
Probiotic Properties and Potentiality of Lactiplantibacillus plantarum Strains for the Biological Control of Chalkbrood Disease

 ,
,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Cultures
2.2. Screening for Antifungal Activity
2.3. Antifungal Activity Determination
2.3.1. Spore Viability and Germination Test
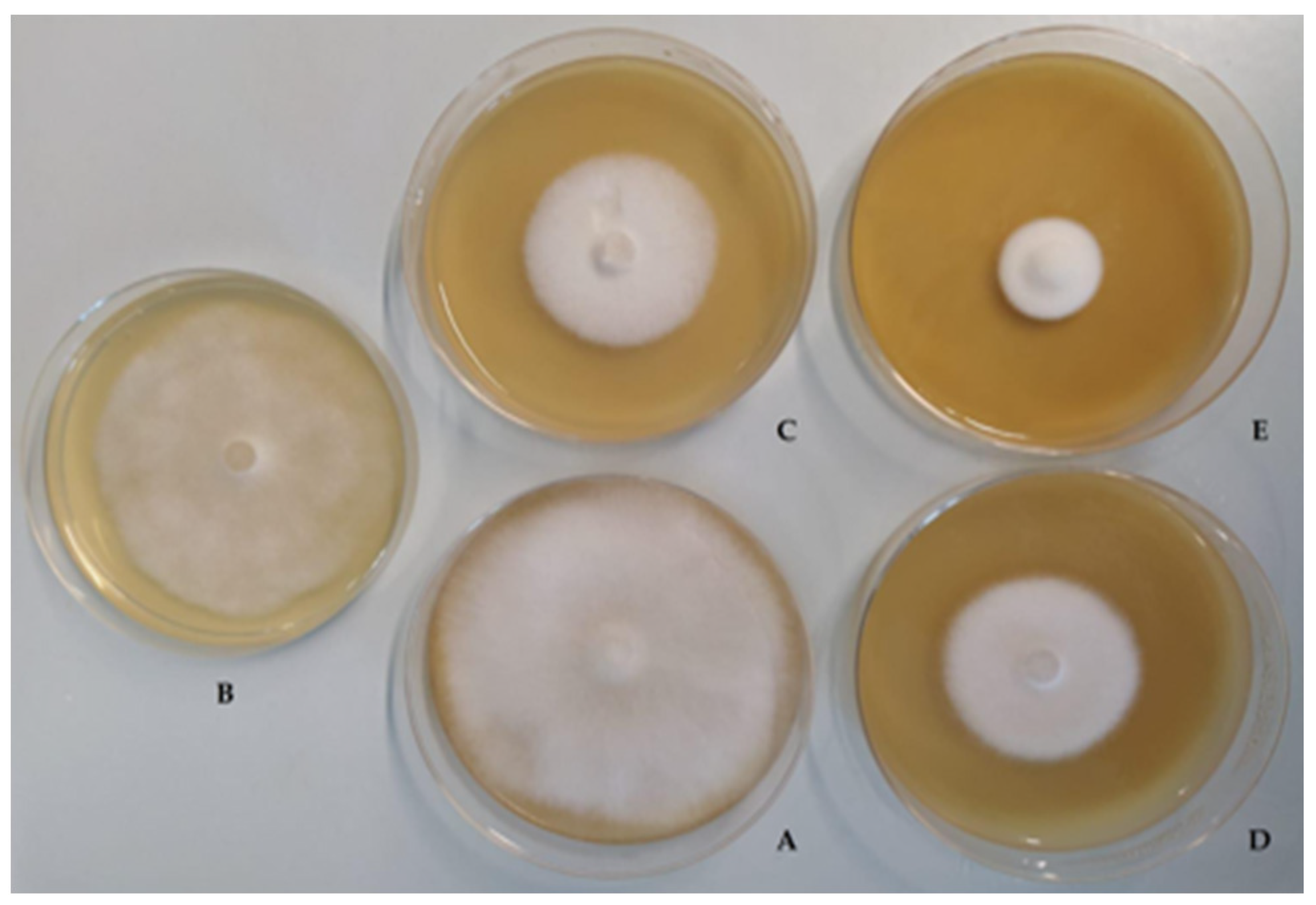
2.3.2. Inhibition of Radial Mycelial Growth
2.4. Biofilm Production
2.5. Exopolysaccharide (EPS) Assay
2.5.1. Production and Isolation of EPSs
2.5.2. Antifungal Activity of EPSs
2.6. Antioxidant Activity
2.6.1. Bacterial Culture Matrices and Cell Protein Assay
2.6.2. Antioxidant Activity Assay
2.7. Statistical Analysis
3. Results
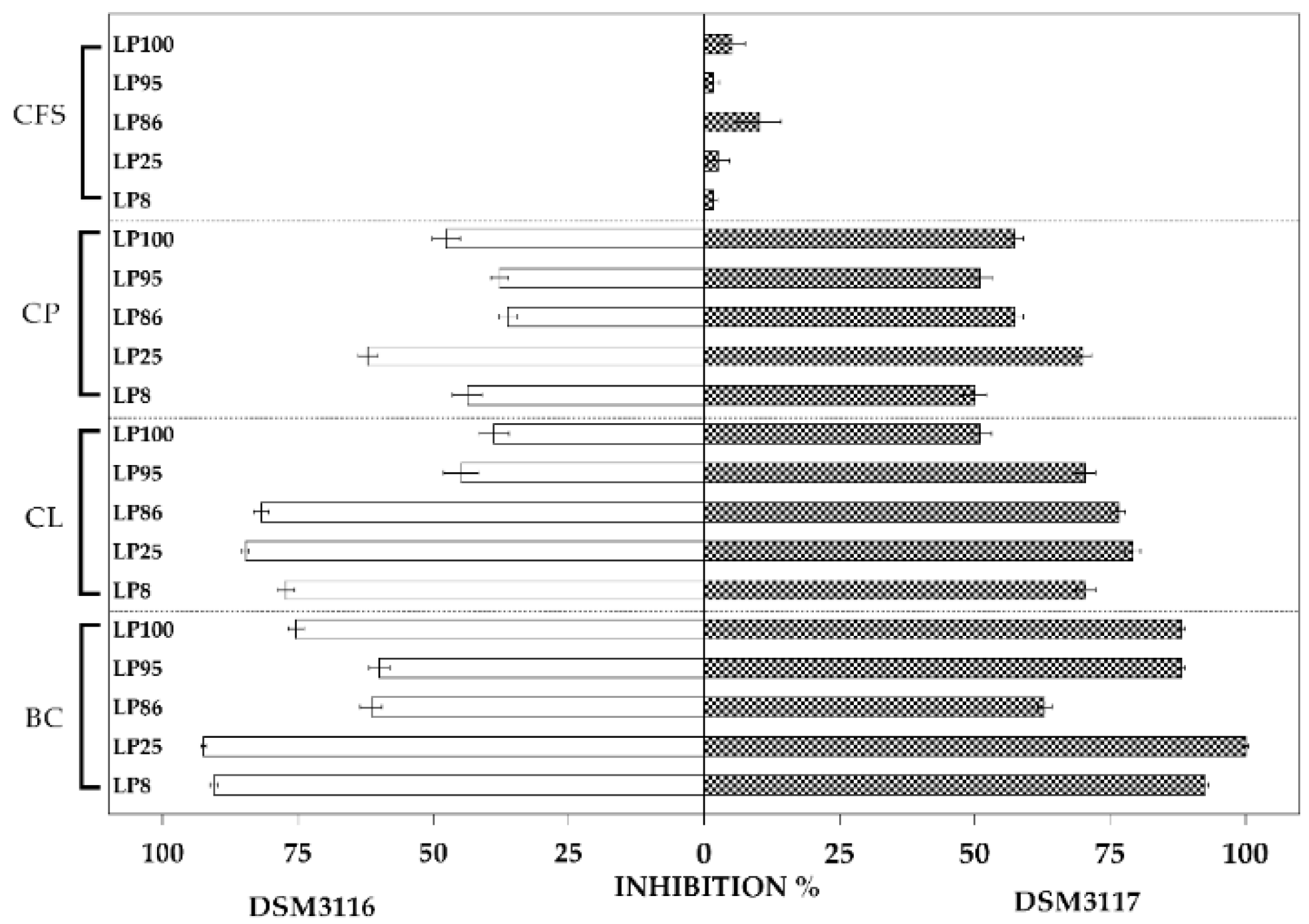
3.1. Antifungal Activity
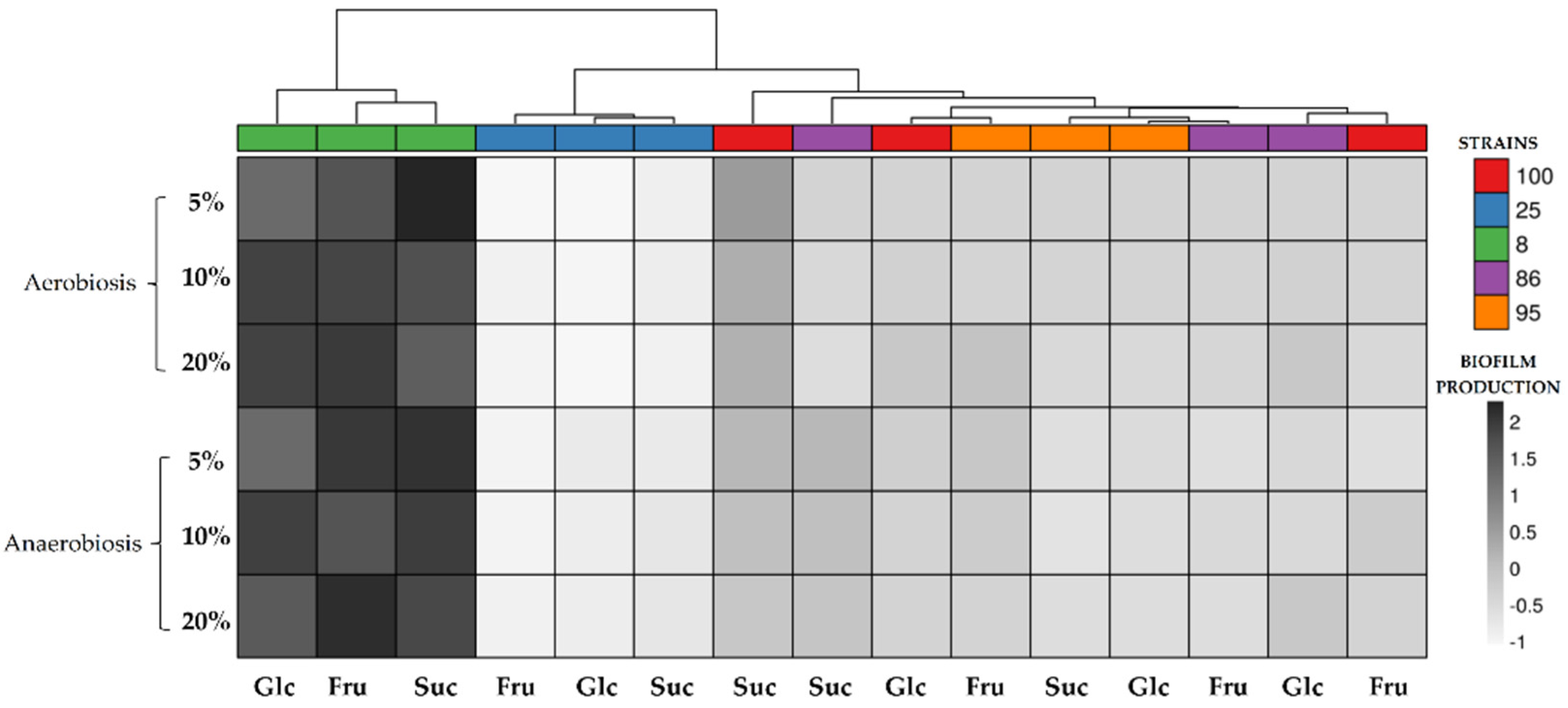
3.2. EPS and Biofilm Production
3.3. Antioxidant Activity
4. Discussion
4.1. Antifungal Activity
4.2. EPS and Biofilm Production
4.3. Antioxidant Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Evison, S.E.; Jensen, A.B. The Biology and Prevalence of Fungal Diseases in Managed and Wild Bees. Ecol. Parasites Parasit. Biol. Control 2018, 26, 105–113. [Google Scholar] [CrossRef]
- Evison, S.E. Chalkbrood: Epidemiological Perspectives from the Host–Parasite Relationship. Soc. Insects Vectors Med. Vet. Entomol. 2015, 10, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Aronstein, K.A.; Murray, K.D. Chalkbrood Disease in Honey Bees. J. Invertebr. Pathol. 2010, 103, S20–S29. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hou, M.; Qiu, Y.; Zhao, B.; Nie, H.; Su, S. Changes in Antioxidant Enzymes Activity and Metabolomic Profiles in the Guts of Honey Bee (Apis Mellifera) Larvae Infected with Ascosphaera Apis. Insects 2020, 11, 419. [Google Scholar] [CrossRef]
- Bamford, S.; Heath, L.A.F. The Effects of Temperature and PH on the Germination of Spores of the Chalkbrood Fungus, Ascosphaera Apis. J. Apic. Res. 1989, 28, 36–40. [Google Scholar] [CrossRef]
- Bailey, L.; Ball, B.V. (Eds.) 5-FUNGI. In Honey Bee Pathology, 2nd ed.; Academic Press: London, UK, 1991; pp. 53–63. ISBN 978-0-12-073481-8. [Google Scholar]
- Spivak, M.; Reuter, G.S. Performance of Hygienic Honey Bee Colonies in a Commercial Apiary. Apidologie 1998, 29, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Invernizzi, C.; Rivas, F.; Bettucci, L. Resistance to Chalkbrood Disease in Apis Mellifera L. (Hymenoptera: Apidae) Colonies with Different Hygienic Behaviour. Neotrop. Entomol. 2011, 40, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Manley, R.; Boots, M.; Wilfert, L. REVIEW: Emerging Viral Disease Risk to Pollinating Insects: Ecological, Evolutionary and Anthropogenic Factors. J. Appl. Ecol. 2015, 52, 331–340. [Google Scholar] [CrossRef]
- Castagnino, G.L.B.; Mateos, A.; Meana, A.; Montejo, L.; Zamorano Iturralde, L.V.; Cutuli de Simón, M.T. Etiology, Symptoms and Prevention of Chalkbrood Disease: A Literature Review. Rev. Bras. Saúde E Produção Anim. 2020, 21. [Google Scholar] [CrossRef]
- Engel, P.; Moran, N.A. Functional and Evolutionary Insights into the Simple yet Specific Gut Microbiota of the Honey Bee from Metagenomic Analysis. Gut Microbes 2013, 4, 60–65. [Google Scholar] [CrossRef]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.-L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A Metagenomic Survey of Microbes in Honey Bee Colony Collapse Disorder. Science 2007, 318, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamdi, C.; Balloi, A.; Essanaa, J.; Crotti, E.; Gonella, E.; Raddadi, N.; Ricci, I.; Boudabous, A.; Borin, S.; Manino, A.; et al. Gut Microbiome Dysbiosis and Honeybee Health. J. Appl. Entomol. 2011, 135, 524–533. [Google Scholar] [CrossRef] [Green Version]
- Alberoni, D.; Gaggìa, F.; Baffoni, L.; Di Gioia, D. Beneficial Microorganisms for Honey Bees: Problems and Progresses. Appl. Microbiol. Biotechnol. 2016, 100, 9469–9482. [Google Scholar] [CrossRef]
