
Characterization of the Highly Efficient Acid-Stable Xylanase and β-Xylosidase System from the Fungus Byssochlamys spectabilis ATHUM 8891 (Paecilomyces variotii ATHUM 8891)

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Growth Conditions
2.2. Enzyme Purification
2.3. Enzyme Assays and Protein Concentration
2.4. Biochemical Characterization
2.5. Hydrolysis Experiments
2.6. Nano LC ESI-MS/MS Analysis and Protein Identification
2.7. Statistical Analysis
3. Results
3.1. Enzyme Production and Purification
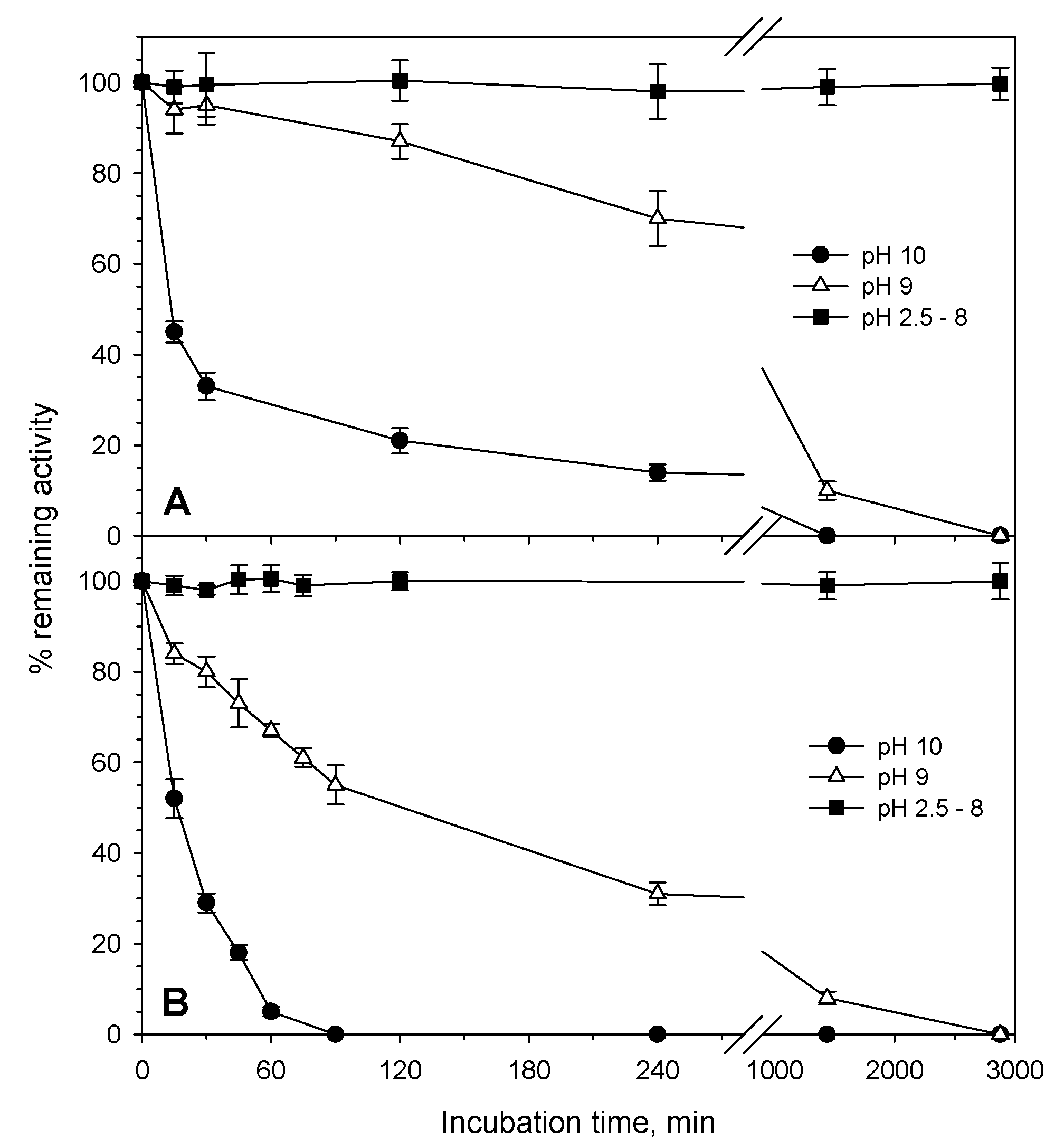
3.2. Activity and Stability Properties
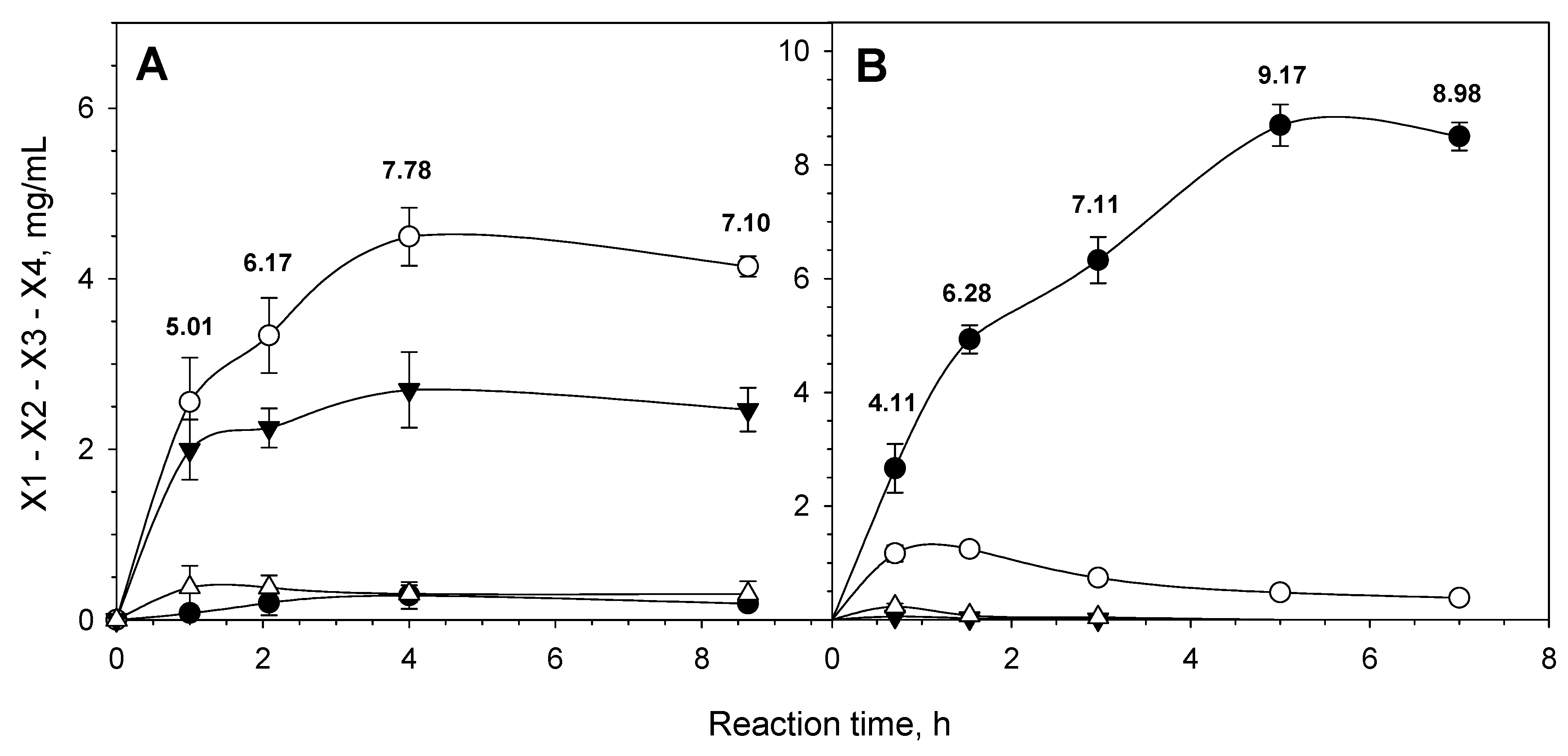
3.3. Substrate Specificity and Action on Xylan
3.4. Sequence Determination: Taxonomic Distribution and CAZy Classification
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guerriero, G.; Hausman, J.-F.; Strauss, J.; Ertan, H.; Siddiqui, K.S. Destructuring plant biomass: Focus on fungal and extremophilic cell wall hydrolases. Plant Sci. 2015, 234, 180–193. [Google Scholar] [CrossRef] [Green Version]
- Saha, B.C. Hemicellulose bioconversion. J. Ind. Microbiol. Biotechnol. 2003, 30, 279–291. [Google Scholar] [CrossRef]
- Subramaniyan, S.; Prema, P. Biotechnology of microbial xylanases: Enzymology, molecular biology, and application. Crit. Rev. Biotechnol. 2002, 22, 33–64. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Microbial xylanases: Engineering, production and industrial applications. Biotechnol. Adv. 2012, 30, 1219–1227. [Google Scholar] [CrossRef]
- Bhattacharya, A.S.; Bhattacharya, A.; Pletschke, B.I. Synergism of fungal and bacterial cellulases and hemicellulases: A novel perspective for enhanced bio-ethanol production. Biotechnol. Lett. 2015, 37, 1117–1129. [Google Scholar] [CrossRef] [PubMed]
- Shallom, D.; Shoham, Y. Microbial hemicellulases. Curr. Opin. Microbiol. 2003, 6, 219–228. [Google Scholar] [CrossRef]
- Olson, D.G.; McBride, J.E.; Shaw, A.J.; Lynd, L.R. Recent progress in consolidated bioprocessing. Curr. Opin. Biotechnol. 2012, 23, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.S.; Nugent, B.; Mullins, E.; Doohan, F.M. Fungal-mediated consolidated bioprocessing: The potential of Fusarium oxysporum for the lignocellulosic ethanol industry. AMB Express 2016, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galanopoulou, A.P.; Hatzinikolaou, D.G. Fungi in consolidated bioprocessing of lignocellulosic materials. In Fungal Applications in Sustainable Environmental Biotechnology; Purchase, D., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 275–305. [Google Scholar] [CrossRef]
