
Ether Oxidation by an Evolved Fungal Heme-Peroxygenase: Insights into Substrate Recognition and Reactivity


Abstract
:1. Introduction
2. Experimental Procedure
2.1. Chemicals and Reagents
2.2. Enzyme Reactions
2.3. Enzyme Kinetics
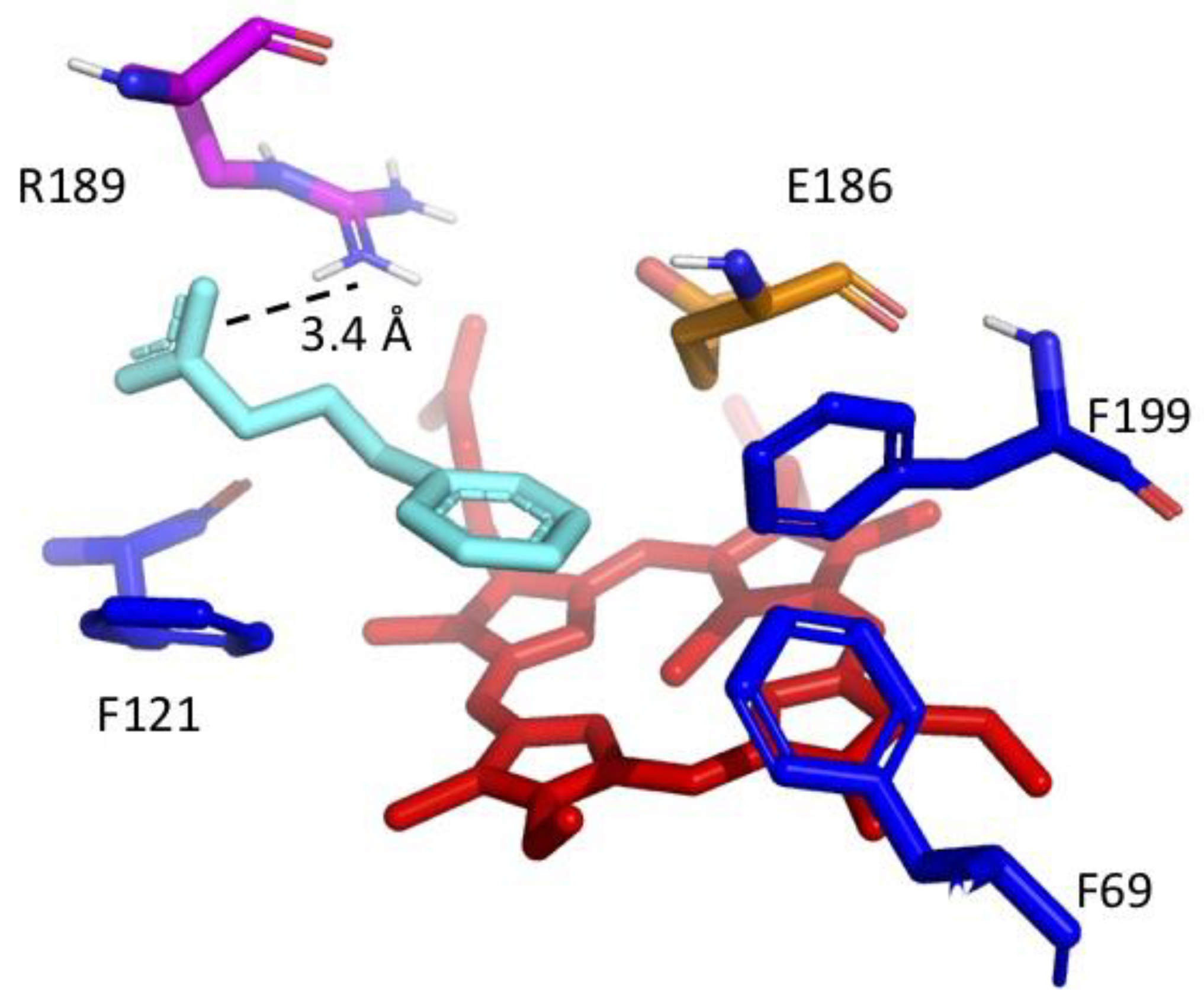
