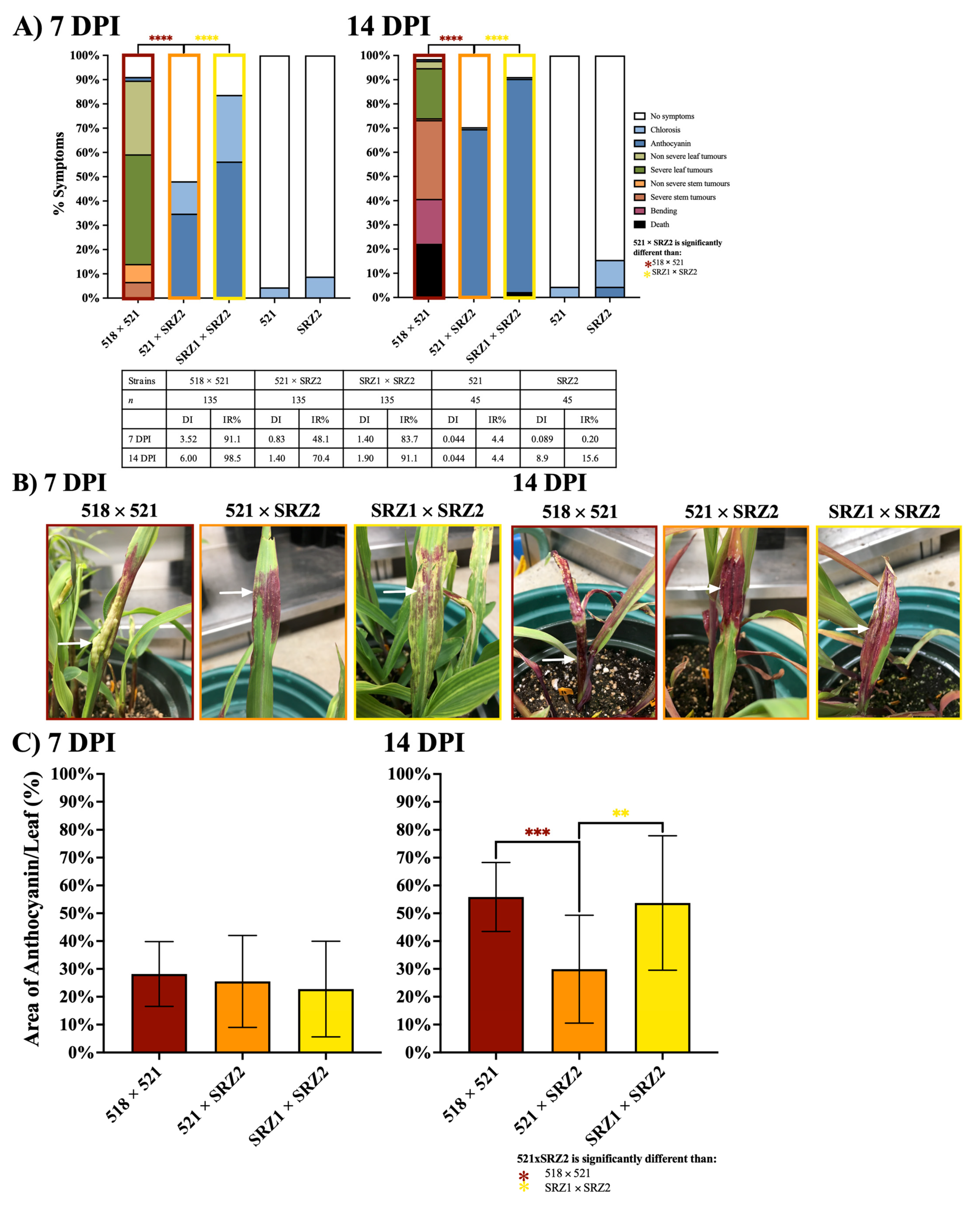
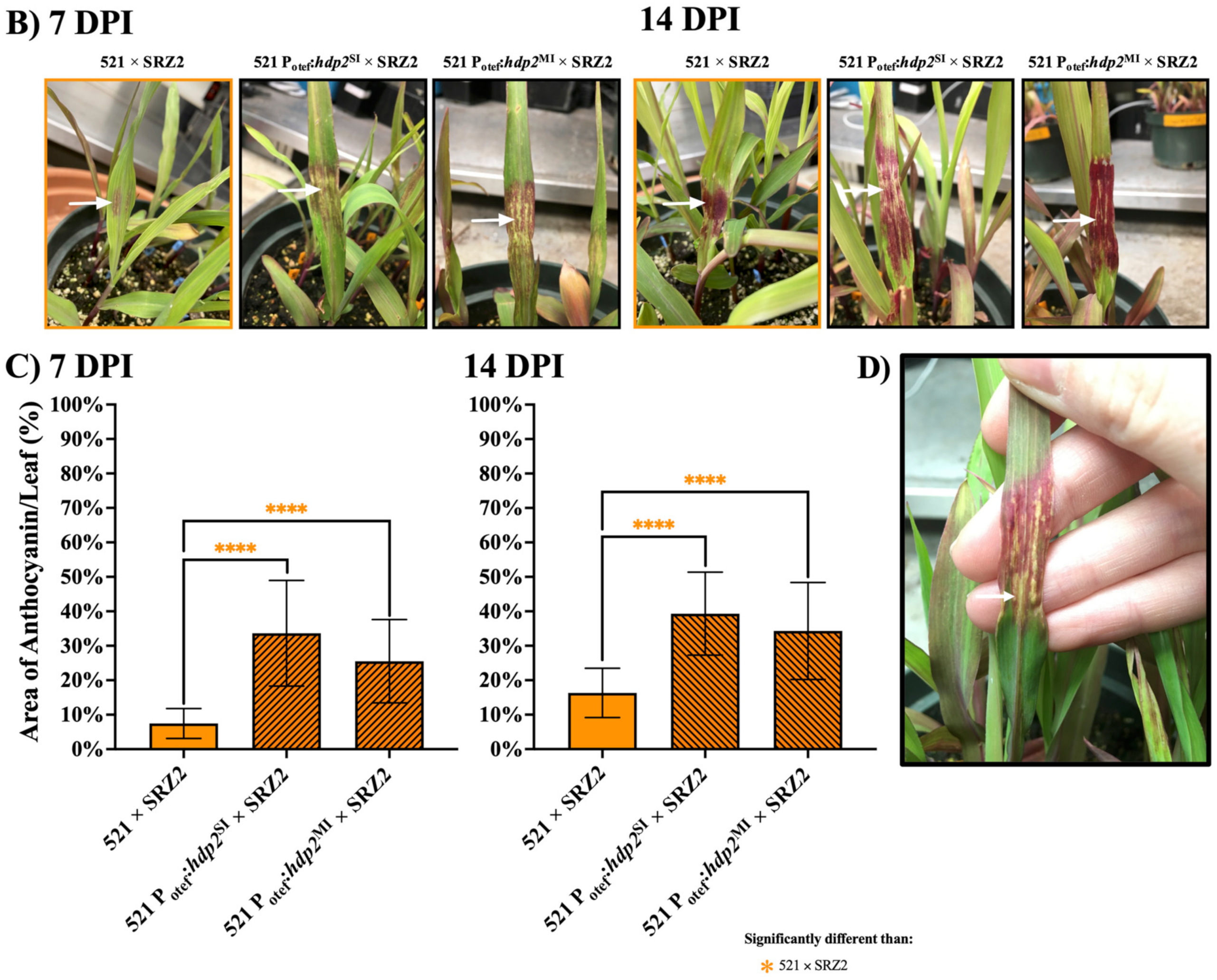
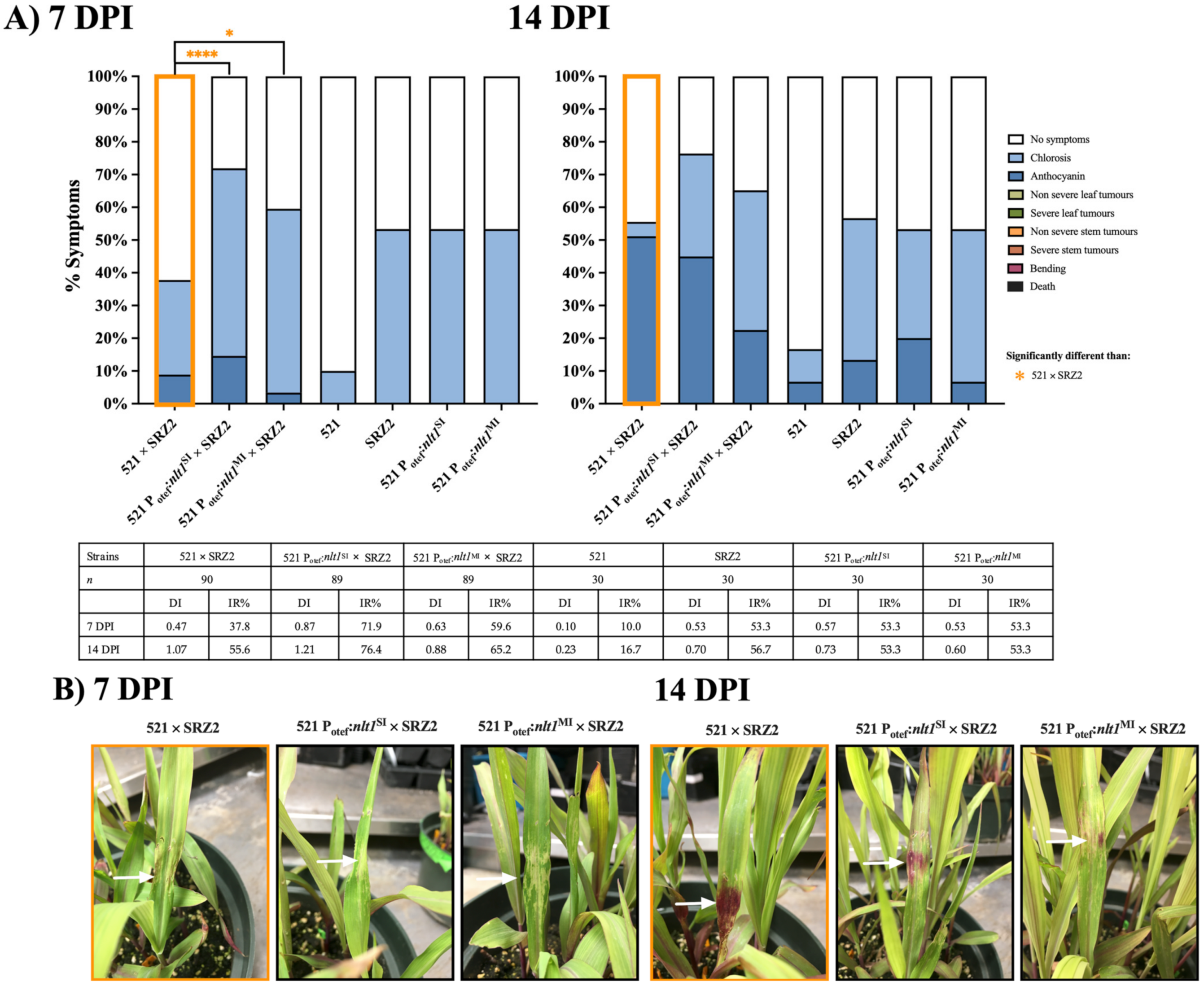
3.1. Induced Filamentation and Virulence Assessment of U. maydis and S. reilianum Hybrids Revealed Sexual Compatibility but Reduced Virulence during Infection
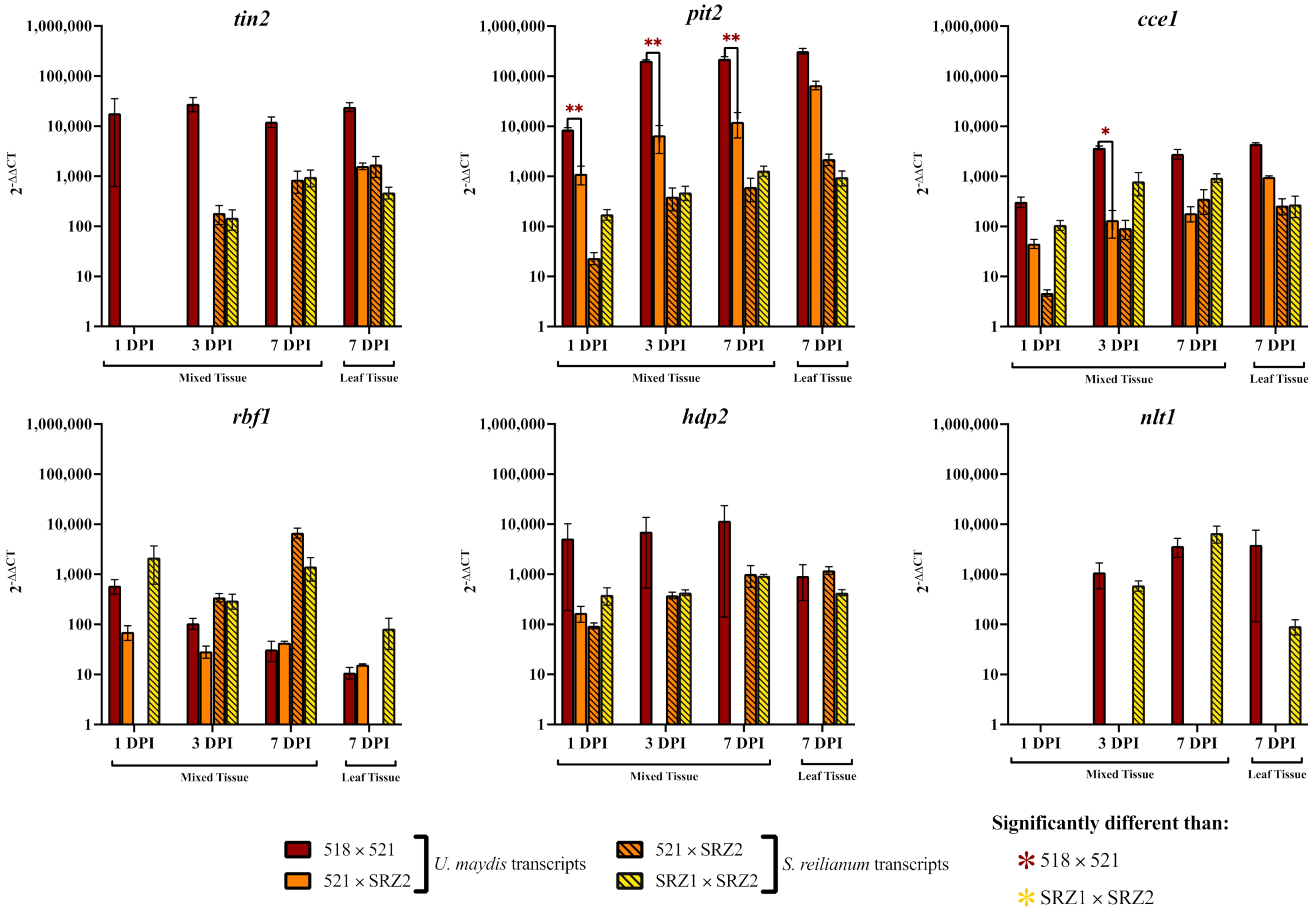
3.2. Reduced Transcript Levels in the Hybrid Dikaryon during Pathogenic Development Revealed Genes to Constitutively Express in the Hybrid
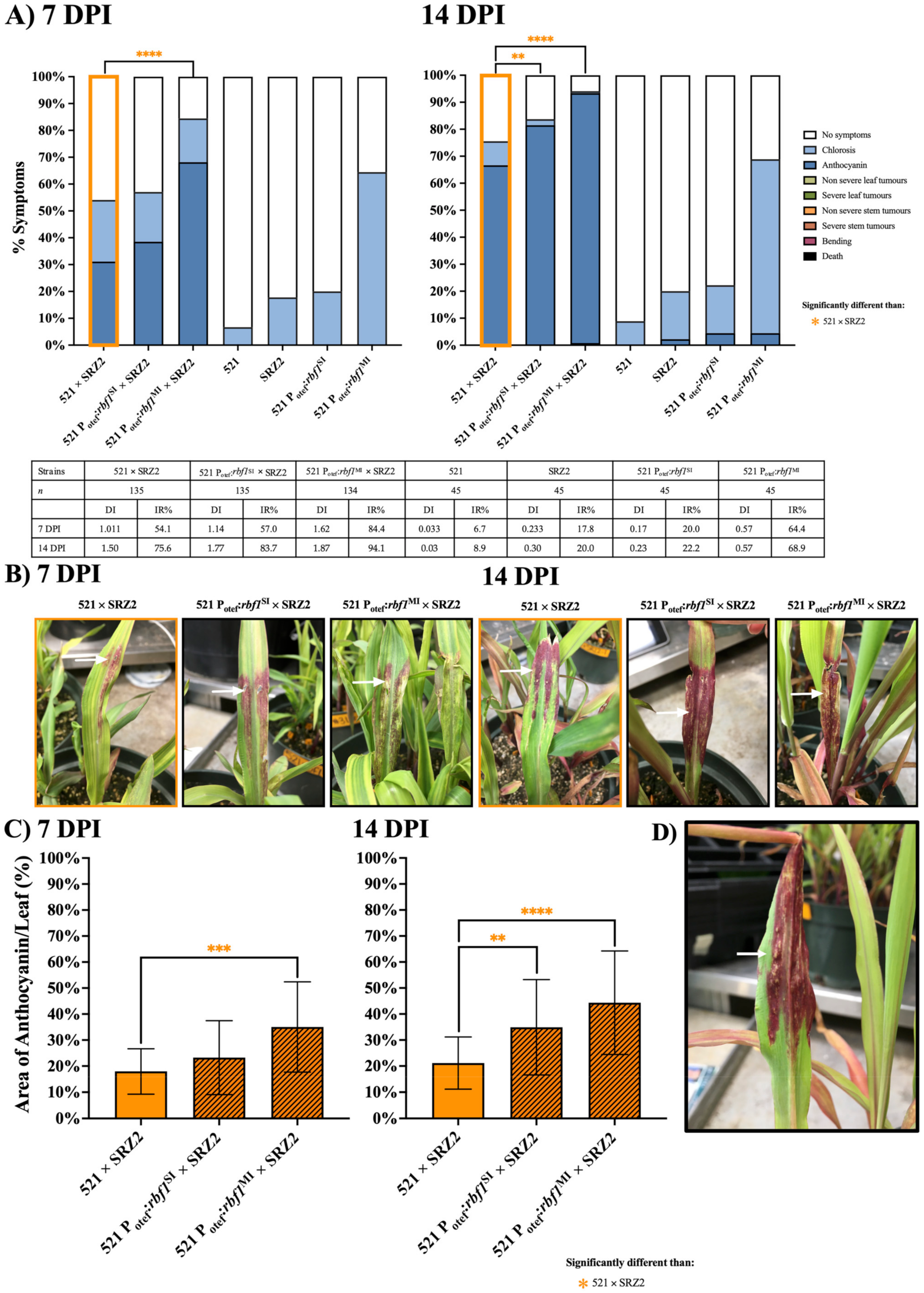
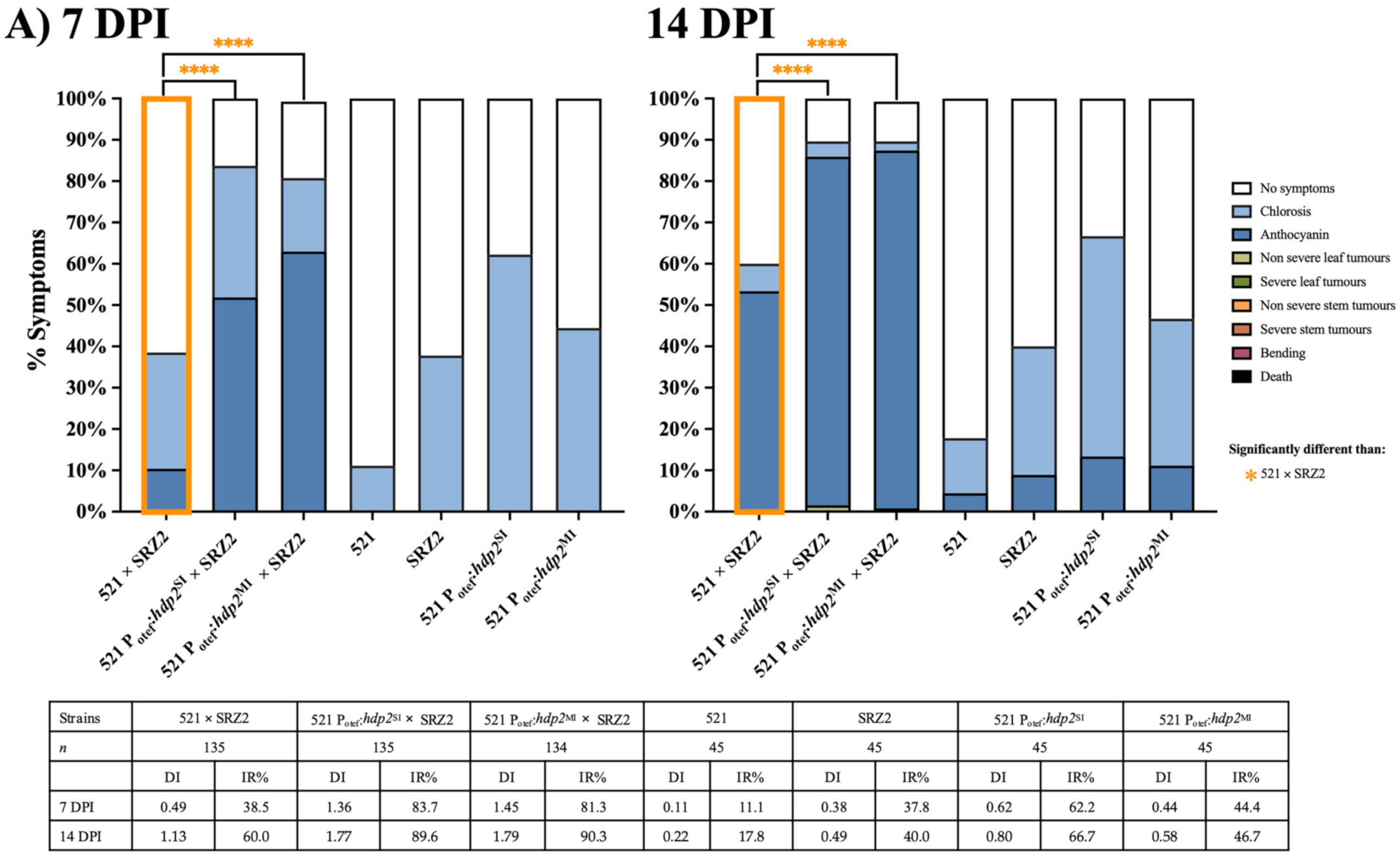
3.3. Constitutively Expressed Virulence Genes within the Hybrid Revealed Transcription Factors Were Capable of Influencing Pathogenic Development
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2013, 484, 1–18. [Google Scholar] [CrossRef]
- Fisher, M.C.; Gow, N.A.R.; Gurr, S.J. Tackling emerging fungal threats to animal health, food security and ecosystem resilience. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20160332. [Google Scholar] [CrossRef] [Green Version]
- Olson, Å.; Stenlid, J. Pathogenic fungal species hybrids infecting plants. Microbes Infect. 2002, 4, 1353–1359. [Google Scholar] [CrossRef]
- Stukenbrock, E.H. The Role of Hybridization in the Evolution and Emergence of New Fungal Plant Pathogens. Phytopathology 2016, 106, 104–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schardl, C.L.; Craven, K.D. Interspecific hybridization in plant-associated fungi and oomycetes: A review. Mol. Ecol. 2003, 12, 2861–2873. [Google Scholar] [CrossRef]
