
Culturable Fungal Community of Pterocladiella capillacea in Keelung, Taiwan: Effects of Surface Sterilization Method and Isolation Medium


Abstract
:1. Introduction
2. Materials and Methods
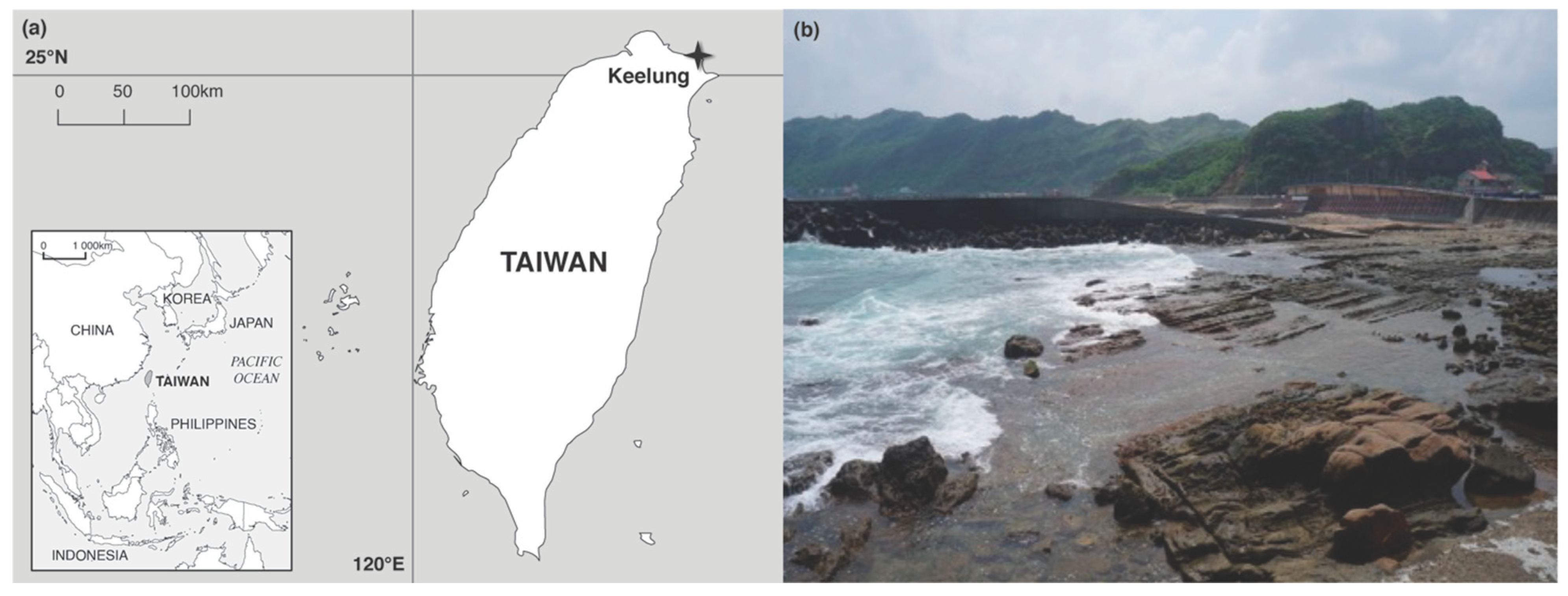
2.1. Collection of Samples
2.2. Surface Washing/Sterilization of Pterocladiella Capillacea
2.3. Fungal Isolation
2.4. Fungal Identification
2.5. Statistics
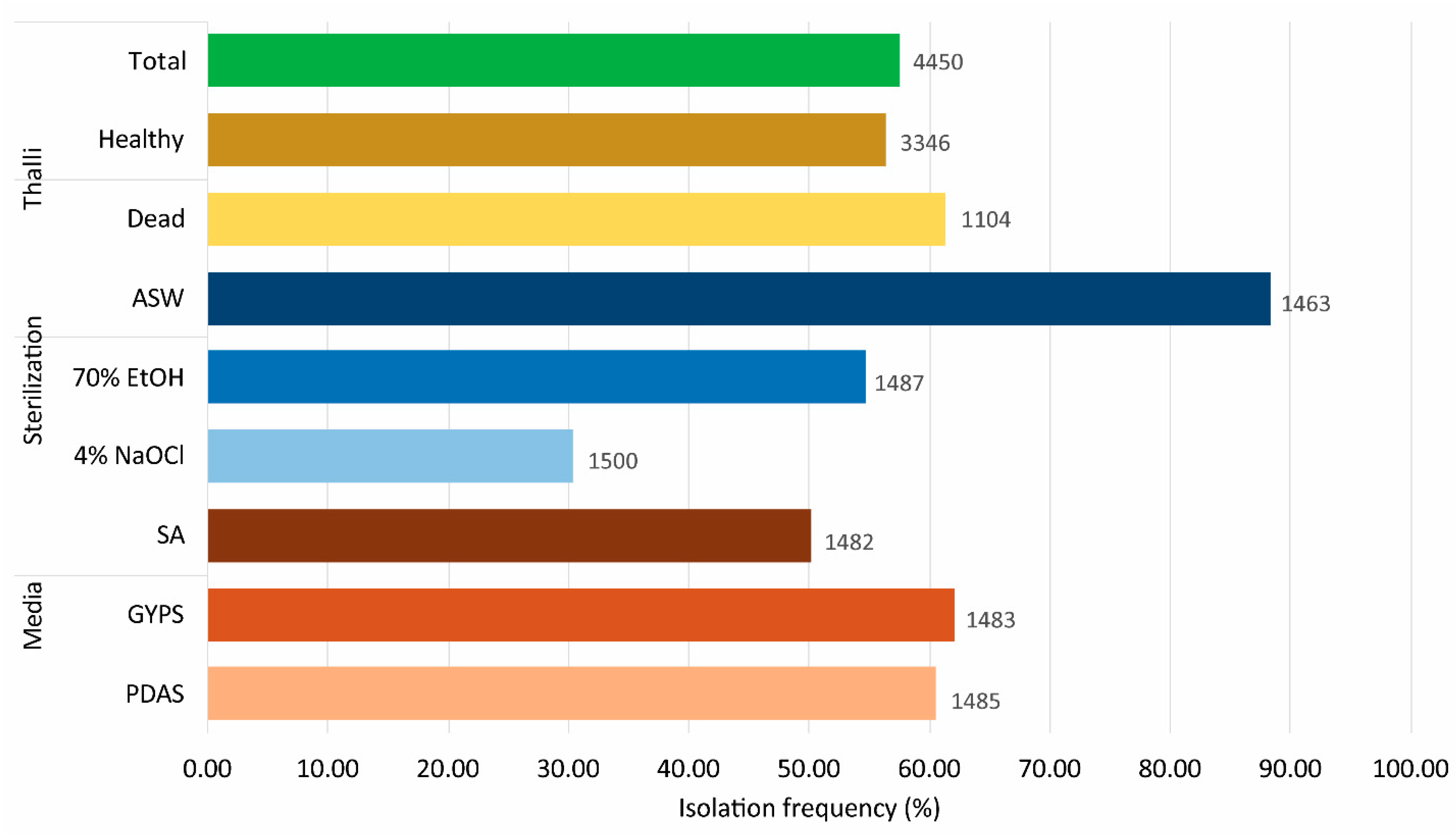
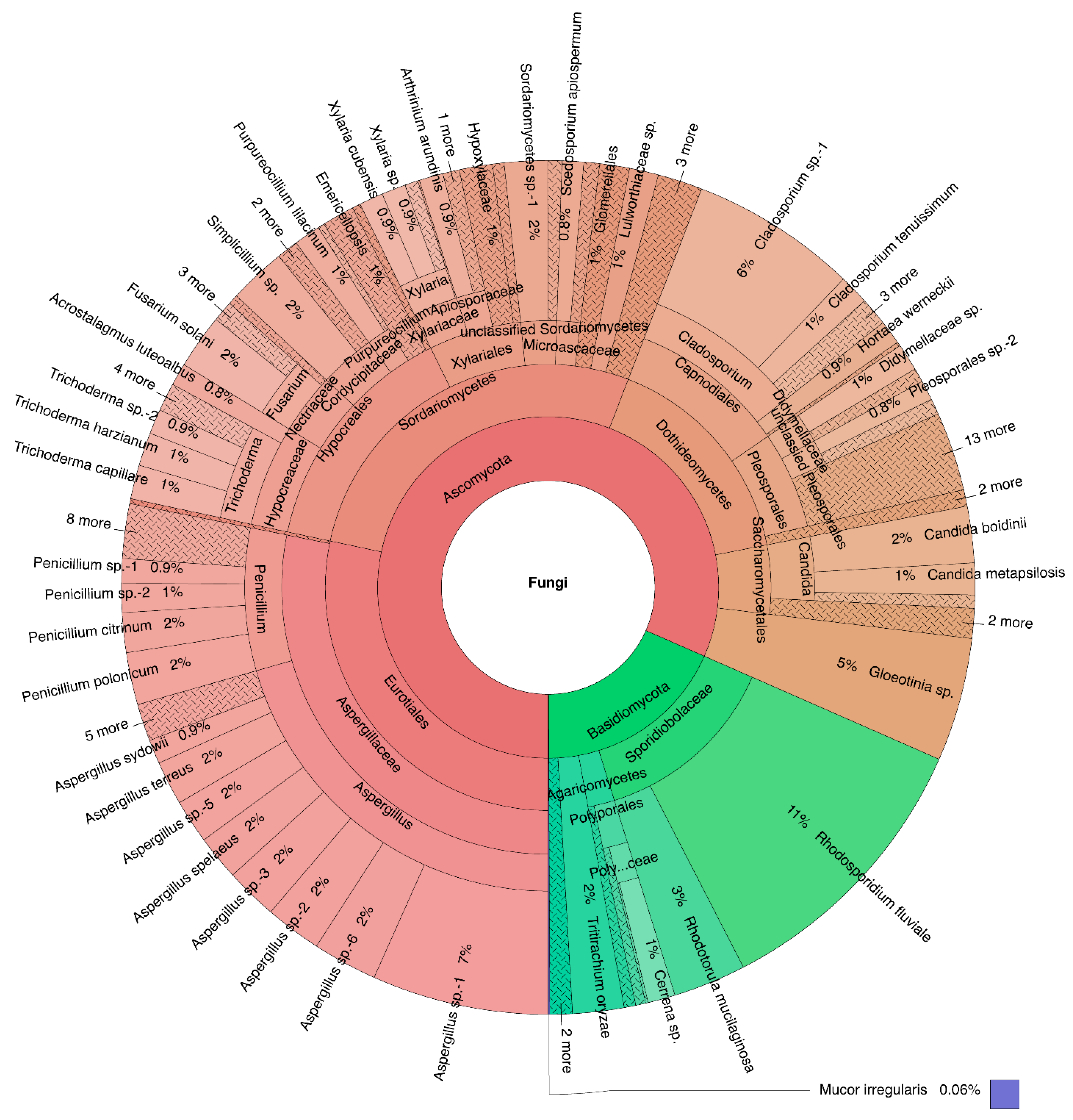
3. Results
3.1. Culturable Fungal Diversity of Pterocladiella Capillacea
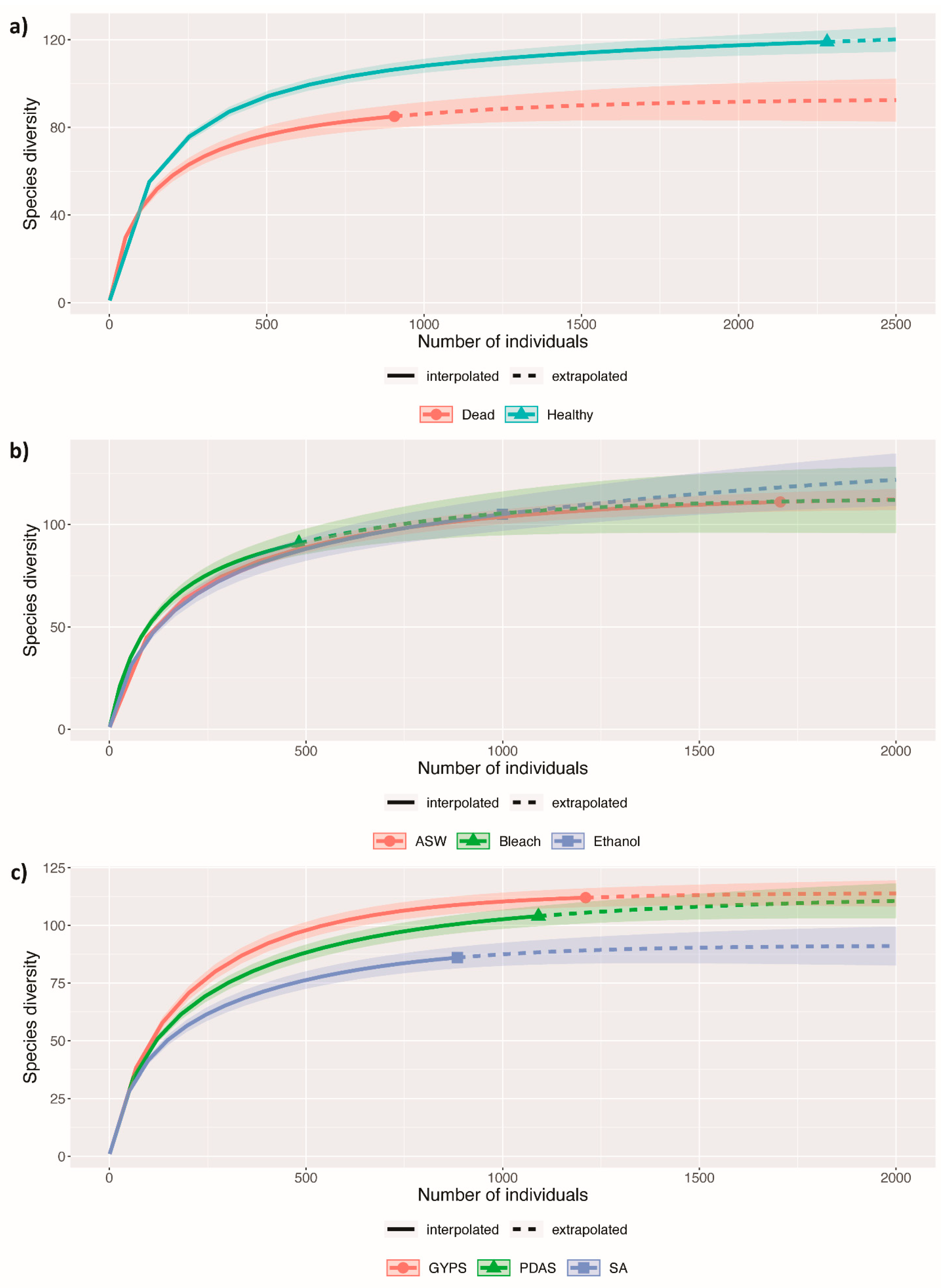
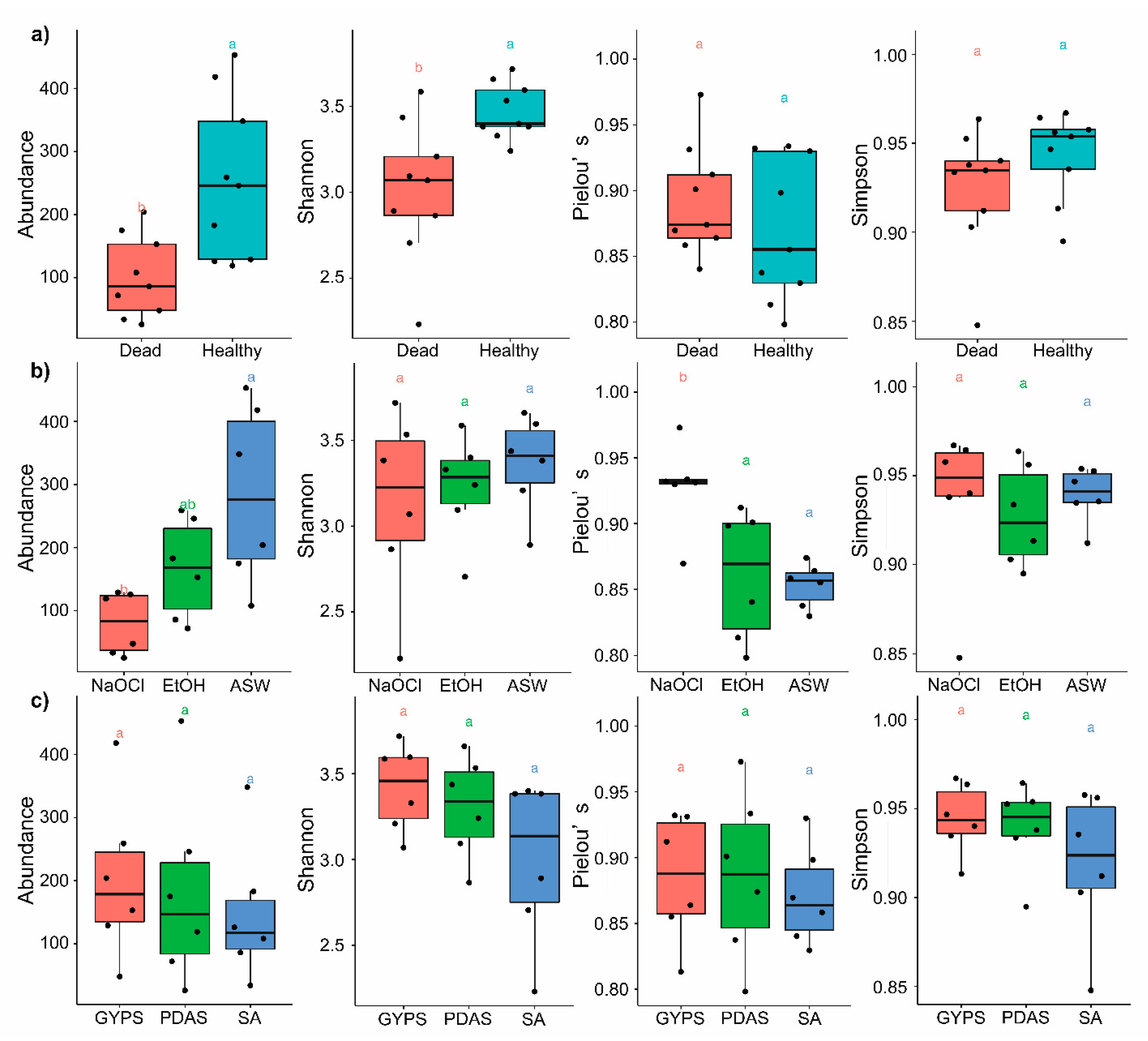
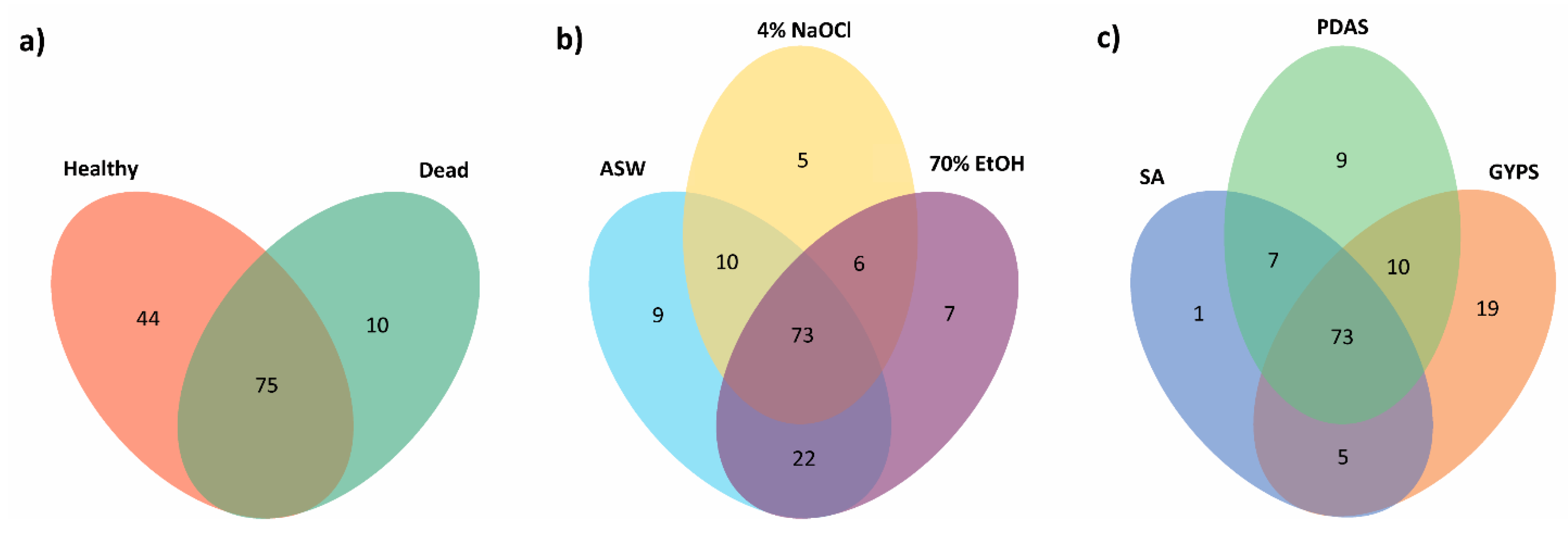
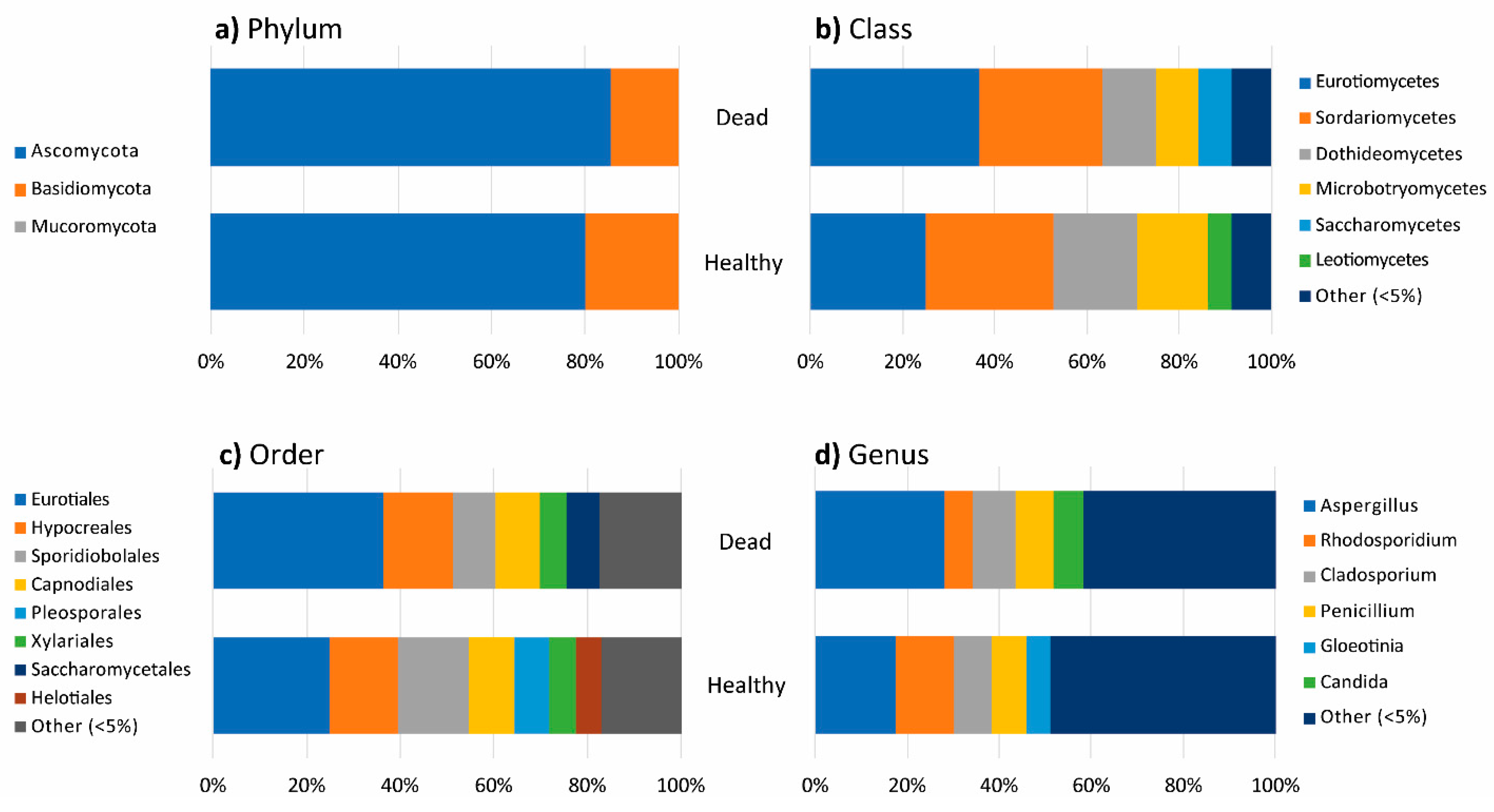
3.2. Fungal Diversity in Healthy and Dead Thalli
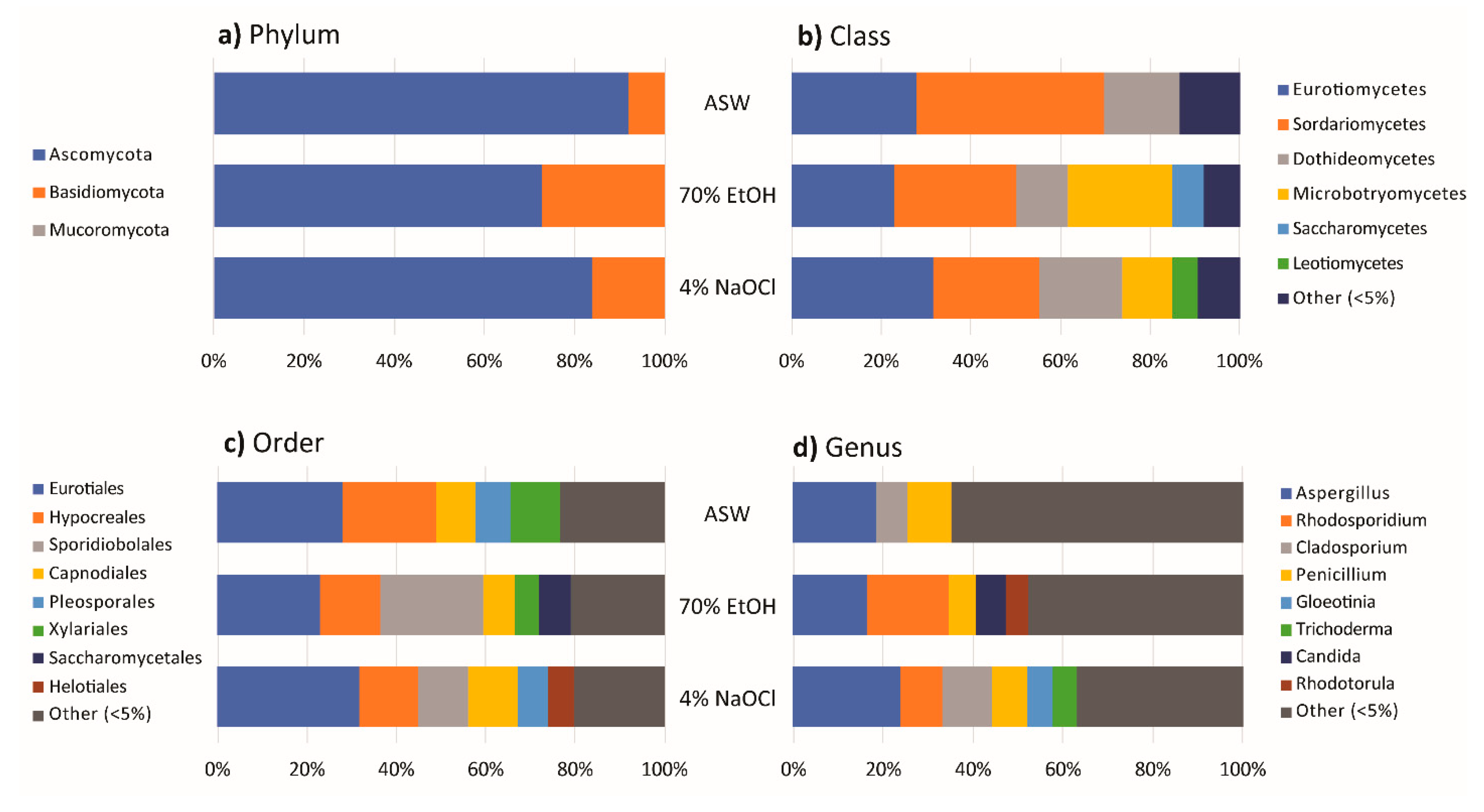
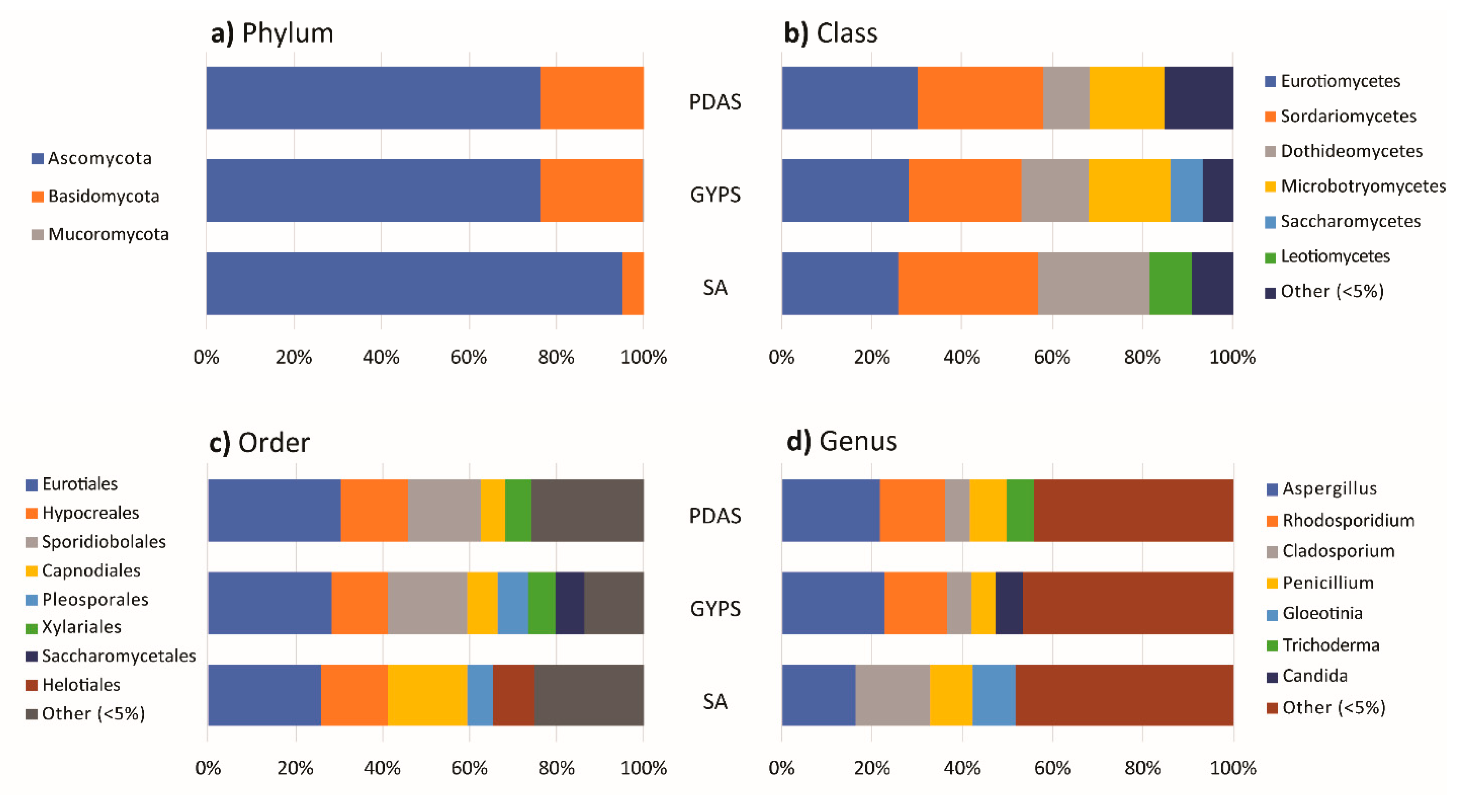
3.3. Effect of Algal Surface Washing/Sterilization Methods on Fungal Diversity
3.4. Effect of Culture Media on Fungal Diversity
4. Discussion
4.1. Fungal Community
4.2. Isolation Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sutherland, G. New marine fungi on Pelvetia. New Phytol. 1915, 14, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, G. Marine fungi imperfecti. New Phytol. 1916, 15, 35–48. [Google Scholar] [CrossRef]
- Vrijmoed, L.L.P. Isolation and culture of higher filamentous fungi. In Marine Mycology—A Practical Approach; Hyde, K.D., Pointing, S.B., Eds.; Fungal Diversity Press: Hong Kong, China, 2000; pp. 1–20. [Google Scholar]
- Zuccaro, A.; Mitchell, J.I. Fungal communities of seaweeds. In The Fungal Community: Its Organisation and Role in the Ecosystem; Dighton, J., Oudemans, P., White, J., Eds.; Marcel Dekker: New York, NY, USA, 2005; pp. 553–579. [Google Scholar]
- Jones, E.B.G.; Pang, K.L.; Stanley, S.J. Fungi from marine algae. In Marine Fungi and Fungal-Like Organisms; Jones, E.B.G., Pang, K.L., Eds.; De Gruyter: Berlin, Germany, 2012; pp. 329–344. [Google Scholar]
- Porter, D.; Farnham, W.F. Mycaureola dilseae, a marine basidiomycete parasite of the red alga, Dilsea carnosa. Trans. Br. Mycol. Soc. 1986, 87, 575–582. [Google Scholar] [CrossRef]