- Amore, A.; Faraco, V. Potential of fungi as category I Consolidated BioProcessing organisms for cellulosic ethanol production. Renew. Sustain. Energy Rev. 2012, 16, 3286–3301. [Google Scholar] [CrossRef]
- Kotzekidou, P. Byssochlamys. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Oxford, UK, 2014; pp. 344–350. [Google Scholar] [CrossRef]
- Moreira, D.C.; Oliveira, M.M.E.; Borba, C.M. Human pathogenic Paecilomyces from food. Microorganisms 2018, 6, 64. [Google Scholar] [CrossRef] [Green Version]
- Herrera Bravo de Laguna, I.; Toledo Marante, F.J.; Mioso, R. Enzymes and bioproducts produced by the ascomycete fungus Paecilomyces variotii. J. Appl. Microbiol. 2015, 119, 1455–1466. [Google Scholar] [CrossRef] [Green Version]
- Mioso, R.; Marante, F.J.T.; de Laguna, I.H.B. The chemical diversity of the Ascomycete Fungus Paecilomyces variotii. Appl. Biochem. Biotechnol. 2015, 177, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Giannoutsou, E.P.; Katsifas, E.A.; Geli, A.; Karagouni, A.D. Protein increase and lysine production by a Paecilomyces variotii strain grown on two-phase olive mill waste. World J. Microbiol. Biotechnol. 2012, 28, 849–856. [Google Scholar] [CrossRef]
- de Morais, T.P.; Barbosa, P.M.G.; Garcia, N.F.L.; da Rosa-Garzon, N.G.; Fonseca, G.G.; da Paz, M.F.; Cabral, H.; Leite, R.S.R. Catalytic and thermodynamic properties of β-glucosidases produced by Lichtheimia corymbifera and Byssochlamys spectabilis. Prep. Biochem. Biotechnol. 2018, 48, 777–786. [Google Scholar] [CrossRef]
- Zerva, A.; Savvides, A.L.; Katsifas, E.A.; Karagouni, A.D.; Hatzinikolaou, D.G. Evaluation of Paecilomyces variotii potential in bioethanol production from lignocellulose through consolidated bioprocessing. Bioresour. Technol. 2014, 162, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.T.; O’Mahony, M.R.; Fogarty, W.M. Extracellular xylanolytic enzymes of Paecilomyces variotii. Biotechnol. Lett. 1989, 11, 885–890. [Google Scholar] [CrossRef]