- Raymann, K.; Moran, N.A. The Role of the Gut Microbiome in Health and Disease of Adult Honey Bee Workers. Ecol. Parasites Parasit. Biol. Control 2018, 26, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Sabaté, D.C.; Carrillo, L.; Carina Audisio, M. Inhibition of Paenibacillus Larvae and Ascosphaera Apis by Bacillus Subtilis Isolated from Honeybee Gut and Honey Samples. Res. Microbiol. 2009, 160, 193–199. [Google Scholar] [CrossRef]
- Li, J.; Zheng, Z.; Hong, S.; Qi, X.; Liang, Q. Isolation and Identification of an Antagonistic Bacterial Strain against Ascosphaera Apis from Honeybee Larvae Infected with Chalkbrood Disease. Sci. Agric. Sin. 2012, 45, 973–980. [Google Scholar]
- Omar, M.O.M.; Moustafa, A.M.; Ansari, M.J.; Anwar, A.M.; Fahmy, B.F.; Al-Ghamdi, A.; Nuru, A. Antagonistic Effect of Gut Bacteria in the Hybrid Carniolan Honey Bee, Apis Mellifera Carnica, Against Ascosphaera Apis, the Causal Organism of Chalkbrood Disease. J. Apic. Sci. 2014, 58, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Heath, L.A.F. Development of Chalk Brood in a Honeybee Colony: A Review. Bee World 1982, 63, 119–130. [Google Scholar] [CrossRef]
- Liu, T.P. Ultrastructural Changes in the Spore and Mycelia of Ascosphaera Apis after Treatment with Benomyl (Benlate 50 W). Mycopathologia 1991, 116, 23–28. [Google Scholar] [CrossRef]
- Glinski, Z.; Chmielewski, M. Imidazole Derivatives in Control of the Honey Bee Brood Mycoses. Pszczel. Zesz. Nauk. Pol. 1996, 40, 165–173. [Google Scholar]
- Davis, C. Control of Chalkbrood Disease with Natural Products: A Report for the Rural Industries Research and Development Corporation/by Craig Davis and Wendy Ward; RIRDC Publication; No. 03/107; Rural Industries Research and Development Corporation: Barton, ACT, Austrilia, 2003. [Google Scholar]
- Frazier, M.; Mullin, C.; Frazier, J.; Ashcraft, S. What Have Pesticides Got to Do with It? Am. Bee J. 2008, 148, 521–524. [Google Scholar]
- Gaggìa, F.; Baffoni, L.; Alberoni, D. Probiotics for honeybees’ health. In Probiotics and Prebiotics in Animal Health and Food Safety; Springer: Berlin/Heidelberg, Germany, 2018; pp. 219–245. [Google Scholar]
- Al-Ghamdi, A.; Ali Khan, K.; Javed Ansari, M.; Almasaudi, S.B.; Al-Kahtani, S. Effect of Gut Bacterial Isolates from Apis Mellifera Jemenitica on Paenibacillus Larvae Infected Bee Larvae. Saudi J. Biol. Sci. 2018, 25, 383–387. [Google Scholar] [CrossRef]
- Tejerina, M.R.; Benítez-Ahrendts, M.R.; Audisio, M.C. Lactobacillus Salivarius A3iob Reduces the Incidence of Varroa Destructor and Nosema Spp. in Commercial Apiaries Located in the Northwest of Argentina. Probiotics Antimicrob. Proteins 2020, 12, 1360–1369. [Google Scholar] [CrossRef]
- Daisley, B.A.; Pitek, A.P.; Chmiel, J.A.; Al, K.F.; Chernyshova, A.M.; Faragalla, K.M.; Burton, J.P.; Thompson, G.J.; Reid, G. Novel Probiotic Approach to Counter Paenibacillus Larvae Infection in Honey Bees. ISME J. 2020, 14, 476–491. [Google Scholar] [CrossRef] [Green Version]
- Ramos, O.Y.; Basualdo, M.; Libonatti, C.; Vega, M.F. Current Status and Application of Lactic Acid Bacteria in Animal Production Systems with a Focus on Bacteria from Honey Bee Colonies. J. Appl. Microbiol. 2020, 128, 1248–1260. [Google Scholar] [CrossRef]
- Tejerina, M.R.; Cabana, M.J.; Benitez-Ahrendts, M.R. Strains of Lactobacillus Spp. Reduce Chalkbrood in Apis Mellifera. J. Invertebr. Pathol. 2021, 178, 107521. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.M.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J. A Taxonomic Note on the Genus Lactobacillus: Description of 23 Novel Genera, Emended Description of the Genus Lactobacillus Beijerinck 1901, and Union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Van Hoorde, K.; Verstraete, T.; Vandamme, P.; Huys, G. Diversity of Lactic Acid Bacteria in Two Flemish Artisan Raw Milk Gouda-Type Cheeses. Food Microbiol. 2008, 25, 929–935. [Google Scholar] [CrossRef]