- Stanley, S.J. Observations on the seasonal occurrence of marine endophytic and parasitic fungi. Can. J. Bot. 1992, 70, 2089–2096. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Bayman, P. Hidden fungi, emergent properties: Endophytes and microbiomes. Annu. Rev. Phytopathol. 2011, 49, 291–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debbab, A.; Aly, A.; Proksch, P. Endophytes and associated marine derived fungi—ecological and chemical perspectives. Fungal Divers. 2012, 57, 45–83. [Google Scholar] [CrossRef]
- Sanders, W.B.; Moe, R.L.; Ascaso, C. The intertidal marine lichen formed by the pyrenomycete fungus Verrucaria tavaresiae (Ascomycotina) and the brown alga Petroderma maculiforme (Phaeophyceae): Thallus organization and symbiont interaction. Am. J. Bot. 2004, 91, 511–522. [Google Scholar] [CrossRef] [Green Version]
- Kohlmeyer, J.; Volkmann-Kohlmeyer, B. Mycophycias, a new genus for the mycobionts of Apophlaea, Ascophyllum and Pelvetia. Syst. Ascomycetum 1998, 16, 1–7. [Google Scholar]
- Zuccaro, A.; Schoch, C.L.; Spatafora, J.W.; Kohlmeyer, J.; Draeger, S.; Mitchell, J.I. Detection and identification of fungi intimately associated with the brown seaweed Fucus serratus. Appl. Environ. Microbiol. 2008, 74, 931–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bugni, T.S.; Ireland, C.M. Marine-derived fungi: A chemically and biologically diverse group of microorganisms. Nat. Prod. Rep. 2004, 21, 143–163. [Google Scholar] [CrossRef]
- Kjer, J.; Debbab, A.; Aly, A.H.; Proksch, P. Methods for isolation of marine-derived endophytic fungi and their bioactive secondary products. Nat. Protoc. 2010, 5, 479–490. [Google Scholar] [CrossRef]
- Loque, C.P.; Medeiros, A.O.; Pellizzari, F.M.; Oliveira, E.C.; Rosa, C.A.; Rosa, L.H. Fungal community associated with marine macroalgae from Antarctica. Polar Biol. 2010, 33, 641–648. [Google Scholar] [CrossRef]
- Godinho, V.M.; Furbino, L.E.; Santiago, I.F.; Pellizzari, F.M.; Yokoya, N.S.; Pupo, D.; Alves, T.M.; Junior, P.A.; Romanha, A.J.; Zani, C.L.; et al. Diversity and bioprospecting of fungal communities associated with endemic and cold-adapted macroalgae in Antarctica. ISME J. 2013, 7, 1434–1451. [Google Scholar] [CrossRef]
