
Structurally Uncommon Secondary Metabolites Derived from Endophytic Fungi

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Alkaloids
3.1.1. Cytochalasans
3.1.2. Indole Alkaloids
3.1.3. Isoindole Derivatives
3.1.4. Pyrrolidone Derivatives
3.1.5. Pyridone and Pyridinyl Derivatives
3.1.6. Diketopiperazine Derivatives
3.1.7. Other Nitrogen-Containing Compounds (Amides and Amines)
3.1.8. Other Types of Alkaloids
3.2. Peptides
3.3. Terpenoids
3.3.1. Sesquiterpenoids
3.3.2. Diterpenoids and Sesterterpenoids
3.3.3. Meroterpenoids
3.4. Polyketides
3.5. Steroids
3.6. Analysis of Neighbor Net and Diversity of Endophytic Fungi Producing Secondary Metabolites with Uncommon Structures
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petrini, O. Fungal Endophytes of Tree Leaves. In Microbial Ecology of Leaves; Andrews, J.H., Hirano, S.S., Eds.; Springer: New York, NY, USA, 1991; pp. 179–197. [Google Scholar]
- Carroll, G. Fungal Endophytes in Stems and Leaves: From Latent Pathogen to Mutualistic Symbiont. Ecology 1988, 69, 2–9. [Google Scholar] [CrossRef]
- Hallmann, J.; Sikora, R.A. Toxicity of fungal endophyte secondary metabolites to plant parasitic nematodes and soil-borne plant pathogenic fungi. Eur. J. Plant Pathol. 1996, 102, 155–162. [Google Scholar] [CrossRef]
- Sturz, A.; Nowak, J. Endophytic communities of rhizobacteria and the strategies required to create yield enhancing associations with crops. Appl. Soil Ecol. 2000, 15, 183–190. [Google Scholar] [CrossRef]
- Azevedo, J.L.; Araujo, W.L. Diversity and applications of endophytic fungi isolated from tropical plants. In Fungi: Multifaceted Microbes; Ganguli, B.N., Deshmukh, S.K., Eds.; CRC Press: New Delhi, India, 2007; pp. 189–207. ISBN 9781420043280. [Google Scholar]
- Torres-Mendoza, D.; Ortega, H.E.; Cubilla-Rios, L. Patents on Endophytic Fungi Related to Secondary Metabolites and Biotransformation Applications. J. Fungi 2020, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Ortega, H.E.; Torres-Mendoza, D.; Cubilla-Rios, L. Patents on Endophytic Fungi for Agriculture and Bio- and Phytoremediation Applications. Microorganisms 2020, 8, 1237. [Google Scholar] [CrossRef] [PubMed]
- Kusari, S.; Hertweck, C.; Spiteller, M. Chemical Ecology of Endophytic Fungi: Origins of Secondary Metabolites. Chem. Biol. 2012, 19, 792–798. [Google Scholar] [CrossRef] [Green Version]
- Kjærbølling, I.; Mortensen, U.H.; Vesth, T.; Andersen, M.R. Strategies to establish the link between biosynthetic gene clusters and secondary metabolites. Fungal Genet. Biol. 2019, 130, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Nisa, H.; Kamili, A.N.; Nawchoo, I.A.; Shafi, S.; Shameem, N.; Bandh, S.A. Fungal endophytes as prolific source of phytochemicals and other bioactive natural products: A review. Microb. Pathog. 2015, 82, 50–59. [Google Scholar] [CrossRef]
- Bertrand, S.; Bohni, N.; Schnee, S.; Schumpp, O.; Gindro, K.; Wolfender, J.-L. Metabolite induction via microorganism co-culture: A potential way to enhance chemical diversity for drug discovery. Biotechnol. Adv. 2014, 32, 1180–1204. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big Effects from Small Changes: Possible Ways to Explore Nature’s Chemical Diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- El-Hawary, S.S.; Moawad, A.S.; Bahr, H.S.; Abdelmohsen, U.R.; Mohammed, R. Natural product diversity from the endophytic fungi of the genus Aspergillus. RSC Adv. 2020, 10, 22058–22079. [Google Scholar] [CrossRef]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal Identification Using Molecular Tools: A Primer for the Natural Products Research Community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef]