- Wouters, D.; Grosu-Tudor, S.; Zamfir, M.; De Vuyst, L. Bacterial Community Dynamics, Lactic Acid Bacteria Species Diversity and Metabolite Kinetics of Traditional Romanian Vegetable Fermentations. J. Sci. Food Agric. 2013, 93, 749–760. [Google Scholar] [CrossRef]
- Gotteland, M.; Cires, M.J.; Carvallo, C.; Vega, N.; Ramirez, M.A.; Morales, P.; Rivas, P.; Astudillo, F.; Navarrete, P.; Dubos, C.; et al. Probiotic Screening and Safety Evaluation of Lactobacillus Strains from Plants, Artisanal Goat Cheese, Human Stools, and Breast Milk. J. Med. Food 2014, 17, 487–495. [Google Scholar] [CrossRef]
- Tajabadi, N.; Mardan, M.; Saari, N.; Mustafa, S.; Bahreini, R.; Manap, M.Y.A. Identification of Lactobacillus Plantarum, Lactobacillus Pentosus and Lactobacillus Fermentum from Honey Stomach of Honeybee. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2014, 44, 717–722. [Google Scholar] [CrossRef] [Green Version]
- Javorský, P.; Fecskeová, L.K.; Hrehová, L.; Sabo, R.; Legáth, J.; Pristas, P. Establishment of Lactobacillus Plantarum Strain in Honey Bee Digestive Tract Monitored Using Gfp Fluorescence. Benef. Microbes 2017, 8, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Parichehreh, S.; Tahmasbi, G.; Sarafrazi, A.; Imani, S.; Tajabadi, N. Isolation and Identification of Lactobacillus Bacteria Found in the Gastrointestinal Tract of the Dwarf Honey Bee, Apis Florea Fabricius, 1973 (Hymenoptera: Apidae). Apidologie 2018, 49, 430–438. [Google Scholar] [CrossRef] [Green Version]
- Iorizzo, M.; Pannella, G.; Lombardi, S.J.; Ganassi, S.; Testa, B.; Succi, M.; Sorrentino, E.; Petrarca, S.; De Cristofaro, A.; Coppola, R.; et al. Inter- and Intra-Species Diversity of Lactic Acid Bacteria in Apis Mellifera Ligustica Colonies. Microorganisms 2020, 8, 1578. [Google Scholar] [CrossRef] [PubMed]
- Corby-Harris, V.; Pontaroli, A.C.; Shimkets, L.J.; Bennetzen, J.L.; Habel, K.E.; Promislow, D.E.L. Geographical Distribution and Diversity of Bacteria Associated with Natural Populations of Drosophila Melanogaster. Appl. Environ. Microbiol. 2007, 73, 3470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, C.; Webster, P.; Finkel, S.E.; Tower, J. Increased Internal and External Bacterial Load during Drosophila Aging without Life-Span Trade-Off. Cell Metab. 2007, 6, 144–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, J.-H.; Kim, S.-H.; Lee, H.-Y.; Bai, J.Y.; Nam, Y.-D.; Bae, J.-W.; Lee, D.G.; Shin, S.C.; Ha, E.-M.; Lee, W.-J. Innate Immune Homeostasis by the Homeobox Gene Caudal and Commensal-Gut Mutualism in Drosophila. Science 2008, 319, 777. [Google Scholar] [CrossRef] [Green Version]
- Sharon, G.; Segal, D.; Ringo, J.M.; Hefetz, A.; Zilber-Rosenberg, I.; Rosenberg, E. Commensal Bacteria Play a Role in Mating Preference of Drosophila Melanogaster. Proc. Natl. Acad. Sci. USA 2010, 107, 20051. [Google Scholar] [CrossRef] [Green Version]
- Storelli, G.; Defaye, A.; Erkosar, B.; Hols, P.; Royet, J.; Leulier, F. Lactobacillus Plantarum Promotes Drosophila Systemic Growth by Modulating Hormonal Signals through TOR-Dependent Nutrient Sensing. Cell Metab. 2011, 14, 403–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niku-Paavola, M.-L.; Laitila, A.; Mattila-Sandholm, T.; Haikara, A. New Types of Antimicrobial Compounds Produced by Lactobacillus Plantarum. J. Appl. Microbiol. 1999, 86, 29–35. [Google Scholar] [CrossRef]