- Kohn, L.M. Mechanisms of Fungal Speciation. Annu. Rev. Phytopathol. 2005, 43, 279–308. [Google Scholar] [CrossRef] [Green Version]
- Menardo, F.; Praz, C.R.; Wyder, S.; Ben-David, R.; Bourras, S.; Matsumae, H.; McNally, K.E.; Parlange, F.; Riba, A.; Roffler, S.; et al. Hybridization of powdery mildew strains gives rise to pathogens on novel agricultural crop species. Nat. Genet. 2016, 48, 201–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newcombe, G.; Stirling, B.; McDonald, S.; Bradshaw, H.D. Melampsora × columbiana, a natural hybrid of M. medusae and M. occidentalis. Mycol. Res. 2000, 104, 261–274. [Google Scholar] [CrossRef]
- Stukenbrock, E.H.; Christiansen, F.B.; Hansen, T.T.; Dutheil, J.Y.; Schierup, M.H. Fusion of two divergent fungal individuals led to the recent emergence of a unique widespread pathogen species. Proc. Natl. Acad. Sci. USA 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertier, L.; Leus, L.; D’hondt, L.; De Cock, A.W.A.M.; Höfte, M. Host adaptation and speciation through hybridization and polyploidy in phytophthora. PLoS ONE 2013, 8, e85385. [Google Scholar] [CrossRef] [Green Version]
- Moon, C.D.; Craven, K.D.; Leuchtmann, A.; Clement, S.L.; Schardl, C.L. Prevalence of interspecific hybrids amongst asexual fungal endophytes of grasses. Mol. Ecol. 2004, 13, 1455–1467. [Google Scholar] [CrossRef]