- Skellam, E. The biosynthesis of cytochalasans. Nat. Prod. Rep. 2017, 34, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhang, X.; Al Anbari, W.H.; Zhou, Q.; Zhou, P.; Zhang, M.; Zeng, F.; Chen, C.; Tong, Q.; Wang, J.; et al. Cysteine Residue Containing Merocytochalasans and 17,18-seco-Aspochalasins from Aspergillus micronesiensis. J. Nat. Prod. 2019, 82, 2653–2658. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Tao, X.; Liu, J.; Chen, R.; Zhang, M.; Li, L.; Fang, X.; Yu, L.Y.; Dai, J. Periconiasin G, a new cytochalasan with unprecedented 7/6/5 tricyclic ring system from the endophytic fungus Periconia sp. Tetrahedron Lett. 2016, 57, 796–799. [Google Scholar] [CrossRef]
- Yan, B.-C.; Wang, W.-G.; Hu, D.-B.; Sun, X.; Kong, L.-M.; Li, X.-N.; Du, X.; Luo, S.H.; Liu, Y.; Li, Y.; et al. Phomopchalasins A and B, Two Cytochalasans with Polycyclic-Fused Skeletons from the Endophytic Fungus Phomopsis sp. shj2. Org. Lett. 2016, 18, 1108–1111. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-X.; Feng, T.; Li, Z.-H.; Li, J.; Ai, H.-L.; Liu, J.-K. Cytochalasins D1 and C1, unique cytochalasans from endophytic fungus Xylaria cf. curta. Tetrahedron Lett. 2019, 60, 150952. [Google Scholar] [CrossRef]
- Huang, S.; Chen, H.; Li, W.; Zhu, X.; Ding, W.; Li, C. Bioactive Chaetoglobosins from the Mangrove Endophytic Fungus Penicillium chrysogenum. Mar. Drugs 2016, 14, 172. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Zhou, D.; Liang, F.; Wu, Z.; She, Z.; Li, C. Penochalasin K, a new unusual chaetoglobosin from the mangrove endophytic fungus Penicillium chrysogenum V11 and its effective semi-synthesis. Fitoterapia 2017, 123, 23–28. [Google Scholar] [CrossRef]
- Yang, M.-H.; Gu, M.-L.; Han, C.; Guo, X.-J.; Yin, G.-P.; Yu, P.; Kong, L.-Y. Aureochaeglobosins A–C, Three [4 + 2] Adducts of Chaetoglobosin and Aureonitol Derivatives from Chaetomium globosum. Org. Lett. 2018, 20, 3345–3348. [Google Scholar] [CrossRef]
- Chen, Z.M.; Chen, H.P.; Li, Y.; Feng, T.; Liu, J.-K. Cytochalasins from cultures of endophytic fungus Phoma multirostrata EA-12. J. Antibiot. 2015, 68, 23–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.X.; Lei, X.; Yang, Y.L.; Li, Z.H.; Ai, H.L.; Li, J.; Feng, T.; Liu, J.K. Xylarichalasin A, a Halogenated Hexacyclic Cytochalasan from the Fungus Xylaria cf. curta. Org. Lett. 2019, 21, 6957–6960. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-H.; Li, G.; Qiao, Y.-N.; Sun, Y.; Peng, X.-P.; Lou, H.-X. Chamiside A, a Cytochalasan with a Tricyclic Core Skeleton from the Endophytic Fungus Chaetomium nigricolor F5. Org. Lett. 2019, 21, 3319–3322. [Google Scholar] [CrossRef]
- Wang, W.X.; Li, Z.H.; Feng, T.; Li, J.; Sun, H.; Huang, R.; Yuan, Q.X.; Ai, H.L.; Liu, J.K. Curtachalasins A and B, Two Cytochalasans with a Tetracyclic Skeleton from the Endophytic Fungus Xylaria curta E10. Org. Lett. 2018, 20, 7758–7761. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-X.; Lei, X.; Ai, H.-L.; Bai, X.; Li, J.; He, J.; Li, Z.-H.; Zheng, Y.-S.; Feng, T.; Liu, J.-K. Cytochalasans from the Endophytic Fungus Xylaria cf. curta with Resistance Reversal Activity against Fluconazole-Resistant Candida albicans. Org. Lett. 2019, 21, 1108–1111. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Gavia, D.J.; Tang, Y. Biosynthesis of fungal indole alkaloids. Nat. Prod. Rep. 2014, 31, 1474–1487. [Google Scholar] [CrossRef] [Green Version]
- Gubiani, J.R.; Oliveira, M.C.S.; Neponuceno, R.A.R.; Camargo, M.J.; Garcez, W.S.; Biz, A.R.; Soares, M.A.; Araujo, A.R.; Bolzani, S.; Lisboa, H.C.F.; et al. Cytotoxic prenylated indole alkaloid produced by the endophytic fungus Aspergillus terreus P63. Phytochem. Lett. 2019, 32, 162–167. [Google Scholar] [CrossRef]
- Zhang, P.; Li, X.-M.; Liu, H.; Li, X.; Wang, B.-G. Two new alkaloids from Penicillium oxalicum EN-201, an endophytic fungus derived from the marine mangrove plant Rhizophora stylosa. Phytochem. Lett. 2015, 13, 160–164. [Google Scholar] [CrossRef]