- Lavermicocca, P.; Valerio, F.; Evidente, A.; Lazzaroni, S.; Corsetti, A.; Gobbetti, M. Purification and Characterization of Novel Antifungal Compounds from the Sourdough Lactobacillus Plantarum Strain 21B. Appl. Environ. Microbiol. 2000, 66, 4084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjögren, J.; Magnusson, J.; Broberg, A.; Schnürer, J.; Kenne, L. Antifungal 3-Hydroxy Fatty Acids from Lactobacillus Plantarum MiLAB 14. Appl. Environ. Microbiol. 2003, 69, 7554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, M.S.; Todorov, S.D.; Ivanova, I.V.; Belguesmia, Y.; Choiset, Y.; Rabesona, H.; Chobert, J.-M.; Haertlé, T.; Franco, B.D.G.M. Characterization of a Two-Peptide Plantaricin Produced by Lactobacillus Plantarum MBSa4 Isolated from Brazilian Salami. Food Control 2016, 60, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Iorizzo, M.; Testa, B.; Lombardi, S.J.; Ganassi, S.; Ianiro, M.; Letizia, F.; Succi, M.; Tremonte, P.; Vergalito, F.; Cozzolino, A.; et al. Antimicrobial Activity against Paenibacillus Larvae and Functional Properties of Lactiplantibacillus Plantarum Strains: Potential Benefits for Honeybee Health. Antibiotics 2020, 9, 442. [Google Scholar] [CrossRef] [PubMed]
- Behera, S.S.; Ray, R.C.; Zdolec, N. Lactobacillus Plantarum with Functional Properties: An Approach to Increase Safety and Shelf-Life of Fermented Foods. BioMed Res. Int. 2018, 2018, 9361614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seddik, H.A.; Bendali, F.; Gancel, F.; Fliss, I.; Spano, G.; Drider, D. Lactobacillus Plantarum and Its Probiotic and Food Potentialities. Probiotics Antimicrob. Proteins 2017, 9, 111–122. [Google Scholar] [CrossRef]
- Royan, M. Mechanisms of Probiotic Action in the Honeybee. Crit. Rev. Eukaryot. Gene Expr. 2019, 29, 95–103. [Google Scholar] [CrossRef]
- Deng, Z.; Luo, X.M.; Liu, J.; Wang, H. Quorum Sensing, Biofilm, and Intestinal Mucosal Barrier: Involvement the Role of Probiotic. Front. Cell. Infect. Microbiol. 2020, 10, 504. [Google Scholar] [CrossRef]
- Bäumler, A.J.; Sperandio, V. Interactions between the Microbiota and Pathogenic Bacteria in the Gut. Nature 2016, 535, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Barzegari, A.; Kheyrolahzadeh, K.; Hosseiniyan Khatibi, S.M.; Sharifi, S.; Memar, M.Y.; Zununi Vahed, S. The Battle of Probiotics and Their Derivatives Against Biofilms. Infect. Drug Resist. 2020, 13, 659–672. [Google Scholar] [CrossRef] [Green Version]
- Vu, B.; Chen, M.; Crawford, R.J.; Ivanova, E.P. Bacterial Extracellular Polysaccharides Involved in Biofilm Formation. Molecules 2009, 14, 535. [Google Scholar] [CrossRef]
- Magnusson, J.; Ström, K.; Roos, S.; Sjögren, J.; Schnürer, J. Broad and Complex Antifungal Activity among Environmental Isolates of Lactic Acid Bacteria. FEMS Microbiol. Lett. 2003, 219, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Testa, B.; Lombardi, S.J.; Macciola, E.; Succi, M.; Tremonte, P.; Iorizzo, M. Efficacy of Olive Leaf Extract (Olea Europaea L. Cv Gentile Di Larino) in Marinated Anchovies (Engraulis Encrasicolus, L.) Process. Heliyon 2019, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, A.B.; Aronstein, K.; Flores, J.M.; Vojvodic, S.; Palacio, M.A.; Spivak, M. Standard Methods for Fungal Brood Disease Research. J. Apic. Res. 2013, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heath, L.A.F.; Gaze, B.M. Carbon Dioxide Activation of Spores of the Chalkbrood Fungus Ascosphaera Apis. J. Apic. Res. 1987, 26, 243–246. [Google Scholar] [CrossRef]
- Iorizzo, M.; Lombardi, S.J.; Ganassi, S.; Testa, B.; Ianiro, M.; Letizia, F.; Succi, M.; Tremonte, P.; Vergalito, F.; Cozzolino, A.; et al. Antagonistic Activity against Ascosphaera Apis and Functional Properties of Lactobacillus Kunkeei Strains. Antibiotics 2020, 9, 262. [Google Scholar] [CrossRef] [PubMed]
- Ricci, A.; Levante, A.; Cirlini, M.; Calani, L.; Bernini, V.; Del Rio, D.; Galaverna, G.; Neviani, E.; Lazzi, C. The Influence of Viable Cells and Cell-Free Extracts of Lactobacillus Casei on Volatile Compounds and Polyphenolic Profile of Elderberry Juice. Front. Microbiol. 2018, 9, 2784. [Google Scholar] [CrossRef]
- Cozzolino, A.; Vergalito, F.; Tremonte, P.; Iorizzo, M.; Lombardi, S.J.; Sorrentino, E.; Luongo, D.; Coppola, R.; Di Marco, R.; Succi, M. Preliminary Evaluation of the Safety and Probiotic Potential of Akkermansia Muciniphila DSM 22959 in Comparison with Lactobacillus Rhamnosus GG. Microorganisms 2020, 8, 189. [Google Scholar] [CrossRef] [Green Version]
