
Bioleaching of Manganese Oxides at Different Oxidation States by Filamentous Fungus Aspergillus niger

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strain
2.2. Manganese Toxicity
2.3. Bioextraction of Manganese
2.4. Analytical Procedures
3. Results
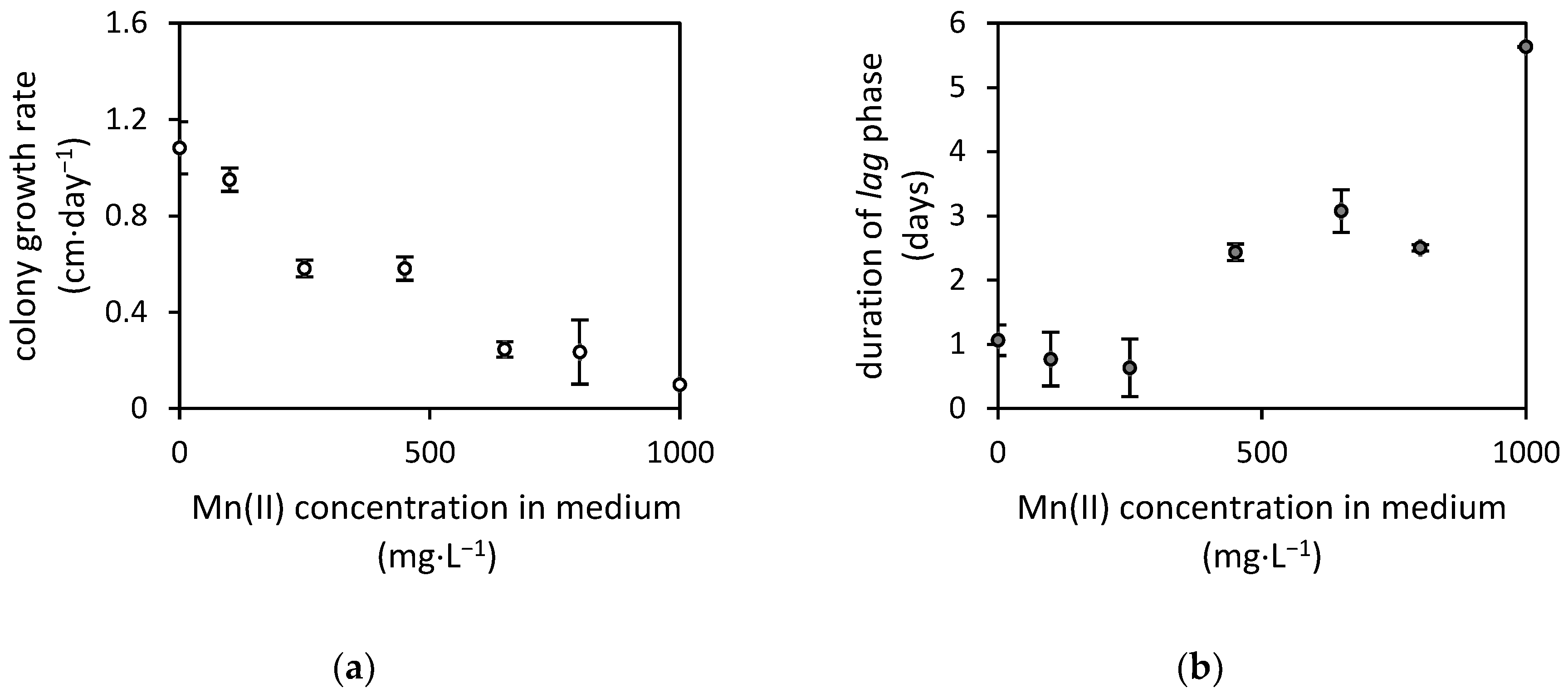
3.1. Fungal Resistance to Manganese(II) and Manganese Oxides
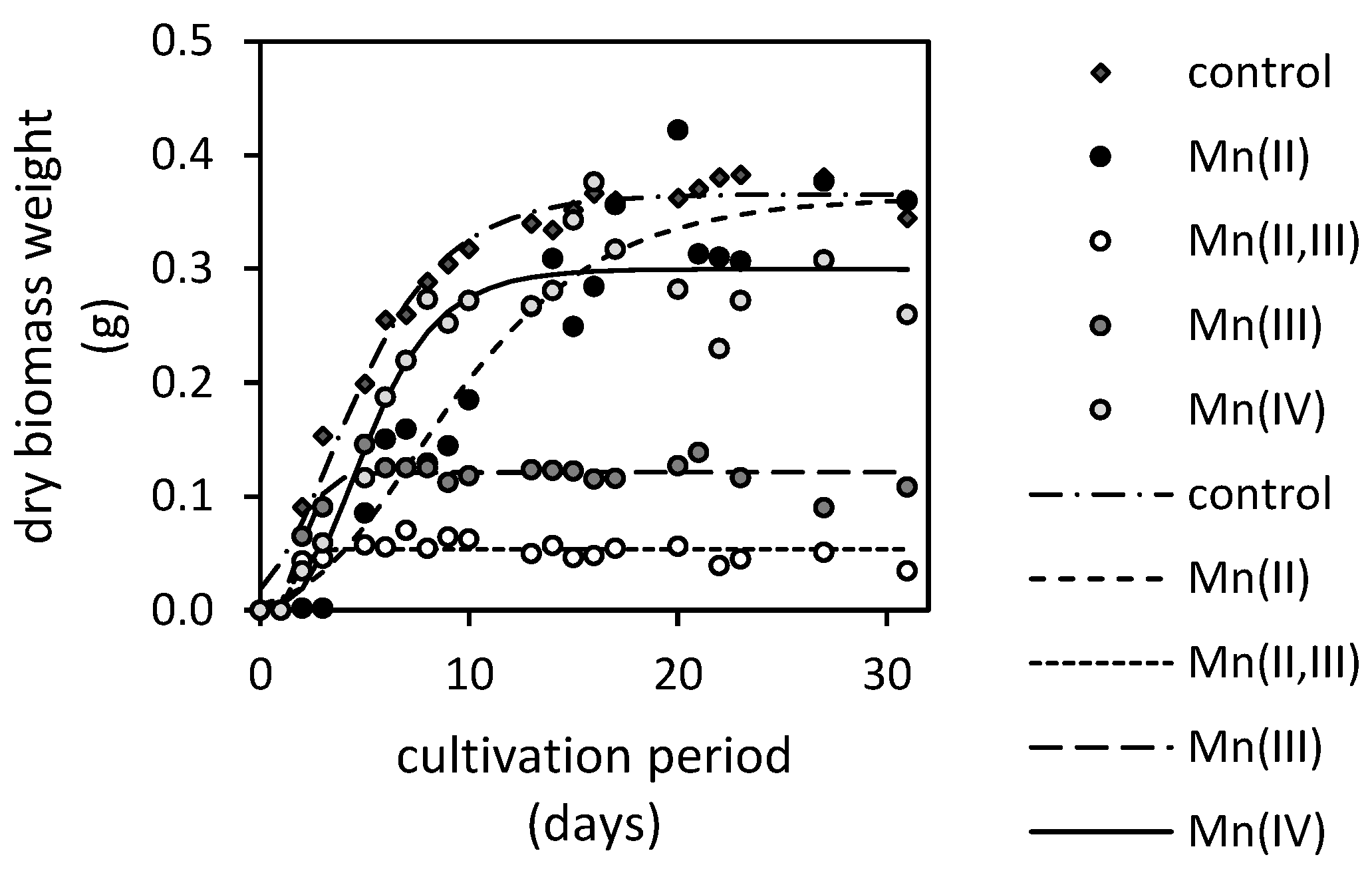
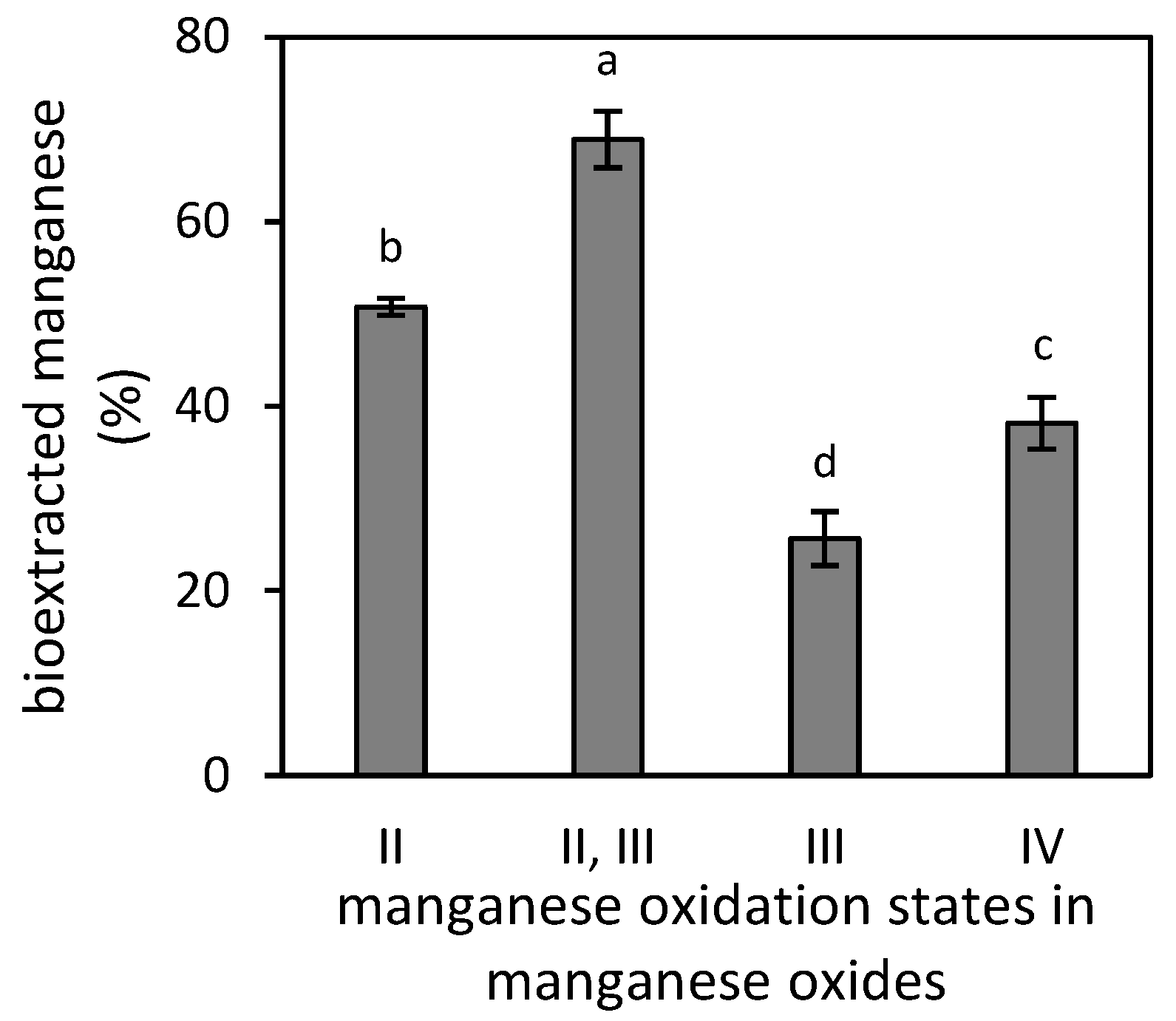
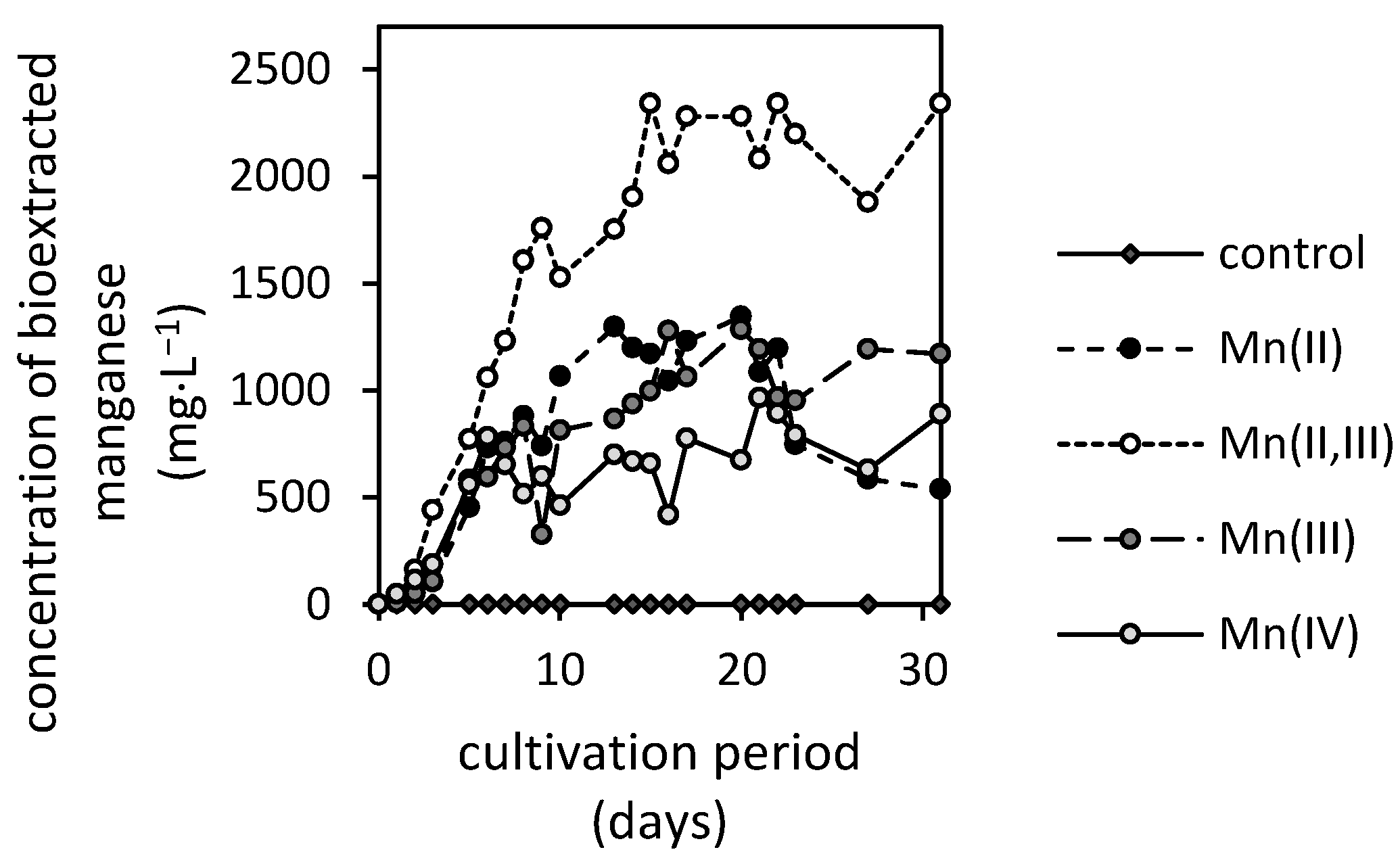
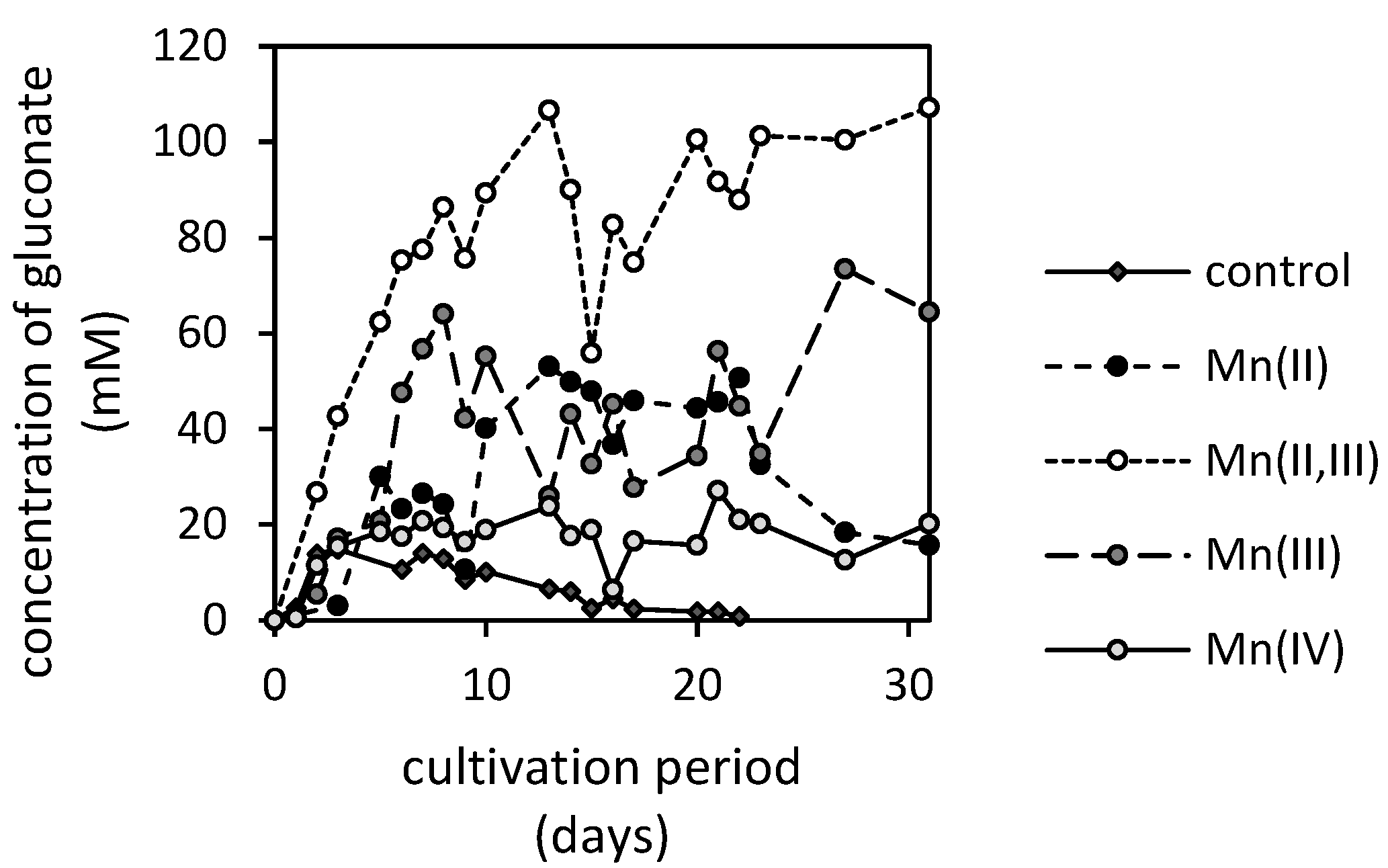
3.2. Manganese Bioextraction
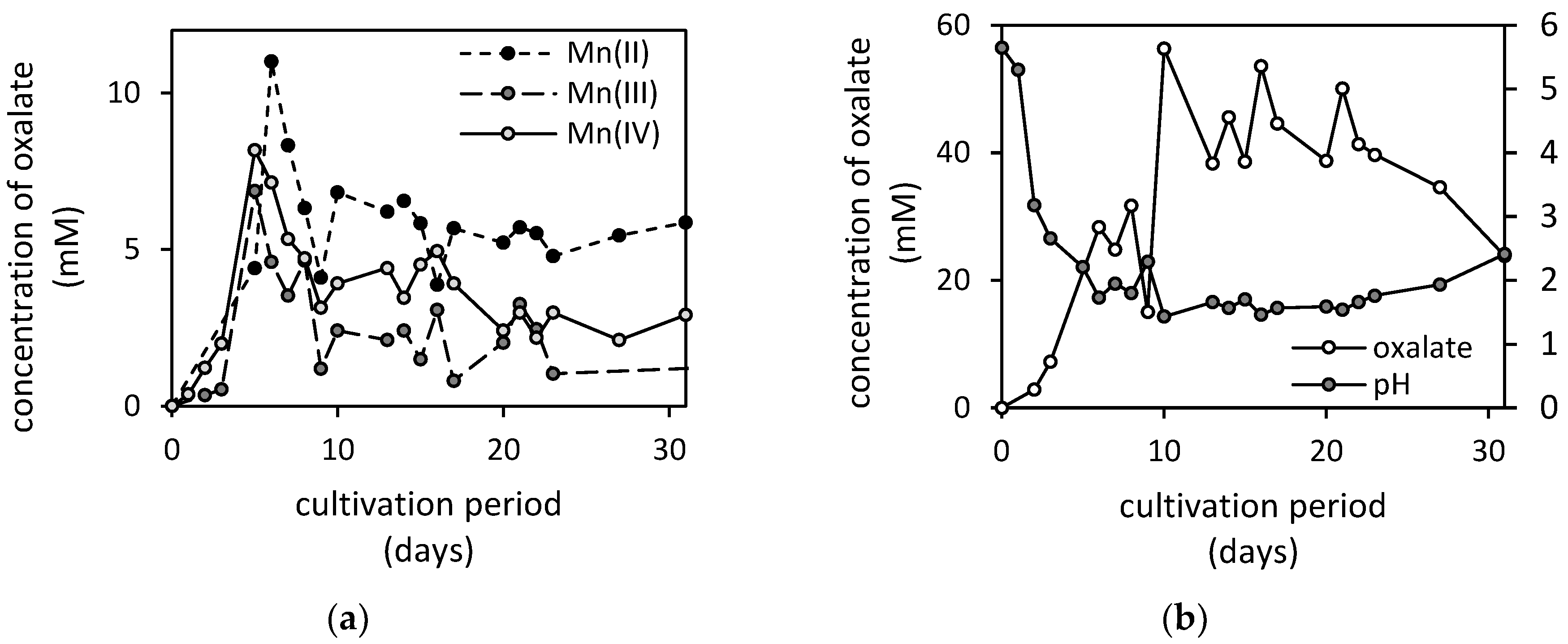
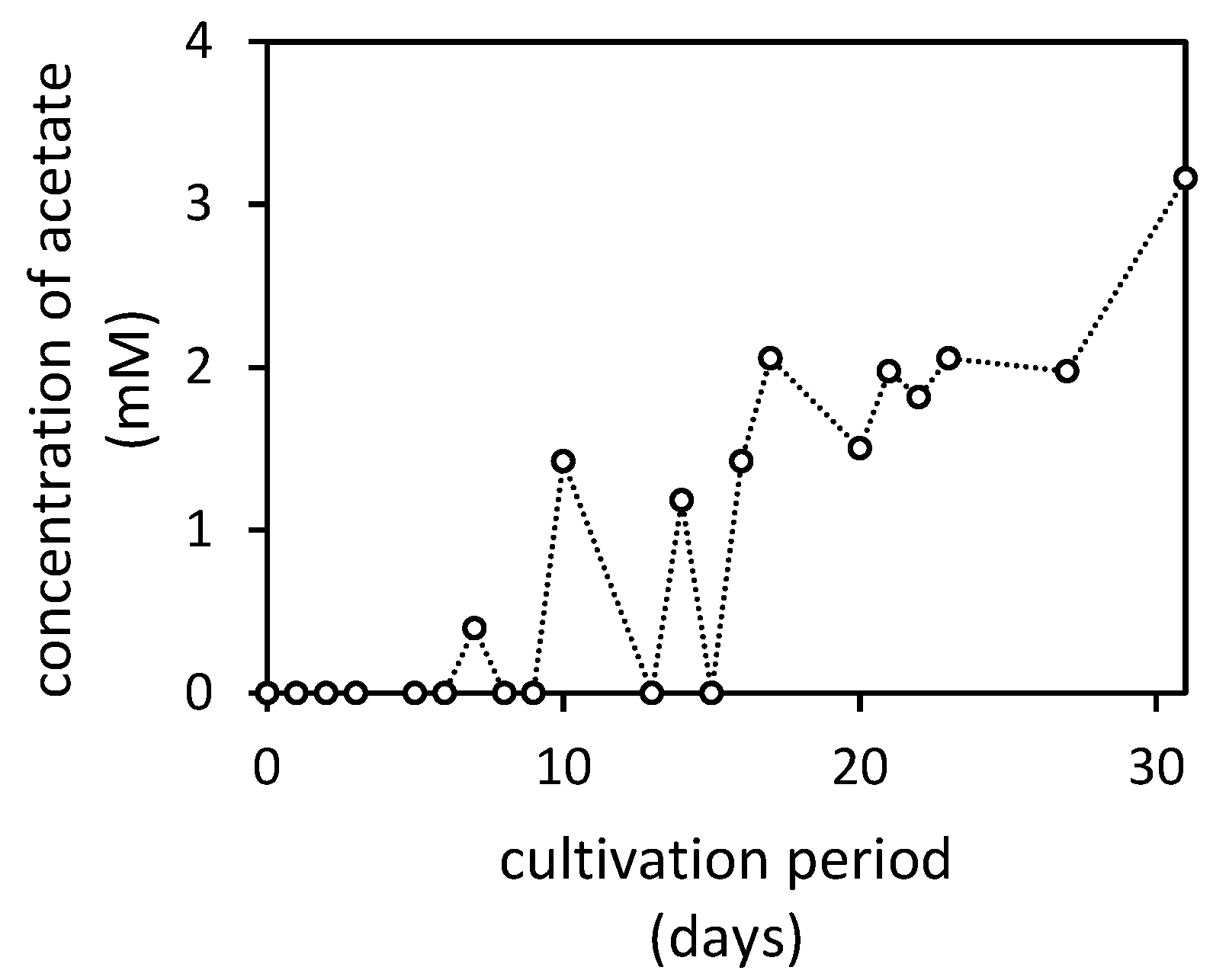
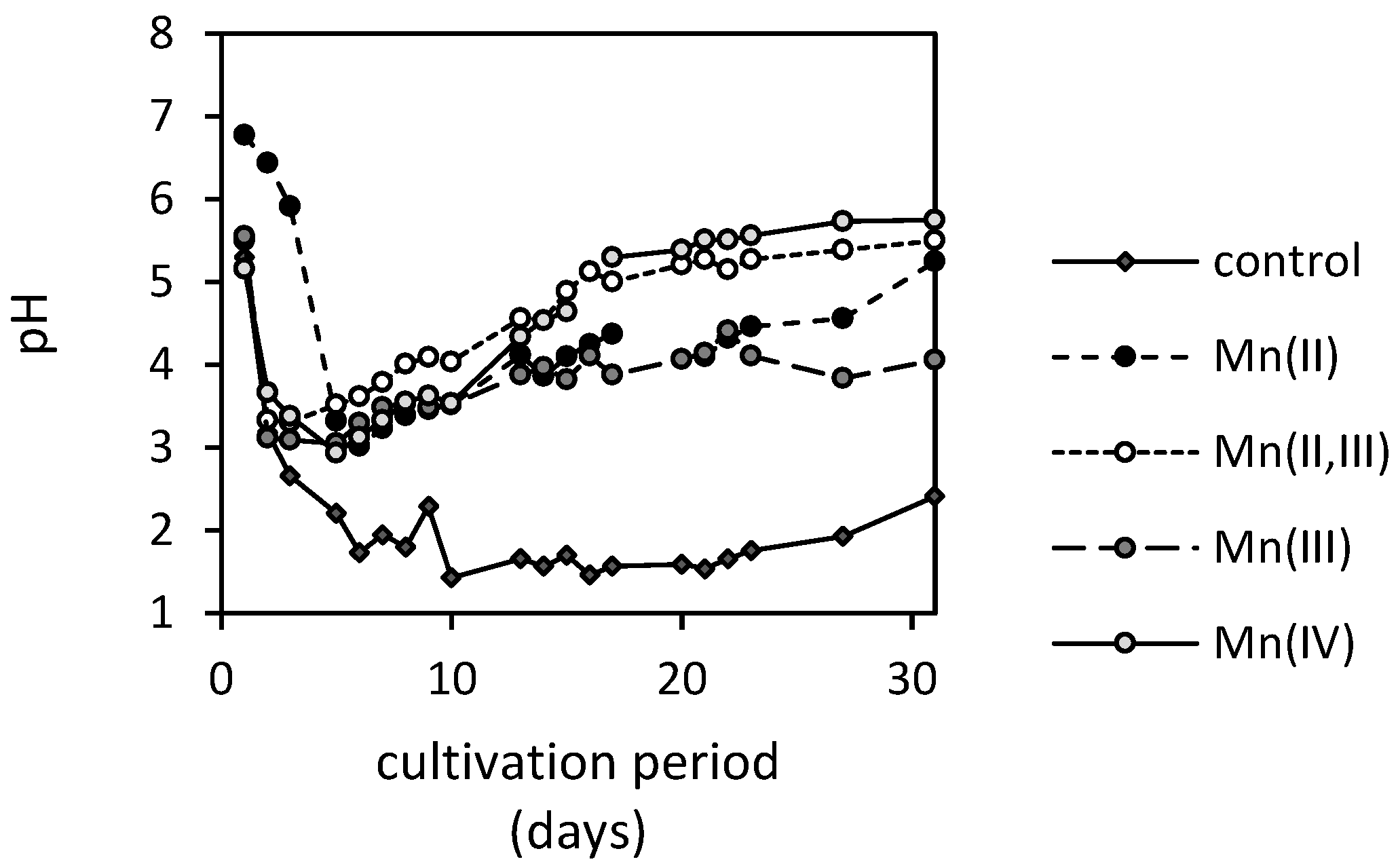
3.3. Changes in pH and Concentrations of Low-Molecular-Weight Organic Acids in the Presence of Manganese Oxides
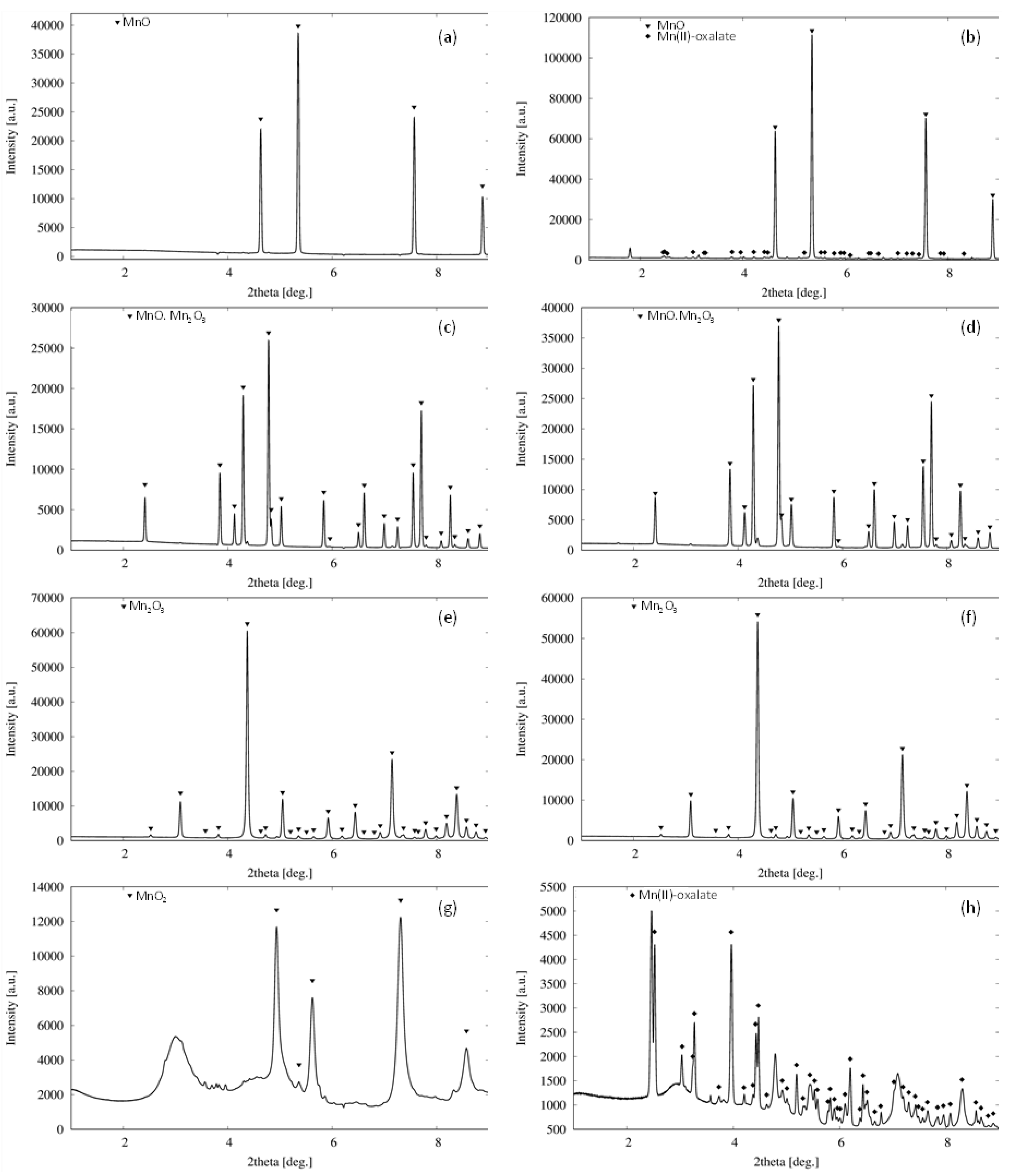
3.4. Formation of Biogenic Mineral
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilkes, R.J.; McKenzie, R.M. Geochemistry and Mineralogy of Manganese in Soils. In Manganese in Soils and Plants, Proceedings of the International Symposium on ‘Manganese in Soils and Plants’, Adelaide, Australia, 22–26 August 1988; Graham, R.D., Hannam, R.J., Uren, N.C., Eds.; Springer: Dordrecht, The Netherlands, 1988; pp. 23–35. [Google Scholar]
- Tebo, B.M.; Bargar, J.R.; Clement, B.G.; Dick, G.J.; Murray, K.J.; Parker, D.; Verity, R.; Webb, S.M. Biogenic manganese oxides: Properties and mechanisms of formation. Annu. Rev. Earth Planet. Sci. 2004, 32, 287–328. [Google Scholar] [CrossRef] [Green Version]