- Krishnamurthy, S.; Vithayathil, P.J. Purification and characterization of endo-1,4-β-xylanase from Paecilomyces varioti Bainier. J. Ferment. Bioeng. 1989, 67, 77–82. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Wiegmann, K.; Hensler, M.; Wöhlbrand, L.; Ulbrich, M.; Schomburg, D.; Rabus, R. Carbohydrate catabolism in Phaeobacter inhibens DSM 17395, a member of the marine roseobacter clade. Appl. Environ. Microbiol. 2014, 80, 4725–4737. [Google Scholar] [CrossRef] [Green Version]
- Kitts, P.A.; Church, D.M.; Thibaud-Nissen, F.; Choi, J.; Hem, V.; Sapojnikov, V.; Smith, R.G.; Tatusova, T.; Xiang, C.; Zherikov, A.; et al. Assembly: A resource for assembled genomes at NCBI. Nucleic Acids Res. 2016, 44, D73–D80. [Google Scholar] [CrossRef]
- Yin, Y.; Mao, X.; Yang, J.; Chen, X.; Mao, F.; Xu, Y. dbCAN: A web resource for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2012, 40, W445–W451. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Tian, H.; Cheng, Y.; Jiang, Z.; Yang, S. Purification and characterization of a thermostable cellulase-free xylanase from the newly isolated Paecilomyces themophila. Enzyme Microb. Technol. 2006, 38, 780–787. [Google Scholar] [CrossRef]
- Zhang, M.; Jiang, Z.; Yang, S.; Hua, C.; Li, L. Cloning and expression of a Paecilomyces thermophila xylanase gene in E. coli and characterization of the recombinant xylanase. Bioresour. Technol. 2010, 101, 688–695. [Google Scholar] [CrossRef]
- Fan, G.; Katrolia, P.; Jia, H.; Yang, S.; Yan, Q.; Jiang, Z. High-level expression of a xylanase gene from the thermophilic fungus Paecilomyces thermophila in Pichia pastoris. Biotechnol. Lett. 2012, 34, 2043–2048. [Google Scholar] [CrossRef]
- Li, X.; Tu, T.; Yao, B.; Xie, X.; Luo, H. A novel bifunctional xylanase/cellulase TcXyn10A from Thermoascus crustaceus JCM12803. Sheng Wu Gong Cheng Xue Bao 2018, 34, 1996–2006. [Google Scholar] [CrossRef]
- Tan, L.U.L.; Mayers, P.; Saddler, J.N. Purification and characterization of a thermostable xylanase from a thermophilic fungus Thermoascus aurantiacus. Can. J. Microbiol. 1987, 33, 689–692. [Google Scholar] [CrossRef]
- Kumar, P.R.; Eswaramoorthy, S.; Vithayathil, P.J.; Viswamitra, A.M.A. The Tertiary Structure at 1.59 A Resolution and the Proposed Amino Acid Sequence of a Family-11 Xylanase from the Thermophilic Fungus Paecilomyces varioti Bainier. J. Mol. Biol. 2000, 295, 581–593. [Google Scholar] [CrossRef]