- Yu, G.; Zhou, G.; Zhu, M.; Wang, W.; Zhu, T.; Gu, Q.; Li, D. Neosartoryadins A and B, Fumiquinazoline Alkaloids from a Mangrove-Derived Fungus Neosartorya udagawae HDN13-313. Org. Lett. 2016, 18, 244–247. [Google Scholar] [CrossRef]
- Zhang, P.; Li, X.-M.; Li, X.; Wang, B.-G. New indole-diterpenoids from the algal-associated fungus Aspergillus nidulans. Phytochem. Lett. 2015, 12, 182–185. [Google Scholar] [CrossRef]
- Gao, S.-S.; Li, X.-M.; Williams, K.; Proksch, P.; Ji, N.-Y.; Wang, B.-G. Rhizovarins A−F, Indole-Diterpenes from the Mangrove-Derived Endophytic Fungus Mucor irregularis QEN-189. J. Nat. Prod. 2016, 79, 2066–2074. [Google Scholar] [CrossRef]
- Prakash, C.R.; Raja, S. Indolinones as Promising Scaffold as Kinase Inhibitors: A Review. Mini Rev. Med. Chem. 2012, 12, 98–119. [Google Scholar] [CrossRef] [PubMed]
- Leoni, A.; Locatelli, A.; Morigi, R.; Rambaldi, M. 2-Indolinone a versatile scaffold for treatment of cancer: A patent review (2008–2014). Expert Opin. Ther. Pat. 2016, 3776, 149–173. [Google Scholar] [CrossRef]
- Speck, K.; Magauer, T. The chemistry of isoindole natural products. Beilstein J. Org. Chem. 2013, 9, 2048–2078. [Google Scholar] [CrossRef]
- Zhou, H.; Sun, X.; Li, N.; Che, Q.; Zhu, T.; Gu, Q.; Li, D. Isoindolone-Containing Meroperpenoids from the Endophytic Fungus Emericella nidulans HDN12-249. Org. Lett. 2016, 18, 4670–4673. [Google Scholar] [CrossRef]
- Cui, H.; Lin, Y.; Luo, M.; Lu, Y.; Huang, X.; She, Z. Diaporisoindoles A−C: Three Isoprenylisoindole Alkaloid Derivatives from the Mangrove Endophytic Fungus Diaporthe sp. SYSU-HQ3. Org. Lett. 2017, 19, 5621–5624. [Google Scholar] [CrossRef]
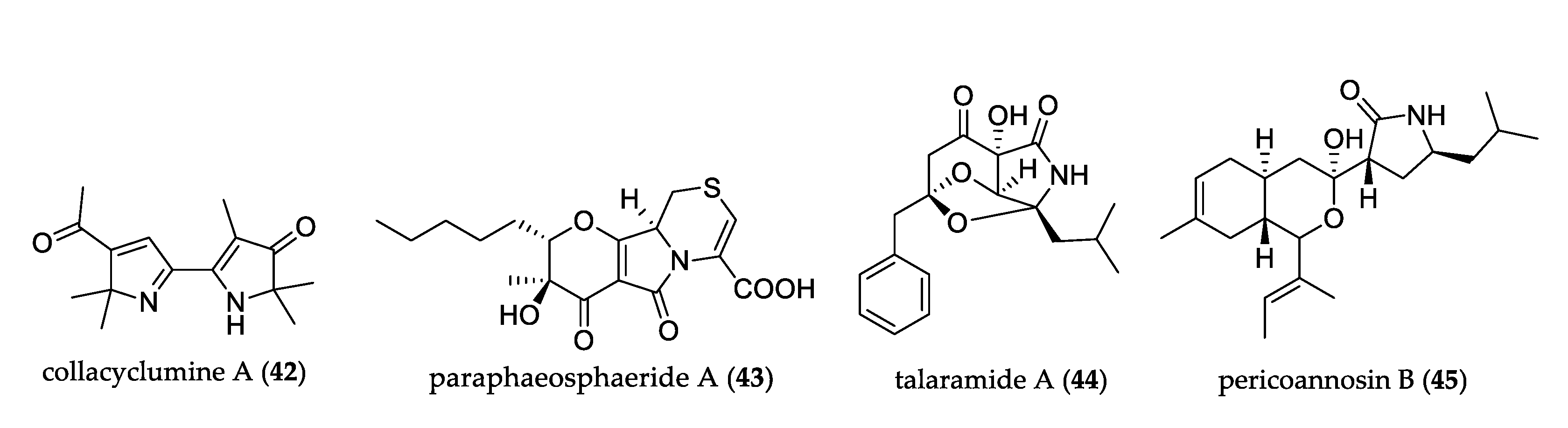
- Lin, X.; Ai, W.; Li, M.; Zhou, X.; Liao, S.; Wang, J.; Liu, J.; Yang, B.; Liu, Y. Collacyclumines A–D from the endophytic fungus Colletotrichum salsolae SCSIO 41021 isolated from the mangrove Kandelia candel. Phytochemistry 2020, 171, 112237. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-S.; Ding, Y.; Yang, B.; Miklossy, G.; Yin, H.-Q.; Walker, L.A.; Turkson, J.; Cao, S. A New Metabolite with a Unique 4-Pyranone-γ-Lactam-1,4-Thiazine Moiety from a Hawaiian-Plant Associated Fungus. Org. Lett. 2015, 17, 3556–3559. [Google Scholar] [CrossRef]
- Chen, S.; He, L.; Chen, D.; Cai, R.; Long, Y.; Lu, Y.; She, Z. Talaramide A, an unusual alkaloid from the mangrove endophytic fungus: Talaromyces sp. (Hz-YX1) as an inhibitor of mycobacterial PknG. New J. Chem. 2017, 41, 4273–4276. [Google Scholar] [CrossRef]
- Zhang, D.W.; Tao, X.Y.; Liu, J.M.; Chen, R.D.; Zhang, M.; Fang, X.M.; Yu, L.Y.; Dai, J.G. A new polyketide synthase-nonribosomal peptide synthetase hybrid metabolite from plant endophytic fungus Periconia sp. Chin. Chem. Lett. 2016, 27, 640–642. [Google Scholar] [CrossRef]
- Qiao, Y.; Xu, Q.; Feng, W.; Tao, L.; Li, X.-N.; Liu, J.; Zhu, H.; Lu, Y.; Wang, J.; Qi, C.; et al. Asperpyridone A: An Unusual Pyridone Alkaloid Exerts Hypoglycemic Activity through the Insulin Signaling Pathway. J. Nat. Prod. 2019, 82, 2925–2930. [Google Scholar] [CrossRef]
- Zhu, M.; Zhang, X.; Feng, H.; Che, Q.; Zhu, T.; Gu, Q.; Li, D. Campyridones A–D, pyridone alkaloids from a mangrove endophytic fungus Campylocarpon sp. HDN13-307. Tetrahedron 2016, 72, 5679–5683. [Google Scholar] [CrossRef]
- Borthwick, A.D. 2,5-Diketopiperazines: Synthesis, Reactions, Medicinal Chemistry, and Bioactive Natural Products. Chem. Rev. 2012, 112, 3641–3716. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Y.; Zhang, X.; Lai, D.; Zhou, L. Structural Diversity and Biological Activities of the Cyclodipeptides from Fungi. Molecules 2017, 22, 2026. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Li, X.-M.; Meng, L.-H.; Konuklugil, B.; Li, X.; Li, H.-L.; Wang, B.-G. Isolation and characterization of three pairs of indolediketopiperazine enantiomers containing infrequent N-methoxy substitution from the marine algal-derived endophytic fungus Acrostalagmus luteoalbus TK-43. Bioorg. Chem. 2019, 90, 103030. [Google Scholar] [CrossRef]