- Robson, A.D. Manganese in Soils and Plants—An Overview. In Manganese in Soils and Plants, Proceedings of the International Symposium on ‘Manganese in Soils and Plants’, Adelaide, Australia, 22–26 August 1988; Graham, R.D., Hannam, R.J., Uren, N.C., Eds.; Springer: Dordrecht, The Netherlands, 1988; pp. 329–333. [Google Scholar]
- Baranowski, R.; Rybak, A.; Baranowska, I. Speciation Analysis of Elements in Soil Samples by XRF. Pol. J. Environ. Stud. 2002, 11, 473–482. [Google Scholar]
- Baranowski, R.; Rybak, A.; Sobczyński, T. X-ray Fluorescence Spectrometry in Speciation Analysis of Bottom Sediments. J. Environ. Stud. 2001, 10, 297–306. [Google Scholar]
- Zerbe, J.; Sobczyński, T.; Elbanowska, H.; Siepak, J. Speciation of heavy metals in bottom sediments of lakes. Pol. J. Environ. Stud. 1999, 8, 331–339. [Google Scholar]
- Remucal, C.K.; Ginder-Vogel, M. A critical review of the reactivity of manganese oxides with organic contaminants. Environ. Sci. Process. Impacts 2014, 16, 1247–1266. [Google Scholar] [CrossRef] [PubMed]
- Archibald, F.S.; Fridovich, I. Manganese and defenses against oxygen toxicity in Lactobacillus plantarum. J. Bacteriol. 1981, 145, 442–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernando, D.R.; Lynch, J.P. Manganese phytotoxicity: New light on an old problem. Ann. Bot. 2015, 116, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Culotta, V.C.; Yang, M.; Hall, M.D. Manganese transport and trafficking: Lessons learned from Saccharomyces cerevisiae. Eukaryot. Cell 2005, 4, 1159–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perel’man, A.I. Geochemical barriers: Theory and practical applications. Appl. Geochem. 1986, 1, 669–680. [Google Scholar] [CrossRef]