2.4. Molecular Docking
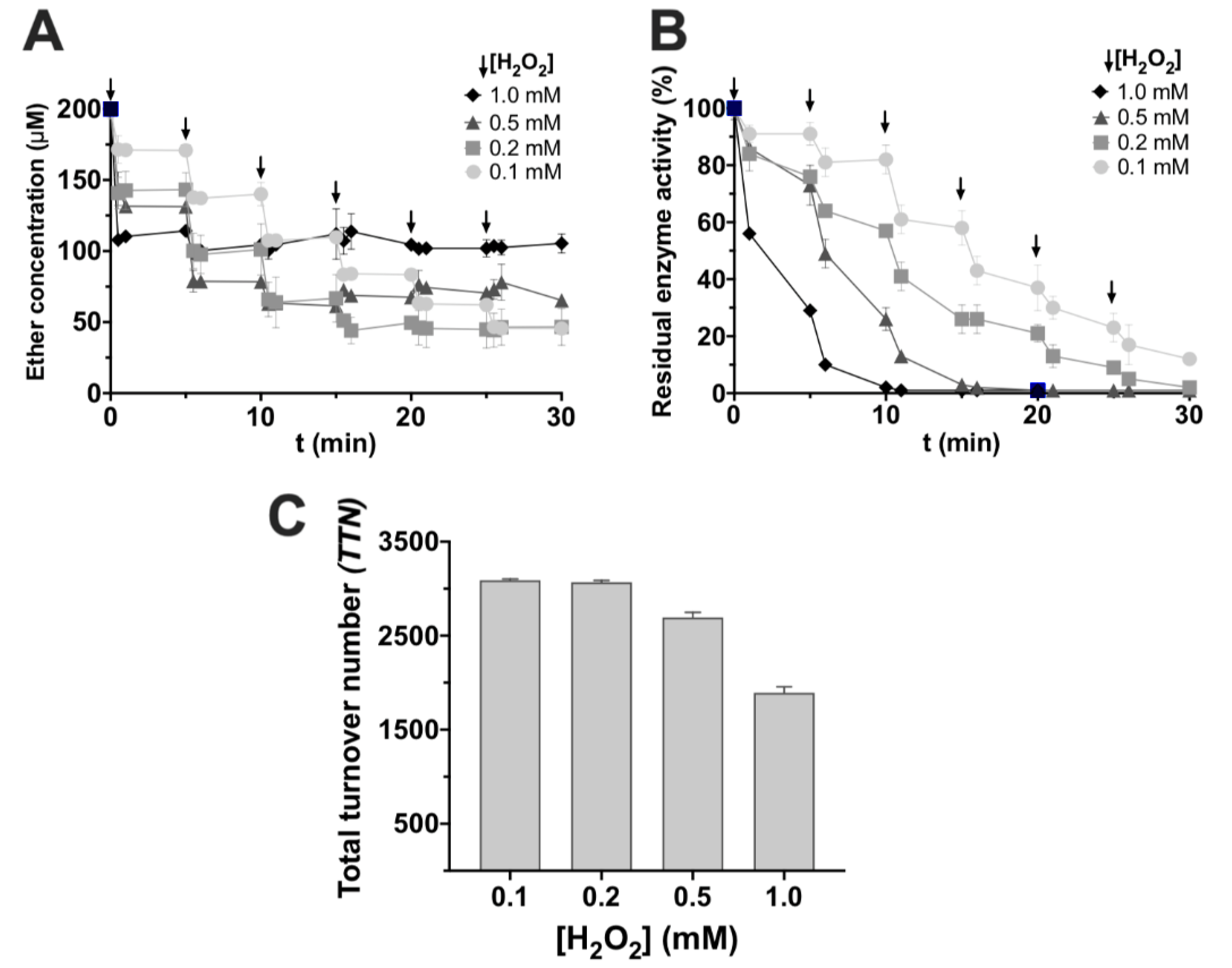
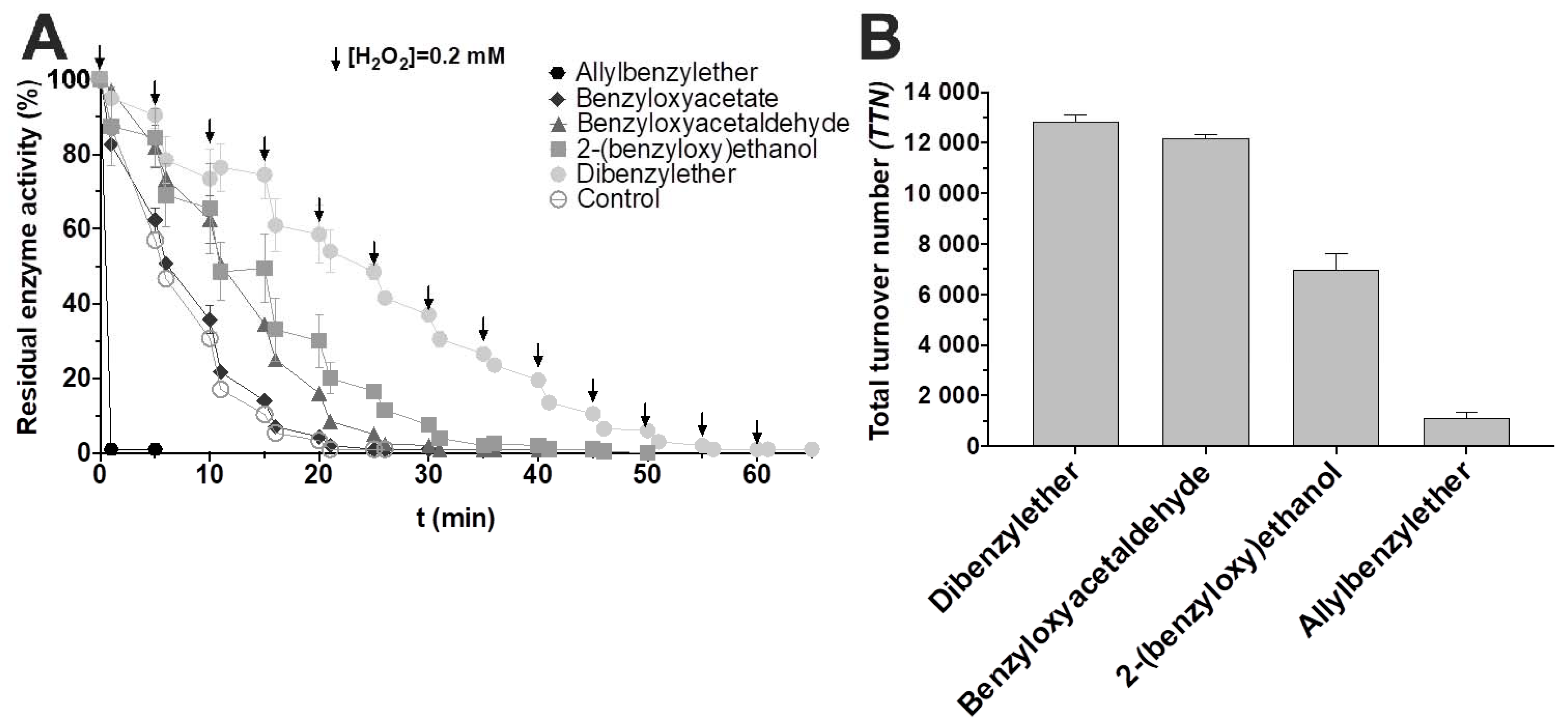
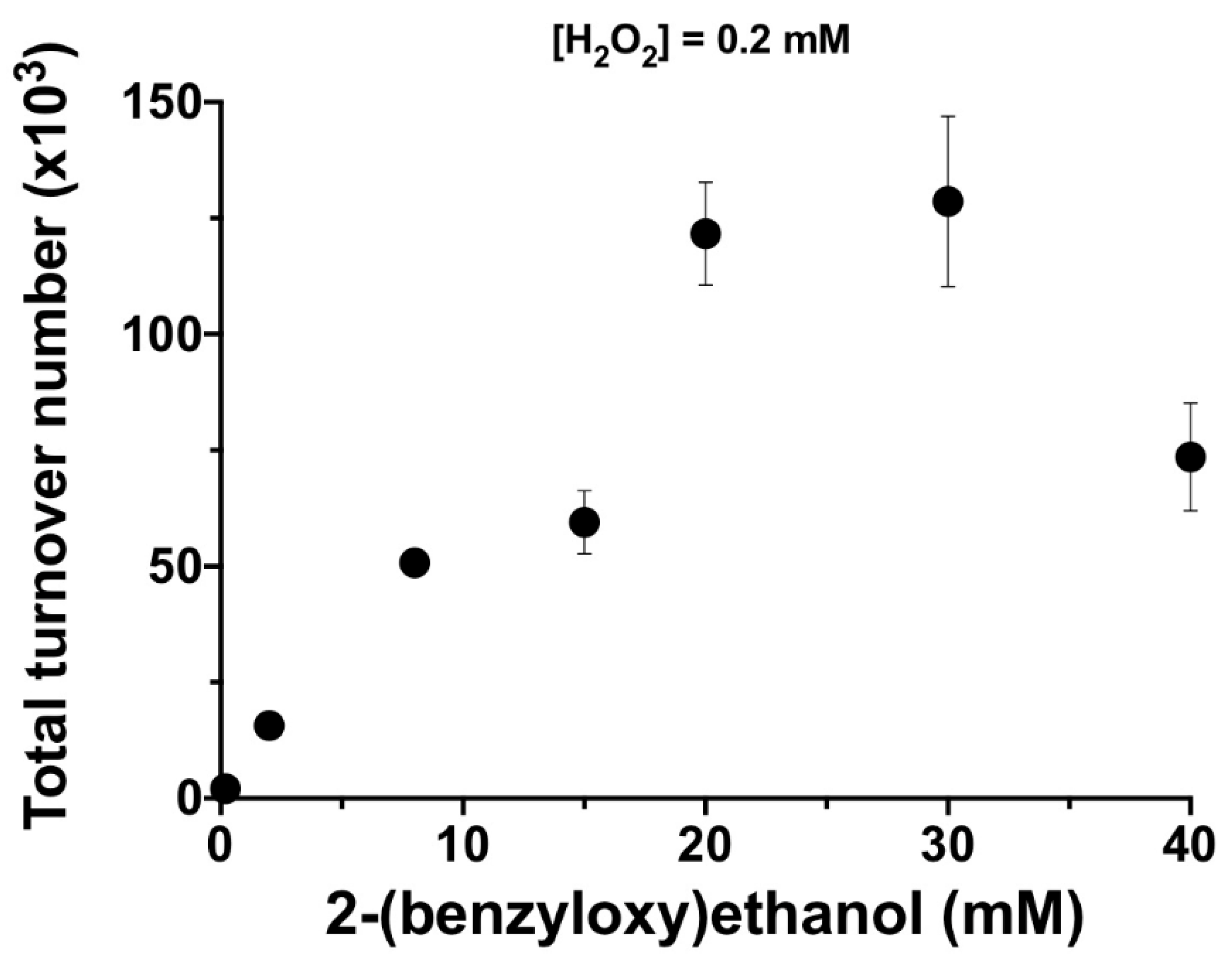
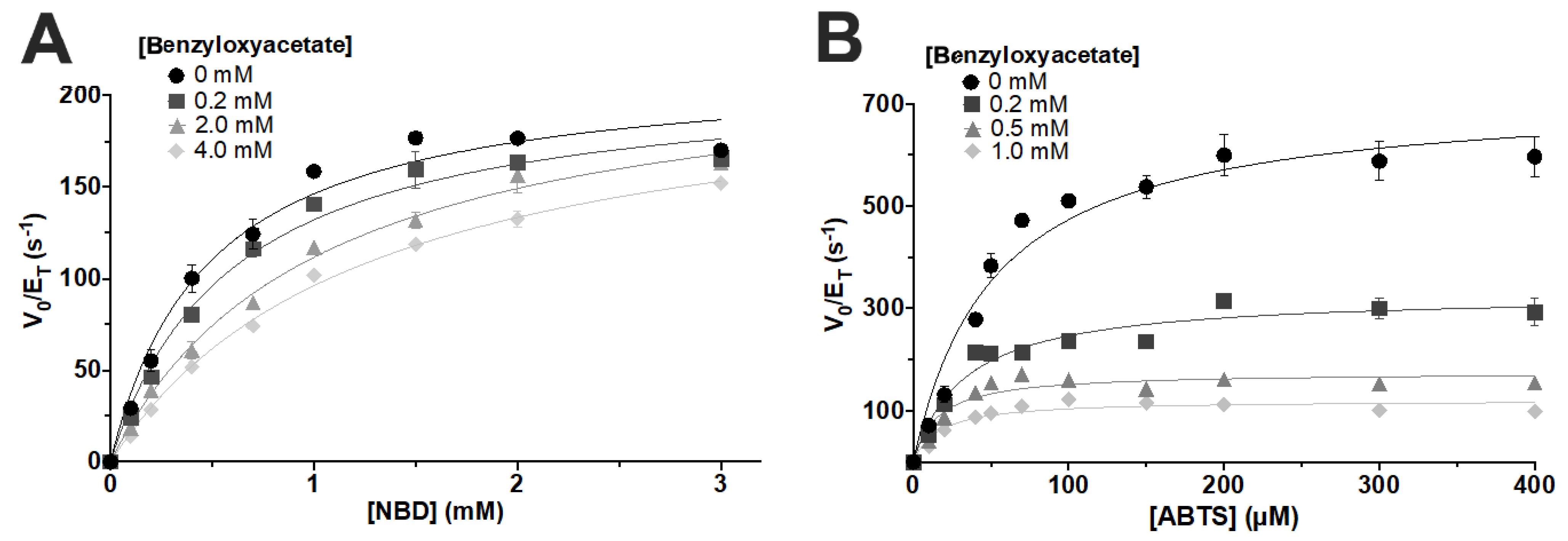
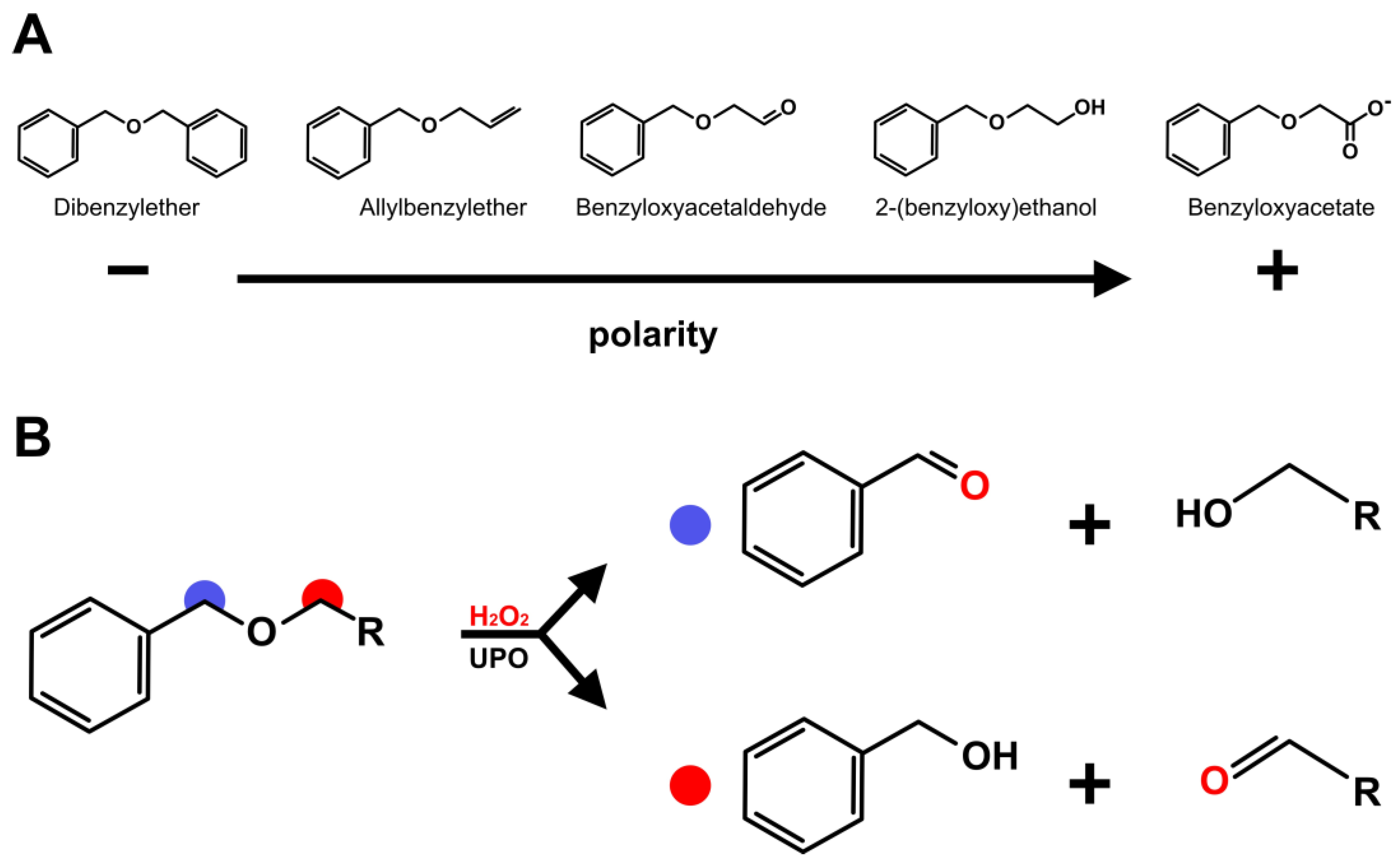
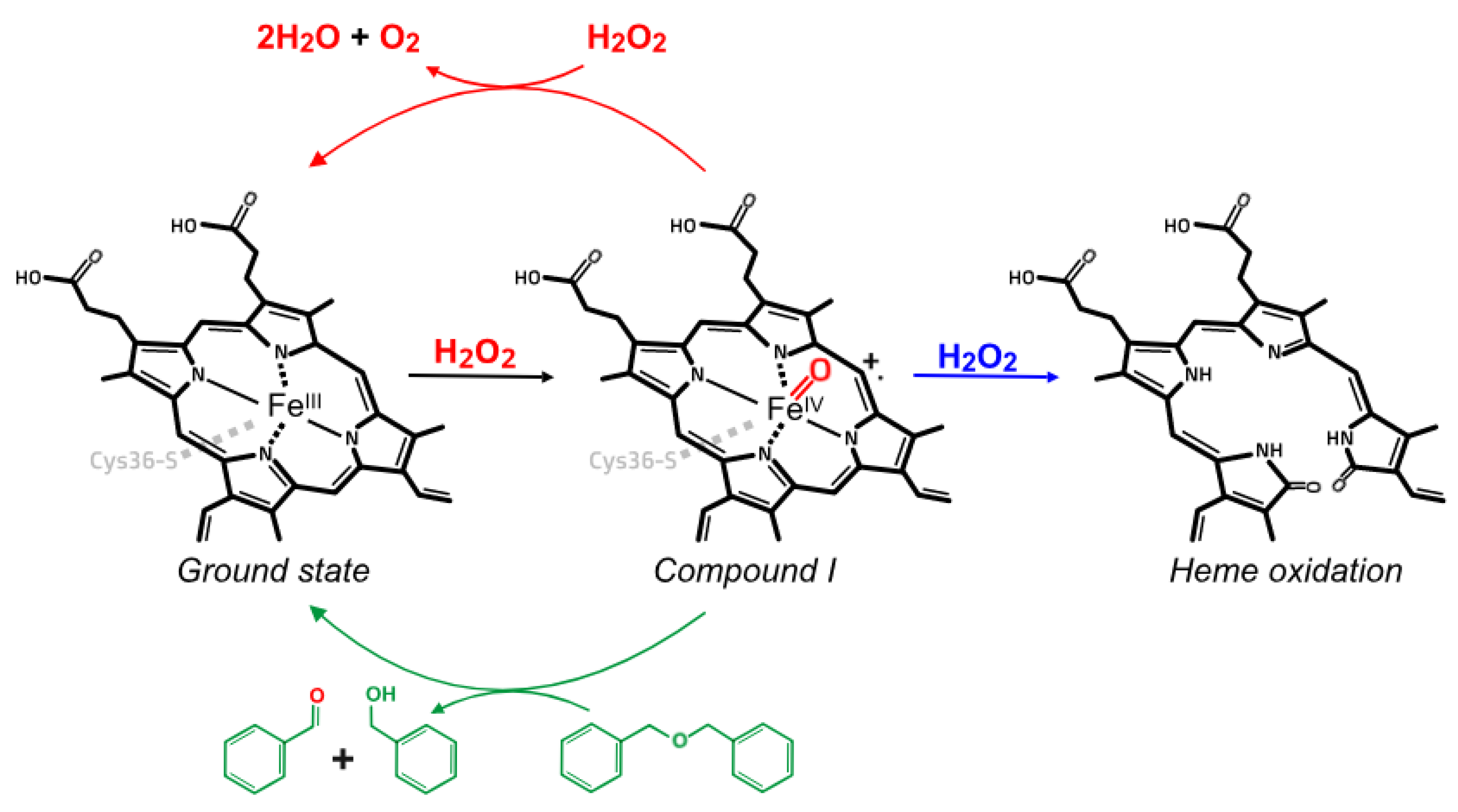
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- White, G.F.; Russell, N.J.; Tidswell, E.C. Bacterial scission of ether bonds. Microbiol. Rev. 1996, 60, 216–232. [Google Scholar] [CrossRef]
- Collinson, S.; Thielemans, W. The catalytic oxidation of biomass to new materials focusing on starch, cellulose and lignin. Co-ord. Chem. Rev. 2010, 254, 1854–1870. [Google Scholar] [CrossRef] [Green Version]
- Weber, R.; Watson, A.; Forter, M.; Oliaei, F. Review Article: Persistent organic pollutants and landfills—A review of past experiences and future challenges. Waste Manag. Res. 2011, 29, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Montes-Grajales, D.; Fennix-Agudelo, M.; Miranda-Castro, W. Occurrence of personal care products as emerging chemicals of concern in water resources: A review. Sci. Total. Environ. 2017, 595, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Environmental Protection Agency. Persistent Organic Pollutants: A Global Issue, A Global Response. 2009. Available online: https://www.epa.gov/international-cooperation/persistent-organic-pollutants-global-issue-global-response (accessed on 1 June 2021).