- Tallon, R.; Bressollier, P.; Urdaci, M.C. Isolation and Characterization of Two Exopolysaccharides Produced by Lactobacillus Plantarum EP56. Res. Microbiol. 2003, 154, 705–712. [Google Scholar] [CrossRef]
- Di Martino, C.; Testa, B.; Letizia, F.; Iorizzo, M.; Lombardi, S.J.; Ianiro, M.; Di Renzo, M.; Strollo, D.; Coppola, R. Effect of Exogenous Proline on the Ethanolic Tolerance and Malolactic Performance of Oenococcus Oeni. J. Food Sci. Technol. 2020, 57, 3973–3979. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A Web Tool for Visualizing Clustering of Multivariate Data Using Principal Component Analysis and Heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Albo, G.N.; Cordoba, S.B.; Reynaldi, F.J. Chalkbrood: Pathogenesis and the Interaction with Honeybee Defenses. Int. J. Environ. Agric. Res. 2017, 3, 71–80. [Google Scholar]
- Maxfield-Taylor, S.A.; Mujic, A.B.; Rao, S. First Detection of the Larval Chalkbrood Disease Pathogen Ascosphaera Apis (Ascomycota: Eurotiomycetes: Ascosphaerales) in Adult Bumble Bees. PLoS ONE 2015, 10, e0124868. [Google Scholar] [CrossRef] [PubMed]
- Landini, P.; Antoniani, D.; Burgess, J.G.; Nijland, R. Molecular Mechanisms of Compounds Affecting Bacterial Biofilm Formation and Dispersal. Appl. Microbiol. Biotechnol. 2010, 86, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Crowley, S.; Mahony, J.; van Sinderen, D. Current Perspectives on Antifungal Lactic Acid Bacteria as Natural Bio-Preservatives. Trends Food Sci. Technol. 2013, 33, 93–109. [Google Scholar] [CrossRef]
- Bulgasem, B.Y.; Lani, M.N.; Hassan, Z.; Yusoff, W.M.W.; Fnaish, S.G. Antifungal Activity of Lactic Acid Bacteria Strains Isolated from Natural Honey against Pathogenic Candida Species. Mycobiology 2016, 44, 302–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marie, K.P.; Ngoufack François, Z.; Edith Marius, F.K.; Ciobotaru, O.; Matei, F.; Cornea, C.P.; Israel-Roming, F. Antifungal Activity of Lactic Acid Bacteria Isolated from Peanuts, Gari, and Orange Fruit Juice against Food Aflatoxigenic Molds. Food Biotechnol. 2018, 32, 237–256. [Google Scholar] [CrossRef]
- Coloretti, F.; Carri, S.; Armaforte, E.; Chiavari, C.; Grazia, L.; Zambonelli, C. Antifungal Activity of Lactobacilli Isolated from Salami. FEMS Microbiol. Lett. 2007, 271, 245–250. [Google Scholar] [CrossRef] [Green Version]
- Voulgari, K.; Hatzikamari, M.; Delepoglou, A.; Georgakopoulos, P.; Litopoulou-Tzanetaki, E.; Tzanetakis, N. Antifungal Activity of Non-Starter Lactic Acid Bacteria Isolates from Dairy Products. Food Control 2010, 21, 136–142. [Google Scholar] [CrossRef]
- Arena, M.P.; Silvain, A.; Normanno, G.; Grieco, F.; Drider, D.; Spano, G.; Fiocco, D. Use of Lactobacillus Plantarum Strains as a Bio-Control Strategy against Food-Borne Pathogenic Microorganisms. Front. Microbiol. 2016, 7, 464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremonte, P.; Pannella, G.; Succi, M.; Luca, T.; Sturchio, M.; Coppola, R.; Luongo, D.; Sorrentino, E. Antimicrobial Activity of Lactobacillus Plantarum Strains Isolated from Different Environments: A Preliminary Study. Int. Food Res. J. 2017, 24, 852–859. [Google Scholar]
- Sorrentino, E.; Tremonte, P.; Succi, M.; Iorizzo, M.; Pannella, G.; Lombardi, S.J.; Sturchio, M.; Coppola, R. Detection of Antilisterial Activity of 3-Phenyllactic Acid Using Listeria Innocua as a Model. Front. Microbiol. 2018, 9, 1373. [Google Scholar] [CrossRef] [PubMed]
- Le Lay, C.; Coton, E.; Le Blay, G.; Chobert, J.-M.; Haertlé, T.; Choiset, Y.; Van Long, N.N.; Meslet-Cladière, L.; Mounier, J. Identification and Quantification of Antifungal Compounds Produced by Lactic Acid Bacteria and Propionibacteria. Int. J. Food Microbiol. 2016, 239, 79–85. [Google Scholar] [CrossRef]
- Cortés-Zavaleta, O.; López-Malo, A.; Hernández-Mendoza, A.; García, H.S. Antifungal Activity of Lactobacilli and Its Relationship with 3-Phenyllactic Acid Production. Int. J. Food Microbiol. 2014, 173, 30–35. [Google Scholar] [CrossRef]