- Hagarová, I. Utilization of biosurfactants in remediation of environmental media contaminated with heavy metals. Chem. Listy 2015, 109, 431–436. [Google Scholar]
- Hagarová, I. Extractions complying with the principles of green chemistry used in trace analysis of metals. Chem. Listy 2015, 109, 269–275. [Google Scholar]
- Boriová, K.; Urík, M.; Bujdoš, M.; Pifková, I.; Matúš, P. Chemical mimicking of bio-assisted aluminium extraction by Aspergillus niger’s exometabolites. Environ. Pollut. 2016, 218, 281–288. [Google Scholar] [CrossRef]
- Zwietering, M.H.; Jongenburger, I.; Rombouts, F.M.; van‘t Riet, K. Modeling of the bacterial growth curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [CrossRef] [Green Version]
- Sádecká, J.; Polonský, J. Determination of organic acids in tobacco by capillary isotachophoresis. J. Chromatogr. A 2003, 988, 161–165. [Google Scholar] [CrossRef]
- Farkas, B.; Kolenčík, M.; Hain, M.; Dobročka, E.; Kratošová, G.; Bujdoš, M.; Feng, H.; Deng, Y.; Yu, Q.; Illa, R.; et al. Aspergillus niger decreases bioavailability of arsenic(V) via biotransformation of manganese oxide into biogenic oxalate minerals. J. Fungi 2020, 6, 270. [Google Scholar] [CrossRef] [PubMed]
- Urík, M.; Farkas, B.; Miglierini, M.B.; Bujdoš, M.; Mitróová, Z.; Kim, H.; Matúš, P. Mobilisation of hazardous elements from arsenic-rich mine drainage ochres by three Aspergillus species. J. Hazard. Mater. 2021, 409, 124938. [Google Scholar] [CrossRef] [PubMed]