- Duarte, A.W.F.; Dayo-Owoyemi, I.; Nobre, F.S.; Pagnocca, F.C.; Chaud, L.C.S.; Pessoa, A.; Felipe, M.G.A.; Sette, L.D. Taxonomic assessment and enzymes production by yeasts isolated from marine and terrestrial Antarctic samples. Extremophiles 2013, 17, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Solis, M.J.L.; Draeger, S.; dela Cruz, T.E.E. Marine-derived fungi from Kappaphycus alvarezii and K. striatum as potential causative agents of ice-ice disease in farmed seaweeds. Bot. Mar. 2010, 53, 587–594. [Google Scholar] [CrossRef]
- Abdel-Gawad, K.M.; Hifney, A.F.; Issa, A.A.; Gomaa, M. Spatio-temporal, environmental factors, and host identity shape culturable-epibiotic fungi of seaweeds in the Red Sea, Egypt. Hydrobiologia 2014, 740, 37–49. [Google Scholar] [CrossRef]
- Zuccaro, A.; Summerbell, R.C.; Gams, W.; Schroers, H.J.; Mitchell, J.I. A new Acremonium species associated with Fucus spp., and its affinity with a phylogenetically distinct marine Emericellopsis clade. Stud. Mycol. 2004, 50, 283–297. [Google Scholar]
- Flewelling, A.J.; Ellsworth, K.T.; Sanford, J.; Forward, E.; Johnson, J.A.; Gray, C.A. Macroalgal Endophytes from the Atlantic coast of Canada: A potential source of antibiotic natural products? Microorganisms 2013, 1, 175–187. [Google Scholar] [CrossRef]
- Flewelling, A.J.; Johnson, J.A.; Gray, C.A. Isolation and bioassay screening of fungal endophytes from North Atlantic marine macroalgae. Bot. Mar. 2013, 56, 287–297. [Google Scholar] [CrossRef]
- Suryanarayanan, T.S.; Venkatachalam, A.; Thirunavukkarasu, N.; Ravishankar, J.P.; Doble, M.; Geetha, V. Internal muconiota of marine macroalgae from the Tamilnadu coast: Distribution, diversity and biotechnological potential. Bot. Mar. 2010, 53, 457–468. [Google Scholar] [CrossRef]
- Kohlmeyer, J.; Volkmann-Kohlmeyer, B. Marine ascomycetes from algae and animal hosts. Bot. Mar. 2003, 46, 285–306. [Google Scholar] [CrossRef]
- Zuccaro, A.; Schulz, B.; Mitchell, J.I. Molecular detection of ascomycetes associated with Fucus serratus. Mycol. Res. 2003, 107, 1451–1466. [Google Scholar] [CrossRef] [PubMed]
- Kohlmeyer, J.; Kohlmeyer, E. Marine Mycology: The Higher Fungi; Academic Press: New York, NY, USA, 1979. [Google Scholar]