- Kleerebezem, M.; Hols, P.; Bernard, E.; Rolain, T.; Zhou, M.; Siezen, R.J.; Bron, P.A. The Extracellular Biology of the Lactobacilli. FEMS Microbiol. Rev. 2010, 34, 199–230. [Google Scholar] [CrossRef]
- Kinoshita, H.; Uchida, H.; Kawai, Y.; Kawasaki, T.; Wakahara, N.; Matsuo, H.; Watanabe, M.; Kitazawa, H.; Ohnuma, S.; Miura, K.; et al. Cell Surface Lactobacillus Plantarum LA 318 Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) Adheres to Human Colonic Mucin. J. Appl. Microbiol. 2008, 104, 1667–1674. [Google Scholar] [CrossRef] [PubMed]
- Åvall-Jääskeläinen, S.; Palva, A. Lactobacillus Surface Layers and Their Applications. FEMS Microbiol. Rev. 2005, 29, 511–529. [Google Scholar] [CrossRef] [Green Version]
- Janashia, I.; Choiset, Y.; Jozefiak, D.; Déniel, F.; Coton, E.; Moosavi-Movahedi, A.A.; Chanishvili, N.; Haertlé, T. Beneficial Protective Role of Endogenous Lactic Acid Bacteria Against Mycotic Contamination of Honeybee Beebread. Probiotics Antimicrob. Proteins 2018, 10, 638–646. [Google Scholar] [CrossRef]
- Siedler, S.; Balti, R.; Neves, A.R. Bioprotective Mechanisms of Lactic Acid Bacteria against Fungal Spoilage of Food. Food Biotechnol. Plant Biotechnol. 2019, 56, 138–146. [Google Scholar] [CrossRef]
- Vásquez, A.; Forsgren, E.; Fries, I.; Paxton, R.J.; Flaberg, E.; Szekely, L.; Olofsson, T.C. Symbionts as Major Modulators of Insect Health: Lactic Acid Bacteria and Honeybees. PLoS ONE 2012, 7, e33188. [Google Scholar] [CrossRef]
- Alberoni, D.; Baffoni, L.; Gaggìa, F.; Ryan, P.M.; Murphy, K.; Ross, P.R.; Stanton, C.; Di Gioia, D. Impact of Beneficial Bacteria Supplementation on the Gut Microbiota, Colony Development and Productivity of Apis Mellifera L. Benef. Microbes 2018, 9, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, R.S.; Moran, N.A.; Evans, J.D. Early Gut Colonizers Shape Parasite Susceptibility and Microbiota Composition in Honey Bee Workers. Proc. Natl. Acad. Sci. USA 2016, 113, 9345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santarmaki, V.; Kourkoutas, Y.; Zoumpopoulou, G.; Mavrogonatou, E.; Kiourtzidis, M.; Chorianopoulos, N.; Tassou, C.; Tsakalidou, E.; Simopoulos, C.; Ypsilantis, P. Survival, Intestinal Mucosa Adhesion, and Immunomodulatory Potential of Lactobacillus Plantarum Strains. Curr. Microbiol. 2017, 74, 1061–1067. [Google Scholar] [CrossRef] [PubMed]
- Tamarit, D.; Ellegaard, K.M.; Wikander, J.; Olofsson, T.; Vásquez, A.; Andersson, S.G.E. Functionally Structured Genomes in Lactobacillus Kunkeei Colonizing the Honey Crop and Food Products of Honeybees and Stingless Bees. Genome Biol. Evol. 2015, 7, 1455–1473. [Google Scholar] [CrossRef] [PubMed]
- Salas-Jara, M.J.; Ilabaca, A.; Vega, M.; García, A. Biofilm Forming Lactobacillus: New Challenges for the Development of Probiotics. Microorganisms 2016, 4, 35. [Google Scholar] [CrossRef]
- Berríos, P.; Fuentes, J.A.; Salas, D.; Carreño, A.; Aldea, P.; Fernández, F.; Trombert, A.N. Inhibitory Effect of Biofilm-Forming Lactobacillus Kunkeei Strains against Virulent Pseudomonas Aeruginosa in Vitro and in Honeycomb Moth (Galleria Mellonella) Infection Model. Benef. Microbes 2018, 9, 257–268. [Google Scholar] [CrossRef]
- Janashia, I.; Choiset, Y.; Rabesona, H.; Hwanhlem, N.; Bakuradze, N.; Chanishvili, N.; Haertlé, T. Protection of Honeybee Apis Mellifera by Its Endogenous and Exogenous Lactic Flora against Bacterial Infections. Ann. Agrar. Sci. 2016, 14, 177–181. [Google Scholar] [CrossRef]
- Lee, I.-C.; Caggianiello, G.; van Swam, I.I.; Taverne, N.; Meijerink, M.; Bron, P.A.; Spano, G.; Kleerebezem, M. Strain-Specific Features of Extracellular Polysaccharides and Their Impact on Lactobacillus Plantarum-Host Interactions. Appl. Environ. Microbiol. 2016, 82, 3959. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.-C.; Neu, T.R.; Wozniak, D.J. The EPS Matrix: The “House of Biofilm Cells”. J. Bacteriol. 2007, 189, 7945. [Google Scholar] [CrossRef] [Green Version]