- Cai, R.; Jiang, H.; Xiao, Z.; Cao, W.; Yan, T.; Liu, Z.; Lin, S.; Long, Y.; She, Z. (−)- And (+)-Asperginulin A, a Pair of Indole Diketopiperazine Alkaloid Dimers with a 6/5/4/5/6 Pentacyclic Skeleton from the Mangrove Endophytic Fungus Aspergillus sp. SK-28. Org. Lett. 2019, 21, 9633–9636. [Google Scholar] [CrossRef]
- Lhamo, S.; Wang, X.B.; Li, T.X.; Wang, Y.; Li, Z.R.; Shi, Y.M.; Yang, M.H.; Kong, L.Y. Three unusual indole diketopiperazine alkaloids from a terrestrial-derived endophytic fungus, Aspergillus sp. Tetrahedron Lett. 2015, 56, 2823–2826. [Google Scholar] [CrossRef]
- Zhang, P.; Li, X.-M.; Wang, J.-N.; Li, X.; Wang, B.-G. Prenylated indole alkaloids from the marine-derived fungus Paecilomyces variotii. Chin. Chem. Lett. 2015, 26, 313–316. [Google Scholar] [CrossRef]
- Meng, L.H.; Wang, C.Y.; Mándi, A.; Li, X.M.; Hu, X.Y.; Kassack, M.U.; Kurtán, T.; Wang, B.G. Three Diketopiperazine Alkaloids with Spirocyclic Skeletons and One Bisthiodiketopiperazine Derivative from the Mangrove-Derived Endophytic Fungus Penicillium brocae MA-231. Org. Lett. 2016, 18, 5304–5307. [Google Scholar] [CrossRef] [PubMed]
- Barakat, F.; Vansteelandt, M.; Triastuti, A.; Jargeat, P.; Jacquemin, D.; Graton, J.; Mejia, K.; Cabanillas, B.; Vendier, L.; Stigliani, J.L.; et al. Thiodiketopiperazines with two spirocyclic centers extracted from Botryosphaeria mamane, an endophytic fungus isolated from Bixa orellana L. Phytochemistry 2019, 158, 142–148. [Google Scholar] [CrossRef]
- Zhu, M.; Zhang, X.; Feng, H.; Dai, J.; Li, J.; Che, Q.; Gu, Q.; Zhu, T.; Li, D. Penicisulfuranols A–F, alkaloids from the mangrove endophytic fungus Penicillium janthinellum HDN13-309. J. Nat. Prod. 2017, 80, 71–75. [Google Scholar] [CrossRef]
- Kajula, M.; Ward, J.M.; Turpeinen, A.; Tejesvi, M.V.; Hokkanen, J.; Tolonen, A.; Häkkänen, H.; Picart, P.; Ihalainen, J.; Sahl, H.G.; et al. Bridged Epipolythiodiketopiperazines from Penicillium raciborskii, an Endophytic Fungus of Rhododendron tomentosum Harmaja. J. Nat. Prod. 2016, 79, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Kong, Z.; Jing, R.; Wu, Y.; Guo, Y.; Geng, Y.; Ji, J.; Qin, L.; Zheng, C. Trichodermadiones A and B from the solid culture of Trichoderma atroviride S361, an endophytic fungus in Cephalotaxus fortunei. Fitoterapia 2018, 127, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Ren, F.; Che, Y.; Liu, G.; Liu, L. New Bergamotane Sesquiterpenoids from the Plant Endophytic Fungus Paraconiothyrium brasiliense. Molecules 2015, 20, 14611–14620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Niu, S.-B.; Li, L.; Ding, G.; Yu, M.; Zhang, G.-S.; Wang, M.-H.; Li, L.-Y.; Zhang, T.; Jia, H.-M.; et al. Trichoderpyrone, a Unique Polyketide Hybrid with a Cyclopentenone—Pyrone Skeleton from the Plant Endophytic Fungus Trichoderma gamsii. J. Nat. Prod. 2017, 80, 1944–1947. [Google Scholar] [CrossRef] [PubMed]
- Elsebai, M.F.; Ghabbour, H.A.; Mehiri, M. Unusual Nitrogenous Phenalenone Derivatives from the Marine-Derived Fungus Coniothyrium cereale. Molecules 2016, 21, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Kusari, S.; Golz, C.; Laatsch, H.; Strohmann, C.; Spiteller, M. Epigenetic Modulation of Endophytic Eupenicillium sp. LG41 by a Histone Deacetylase Inhibitor for Production of Decalin-Containing Compounds. J. Nat. Prod. 2017, 80, 983–988. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.-W.; Lv, M.-M.; Li, X.-S.; Zhang, L.; Liu, C.X.; Guo, Z.-Y.; Deng, Z.-S.; Zou, K.; Proksch, P. Penicitroamide, an antimicrobial metabolite with high carbonylization from the endophytic fungus Penicillium sp. (NO. 24). Molecules 2016, 21, 1438. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Sun, Z.-H.; Chen, Y.-C.; Liu, H.-X.; Li, H.-H.; Tan, G.-H.; Li, S.-N.; Guo, X.-L.; Zhang, W.-M. Cytotoxic Cochlioquinone Derivatives from the Endophytic Fungus Bipolaris sorokiniana Derived from Pogostemon cablin. Fitoterapia 2016, 110, 77–82. [Google Scholar] [CrossRef]