- Kohlmeyer, J.; Volkmann-Kohlmeyer, B. Illustrated key to the filamentous marine fungi. Bot. Mar. 1991, 34, 1–61. [Google Scholar] [CrossRef]
- Wainwright, B.J.; Zahn, G.L.; Zushi, J.; Lee, N.L.Y.; Ooi, J.L.S.; Lee, J.N.; Huang, D. Seagrass-associated fungal communities show distance decay of similarity that has implications for seagrass management and restoration. Ecol. Evol. 2019, 9, 11288–11297. [Google Scholar] [CrossRef] [PubMed]
- Guiry, M.D. How many species of Algae are there? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef]
- Teixeira, T.R.; Santos, G.S.; Turatti, I.C.C.; Paziani, M.H.; von Zeska Kress, M.R.; Colepicolo, P.; Debonsi, H.M. Characterization of the lipid profile of Antarctic brown seaweeds and their endophytic fungi by gas chromatography–mass spectrometry (GC–MS). Polar Biol. 2019, 42, 1431–1444. [Google Scholar] [CrossRef]
- Sieber, T.N. Fungal root endophytes. In Plant Roots: The Hidden Half; Waisel, Y., Eshel, A., Kafkafi, U., Eds.; Marcel Dekker: New York, NY, USA, 2002; pp. 887–917. [Google Scholar]
- Wong Chin, J.M.; Puchooa, D.; Bahorun, T.; Jeewon, R. Antimicrobial properties of marine fungi from sponges and brown algae of Mauritius. Mycology 2021, 12, 1–14. [Google Scholar] [CrossRef]
- Deutsch, Y.; Gur, L.; Berman Frank, I.; Ezra, D. Endophytes from Algae, a Potential Source for New Biologically Active Metabolites for Disease Management in Aquaculture. Front. Mar. Sci. 2021, 8, 333. [Google Scholar] [CrossRef]
- Kamat, S.; Kumari, M.; Taritla, S.; Jayabaskaran, C. Endophytic fungi of marine alga from Konkan coast, India—A rich source of bioactive material. Front. Mar. Sci. 2020, 7, 31. [Google Scholar] [CrossRef]
- Lee, S.; Park, M.S.; Lee, H.; Kim, J.J.; Eimes, J.A.; Lim, Y.W. Fungal diversity and enzyme activity associated with the macroalgae, Agarum clathratum. Mycobiology 2019, 47, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Gnavi, G.; Garzoli, L.; Poli, A.; Prigione, V.; Burgaud, G.; Varese, G.C. The culturable mycobiota of Flabellia petiolata: First survey of marine fungi associated to a Mediterranean green alga. PLoS ONE 2017, 12, e0175941. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-coverage ITS primers for the DNA-based identification of ascomycetes and basidiomycetes in environmental samples. PLoS ONE 2012, 7, e40863. [Google Scholar]