- Singh, R.P.; Hodson, D.P.; Huerta-Espino, J.; Jin, Y.; Bhavani, S.; Njau, P.; Herrera-Foessel, S.; Singh, P.K.; Singh, S.; Govindan, V. The Emergence of Ug99 Races of the Stem Rust Fungus is a Threat to World Wheat Production. Annu. Rev. Phytopathol. 2011, 49, 465–481. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Upadhyaya, N.M.; Sperschneider, J.; Matny, O.; Nguyen-, H.; Mago, R.; Raley, C.; Miller, M.E.; Silverstein, K.A.; Henningsen, E.; et al. Emergence of the Ug99 lineage of the wheat stem rust pathogen through somatic hybridisation. Nat. Commun. 2019, 10, 5068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begerow, D.; Stoll, M.; Bauer, R. A phylogenetic hypothesis of Ustilaginomycotina based on multiple gene analyses and morphological data. Mycologia 2006, 98, 906–916. [Google Scholar] [CrossRef]
- Stoll, M.; Begerow, D.; Oberwinkler, F. Molecular phylogeny of Ustilago, Sporisorium, and related taxa based on combined analyses of rDNA sequences. Mycol. Res. 2005, 109, 342–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Espinoza, A.D.; García-Pedrajas, M.D.; Gold, S.E. The Ustilaginales as plant pests and model systems. Fungal Genet. Biol. 2002, 35, 1–20. [Google Scholar] [CrossRef]
- Raudaskoski, M.; Kothe, E. Basidiomycete mating type genes and pheromone signaling. Eukaryot. Cell 2010, 9, 847–859. [Google Scholar] [CrossRef] [Green Version]
- Hu, G.; Kamp, A.; Linning, R.; Naik, S.; Bakkeren, G. Complementation of Ustilago maydis MAPK mutants by a wheat leaf rust, Puccinia triticina homolog: Potential for functional analyses of rust genes. Mol. Plant-Microbe Interact. 2007, 20, 637–647. [Google Scholar] [CrossRef] [Green Version]