- Yan, Q.J.; Wang, L.; Jiang, Z.Q.; Yang, S.Q.; Zhu, H.F.; Li, L.T. A xylose-tolerant β-xylosidase from Paecilomyces thermophila: Characterization and its co-action with the endogenous xylanase. Bioresour. Technol. 2008, 99, 5402–5410. [Google Scholar] [CrossRef] [PubMed]
- Teng, C.; Jia, H.; Yan, Q.; Zhou, P.; Jiang, Z. High-level expression of extracellular secretion of a β-xylosidase gene from Paecilomyces thermophila in Escherichia coli. Bioresour. Technol. 2011, 102, 1822–1830. [Google Scholar] [CrossRef] [PubMed]
- Juturu, V.; Wu, J.C. Heterologous expression of β-xylosidase gene from Paecilomyces thermophila in Pichia pastoris. World J. Microbiol. Biotechnol. 2013, 29, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, M.; Endou, A.; Okada, T.; Yamaoka, Y. Purification and characterization of β-xylosidase from Thermoascus sp. J. Ferment. Bioeng. 1998, 86, 403–405. [Google Scholar] [CrossRef]
- Wu, J.F.; Lastick, S.M.; Updegraff, D.M. Ethanol production from sugars derived from plant biomass by a novel fungus. Nature 1986, 321, 887–888. [Google Scholar] [CrossRef]
- Mustafa, G.; Kousar, S.; Rajoka, M.I.; Jamil, A. Molecular cloning and comparative sequence analysis of fungal β-xylosidases. AMB Express 2016, 6, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaro-Reyes, A.; García-Almendárez, B.E.; Vázquez-Mandujano, D.G.; Amaya-Llano, S.; Castaño-Tostado, E.; Guevara-González, R.G.; Loera, O.; Regalado, C. Homologue expression of a β-xylosidase from native Aspergillus niger. J. Ind. Microbiol. Biotechnol. 2011, 38, 1311–1319. [Google Scholar] [CrossRef]
- Terrasan, C.R.F.; Guisan, J.M.; Carmona, E.C. Xylanase and β-xylosidase from Penicillium janczewskii: Purification, characterization and hydrolysis of substrates. Elecron. J. Biotechnol. 2016, 23, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Wakiyama, M.; Yoshihara, K.; Hayashi, S.; Ohta, K. Purification and properties of an extracellular β-xylosidase from Aspergillus japonicus and sequence analysis of the encoding gene. J. Biosci. Bioeng. 2008, 106, 398–404. [Google Scholar] [CrossRef]
- Michelin, M.; Peixoto-Nogueira, S.C.; Silva, T.M.; Jorge, J.A.; Terenzi, H.F.; Teixeira, J.A.; Polizeli, M. de L.T.M. A novel xylan degrading β-D-xylosidase: Purification and biochemical characterization. World J. Microbiol. Biotechnol. 2012, 28, 3179–3186. [Google Scholar] [CrossRef] [Green Version]