- Beenken, L.; Lutz, M.; Scholler, M. DNA barcoding and phylogenetic analyses of the genus Coleosporium (Pucciniales) reveal that the North American goldenrod rust C. solidaginis is a neomycete on introduced and native Solidago species in Europe. Mycol. Prog. 2017, 16, 1073–1085. [Google Scholar] [CrossRef]
- Petrini, O.; Fisher, P.J.; Petrini, L.E. Fungal endophytes of bracken (Pteridium aquilinum), with some reflections on their use in biological control. Sydowia 1992, 44, 282–293. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; The University of Illinois Press: Urbana, IL, USA, 1949; pp. 3–24. [Google Scholar]
- Margalef, R. Perspectives in Ecological Theory; University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for interpolation and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Ondov, B.D.; Nicholas, H.B.; Adam, M.P. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. Jvenn: An interactive Venn diagram viewer. BMC Bioinform. 2014, 15, 293. [Google Scholar] [CrossRef] [Green Version]
- Gunde-Cimerman, N.; Ramos, J.; Plemenitaš, A. Halotolerant and halophilic fungi. Mycol. Res. 2009, 113, 1231–1241. [Google Scholar] [CrossRef]
- Becker, K.; Stadler, M. Recent progress in biodiversity research on the Xylariales and their secondary metabolism. J. Antibiot. 2021, 74, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Sakayaroj, J.; Preedanon, S.; Phongpaichit, S.; Buatong, J.; Chaowalit, P.; Rukachaisirikul, V. 16 Diversity of endophytic and marine-derived fungi associated with marine plants and animals. In Marine Fungi; Gareth Jones, E.B., Pang, K.-L., Eds.; De Gruyter: Berlin, Germany, 2012; pp. 291–328. [Google Scholar]
- De Felício, R.; Pavão, G.B.; de Oliveira, A.L.L.; Erbert, C.; Conti, R.; Pupo, M.T.; Furtado, N.A.; Ferreira, E.G.; Costa-Lotufo, L.V.; Young, M.C.M.; et al. Antibacterial, antifungal and cytotoxic activities exhibited by endophytic fungi from the Brazilian marine red alga Bostrychia tenella (Ceramiales). Rev. Bras. Farmacogn. 2015, 25, 641–650. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.S.; Wang, S.W.; Wang, G.J.; Pang, K.L.; Lee, C.K.; Kuo, Y.H.; Cha, H.J.; Lin, R.K.; Lee, T.H. Angiogenesis inhibitors and anti-inflammatory agents from Phoma sp. NTOU4195. J. Nat. Prod. 2016, 79, 2983–2990. [Google Scholar] [CrossRef]