- Grangeon, S.; Bataillard, P.; Coussy, S. The Nature of Manganese Oxides in Soils and Their Role as Scavengers of Trace Elements: Implication for Soil Remediation. In Environmental Soil Remediation and Rehabilitation: Existing and Innovative Solutions; van Hullebusch, E.D., Huguenot, D., Pechaud, Y., Simonnot, M.-O., Colombano, S., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 399–429. [Google Scholar]
- Farkas, B.; Urík, M.; Matúš, P. Manganese biotransformation by microorganisms. Chem. Listy 2020, 114, 841–846. [Google Scholar]
- Das, A.P.; Sukla, L.B.; Pradhan, N.; Nayak, S. Manganese biomining: A review. Bioresour. Technol. 2011, 102, 7381–7387. [Google Scholar] [CrossRef]
- Acharya, C.; Kar, R.N.; Sukla, L.B. Studies on reaction mechanism of bioleaching of manganese ore. Miner. Eng. 2003, 16, 1027–1030. [Google Scholar] [CrossRef]
- Mehta, K.D.; Das, C.; Pandey, B.D. Leaching of copper, nickel and cobalt from Indian Ocean manganese nodules by Aspergillus niger. Hydrometallurgy 2010, 105, 89–95. [Google Scholar] [CrossRef]
- Keshavarz, S.; Faraji, F.; Rashchi, F.; Mokmeli, M. Bioleaching of manganese from a low-grade pyrolusite ore using Aspergillus niger: Process optimization and kinetic studies. J. Environ. Manag. 2021, 285, 112153. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-J.; Seo, J.-Y.; Choi, Y.-S.; Kim, G.-H. Bioleaching of spent Zn–Mn or Ni–Cd batteries by Aspergillus species. Waste Manag. 2016, 51, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Polák, F.; Urík, M.; Matúš, P. Low molecular weight organic acids in soil environment. Chem. Listy 2019, 113, 307–314. [Google Scholar]