- Luo, Y.-R. Comprehensive Handbook of Chemical Bond Energies; CRC Press: Boca Ratón, FL, USA, 2007. [Google Scholar]
- Kluge, M.G.; Ullrich, R.; Scheibner, K.; Hofrichter, M. Spectrophotometric assay for detection of aromatic hydroxylation catalyzed by fungal haloperoxidase–peroxygenase. Appl. Microbiol. Biotechnol. 2007, 75, 1473–1478. [Google Scholar] [CrossRef] [PubMed]
- Hofrichter, M.; Kellner, H.; Herzog, R.; Karich, A.; Liers, C.; Scheibner, K.; Kimani, V.W.; Ullrich, R. Fungal Peroxygenases: A Phylogenetically Old Superfamily of Heme Enzymes with Promiscuity for Oxygen Transfer Reactions. In Grand Challenges in Fungal Biotechnology; Nevalainen, H., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 369–403. [Google Scholar]
- Kinne, M.; Poraj-Kobielska, M.; Ralph, S.A.; Ullrich, R.; Hofrichter, M.; Hammel, K.E. Oxidative Cleavage of Diverse Ethers by an Extracellular Fungal Peroxygenase. J. Biol. Chem. 2009, 284, 29343–29349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eltis, L.; Karlson, U.; Timmis, K.N. Purification and characterization of cytochrome P450RR1 from Rhodococcus rhodochrous. Eur. J. Biochem. 1993, 213, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Mallinson, S.J.B.; Machovina, M.M.; Silveira, R.L.; Garcia-Borràs, M.; Gallup, N.; Johnson, C.W.; Allen, M.D.; Skaf, M.S.; Crowley, M.F.; Neidle, E.L.; et al. A promiscuous cytochrome P450 aromatic O-demethylase for lignin bioconversion. Nat. Commun. 2018, 9, 2487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poraj-Kobielska, M.; Kinne, M.; Ullrich, R.; Scheibner, K.; Kayser, G.; Hammel, K.E.; Hofrichter, M. Preparation of human drug metabolites using fungal peroxygenases. Biochem. Pharmacol. 2011, 82, 789–796. [Google Scholar] [CrossRef] [Green Version]
- Karich, A.; Ullrich, R.; Scheibner, K.; Hofrichter, M. Fungal Unspecific Peroxygenases Oxidize the Majority of Organic EPA Priority Pollutants. Front. Microbiol. 2017, 8, 1463. [Google Scholar] [CrossRef]
- Ramirez-Ramirez, J.; Martin-Diaz, J.; Pastor, N.; Alcalde, M.; Ayala, M. Exploring the role of phenylalanine residues in modulating the flexibility and topography of the active site in the peroxygenase variant PaDa-I. Int. J. Mol. Sci. 2020, 21, 5734. [Google Scholar] [CrossRef]
- Molina-Espeja, P.; Garcia-Ruiz, E.; Gonzalez-Perez, D.; Ullrich, R.; Hofrichter, M.; Alcalde, M. Directed Evolution of Unspecific Peroxygenase from Agrocybe aegerita. Appl. Environ. Microbiol. 2014, 80, 3496–3507. [Google Scholar] [CrossRef] [Green Version]
- Poraj-Kobielska, M.; Kinne, M.; Ullrich, R.; Scheibner, K.; Hofrichter, M. A spectrophotometric assay for the detection of fungal peroxygenases. Anal. Biochem. 2012, 421, 327–329. [Google Scholar] [CrossRef] [PubMed]
- Dixon, M. The determination of enzyme inhibitor constants. Biochem. J. 1953, 55, 170–171. [Google Scholar] [CrossRef] [PubMed]