- Limoli, D.H.; Jones, C.J.; Wozniak, D.J. Bacterial Extracellular Polysaccharides in Biofilm Formation and Function. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Neu, T.R.; Lawrence, J.R. Chapter 37-Extracellular polymeric substances in microbial biofilms. In Microbial Glycobiology; Holst, O., Brennan, P.J., von Itzstein, M., Moran, A.P., Eds.; Academic Press: San Diego, CA, USA, 2010; pp. 733–758. ISBN 978-0-12-374546-0. [Google Scholar]
- Butler, È.; Alsterfjord, M.; Olofsson, T.C.; Karlsson, C.; Malmström, J.; Vásquez, A. Proteins of Novel Lactic Acid Bacteria from Apis Mellifera Mellifera: An Insight into the Production of Known Extra-Cellular Proteins during Microbial Stress. BMC Microbiol. 2013, 13, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karatan, E.; Watnick, P. Signals, Regulatory Networks, and Materials That Build and Break Bacterial Biofilms. Microbiol. Mol. Biol. Rev. 2009, 73, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caggianiello, G.; Kleerebezem, M.; Spano, G. Exopolysaccharides Produced by Lactic Acid Bacteria: From Health-Promoting Benefits to Stress Tolerance Mechanisms. Appl. Microbiol. Biotechnol. 2016, 100, 3877–3886. [Google Scholar] [CrossRef]
- Weirich, G.F.; Collins, A.M.; Williams, V.P. Antioxidant Enzymes in the Honey Bee, Apis Mellifera. Apidologie 2002, 33, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Nikolić, T.V.; Purać, J.; Orčić, S.; Kojić, D.; Vujanović, D.; Stanimirović, Z.; Gržetić, I.; Ilijević, K.; Šikoparija, B.; Blagojević, D.P. Environmental effects on superoxide dismutase and catalase activity and expression in honey bee. Arch. Insect Biochem. Physiol. 2015, 90, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.M.; Williams, V.; Evans, J.D. Sperm Storage and Antioxidative Enzyme Expression in the Honey Bee, Apis Mellifera. Insect Mol. Biol. 2004, 13, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Balieira, K.V.B.; Mazzo, M.; Bizerra, P.F.V.; Guimarães, A.R.D.J.S.; Nicodemo, D.; Mingatto, F.E. Imidacloprid-Induced Oxidative Stress in Honey Bees and the Antioxidant Action of Caffeine. Apidologie 2018, 49, 562–572. [Google Scholar] [CrossRef] [Green Version]
- Morimoto, T.; Kojima, Y.; Toki, T.; Komeda, Y.; Yoshiyama, M.; Kimura, K.; Nirasawa, K.; Kadowaki, T. The Habitat Disruption Induces Immune-Suppression and Oxidative Stress in Honey Bees. Ecol. Evol. 2011, 1, 201–217. [Google Scholar] [CrossRef]
- Banerjee, B.D.; Seth, V.; Ahmed, R.S. Pesticide-Induced Oxidative Stress: Perspective and Trends. Rev. Environ. Health 2001, 16, 1–40. [Google Scholar] [CrossRef]
- Simone-Finstrom, M.; Li-Byarlay, H.; Huang, M.H.; Strand, M.K.; Rueppell, O.; Tarpy, D.R. Migratory Management and Environmental Conditions Affect Lifespan and Oxidative Stress in Honey Bees. Sci. Rep. 2016, 6, 32023. [Google Scholar] [CrossRef] [PubMed]
- Claudianos, C.; Ranson, H.; Johnson, R.M.; Biswas, S.; Schuler, M.A.; Berenbaum, M.R.; Feyereisen, R.; Oakeshott, J.G. A Deficit of Detoxification Enzymes: Pesticide Sensitivity and Environmental Response in the Honeybee. Insect Mol. Biol. 2006, 15, 615–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belsky, J.; Joshi, N.K. Impact of Biotic and Abiotic Stressors on Managed and Feral Bees. Insects 2019, 10, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drossart, M.; Gérard, M. Beyond the Decline of Wild Bees: Optimizing Conservation Measures and Bringing Together the Actors. Insects 2020, 11, 649. [Google Scholar] [CrossRef]
- Blot, N.; Veillat, L.; Rouzé, R.; Delatte, H. Glyphosate, but Not Its Metabolite AMPA, Alters the Honeybee Gut Microbiota. PLoS ONE 2019, 14, e0215466. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ma, W.; Shen, J.; Long, D.; Feng, Y.; Su, W.; Xu, K.; Du, Y.; Jiang, Y. Tolerance and Response of Two Honeybee Species Apis Cerana and Apis Mellifera to High Temperature and Relative Humidity. PLoS ONE 2019, 14, e0217921. [Google Scholar] [CrossRef] [Green Version]