- Bagar, T.; Altenbach, K.; Read, N.D.; Bencina, M. Live-Cell imaging and measurement of intracellular pH in filamentous fungi using a genetically encoded ratiometric probe. Eukaryot. Cell 2009, 8, 703–712. [Google Scholar] [CrossRef] [Green Version]
- Serrano, R. Structure and function of proton translocating ATPase in plasma membranes of plants and fungi. Biochim. Biophys. Acta Rev. Biomembr. 1988, 947, 1–28. [Google Scholar] [CrossRef]
- Urík, M.; Polák, F.; Bujdoš, M.; Pifková, I.; Kořenková, L.; Littera, P.; Matúš, P. Aluminium Leaching by Heterotrophic Microorganism Aspergillus niger: An Acidic Leaching? Arab. J. Sci. Eng. 2018, 43, 2369–2374. [Google Scholar] [CrossRef]
- Ruijter, G.J.G.; van de Vondervoort, P.J.I.; Visser, J. Oxalic acid production by Aspergillus niger: An oxalate-non-producing mutant produces citric acid at pH 5 and in the presence of manganese. Microbiology 1999, 145, 2569–2576. [Google Scholar] [CrossRef] [Green Version]
- Xyla, A.G.; Sulzberger, B.; Luther, G.W.; Hering, J.G.; Van Cappellen, P.; Stumm, W. Reductive dissolution of manganese(III, IV) (hydr)oxides by oxalate: The effect of pH and light. Langmuir 1992, 8, 95–103. [Google Scholar] [CrossRef]
- Miyata, N.; Tani, Y.; Iwahori, K.; Soma, M. Enzymatic formation of manganese oxides by an Acremonium-like hyphomycete fungus, strain KR21-2. FEMS Microbiol. Ecol. 2004, 47, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Höfer, C.; Schlosser, D. Novel enzymatic oxidation of Mn2+ to Mn3+ catalyzed by a fungal laccase. FEBS Lett. 1999, 451, 186–190. [Google Scholar] [CrossRef] [Green Version]
