
Microbiome Status of Cider-Apples, from Orchard to Processing, with a Special Focus on Penicillium expansum Occurrence and Patulin Contamination

 ,
, 
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Overview of the Apple Sampling and Handling Processes
2.2. Culture Dependent (Plating) Analysis of the Apple Surface Microbiota
2.3. Extraction and Analysis of Patulin in Apples
2.4. DNA Isolation from the Surface of the Apples
2.5. Quantification of P. expansum on the Surface of Apples Using Real-Time PCR
2.6. DNA Amplification and Sequencing
2.6.1. Library Construction and Sequencing
2.6.2. Data Analysis and Statistics
3. Results
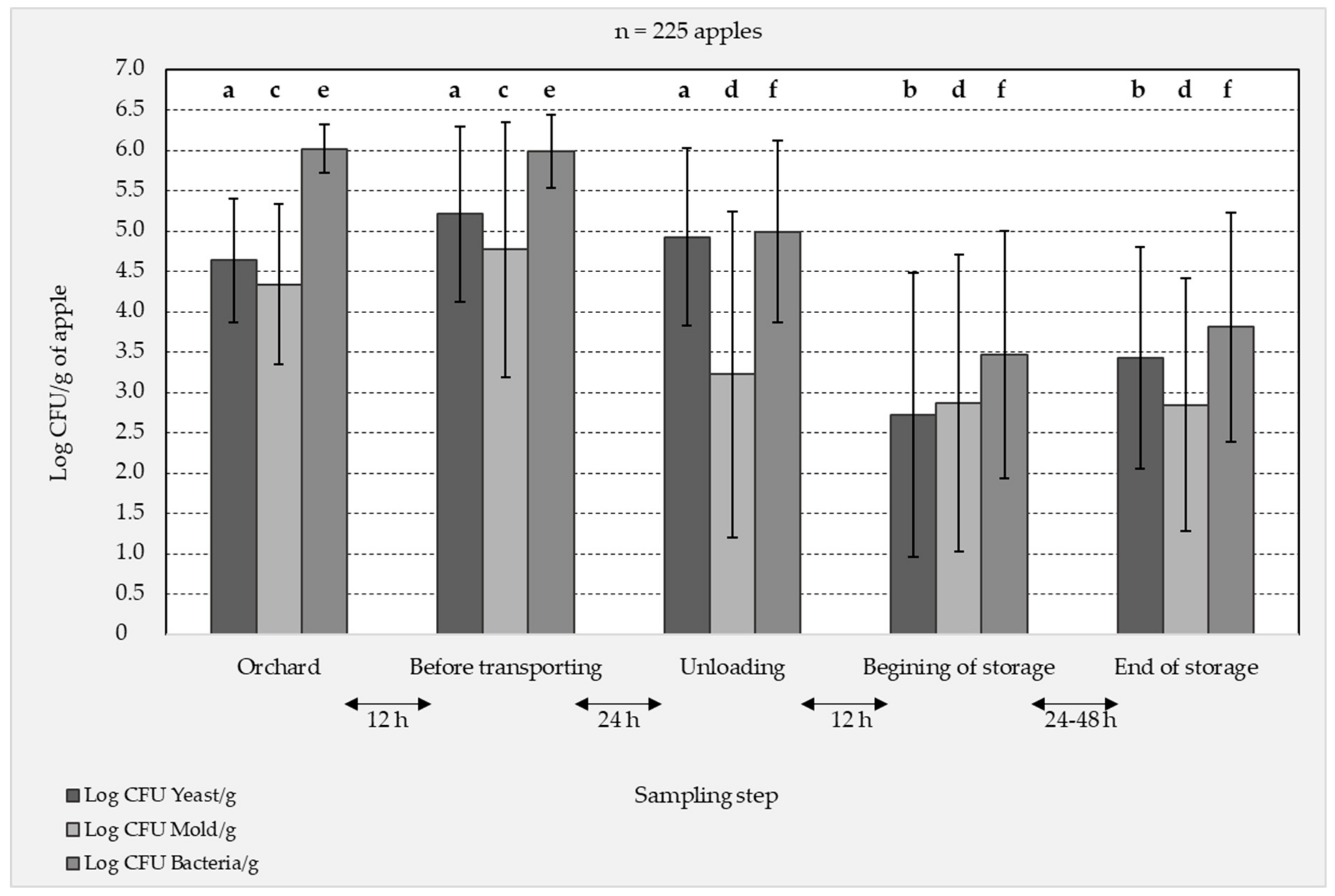
3.1. Culture Dependent (Plating) Analysis of the Apple Surface Microbiota
3.2. Quantification of Patulin Content in Apples by HPLC-MS/MS
3.3. Quantification of P. expansum on the Surface of Apples Using Real-Time PCR
3.4. General Structure of the Bacterial and Fungal Microbiota of the Surface of Cider-Apple
3.4.1. Sequencing Results
3.4.2. Epiphytic Fungal and Bacterial Microbiota of Cider-Apples
3.4.3. Compositional Differences in the Diversity and the Taxonomy of Fungal and Bacterial Microbiota Induced by the Different Sampling Steps
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| P. expansum | Penicillium expansum |
| IARC | International Agency for Research on Cancer |
| EU | European Union |
| CFU | Colony-forming unit |
| ITS | Internal transcribed spacer |
| ASV | Amplicon sequence variant |
| RA | Relative abundance |
| PGPR | Plant growth promoting rhizobacteria |
| AAB | Acetic acid bacteria |
References
- Cousin, F.J.; Le Guellec, R.; Schlusselhuber, M.; Dalmasso, M.; LaPlace, J.-M.; Cretenet, M. Microorganisms in Fermented Apple Beverages: Current Knowledge and Future Directions. Microorganisms 2017, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Lea, A.G.H.; Piggott, J.R. Fermented Beverage Production, 2nd ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2003. [Google Scholar]
- European Cider and Fruit Wine Association (AICV). AICV Cider Trends. 2020. Available online: https://aicv.org/en/publications (accessed on 9 November 2020).