- Hsiao, G.; Chi, W.C.; Pang, K.L.; Chen, J.J.; Kuo, Y.H.; Wang, Y.K.; Cha, H.J.; Chou, S.C.; Lee, T.H. Hirsutane-type sesquiterpenes with inhibitory activity of microglial nitric oxide production from the red alga derived fungus Chondrostereum sp. NTOU4196. J. Nat. Prod. 2017, 80, 1615–1622. [Google Scholar] [CrossRef] [PubMed]
- Alva, P.; McKenzie, E.H.C.; Pointing, S.B.; Pena-Muralla, R.; Hyde, K.D. Do sea grasses harbour endophytes? Fungal Divers. Res. Ser. 2002, 7, 167–178. [Google Scholar]
- Jones, E.B.G.; Suetrong, S.; Sakayaroj, J.; Bahkali, A.H.; Abdel-Wahab, M.A.; Boekhout, T.; Pang, K.L. Classification of marine Ascomycota, Basidiomycota, Blastocladiomycota and Chytridiomycota. Fungal Divers. 2015, 73, 1–72. [Google Scholar] [CrossRef]
- Hyde, K.D.; Soytong, K. Understanding microfungal diversity: A critique. Cyptogamie Mycologie 2007, 28, 281–289. [Google Scholar]
- Das, S.; Lyla, P.S.; Khan, S.A. Filamentous fungal population and species diversity from the continental slope of Bay of Bengal, India. Acta Oecol. 2009, 35, 269–279. [Google Scholar] [CrossRef]
- Matallah-Boutiba, A.; Ruiz, N.; Sallenave-Namont, C.; Grovel, O.; Amiard, J.C.; Pouchus, Y.F.; Boutiba, Z. Screening for toxigenic marine-derived fungi in Algerian mussels and their immediate environment. Aquaculture 2012, 342, 75–79. [Google Scholar] [CrossRef]
- König, G.M.; Kehraus, S.; Seibert, S.F.; Abdel-Lateff, A.; Müller, D. Natural products from marine organisms and their associated microbes. ChemBioChem 2006, 7, 229–238. [Google Scholar] [CrossRef]
- Sarasan, M.; Job, N.; Puthumana, J.; Ravinesh, R.; Prabhakaran, M.P.; Thomas, L.C.; Philip, R. Exploration and profiling of hidden endophytic mycota of marine macroalgae with potential drug leads. FEMS Microbiol. Lett. 2020, 367, fnaa078. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, A.; Thirunavukkarasu, N.; Suryanarayanan, T.S. Distribution and diversity of endophytes in seagrasses. Fungal Ecol. 2015, 13, 60–65. [Google Scholar] [CrossRef]