- Zeiner, C.A.; Purvine, S.O.; Zink, E.; Wu, S.; Paša-Tolić, L.; Chaput, D.L.; Santelli, C.M.; Hansel, C.M. Mechanisms of Manganese(II) Oxidation by Filamentous Ascomycete Fungi Vary with Species and Time as a Function of Secretome Composition. Front. Microbiol. 2021, 12, 81. [Google Scholar] [CrossRef]
- Tang, Y.; Zeiner, C.A.; Santelli, C.M.; Hansel, C.M. Fungal oxidative dissolution of the Mn(II)-bearing mineral rhodochrosite and the role of metabolites in manganese oxide formation. Environ. Microbiol. 2013, 15, 1063–1077. [Google Scholar] [CrossRef]
- Johnson, D.B.; Pakostova, E. Dissolution of Manganese (IV) Oxide Mediated by Acidophilic Bacteria, and Demonstration That Manganese (IV) Can Act as Both a Direct and Indirect Electron Acceptor for Iron-Reducing Acidithiobacillus spp. Geomicrobiol. J. 2021, 38, 570–576. [Google Scholar] [CrossRef]
- Wei, Z.; Hillier, S.; Gadd, G.M. Biotransformation of manganese oxides by fungi: Solubilization and production of manganese oxalate biominerals. Environ. Microbiol. 2012, 14, 1744–1753. [Google Scholar] [CrossRef]
- Gadd, G.M. Metals, minerals and microbes: Geomicrobiology and bioremediation. Microbiology 2010, 156, 609–643. [Google Scholar] [CrossRef]
- Das, A.P.; Swain, S.; Panda, S.; Pradhan, N.; Sukla, L.B. Reductive Acid Leaching of Low Grade Manganese Ores. Geomaterials 2012, 2, 70–72. [Google Scholar] [CrossRef]
- Karaffa, L.; Kubicek, C. Production of Organic Acids by Fungi. In The Mycota. Industrial Applications; Hofrichter, M., Ed.; Springer: Berlin/Heidelberg, Germay, 2020; pp. 215–234. [Google Scholar]