- Schirawski, J.; Heinze, B.; Wagenknecht, M.; Kahmann, R. Mating type loci of Sporisorium reilianum: Novel pattern with three a and multiple b specificities. Eukaryot. Cell 2005, 4, 1317–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakkeren, G.; Kronstad, J.W. The pheromone cell signaling components of the Ustilago a mating-type loci determine intercompatibility between species. Genetics 1996, 143, 1601–1613. [Google Scholar] [CrossRef]
- Kellner, R.; Vollmeister, E.; Feldbrugge, M.; Begerow, D. Interspecific Sex in Grass Smuts and the Genetic Diversity of Their Pheromone-Receptor System. PLoS Genet. 2011, 7, e1002436. [Google Scholar] [CrossRef] [Green Version]
- Rabe, F.; Bosch, J.; Stirnberg, A.; Guse, T.; Bauer, L.; Seitner, D.; Rabanal, F.A.; Czedik-Eysenberg, A.; Uhse, S.; Bindics, J.; et al. A complete toolset for the study of Ustilago bromivora and Brachypodium sp. as a fungal-temperate grass pathosystem. Elife 2016, 5, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Bosch, J.; Czedik-Eysenberg, A.; Hastreiter, M.; Khan, M.; Güldener, U.; Djamei, A. Two Is Better Than One: Studying Ustilago bromivora-Brachypodium Compatibility by Using a Hybrid Pathogen. Mol. Plant. Microbe Interact. 2019, 32, 1623–1634. [Google Scholar] [CrossRef] [Green Version]