- Knob, A.; Carmona, E.C. Purification and characterization of two extracellular xylanases from Penicillium sclerotiorum: A novel acidophilic xylanase. Appl. Biochem. Biotechnol. 2010, 162, 429–443. [Google Scholar] [CrossRef]
- Lee, J.-W.; Park, J.-Y.; Kwon, M.; Choi, I.-G. Purification and characterization of a thermostable xylanase from the brown-rot fungus Laetiporus sulphureus. J. Biosci. Bioeng. 2009, 107, 33–37. [Google Scholar] [CrossRef]
- Luo, H.; Li, J.; Yang, J.; Wang, H.; Yang, Y.; Huang, H.; Shi, P.; Yuan, T.; Fan, Y.; Yao, B. A thermophilic and acid stable family-10 xylanase from the acidophilic fungus Bispora sp. MEY-1. Extremophiles 2009, 13, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Sulistyo, J.; Kamiyama, Y.; Yasui, T. Purification and some properties of Aspergillus pulverulentus β-xylosidase with transxylosylation capacity. J. Ferment. Bioeng. 1995, 79, 17–22. [Google Scholar] [CrossRef]
- Iembo, T.; da Silva, R.; Pagnocca, F.C.; Gomes, E. Production, characterization, and properties of β-glucosidase and β-xylosidase from a strain of Aureobasidium sp. Appl. Biochem. Microbiol. 2002, 38, 549–552. [Google Scholar] [CrossRef]
- Ohta, K.; Fujimoto, H.; Fujii, S.; Wakiyama, M. Cell-associated β-xylosidase from Aureobasidium pullulans ATCC 20524: Purification, properties, and characterization of the encoding gene. J. Biosci. Bioeng. 2010, 110, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Knob, A.; Carmona, E.C. Purification and properties of an acid β-xylosidase from Penicillium sclerotiorum. Ann. Microbiol. 2011, 62, 501–508. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Xylanase | β-Xylosidase | ||||
|---|---|---|---|---|---|---|
| Specific Activity (nkat mg−1) | Enrichment | Recovery (%) | Specific Activity (nkat mg−1) | Enrichment | Recovery (%) | |
| Extracellular medium | 1259 | 1.0 | 100 | 67 | 1 | 100 |
| (NH4)2SO4 precipitation | 1418 | 1.1 | 92 | 89 | 1.3 | 88 |
| Q-Sepharose anion exchange | 3957 | 3.1 | 76 | 504 | 7.5 | 84 |
| SP-Sepharose cation exchange | 29,781 | 23.7 | 72 | 3359 | 50.1 | 62 |
| Sephacryl S-200 gel filtration | 75,847 | 60.2 | 62 | 5200 | 77.6 | 51 |
| Compound | PvXyn11A | PvXyd3A | ||
|---|---|---|---|---|
| 1 mM | 10 mM | 1 mM | 10 mM | |
| Control | 100 | 100 | 100 | 100 |