- Šimonovičová, A.; Kupka, D.; Nosalj, S.; Kraková, L.; Drahovská, H.; Bártová, Z.; Vojtková, H.; Boturová, K.; Pangallo, D. Differences in metabolites production using the Biolog FF Microplate™ system with an emphasis on some organic acids of Aspergillus niger wild type strains. Biologia 2020, 75, 1537–1546. [Google Scholar] [CrossRef]
- Show, P.L.; Oladele, K.O.; Siew, Q.Y.; Aziz Zakry, F.A.; Lan, J.C.-W.; Ling, T.C. Overview of citric acid production from Aspergillus niger. Front. Life Sci. 2015, 8, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.S.; Farag, S.S.; Hassan, I.A.; Botros, H.W. Production of gluconic acid by using some irradiated microorganisms. J. Radiat. Res. Appl. Sci. 2015, 8, 374–380. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, K.; Peter, V.; Meinert, S.; Kornfeld, G.; Hardiman, T.; Wiechert, W.; Noack, S. Simultaneous utilization of glucose and gluconate in Penicillium chrysogenum during overflow metabolism. Biotechnol. Bioeng. 2013, 110, 3235–3243. [Google Scholar] [CrossRef]
- Ferrier, J.; Yang, Y.; Csetenyi, L.; Gadd, G.M. Colonization, penetration and transformation of manganese oxide nodules by Aspergillus niger. Environ. Microbiol. 2019, 21, 1821–1832. [Google Scholar] [CrossRef] [Green Version]