- Li, W.-S.; Hu, H.-B.; Huang, Z.-H.; Yan, R.-J.; Tian, L.-W.; Wu, J. Phomopsols A and B from the Mangrove Endophytic Fungus Phomopsis sp. xy21: Structures, Neuroprotective Effects, and Biogenetic Relationships. Org. Lett. 2019, 21, 7919–7922. [Google Scholar] [CrossRef]
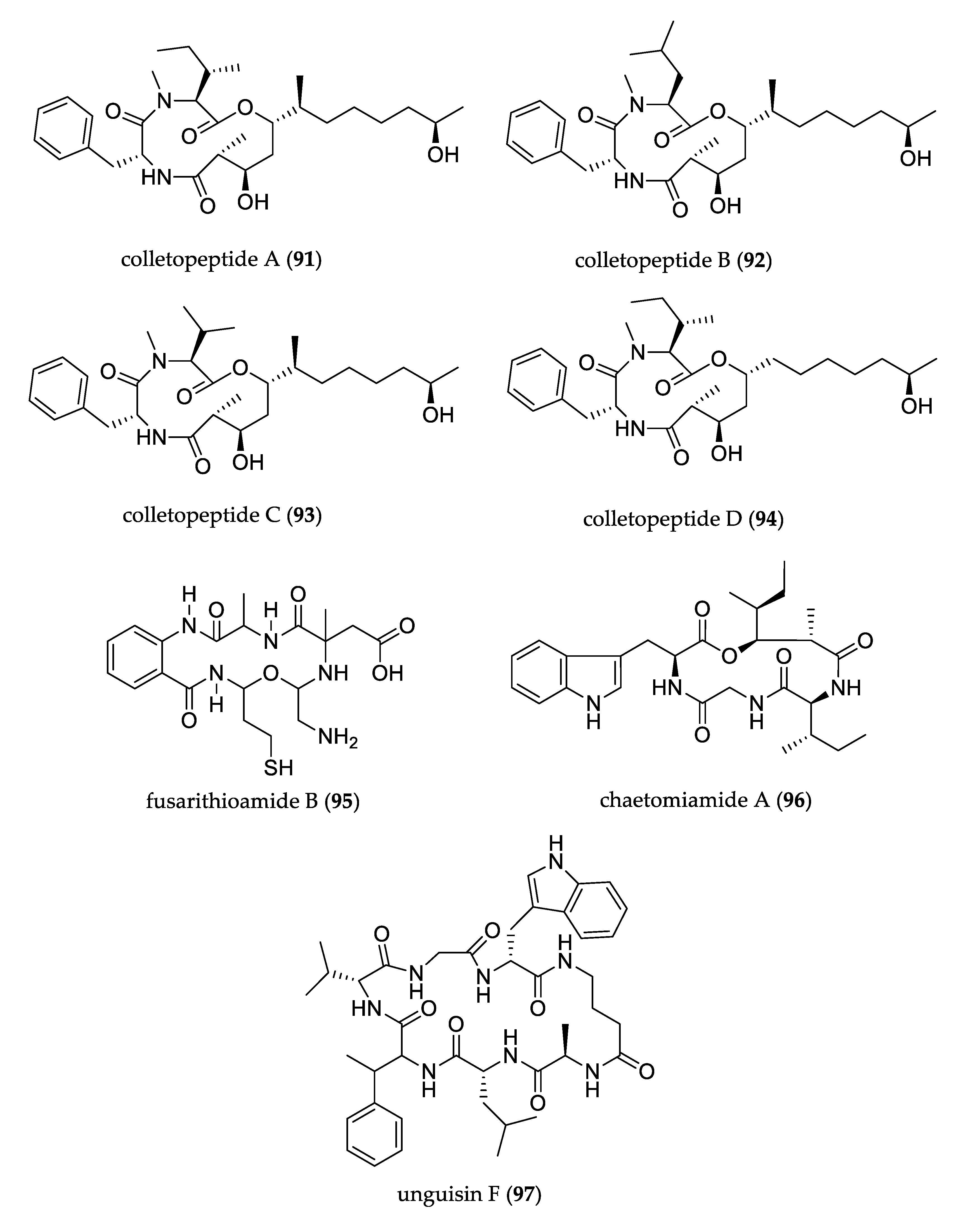
- Feng, L.; Wang, J.; Liu, S.; Zhang, X.-J.; Bi, Q.-R.; Hu, Y.-Y.; Wang, Z.; Tan, N.-H. Colletopeptides A–D, Anti-inflammatory Cyclic Tridepsipeptides from the Plant Endophytic Fungus Colletotrichum sp. S8. J. Nat. Prod. 2019, 82, 1434–1441. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Al Haidari, R.A.; Zayed, M.F.; El-Kholy, A.A.; Elkhayat, E.S.; Ross, S.A. Fusarithioamide B, a new benzamide derivative from the endophytic fungus Fusarium chlamydosporium with potent cytotoxic and antimicrobial activities. Bioorg. Med. Chem. 2018, 26, 786–790. [Google Scholar] [CrossRef]
- Wang, F.; Jiang, J.; Hu, S.; Ma, H.; Zhu, H.; Tong, Q.; Cheng, L.; Hao, X.; Zhang, G.; Zhang, Y. Secondary metabolites from endophytic fungus Chaetomium sp. induce colon cancer cell apoptotic death. Fitoterapia 2017, 121, 86–93. [Google Scholar] [CrossRef]
- Akone, S.H.; Daletos, G.; Lin, W.; Proksch, P. Unguisin F, a new cyclic peptide from the endophytic fungus Mucor irregularis. Z. Nat. C 2016, 71, 15–19. [Google Scholar] [CrossRef]
- Sun, L.-T.; Chen, Y.; Yang, H.-X.; Li, Z.-H.; Liu, J.-K.; Wang, G.-K.; Feng, T. Bisabolane sesquiterpenes and α-pyrone derivative from endophytic fungus Zopfiella sp. Phytochem. Lett. 2020, 37, 29–32. [Google Scholar] [CrossRef]
- Guo, Z.-Y.; Tan, M.-H.; Liu, C.-X.; Lv, M.-M.; Deng, Z.-S.; Cao, F.; Zou, K.; Proksch, P. Aspergoterpenins A–D: Four New Antimicrobial Bisabolane Sesquiterpenoid Derivatives from an Endophytic Fungus Aspergillus versicolor. Molecules 2018, 23, 1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wibowo, M.; Prachyawarakorn, V.; Aree, T.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Cytotoxic sesquiterpenes from the endophytic fungus Pseudolagarobasidium acaciicola. Phytochemistry 2016, 122, 126–138. [Google Scholar] [CrossRef]
- Xu, L.-L.; Chen, H.-L.; Hai, P.; Gao, Y.; Xie, C.-D.; Yang, X.-L.; Abe, I. (+)- and (−)-Preuisolactone A: A Pair of Caged Norsesquiterpenoidal Enantiomers with a Tricy-clo[4.4.01,6.02,8]decane Carbon Skeleton from the Endophytic Fungus Preussia isomera. Org. Lett. 2019, 21, 1078–1081. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Lin, L.-B.; Hu, J.-Y.; Duan, D.-Z.; Shi, W.; Zhang, Q.; Han, W.-B.; Wang, L.; Wang, X.-L. Pestalustaines A and B, unprecedented sesquiterpene and coumarin derivatives from endophytic fungus Pestalotiopsis adusta. Tetrahedron Lett. 2018, 59, 1772–1775. [Google Scholar] [CrossRef]
- Wang, Y.-N.; Xia, G.-Y.; Wang, L.-Y.; Ge, G.-B.; Zhang, H.-W.; Zhang, J.-F.; Wu, Y.-Z.; Lin, S. Purpurolide A, 5/5/5 Spirocyclic Sesquiterpene Lactone in Nature from the Endophytic Fungus Penicillium purpurogenum. Org. Lett. 2018, 20, 7341–7344. [Google Scholar] [CrossRef]
- Xie, S.; Wu, Y.; Qiao, Y.; Guo, Y.; Wang, J.; Hu, Z.; Zhang, Q.; Li, X.; Huang, J.; Zhou, Q.; et al. Protoilludane, Illudalane, and Botryane Sesquiterpenoids from the Endophytic Fungus Phomopsis sp. TJ507A. J. Nat. Prod. 2018, 81, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Tao, Y.; Tao, X.; Su, Q.; Cai, J.; Qin, C.; Ding, W.; Li, C. Sesquiterpenes with Phytopathogenic Fungi Inhibitory Activities from Fungus Trichoderma virens from Litchi chinensis Sonn. J. Agric. Food Chem. 2019, 67, 10646–10652. [Google Scholar] [CrossRef]