- Lanver, D.; Müller, A.N.; Happel, P.; Schweizer, G.; Haas, F.B.; Franitza, M.; Pellegrin, C.; Reissmann, S.; Altmüller, J.; Rensing, S.A.; et al. The biotrophic development of Ustilago maydis studied by RNAseq analysis. Plant Cell 2018, 30, 300–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanver, D.; Tollot, M.; Schweizer, G.; Presti, L.L.; Reissmann, S.; Ma, L.; Schuster, M.; Tanaka, S.; Liang, L.; Ludwig, N.; et al. Ustilago maydis effectors and their impact on virulence. Nat. Publ. Gr. 2017, 15, 409–421. [Google Scholar] [CrossRef]
- Kamper, J.; Kahmann, R.; Bolker, M.; Ma, L.-J.; Brefort, T.; Saville, B.J.; Banuett, F.; Kronstad, J.; Gold, S.; Müller, O.; et al. Insights from the genome of the biotrophic fungal plant pathogen Ustilago maydis. Nature 2006, 444, 97–101. [Google Scholar] [CrossRef]
- Schirawski, J.; Mannhaupt, G.; Münch, K.; Brefort, T.; Schipper, K.; Doehlemann, G.; Di Stasio, M.; Rössel, N.; Mendoza-Mendoza, A.; Pester, D.; et al. Pathogenicity determinants in smut fungi revealed by genome comparison. Science 2010, 330, 1546–1548. [Google Scholar] [CrossRef]
- Brefort, T.; Doehlemann, G.; Mendoza-Mendoza, A.; Reissmann, S.; Djamei, A.; Kahmann, R. Ustilago maydis as a Pathogen. Annu. Rev. Phytopathol. 2009, 47, 423–445. [Google Scholar] [CrossRef] [Green Version]
- Christensen, J.J. Corn Smut Caused by Ustilago maydis (Monograph No. 2); American Phytopathological Society: St. Paul, MN, USA, 1963. [Google Scholar]
- Poloni, A.; Schirawski, J. Host specificity in Sporisorium reilianum is determined by distinct mechanisms in maize and sorghum. Mol. Plant Pathol. 2016, 17, 741–754. [Google Scholar] [CrossRef]