- Höller, U.; Wright, A.D.; Matthee, G.F.; Konig, G.M.; Draeger, S.; Hans-Jürgen, A.U.S.T.; Schulz, B. Fungi from marine sponges: Diversity, biological activity and secondary metabolites. Mycol. Res. 2000, 104, 1354–1365. [Google Scholar] [CrossRef]
- Baker, P.W.; Kennedy, J.; Dobson, A.D.; Marchesi, J.R. Phylogenetic diversity and antimicrobial activities of fungi associated with Haliclona simulans isolated from Irish coastal waters. Mar. Biotechnol. 2009, 11, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Pang, K.L.; Chiang, M.W.-L.; Guo, S.-Y.; Shih, C.-Y.; Dahms, H.U.; Hwang, J.-S.; Cha, H.-J. Growth study under combined effects of temperature, pH and salinity and transcriptome analysis revealed adaptations of Aspergillus terreus NTOU4989 to the extreme conditions at Kueishan Island Hydrothermal Vent Field, Taiwan. PLoS ONE 2020, 15, e0233621. [Google Scholar] [CrossRef]
- Furbino, L.; Pellizzari, F.M.; Neto, P.C.; Rosa, C.A.; Rosa, L.H. Isolation of fungi associated with macroalgae from maritime Antarctica and their production of agarolytic and carrageenolytic activities. Polar Biol. 2017, 41, 527–535. [Google Scholar] [CrossRef]
- Garzoli, L.; Gnavi, G.; Varese, G.C.; Picco, A.M. Mycobiota associated with the rhodophyte alien species Asparagopsis taxiformis (Delile) Trevisan de Saint-Léon in the Mediterranean Sea. Mar. Ecol. 2015, 36, 959–968. [Google Scholar] [CrossRef]
- Fell, J.W.; Boekhout, T.; Fonseca, A.; Scorzetti, G.; Statzell-Tallman, A. Biodiversity and systematics of basidiomycetous yeasts as determined by large subunit rDNA D1/D2 domain sequence analysis. Int. J. Syst. Evol. Microbiol. 2000, 50, 1351–1371. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Choeyklin, R. Ecology of marine and freshwater basidiomycetes. In Ecology of Saprotrophic Basidiomycetes; Boddy, L., Frankland, J.C., van West, P., Eds.; Elsevier: London, UK, 2008; pp. 301–324. [Google Scholar]
- Pang, K.L.; Guo, S.Y.; Chen, I.A.; Burgaud, G.; Luo, Z.H.; Dahms, H.U.; Hwang, J.S.; Lin, Y.L.; Huang, J.S.; Ho, T.W.; et al. Insights into fungal diversity of a shallow-water hydrothermal vent field at Kueishan Island, Taiwan by culture-based and metabarcoding analyses. PLoS ONE 2019, 14, e0226616. [Google Scholar]