| CuCl2 | 60 ± 5 | 39 ± 4 | 95 ± 3 | 92 ± 3 |
| CoCl2 | 90 ± 6 | 78 ± 5 | 105 ± 7 | 97 ± 6 |
| NaCl | 102 ± 4 | 109 ± 8 | 95 ± 6 | 95 ± 9 |
| BaCl2 | 98 ± 5 | 103 ± 2 | 88 ± 3 | 86 ± 5 |
| CaCl2 | 107 ± 4 | 110 ± 7 | 95 ± 7 | 92 ± 5 |
| HgCl2 | 86 ± 6 | 7 ± 3 | 95 ± 3 | 66 ± 7 |
| MgSO4 | 90 ± 7 | 93 ± 8 | 103 ± 6 | 99 ± 3 |
| Pb(CH3COO)2 | 106 ± 7 | 95 ± 5 | 106 ± 3 | 99 ± 4 |
| MnSO4 | 94 ± 5 | 88 ± 6 | 104 ± 5 | 138 ± 5 |
| ZnSO4 | 88 ± 10 | 85 ± 7 | 106 ± 9 | 122 ± 6 |
| SDS | 84 ± 5 | 80 ± 2 | 81 ± 3 | 72 ± 5 |
| EDTA | 94 ± 5 | 90 ± 4 | 99 ± 2 | 95 ± 3 |
| DDT | 98 ± 6 | 103 ± 7 | 81 ± 4 | 50 ± 7 |
| P. thermophila J18 | P. thermophila J18 | P. thermophila J18 | Thermoascus crustaceus JCM12803 | Thermoascus auranticus | P. variotii | P. variotii IMD RK 032 | B. spectabilis ATHUM8891 | |
|---|---|---|---|---|---|---|---|---|
| Production Source | WT | E. coli | P. Pastoris | P. Pastoris | WT | WT | WT | WT |
| MW | 26 kDa | 28 kDa | 29 kDa | ND | 32 | 25 kDa | 20 kDa | 23 kDa |
| pI | ND | 4.43 | ND | ND | 7.1 | 3.9 | 5.2 | >3 & <4 |
| C(H2O) content | 21% | ND | ND | ND | ND | 4.5% | No glycosylation | 7.1% |
| Specific activity | 20,500 nkat/mg (BiX) | 11,800 nkat/mg (BiX) | 108,950 nkat/mg (BiX) | 24,716 nkat/mg (BeX) | 1064 nkat/mg (OAX) | 8200 (LX) | 816 nkat/mg (xylan) | 75,800 nkat/mg (BeX) |
| T optimum | 75 °C | 75 °C | 75 °C | 65–70 °C | 80 °C | 65 °C | 50 °C | 60 °C |
| T stability | Stable up to 75 °C for 30 min | t1/2 174.8, 137.9, 107.4 and 68.2 min at 70, 75, 80 and 85 °C, resp. | Stable up to 70 °C for 30 min. 75% residual act. at 75 °C and 65% at 80 °C (30 min) | Stable for 60 min at least at 60 °C | 60 min at 80 °C. Stable up to 70 °C for days | Stable up to 60 °C. 65% residual activity after 60 min at 70 °C, inactivation after 40 min at 80 °C | ND | t1/2 of 17 and 60 min at 65 and 60 °C. Stable up to 55 °C for 2 days at least |
| pH optimum | 6.5 | 7 | 7 | 5 | 5 | 5.5–7 | 4 | 3.5 |
| pH stability | Stable for 30 min at least at pH 6.0–11.0 | Stable for 30 min at least at pH 6.5–10.5 | Stable for 30 min at least at pH 4.5–11 | Stable at pH 3–11 | ND | Stable at pH 3–10 | ND | Stable for 2 days at least at pH 2.5–8 |
| KM | ND | 4.4 (BiX), 3.6 mg/mL (BeX), 9.7 mg/mL (OAX) | ND | ND | 1.7 mg/mL (OAX) | 2.5 mg/mL (LX) | 49.5 mg/mL (xylan) | 2.52 ± 0.45 mg/mL (BeX) |