- Wollenberg, T.; Schirawski, J. Comparative Genomics of Plant Fungal Pathogens: The Ustilago-Sporisorium Paradigm. PLoS Pathog. 2014, 10, e1004218. [Google Scholar] [CrossRef] [Green Version]
- Holliday, R. The genetics of Ustilago maydis. Genet. Res. (Camb.) 1961, 2, 204–230. [Google Scholar] [CrossRef]
- Cheung, H.Y.K.; Donaldson, M.; Storfie, E.; Spence, K.; Fetsch, J.; Harrison, M.; Saville, B.J. Zfp1, a putative Zn(II)2Cys6 transcription factor, influences Ustilago maydis pathogenesis at multiple stages. Plant Pathol. 2021, 70, 1626–1639. [Google Scholar] [CrossRef]
- Morrison, E.N.; Donaldson, M.E.; Saville, B.J. Identification and analysis of genes expressed in the Ustilago maydis dikaryon: Uncovering a novel class of pathogenesis genes. Can. J. Plant Pathol. 2012, 34, 417–435. [Google Scholar] [CrossRef]
- Gold, S.E.; Brogdon, S.M.; Mayorga, M.E.; Kronstad, J.W. The Ustilago maydis regulatory subunit of a cAMP-dependent protein kinase is required for gall formation in maize. Plant Cell 1997, 9, 1585–1594. [Google Scholar] [CrossRef] [Green Version]
- Goulet, K.M.; Storfie, E.R.; Saville, B.J. Exploring links between antisense RNAs and pathogenesis in Ustilago maydis through transcript and gene characterization. Fungal Genet. Biol. 2020, 134, 103283. [Google Scholar] [CrossRef]
- ImageJ 1.53a. National Institutes of Health. Available online: https://imagej.nih.gov/ij/index.html (accessed on 30 July 2021).