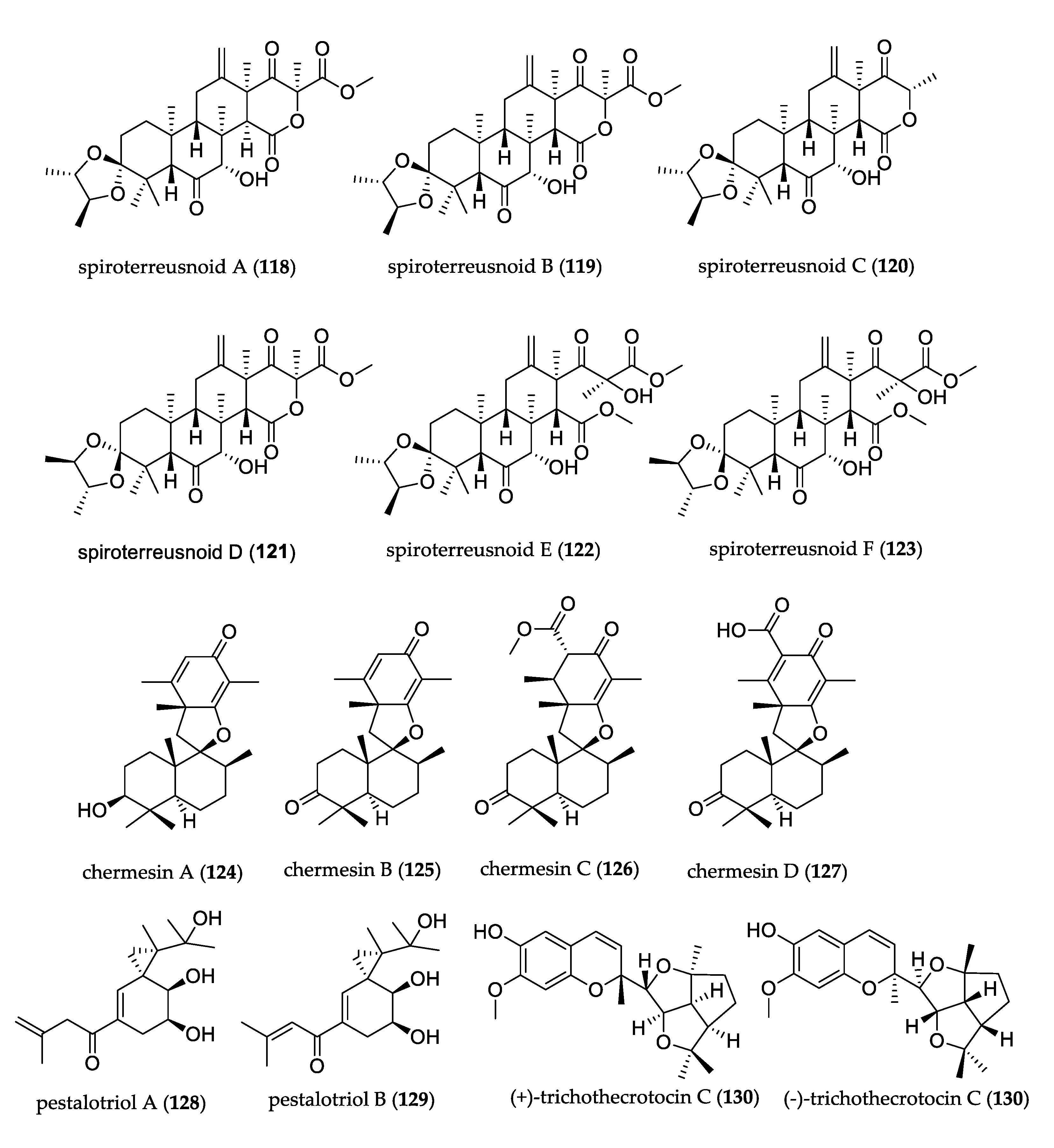
- Yang, H.-X.; Ai, H.-L.; Feng, T.; Wang, W.-X.; Wu, B.; Zheng, Y.-S.; Sun, H.; He, J.; Li, Z.-H.; Liu, J.-K. Trichothecrotocins A−C, Antiphytopathogenic Agents from Potato Endophytic Fungus Trichothecium crotocinigenum. Org. Lett. 2018, 20, 8069–8072. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, J.-M.; Zhao, J.-L.; Li, N.; Chen, R.-D.; Xie, K.-B.; Zhang, W.-J.; Feng, K.-P.; Yan, Z.; Wang, N.; et al. Two new diterpenoids from the endophytic fungus Trichoderma sp. Xy24 isolated from mangrove plant Xylocarpus granatum. Chin. Chem. Lett. 2016, 27, 957–960. [Google Scholar] [CrossRef]
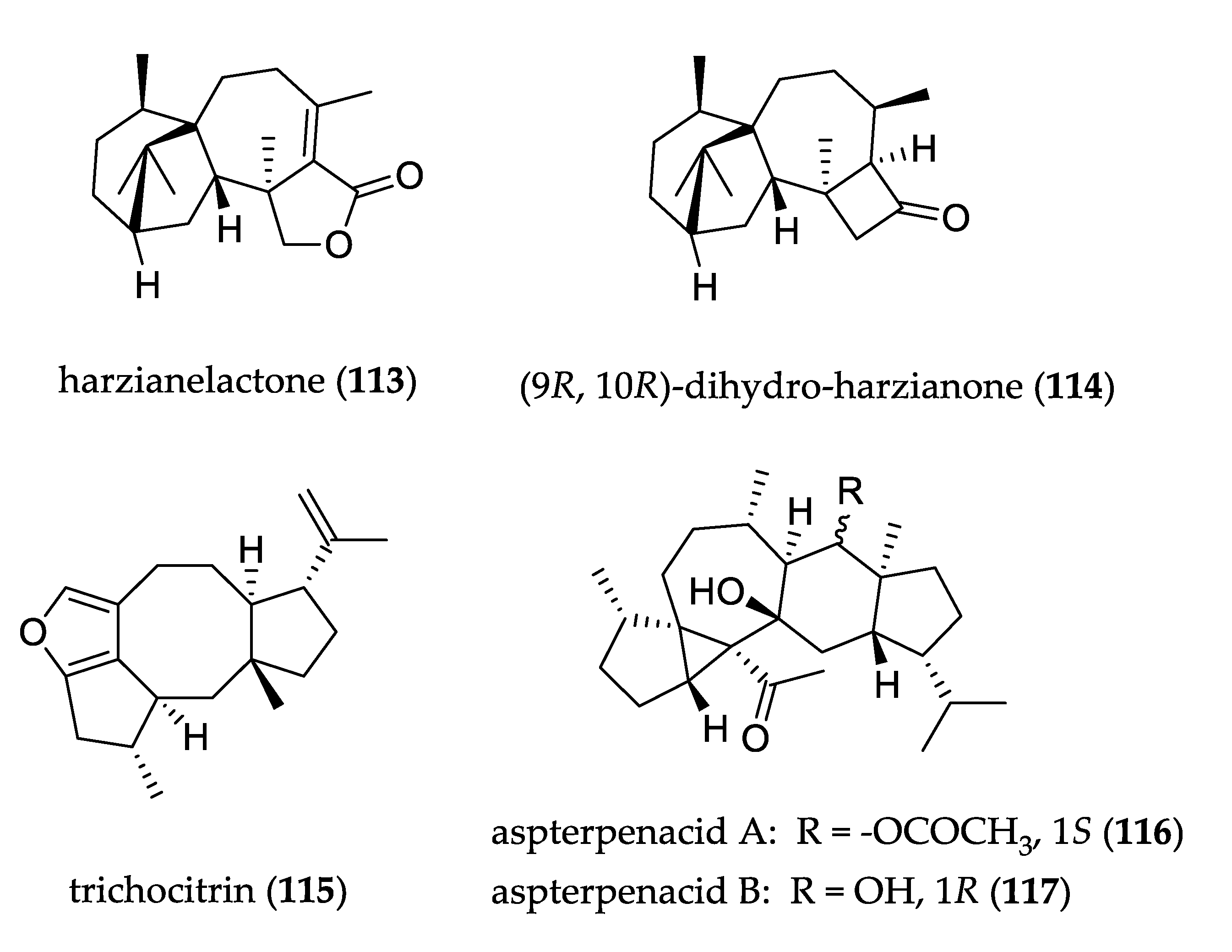
- Liang, X.-R.; Miao, F.-P.; Song, Y.-P.; Guo, Z.-Y.; Ji, N.-Y. Trichocitrin, a new fusicoccane diterpene from the marine brown alga-endophytic fungus Trichoderma citrinoviride cf-27. Nat. Prod. Res. 2016, 30, 1605–1610. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, Y.; Chen, S.; Liu, Y.; Lu, Y.; Chen, D.; Lin, Y.; Huang, X.; She, Z. Aspterpenacids A and B, Two Sesterterpenoids from a Mangrove Endophytic Fungus Aspergillus terreus H010. Org. Lett. 2016, 18, 1406–1409. [Google Scholar] [CrossRef] [PubMed]
- Qi, C.; Zhou, Q.; Gao, W.; Liu, M.; Chen, C.; Li, X.N.; Lai, Y.; Zhou, Y.; Li, D.; Hu, Z.; et al. Anti-BACE1 and anti-AchE activities of undescribed spiro-dioxolane-containing meroterpenoids from the endophytic fungus Aspergillus terreus Thom. Phytochemistry 2019, 165, 112041. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, X.-M.; Liu, Y.; Zhang, P.; Wang, J.-N.; Wang, B.-G. Chermesins A–D: Meroterpenoids with a Drimane-Type Spirosesquiterpene Skeleton from the Marine Algal-Derived Endophytic Fungus Penicillium chermesinum EN-480. J. Nat. Prod. 2016, 79, 806–811. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, C.; Li, L.; Guo, L.; Che, Y. Pestalotriols A and B, new spiro[2.5]octane derivatives from the endophytic fungus Pestalotiopsis fici. RSC Adv. 2015, 5, 78708–78711. [Google Scholar] [CrossRef]
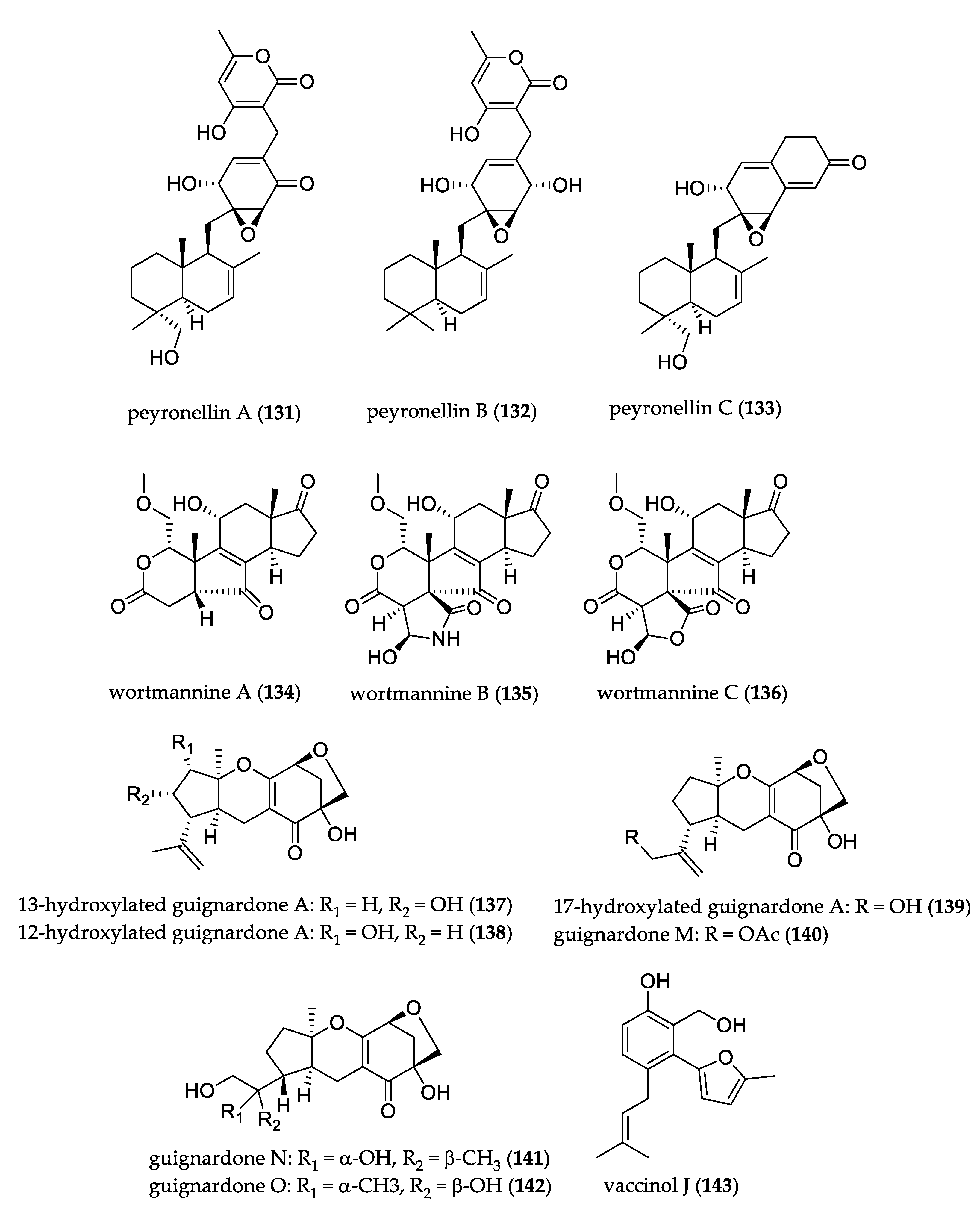
- Li, C.-S.; Ren, G.; Yang, B.-J.; Miklossy, G.; Turkson, J.; Fei, P.; Ding, Y.; Walker, L.A.; Cao, S. Meroterpenoids with Antiproliferative Activity from a Hawaiian-Plant Associated Fungus Peyronellaea coffeae-arabicae FT238. Org. Lett. 2016, 18, 2335–2338. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.-C.; Yang, Z.-D.; Zhou, S.-Y.; Li, X.-M.; Yu, H.-T.; Yao, X.-J.; Fang, J.-G.; Shu, Z.-M.; Xue, H.-Y.; Wang, Y.-G. Wortmannines A–C, three novel wortmannin derivatives with an unusual five-membered B ring from the endophytic fungus Talaromyces wortmannii LGT-4. Tetrahedron Lett. 2016, 57, 4608–4611. [Google Scholar] [CrossRef]
- Li, T.-X.; Yang, M.-H.; Wang, X.-B.; Wang, Y.; Kong, L.-Y. Synergistic Antifungal Meroterpenes and Dioxolanone Derivatives from the Endophytic Fungus Guignardia sp. J. Nat. Prod. 2015, 78, 2511–2520. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-F.; Liang, R.; Liao, S.-R.; Yang, B.; Tu, Z.-C.; Lin, X.-P.; Wang, B.-G.; Liu, Y. Vaccinols J–S, ten new salicyloid derivatives from the marine mangrove-derived endophytic fungus Pestalotiopsis vaccinii. Fitoterapia 2017, 120, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Zhao, J.-Y.; Fang, X.-M.; Zhang, T.; Zhang, D.-W.; Liu, H.-Y.; Su, J.; Cen, S.; Yu, L.-Y. Metabolites from the Plant Endophytic Fungus Aspergillus sp. CPCC 400735 and Their Anti-HIV Activities. J. Nat. Prod. 2017, 80, 2595–2601. [Google Scholar] [CrossRef] [PubMed]