- Paris, L.; Roussel, M.; Pereira, B.; Delbac, F.; Diogon, M. Disruption of Oxidative Balance in the Gut of the Western Honeybee Apis Mellifera Exposed to the Intracellular Parasite Nosema Ceranae and to the Insecticide Fipronil. Microb. Biotechnol. 2017, 10, 1702–1717. [Google Scholar] [CrossRef]
- Feldhaar, H.; Otti, O. Pollutants and Their Interaction with Diseases of Social Hymenoptera. Insects 2020, 11, 153. [Google Scholar] [CrossRef] [Green Version]
- Seehuus, S.-C.; Norberg, K.; Gimsa, U.; Krekling, T.; Amdam, G.V. Reproductive Protein Protects Functionally Sterile Honey Bee Workers from Oxidative Stress. Proc. Natl. Acad. Sci. USA 2006, 103, 962. [Google Scholar] [CrossRef] [Green Version]
- Feng, T.; Wang, J. Oxidative Stress Tolerance and Antioxidant Capacity of Lactic Acid Bacteria as Probiotic: A Systematic Review. Gut Microbes 2020, 12, 1801944. [Google Scholar] [CrossRef]
- Lin, M.-Y.; Yen, C.-L. Antioxidative Ability of Lactic Acid Bacteria. J. Agric. Food Chem. 1999, 47, 1460–1466. [Google Scholar] [CrossRef] [PubMed]
- Mishra, V.; Shah, C.; Mokashe, N.; Chavan, R.; Yadav, H.; Prajapati, J. Probiotics as Potential Antioxidants: A Systematic Review. J. Agric. Food Chem. 2015, 63, 3615–3626. [Google Scholar] [CrossRef] [PubMed]
- Das, D. Arun Goyal Antioxidant Activity and γ-Aminobutyric Acid (GABA) Producing Ability of Probiotic Lactobacillus Plantarum DM5 Isolated from Marcha of Sikkim. Lebensm. Wiss. Ie Technol. Food Sci. Technol. Sci. Technol. Aliment. 2015, 61, 263–268. [Google Scholar] [CrossRef]
- Kullisaar, T.; Zilmer, M.; Mikelsaar, M.; Vihalemm, T.; Annuk, H.; Kairane, C.; Kilk, A. Two Antioxidative Lactobacilli Strains as Promising Probiotics. Int. J. Food Microbiol. 2002, 72, 215–224. [Google Scholar] [CrossRef]
- Chanput, W.; Krueyos, N.; Ritthiruangdej, P. Anti-Oxidative Assays as Markers for Anti-Inflammatory Activity of Flavonoids. Int. Immunopharmacol. 2016, 40, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Cano, A.; Acosta, M. The Hydrophilic and Lipophilic Contribution to Total Antioxidant Activity. Food Chem. 2001, 73, 239–244. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| L. plantarum Strains | |||||
|---|---|---|---|---|---|
| LP8 | LP25 | LP86 | LP95 | LP100 | |
| CFSLM | 37.45 ± 0.40 c | 36.88 ± 0.40 c | 25.73 ± 0.81 b | 22.30 ± 0.05 a | 20.01 ± 0.81 a |
| antioxidant activity * | |||||
| CES * | 0.17 ± 0.02 b | 0.16 ± 0.01 b | 0.12 ± 0.01 a | 0.14 ± 0.01 a | 0.11 ± 0.00 a |
| antioxidant activity ** | |||||
| EPS-r | 1.40 ± 0.03 b | 1.56 ± 0.50 c | 1.28 ± 0.10 b | 0.82 ± 0.07 a | 1.49 ± 0.06 b |
| EPS-b | 8.82 ± 0.11 d | 2.23 ± 0.09 a | 5.05 ± 0.12 b | 1.96 ± 0.08 a | 5.54 ± 0.10 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iorizzo, M.; Testa, B.; Ganassi, S.; Lombardi, S.J.; Ianiro, M.; Letizia, F.; Succi, M.; Tremonte, P.; Vergalito, F.; Cozzolino, A.; et al. Probiotic Properties and Potentiality of Lactiplantibacillus plantarum Strains for the Biological Control of Chalkbrood Disease. J. Fungi 2021, 7, 379. https://doi.org/10.3390/jof7050379
Iorizzo M, Testa B, Ganassi S, Lombardi SJ, Ianiro M, Letizia F, Succi M, Tremonte P, Vergalito F, Cozzolino A, et al. Probiotic Properties and Potentiality of Lactiplantibacillus plantarum Strains for the Biological Control of Chalkbrood Disease. Journal of Fungi. 2021; 7(5):379. https://doi.org/10.3390/jof7050379
Chicago/Turabian StyleIorizzo, Massimo, Bruno Testa, Sonia Ganassi, Silvia Jane Lombardi, Mario Ianiro, Francesco Letizia, Mariantonietta Succi, Patrizio Tremonte, Franca Vergalito, Autilia Cozzolino, and et al. 2021. "Probiotic Properties and Potentiality of Lactiplantibacillus plantarum Strains for the Biological Control of Chalkbrood Disease" Journal of Fungi 7, no. 5: 379. https://doi.org/10.3390/jof7050379