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2-A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Sigrist, C.J.A.; De Castro, E.; Cerutti, L.; Cuche, B.A.; Hulo, N.; Bridge, A.; Bougueleret, L.; Xenarios, I. New and continuing developments at PROSITE. Nucleic Acids Res. 2013, 41, 344–347. [Google Scholar] [CrossRef] [Green Version]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Zahiri, A.R.; Babu, M.R.; Saville, B.J. Differential gene expression during teliospore germination in Ustilago maydis. Mol. Genet. Genom. 2005, 273, 394–403. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ho, E.; Cahill, M.; Saville, B.J. Gene discovery and transcript analyses in the corn smut pathogen Ustilago maydis: Expressed sequence tag and genome sequence comparison. BMC Genom. 2007, 8, 334. [Google Scholar] [CrossRef] [Green Version]
- Ho, E.; Donaldson, M.E.; Saville, B.J. Detection of Antisense RNA Transcripts by Strand-Specific RT-PCR. In Methods in Molecular Biology; King, N., Ed.; Springer Science + Business Media (Now Part of Springer Nature): New York, NY, USA, 2010; pp. 125–138. [Google Scholar]
- Wang, J.; Holden, D.; Leong, S. Gene transfer system for the phytopathogenic fungus Ustilago maydis. Proc. Natl. Acad. Sci. USA 1988, 85, 865–869. [Google Scholar] [CrossRef] [Green Version]
- Ostrowski, L.A.; Saville, B.J. Natural antisense transcripts to nucleus-encoded mitochondrial genes are linked to Ustilago maydis teliospore dormancy. Mol. Microbiol. 2016, 103, 745–763. [Google Scholar] [CrossRef] [Green Version]
- Yee, A.R. Specificity at the b Mating Type Locus of Ustilago maydis; University of British Columbia: Vancouver, BC, Cananda, 1998. [Google Scholar]
- Hoffman, C.; Winston, F. A ten-minute DNA preparation from yeast efficiently releases autonomous plasmids for transformation of Escherichia coli. Gene 1987, 57, 267–272. [Google Scholar] [CrossRef]
- Banuett, F.; Herskowitz, I. Morphological Transitions in the Life Cycle of Ustilago maydis and Their Genetic Control by the a and b Loci. Exp. Mycol. 1994, 18, 247–266. [Google Scholar] [CrossRef]
- Banuett, F.; Herskowitz, I. Discrete developmental stages during teliospore formation in the corn smut fungus, Ustilago maydis. Development 1996, 122, 2965–2976. [Google Scholar] [CrossRef]
- Tanaka, S.; Rössel, N.; Fukada, F.; Schweizer, G.; Thines, M.; Kahmann, R. Neofunctionalization of a secreted effector in a fungal pathogen. Nat. Microbiol. 2019, 4, 251–257. [Google Scholar] [CrossRef]
- Chalker-Scott, L. Environmental significance of anthocyanins in plant stress responses. Photochem. Photobiol. 1999, 70, 1–9. [Google Scholar] [CrossRef]
- Banuett, F. Genetics of Ustilago maydis, a fungal pathogen that induces tumors in maize. Annu. Rev. Genet. 1990, 29, 179–208. [Google Scholar] [CrossRef]
- Tanaka, S.; Brefort, T.; Neidig, N.; Djamei, A.; Kahnt, J.; Vermerris, W.; Koenig, S.; Feussner, K.; Feussner, I.; Kahmann, R. A secreted Ustilago maydis effector promotes virulence by targeting anthocyanin biosynthesis in maize. Elife 2014, 2014, 1–27. [Google Scholar] [CrossRef]
- Doehlemann, G.; Reissmann, S.; Aßmann, D.; Fleckenstein, M.; Kahmann, R. Two linked genes encoding a secreted effector and a membrane protein are essential for Ustilago maydis-induced tumour formation. Mol. Microbiol. 2011, 81, 751–766. [Google Scholar] [CrossRef]
- Seitner, D.; Uhse, S.; Gallei, M.; Djamei, A. The core effector Cce1 is required for early infection of maize by Ustilago maydis. Mol. Plant Pathol. 2018, 19, 2277–2287. [Google Scholar] [CrossRef] [Green Version]