- Gombodorj, S.; Yang, M.-H.; Shang, Z.-C.; Liu, R.-H.; Li, T.-X.; Yin, G.-P.; Kong, L.-Y. New phenalenone derivatives from Pinellia ternata tubers derived Aspergillus sp. Fitoterapia 2017, 120, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.-L.; Wei, W.-J.; Li, H.-Y.; Wang, L.-D.; Dong, S.-H.; Gao, K. Phomotide A, a novel polyketide, from the endophytic fungus Phomopsis sp. CFS42. Tetrahedron Lett. 2020, 61, 151468. [Google Scholar] [CrossRef]
- Tang, J.-W.; Xu, H.-C.; Wang, W.-G.; Hu, K.; Zhou, Y.-F.; Chen, R.; Li, X.-N.; Du, X.; Sun, H.-D.; Puno, P.-T. (+)- and (−)-Alternarilactone A: Enantiomers with a Diepoxy-Cage-like Scaffold from an Endophytic Alternaria sp. J. Nat. Prod. 2019, 82, 735–740. [Google Scholar] [CrossRef]
- Darsih, C.; Prachyawarakorn, V.; Wiyakrutta, S.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Cytotoxic metabolites from the endophytic fungus Penicillium chermesinum: Discovery of a cysteine-targeted Michael acceptor as a pharmacophore for fragment-based drug discovery, bioconjugation and click reactions. RSC Adv. 2015, 5, 70595–70603. [Google Scholar] [CrossRef]
- Sang, X.-N.; Chen, S.-F.; Chen, G.; An, X.; Li, S.-G.; Lu, X.-J.; Zhao, D.; Bai, J.; Wang, H.-F.; Pei, Y.-H. Two pairs of enantiomeric α-pyrone dimers from the endophytic fungus Phoma sp. YN02-P-3. RSC Adv. 2017, 7, 1943–1946. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Zhou, K.; He, P.; Wang, K.-M.; Zhu, R.-Z.; Wang, Y.-D.; Dong, W.; Li, G.-P.; Yang, H.-Y.; Ye, Y.-Q.; et al. Antiviral and Cytotoxic Isocoumarin Derivatives from an Endophytic Fungus Aspergillus oryzae. Planta Med. 2016, 82, 414–417. [Google Scholar] [CrossRef] [Green Version]
- Cai, R.; Wu, Y.; Chen, S.; Cui, H.; Liu, Z.; Li, C.; She, Z. Peniisocoumarins A-J: Isocoumarins from Penicillium commune QQF-3, an Endophytic Fungus of the Mangrove Plant Kandelia candel. J. Nat. Prod. 2018, 81, 1376–1383. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.-J.; Zhang, S.-B.; Chen, H.-L.; Zhao, W.-T.; Yang, D.-F.; Xian, P.-J.; Xu, L.-L.; Tao, Y.-D.; Fu, H.-Y.; Yang, X.-L. Emericelactones A-D: Four novel polyketides produced by Emericella sp. XL 029, a fungus associated the leaves of Panax notoginseng. Tetrahedron Lett. 2018, 59, 4566–4570. [Google Scholar] [CrossRef]
- Shiono, Y.; Miyazaki, N.; Murayama, T.; Koseki, T.; Harizon; Katja, D.G.; Supratman, U.; Nakata, J.; Kakihara, Y.; Saeki, M.; et al. GSK-3B inhibitory activities of novel dichroloresorcinol derivatives from Cosmospora vilior isolated from a mangrove plant. Phytochem. Lett. 2016, 18, 122–127. [Google Scholar] [CrossRef]
- Nalli, Y.; Mirza, D.N.; Wani, Z.A.; Wadhwa, B.; Mallik, F.A.; Raina, C.; Chaubey, A.; Riyaz-Ul-Hassan, S.; Ali, A. Phialomustin A–D, new antimicrobial and cytotoxic metabolites from an endophytic fungus, Phialophora mustea. RSC Adv. 2015, 5, 95307–95312. [Google Scholar] [CrossRef]
- Li, T.X.; Liu, R.H.; Wang, X.B.; Luo, J.; Luo, J.G.; Kong, L.Y.; Yang, M.H. Hypoxia-Protective Azaphilone Adducts from Peyronellaea glomerata. J. Nat. Prod. 2018, 81, 1148–1153. [Google Scholar] [CrossRef]
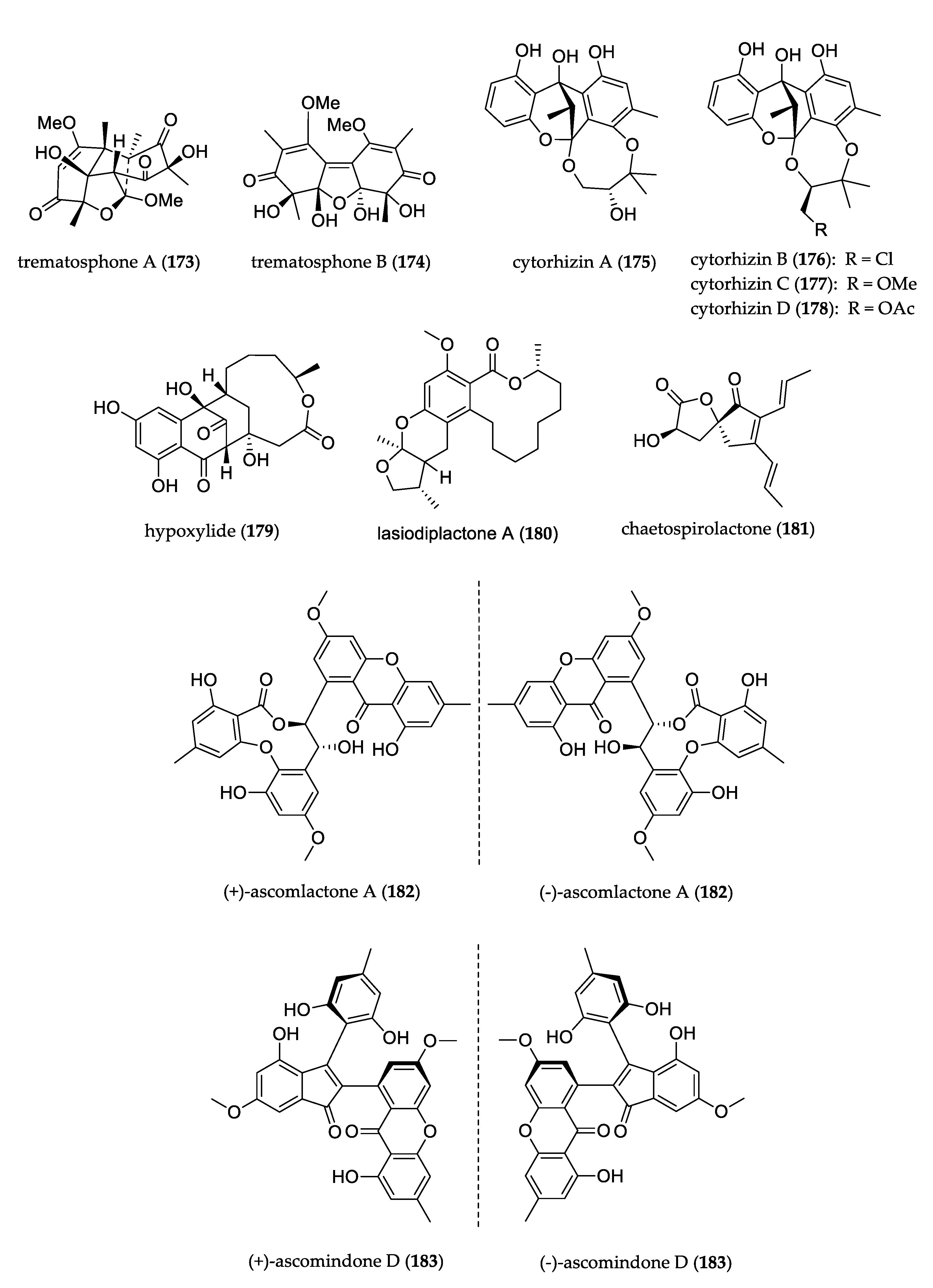
- Song, B.; Li, L.-Y.; Shang, H.; Liu, Y.; Yu, M.; Ding, G.; Zou, Z.-M. Trematosphones A and B, two unique dimeric structures from the desert plant endophytic fungus Trematosphaeria terricola. Org. Lett. 2019, 21, 2139–2142. [Google Scholar] [CrossRef]
- Liu, H.; Tan, H.; Chen, Y.; Guo, X.; Wang, W.; Guo, H.; Liu, Z.; Zhang, W. Cytorhizins A–D, Four Highly Structure-Combined Benzophenones from the Endophytic Fungus Cytospora rhizophorae. Org. Lett. 2019, 21, 1063–1067. [Google Scholar] [CrossRef]
- Liu, Y.; Kurtán, T.; Mándi, A.; Weber, H.; Wang, C.; Hartmann, R.; Lin, W.; Daletos, G.; Proksch, P. A novel 10-membered macrocyclic lactone from the mangrove-derived endophytic fungus Annulohypoxylon sp. Tetrahedron Lett. 2018, 59, 632–636. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Liu, Z.; Liu, H.; Long, Y.; Chen, D.; Lu, Y.; She, Z. Lasiodiplactone A, a novel lactone from the mangrove endophytic fungus Lasiodiplodia theobromae ZJ-HQ1. Org. Biomol. Chem. 2017, 15, 6338–6341. [Google Scholar] [CrossRef]