- Zaky, A.S.; Tucker, G.A.; Daw, Z.Y.; Du, C. Marine yeast isolation and industrial application. FEMS Yeast Res. 2014, 14, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.W.; Sparrow, F.K. Fungi in Oceans and Estuaries; Cramer: Weinheim, Germany, 1961. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| . | Thalli | Surface Washing/Sterilization | Medium | Total | |||||
|---|---|---|---|---|---|---|---|---|---|
| Healthy | Dead | ASW | 70% EtOH | 4% NaOCl | SA | GYPS | PDAS | ||
| Total No. of isolates (Total Abundance), N | 2281 | 906 | 1706 | 999 | 482 | 885 | 1211 | 1091 | 3187 |
| Richness (Total number of Taxa in the community), S | 119 | 85 | 111 | 105 | 91 | 86 | 112 | 104 | 129 |
| Species Richness (Margalef): d | 15.2605 | 12.3365 | 14.7812 | 15.0577 | 14.5680 | 12.5265 | 15.6356 | 14.7251 | 15.8674 |
| Shannon-Wiener Diversity Index: H′ | 4.0335 | 3.8510 | 3.9623 | 3.8180 | 4.0923 | 3.7189 | 4.0107 | 3.8901 | 4.0982 |
| Pielou’s Evenness: J′ | 0.8440 | 0.8668 | 0.8413 | 0.8204 | 0.9072 | 0.8349 | 0.8500 | 0.8376 | 0.8433 |
| Simpson’s Dominance Index: D | 0.0349 | 0.0312 | 0.0342 | 0.0489 | 0.0233 | 0.0450 | 0.0379 | 0.0408 | 0.0310 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cha, H.-J.; Chiang, M.W.L.; Guo, S.-Y.; Lin, S.-M.; Pang, K.-L. Culturable Fungal Community of Pterocladiella capillacea in Keelung, Taiwan: Effects of Surface Sterilization Method and Isolation Medium. J. Fungi 2021, 7, 651. https://doi.org/10.3390/jof7080651
Cha H-J, Chiang MWL, Guo S-Y, Lin S-M, Pang K-L. Culturable Fungal Community of Pterocladiella capillacea in Keelung, Taiwan: Effects of Surface Sterilization Method and Isolation Medium. Journal of Fungi. 2021; 7(8):651. https://doi.org/10.3390/jof7080651
Chicago/Turabian StyleCha, Hyo-Jung, Michael W. L. Chiang, Sheng-Yu Guo, Showe-Mei Lin, and Ka-Lai Pang. 2021. "Culturable Fungal Community of Pterocladiella capillacea in Keelung, Taiwan: Effects of Surface Sterilization Method and Isolation Medium" Journal of Fungi 7, no. 8: 651. https://doi.org/10.3390/jof7080651