- Mueller, A.N.; Ziemann, S.; Treitschke, S.; Aßmann, D.; Doehlemann, G. Compatibility in the Ustilago maydis-Maize Interaction Requires Inhibition of Host Cysteine Proteases by the Fungal Effector Pit2. PLoS Pathog. 2013, 9, e1003177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweizer, G.; Münch, K.; Mannhaupt, G.; Schirawski, J.; Kahmann, R.; Dutheil, J.Y. Positively Selected Effector Genes and Their Contribution to Virulence in the Smut Fungus Sporisorium reilianum. Genome Biol. Evol. 2018, 10, 629–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djamei, A.; Kahmann, R. Ustilago maydis: Dissecting the Molecular Interface between Pathogen and Plant. PLoS Pathog. 2012, 8, e1002955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heimel, K.; Scherer, M.; Vranes, M.; Wahl, R.; Pothiratana, C. The Transcription Factor Rbf1 Is the Master Regulator for b-Mating Type Controlled Pathogenic Development in Ustilago maydis. PLoS Pathog. 2010, 6, e1001035. [Google Scholar] [CrossRef] [Green Version]
- Castanheira, S.; Mielnichuk, N.; Pérez-Martín, J. Programmed cell cycle arrest is required for infection of corn plants by the fungus Ustilago maydis. Dev. 2014, 141, 4817–4826. [Google Scholar] [CrossRef] [Green Version]
- Lanver, D.; Berndt, P.; Tollot, M.; Naik, V.; Vranes, M.; Warmann, T.; Münch, K.; Rössel, N.; Kahmann, R. Plant Surface Cues Prime Ustilago maydis for Biotrophic Development. PLoS Pathog. 2014, 10, e1004272. [Google Scholar] [CrossRef]
- Mendoza-Mendoza, A.; Berndt, P.; Djamei, A.; Weise, C.; Linne, U.; Marahiel, M.; Vraneš, M.; Kämper, J.; Kahmann, R. Physical-chemical plant-derived signals induce differentiation in Ustilago maydis. Mol. Microbiol. 2009, 71, 895–911. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, M.E.; Ostrowski, L.A.; Goulet, K.M.; Saville, B.J. Transcriptome analysis of smut fungi reveals widespread intergenic transcription and conserved antisense transcript expression. BMC Genom. 2017, 18, 340. [Google Scholar] [CrossRef] [PubMed]
- Doehlemann, G.; Wahl, R.; Horst, R.J.; Voll, L.M.; Usadel, B.; Poree, F.; Stitt, M.; Pons-Kühnemann, J.; Sonnewald, U.; Kahmann, R.; et al. Reprogramming a maize plant: Transcriptional and metabolic changes induced by the fungal biotroph Ustilago maydis. Plant J. 2008, 56, 181–195. [Google Scholar] [CrossRef]
- Doehlemann, G.; Wahl, R.; Vranes, M.; de Vries, R.P.; Kämper, J.; Kahmann, R. Establishment of compatibility in the Ustilago maydis/maize pathosystem. J. Plant Physiol. 2008, 165, 29–40. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Haploid Strain a | Relevant Genotype | Source |
|---|---|---|
| 518 | a2b2 | Holliday [32] |
| 521 b | a1b1 | Holliday [32] |
| SRZ1 | a1b1 | Schirawski et al. [19] |
| SRZ2 b | a2b2 | Schirawski et al. [19] |
| 521 Potef:rbf1SI | a1b1 ipR[Potef:rbf1SI]ipS | This Study |
| 521 Potef:rbf1MI | a1b1 ipR[Potef:rbf1MI]ipS | This Study |
| 521 Potef:hdp2SI | a1b1 ipR[Potef:hdp2SI]ipS | This Study |
| 521 Potef:hdp2MI | a1b1 ipR[Potef:hdp2MI]ipS | This Study |
| 521 Potef:nlt1SI | a1b1 ipR[Potef:nlt1SI]ipS | Meade et al. [unpublished] This Study |
| 521 Potef:nlt1MI | a1b1 ipR[Potef:nlt1MI]ipS | Meade et al. [unpublished] This Study |
| 521 Potef:tin2SI | a1b1 ipR[Potef:tin2SI]ipS | Cheung et al. [33] This Study |
| 521 Potef:tin2MI | a1b1 ipR[Potef:tin2MI]ipS | Cheung et al. [33] This Study |
| 521 Potef:pit2SI | a1b1 ipR[Potef:pit2SI]ipS | This Study |
| 521 Potef:pit2MI | a1b1 ipR[Potef:pit2MI]ipS | This Study |
| 521 Potef:cce1SI | a1b1 ipR[Potef:cce1SI]ipS | This Study |
| 521 Potef:cce1MI | a1b1 ipR[Potef:cce1MI]ipS | This Study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Storfie, E.R.M.; Saville, B.J. Fungal Pathogen Emergence: Investigations with an Ustilago maydis × Sporisorium reilianum Hybrid. J. Fungi 2021, 7, 672. https://doi.org/10.3390/jof7080672
Storfie ERM, Saville BJ. Fungal Pathogen Emergence: Investigations with an Ustilago maydis × Sporisorium reilianum Hybrid. Journal of Fungi. 2021; 7(8):672. https://doi.org/10.3390/jof7080672
Chicago/Turabian StyleStorfie, Emilee R. M., and Barry J. Saville. 2021. "Fungal Pathogen Emergence: Investigations with an Ustilago maydis × Sporisorium reilianum Hybrid" Journal of Fungi 7, no. 8: 672. https://doi.org/10.3390/jof7080672