- Xu, Q.-L.; Xiao, Y.-S.; Shen, Y.; Wu, H.-M.; Zhang, X.; Deng, X.-Z.; Wang, T.-T.; Li, W.; Tan, R.-X.; Jiao, R.-H.; et al. Novel chaetospirolactone and orsellide F from an endophytic fungus Chaetomium sp. J. Asian Nat. Prod. Res. 2018, 20, 234–241. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, S.; Qiu, P.; Tan, C.; Long, Y.; Lu, Y.; She, Z. (+)- and (−)-Ascomlactone A, a pair of novel dimeric polyketides from a mangrove endophytic fungus Ascomycota sp. SK2YWS-L. Org. Biomol. Chem. 2017, 15, 10276–10280. [Google Scholar] [CrossRef]
- Liu, Z.; Qiu, P.; Li, J.; Chen, G.; Chen, Y.; Liu, H.; She, Z. Anti-inflammatory polyketides from the mangrove-derived fungus Ascomycota sp. SK2YWS-L. Tetrahedron 2018, 74, 746–751. [Google Scholar] [CrossRef]
- Liu, L.; Han, Y.; Xiao, J.; Li, L.; Guo, L.; Jiang, X.; Kong, L.; Che, Y. Chlorotheolides A and B, Spiroketals Generated via Diels−Alder Reactions in the Endophytic Fungus Pestalotiopsis theae. J. Nat. Prod. 2016, 79, 2616–2623. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Grimaldo, M.; Rivero-Cruz, I.; Madariaga-Mazón, A.; Figueroa, M.; Mata, R. α-Glucosidase Inhibitors from Preussia minimoides. J. Nat. Prod. 2017, 80, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, W.; El-Neketi, M.; Lewald, L.I.; Orfali, R.S.; Lin, W.; Rehberg, N.; Kalscheuer, R.; Daletos, G.; Proksch, P. Metabolites from the Fungal Endophyte Aspergillus austroafricanus in Axenic Culture and in Fungal—Bacterial Mixed Cultures. J. Nat. Prod. 2016, 79, 914–922. [Google Scholar] [CrossRef]
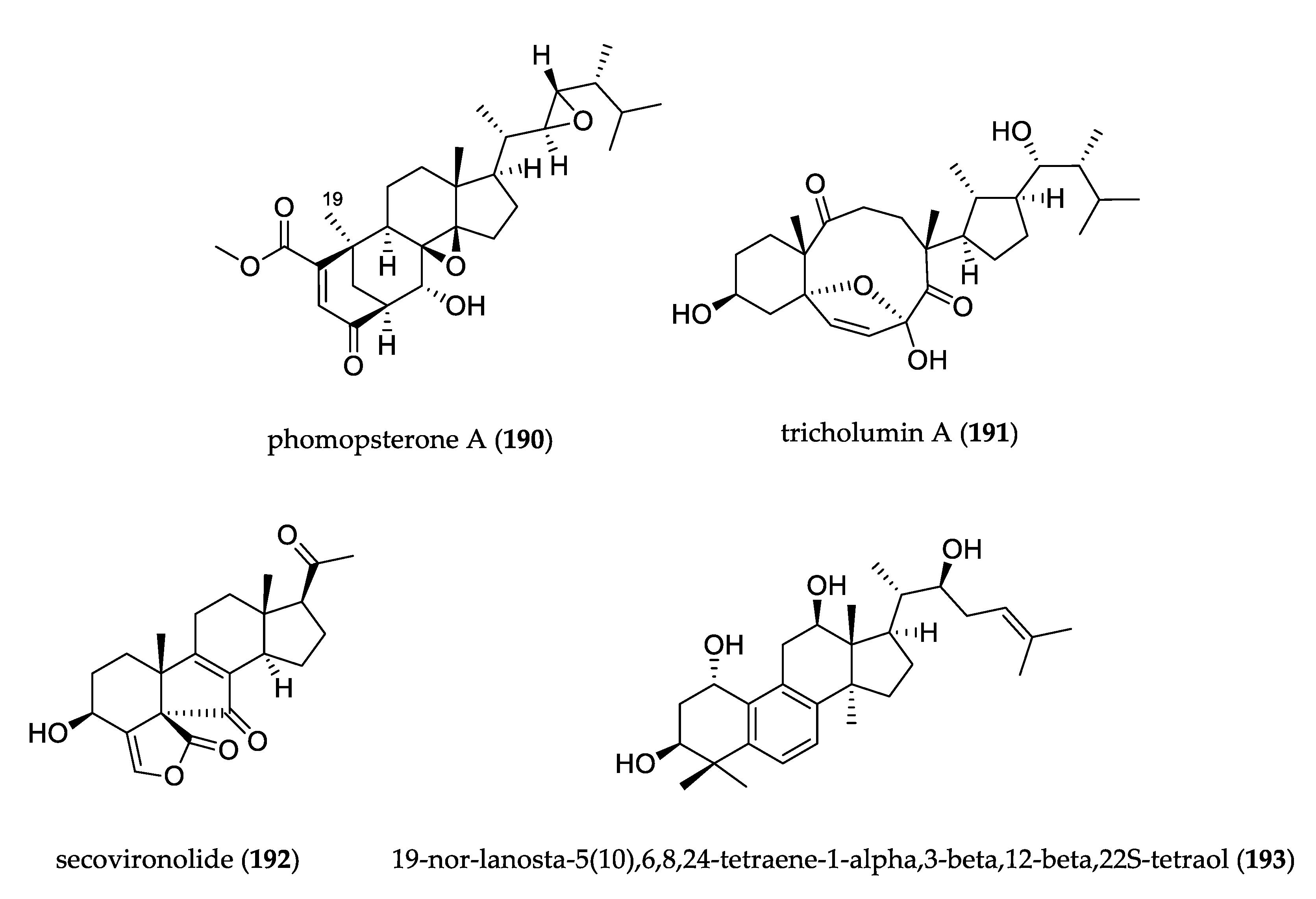
- Hu, Z.; Wu, Y.; Xie, S.; Sun, W.; Guo, Y.; Li, X.-N.; Liu, J.; Li, H.; Wang, J.; Luo, Z.; et al. Phomopsterones A and B, two functionalized ergostane-type steroids from the endophytic fungus Phomopsis sp. TJ507A. Org. Lett. 2017, 19, 258–261. [Google Scholar] [CrossRef]
- Song, Y.-P.; Shi, Z.-Z.; Miao, F.-P.; Fang, S.-T.; Yin, X.-L.; Ji, N.-Y. Tricholumin A, a Highly Transformed Ergosterol Derivative from the Alga-Endophytic Fungus Trichoderma asperellum. Org. Lett. 2018, 20, 6306–6309. [Google Scholar] [CrossRef]
- Ding, H.-E.; Yang, Z.-D.; Sheng, L.; Zhou, S.-Y.; Li, S.; Yao, X.-J.; Zhi, K.-K.; Wang, Y.-G.; Zhang, F. Secovironolide, a novel furanosteroid scaffold with a five-membered B ring from the endophytic fungus Talaromyces wortmannii LGT-4. Tetrahedron Lett. 2015, 56, 6754–6757. [Google Scholar] [CrossRef]
- Li, G.; Kusari, S.; Kusari, P.; Kayser, O.; Spiteller, M. Endophytic Diaporthe sp. LG23 Produces a Potent Antibacterial Tetracyclic Triterpenoid. J. Nat. Prod. 2015, 78, 2128–2132. [Google Scholar] [CrossRef]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions Through Comparative Studies of Nucleotides Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.-W.; Wang, S.-W.; Wu, B.; Wei, J.-G. Bioactive natural compounds from mangrove endophytic fungi. Mini Rev. Med. Chem. 2014, 14, 370–391. [Google Scholar] [CrossRef] [PubMed]





















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortega, H.E.; Torres-Mendoza, D.; Caballero E., Z.; Cubilla-Rios, L. Structurally Uncommon Secondary Metabolites Derived from Endophytic Fungi. J. Fungi 2021, 7, 570. https://doi.org/10.3390/jof7070570
Ortega HE, Torres-Mendoza D, Caballero E. Z, Cubilla-Rios L. Structurally Uncommon Secondary Metabolites Derived from Endophytic Fungi. Journal of Fungi. 2021; 7(7):570. https://doi.org/10.3390/jof7070570
Chicago/Turabian StyleOrtega, Humberto E., Daniel Torres-Mendoza, Zuleima Caballero E., and Luis Cubilla-Rios. 2021. "Structurally Uncommon Secondary Metabolites Derived from Endophytic Fungi" Journal of Fungi 7, no. 7: 570. https://doi.org/10.3390/jof7070570