- Fomina, M.; Hillier, S.; Charnock, J.M.; Melville, K.; Alexander, I.J.; Gadd, G.M. Role of Oxalic Acid Overexcretion in Transformations of Toxic Metal Minerals by Beauveria caledonica. Appl. Environ. Microbiol. 2005, 71, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Cheung, H.Y.; Vitkovič, L.; Brown, M.R.W. Toxic Effect of Manganese on Growth and Sporulation of Bacillus stearothermophilus. Microbiology 1982, 128, 2395–2402. [Google Scholar] [CrossRef] [Green Version]
- Green Iii, F.; Clausen, C.A. Copper tolerance of brown-rot fungi: Time course of oxalic acid production. Int. Biodeterior. Biodegrad. 2003, 51, 145–149. [Google Scholar] [CrossRef]
- Li, Q.; Liu, D.; Jia, Z.; Csetenyi, L.; Gadd, G.M. Fungal Biomineralization of Manganese as a Novel Source of Electrochemical Materials. Curr. Biol. 2016, 26, 950–955. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farkas, B.; Bujdoš, M.; Polák, F.; Matulová, M.; Cesnek, M.; Duborská, E.; Zvěřina, O.; Kim, H.; Danko, M.; Kisová, Z.; et al. Bioleaching of Manganese Oxides at Different Oxidation States by Filamentous Fungus Aspergillus niger. J. Fungi 2021, 7, 808. https://doi.org/10.3390/jof7100808
Farkas B, Bujdoš M, Polák F, Matulová M, Cesnek M, Duborská E, Zvěřina O, Kim H, Danko M, Kisová Z, et al. Bioleaching of Manganese Oxides at Different Oxidation States by Filamentous Fungus Aspergillus niger. Journal of Fungi. 2021; 7(10):808. https://doi.org/10.3390/jof7100808
Chicago/Turabian StyleFarkas, Bence, Marek Bujdoš, Filip Polák, Michaela Matulová, Martin Cesnek, Eva Duborská, Ondřej Zvěřina, Hyunjung Kim, Martin Danko, Zuzana Kisová, and et al. 2021. "Bioleaching of Manganese Oxides at Different Oxidation States by Filamentous Fungus Aspergillus niger" Journal of Fungi 7, no. 10: 808. https://doi.org/10.3390/jof7100808