| GH family | ND | 11 | ND | ND | ND | ND | ND | 11 |
| Ref. | [26] | [27] | [28] | [29] | [30] | [19,31] | [18] | Present work |
| P. thermophila J18 | P. thermophila J18 | P. thermophila J18 | Thermosascus sp. | P. variotii IMD RK 032 | B. spectabilis ATHUM 8891 | |
|---|---|---|---|---|---|---|
| Production source | WT | E. coli | P. pastoris | WT | WT | WT |
| MW | 53.5 kDa | 52. kDa | 52.3 kDa | 100 kDa | 67 kDa | 78 kDa |
| pI | ND | ND | ND | ND | 4 | >3 & <4 |
| Specific activity | 724 nkat/mg (pNPX) | 765 nkat/mg (pNPX) | 4.4 nkat/mg (pNPX) | 116.7 nkat/mg (pNPX) | ND | 5200 nkat/mg (pNPX) |
| T optimum | 55 °C | 55 °C | 60 °C | 55 °C | 60 °C | 70 °C |
| T stability | Stable up to 55 °C for 30 min | t1/2 of 1160, 605 and 15 min at 50, 55 and 60 °C | ND | Stable up to 60 °C for 1 h. | ND | t1/2 of 25 min at 70. Stable up to 60 °C for 2 days at least |
| pH optimum | 6.5 | 7 | 7 | 4.5 | 4 | 3.5 |
| pH stability | Stable 30 min from 6 to 9 at 50 °C | Stable 30 min from 6 to 9 at 50 °C | ND | ND | ND | Stable for 2 days at least at pH 2.5–8 |
| KM (pnpX) | 4.3 mM | 4.5 mM | 8 mM | ND | 5.4 mM | 0.45 ± 0.06 mM |
| KI (xylose) | 139 mM | ND | ND | ND | ND | 10.2 mM |
| GH | ND | GH43 | ND | ND | ND | 3 |
| Ref. | [32] | [33] | [34] | [35] | [18] | Present work |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galanopoulou, A.P.; Haimala, I.; Georgiadou, D.N.; Mamma, D.; Hatzinikolaou, D.G. Characterization of the Highly Efficient Acid-Stable Xylanase and β-Xylosidase System from the Fungus Byssochlamys spectabilis ATHUM 8891 (Paecilomyces variotii ATHUM 8891). J. Fungi 2021, 7, 430. https://doi.org/10.3390/jof7060430
Galanopoulou AP, Haimala I, Georgiadou DN, Mamma D, Hatzinikolaou DG. Characterization of the Highly Efficient Acid-Stable Xylanase and β-Xylosidase System from the Fungus Byssochlamys spectabilis ATHUM 8891 (Paecilomyces variotii ATHUM 8891). Journal of Fungi. 2021; 7(6):430. https://doi.org/10.3390/jof7060430
Chicago/Turabian StyleGalanopoulou, Anastasia P., Irini Haimala, Daphne N. Georgiadou, Diomi Mamma, and Dimitris G. Hatzinikolaou. 2021. "Characterization of the Highly Efficient Acid-Stable Xylanase and β-Xylosidase System from the Fungus Byssochlamys spectabilis ATHUM 8891 (Paecilomyces variotii ATHUM 8891)" Journal of Fungi 7, no. 6: 430. https://doi.org/10.3390/jof7060430