- Cornish-Bowden, A. A simple graphical method for determining the inhibition constants of mixed, uncompetitive and non-competitive inhibitors. Biochem. J. 1974, 137, 143–144. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Hobisch, M.; Holtmann, D.; de Santos, P.G.; Alcalde, M.; Hollmann, F.; Kara, S. Recent developments in the use of peroxygenases—Exploring their high potential in selective oxyfunctionalisations. Biotechnol. Adv. 2020, 107615. [Google Scholar] [CrossRef] [PubMed]
- Valderrama, B.; Ayala, M.; Vazquez-Duhalt, R. Suicide Inactivation of Peroxidases and the Challenge of Engineering More Robust Enzymes. Chem. Biol. 2002, 9, 555–565. [Google Scholar] [CrossRef] [Green Version]
- Ayala, M.; Batista, C.; Vazquez-Duhalt, R. Heme destruction, the main molecular event during the peroxide-mediated inactivation of chloroperoxidase from Caldariomyces fumago. J. Biol. Inorg. Chem. 2010, 16, 63–68. [Google Scholar] [CrossRef]
- Karich, A.; Scheibner, K.; Ullrich, R.; Hofrichter, M. Exploring the catalase activity of unspecific peroxygenases and the mechanism of peroxide-dependent heme destruction. J. Mol. Catal. B Enzym. 2016, 134, 238–246. [Google Scholar] [CrossRef]
- Peter, S.; Kinne, M.; Ullrich, R.; Kayser, G.; Hofrichter, M. Epoxidation of linear, branched and cyclic alkenes catalyzed by unspecific peroxygenase. Enzym. Microb. Technol. 2013, 52, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Dexter, A.F.; Hager, L.P. Transient Heme N-Alkylation of Chloroperoxidase by Terminal Alkenes and Alkynes. J. Am. Chem. Soc. 1995, 117, 817–818. [Google Scholar] [CrossRef]
- Ramirez-Escudero, M.; Molina-Espeja, P.; de Santos, P.G.; Hofrichter, M.; Sanz-Aparicio, J.; Alcalde, M. Structural Insights into the Substrate Promiscuity of a Laboratory-Evolved Peroxygenase. ACS Chem. Biol. 2018, 13, 3259–3268. [Google Scholar] [CrossRef] [PubMed]
- Manoj, K.M.; Hager, L.P. Chloroperoxidase, a Janus Enzyme. Biochemistry 2008, 47, 2997–3003. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2-(benzyloxy)ethanol Concentration (mM) | |||
|---|---|---|---|
| 0 | 2 | 4 | |
| kcat (s−1) | 388 ± 12 | 252 ± 10 | 231 ± 13 |
| Km (μM) | 757 ± 84 | 1111 ± 341 | 2498 ± 331 |
| kcat/Km (M−1s−1) | 5.1 × 105 | 2.3 × 105 | 9.2 × 104 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mireles, R.; Ramirez-Ramirez, J.; Alcalde, M.; Ayala, M. Ether Oxidation by an Evolved Fungal Heme-Peroxygenase: Insights into Substrate Recognition and Reactivity. J. Fungi 2021, 7, 608. https://doi.org/10.3390/jof7080608
Mireles R, Ramirez-Ramirez J, Alcalde M, Ayala M. Ether Oxidation by an Evolved Fungal Heme-Peroxygenase: Insights into Substrate Recognition and Reactivity. Journal of Fungi. 2021; 7(8):608. https://doi.org/10.3390/jof7080608
Chicago/Turabian StyleMireles, Raul, Joaquin Ramirez-Ramirez, Miguel Alcalde, and Marcela Ayala. 2021. "Ether Oxidation by an Evolved Fungal Heme-Peroxygenase: Insights into Substrate Recognition and Reactivity" Journal of Fungi 7, no. 8: 608. https://doi.org/10.3390/jof7080608