- Robinson, R.K.; Batt, C.A. Encyclopedia of Food Microbiology; Elsevier Science: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Morales, H.; Sanchis, V.; Usall, J.; Ramos, A.J.; Marín, S. Effect of biocontrol agents Candida sake and Pantoea agglomerans on Penicillium expansum growth and patulin accumulation in apples. Int. J. Food Microbiol. 2008, 122, 61–67. [Google Scholar] [CrossRef]
- Sydenham, E.W.; Vismer, H.F.; Marasas, W.F.; Brown, N.; Schlechter, M.; Van Der Westhuizen, L.; Rheeder, J.P. Reduction of patulin in apple juice samples—Influence of initial processing. Food Control 1995, 6, 195–200. [Google Scholar] [CrossRef]
- Sanzani, S.; Reverberi, M.; Punelli, M.; Ippolito, A.; Fanelli, C. Study on the role of patulin on pathogenicity and virulence of Penicillium expansum. Int. J. Food Microbiol. 2012, 153, 323–331. [Google Scholar] [CrossRef]
- Doores, S.; Splittstoesser, D.F. The microbiology of apples and apple products. Crit. Rev. Food Sci. Nutr. 1983, 19, 133–149. [Google Scholar] [CrossRef] [PubMed]
- Soliman, S.; Li, X.-Z.; Shao, S.; Behar, M.; Svircev, A.M.; Tsao, R.; Zhou, T. Potential mycotoxin contamination risks of apple products associated with fungal flora of apple core. Food Control 2015, 47, 585–591. [Google Scholar] [CrossRef]
- Sant’Ana, A.S.; Simas, R.C.; Almeida, C.A.; Cabral, E.C.; Rauber, R.H.; Mallmann, C.A.; Eberlin, M.N.; Rosenthal, A.; Massaguer, P.R. Influence of package, type of apple juice and temperature on the production of patulin by Byssochlamys nivea and Byssochlamys fulva. Int. J. Food Microbiol. 2010, 142, 156–163. [Google Scholar] [CrossRef] [PubMed]
- McKinley, E.; Carlton, W. Patulin mycotoxicosis in Swiss ICR mice. Food Cosmet. Toxicol. 1980, 18, 181–187. [Google Scholar] [CrossRef]
- Mckinley, E.R.; Carlton, W.W.; Boon, G.D. Patulin mycotoxicosis in the rat: Toxicology, pathology, and clinical pathology. Food Chem. Toxicol. 1982, 20, 289–300. [Google Scholar] [CrossRef]
- Pal, S.; Singh, N.; Ansari, K.M. Toxicological effects of patulin mycotoxin on the mammalian system: An overview. Toxicol. Res. 2017, 6, 764–771. [Google Scholar] [CrossRef] [Green Version]
- Puel, O.; Galtier, P.; Oswald, I.P. Biosynthesis and Toxicological Effects of Patulin. Toxins 2010, 2, 613–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, A.; Ouhibi, S.; Ghali, R.; Hedhili, A.; De Saeger, S.; De Boevre, M. The mycotoxin patulin: An updated short review on occurrence, toxicity and analytical challenges. Food Chem. Toxicol. 2019, 129, 249–256. [Google Scholar] [CrossRef]
- The Commission of the European Communities (EC) No 1425/2003. The prevention and reduction of patulin contamination in apple juice and apple juice having regard to the treaty establishing the European Community, and in particular the second indent of Artic, 11 August 2003. Off. J. Eur. Union 2003, L203, 54–59. [Google Scholar]
- The Commission of the European Communities (EC) No 1881/2006 of 20.12.2006. Setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, L 364, 5–24. [Google Scholar]
- IFPC. Pollution du Fruit et Patuline Cidre, Pomme, Pommier. Available online: http://www.ifpc.eu/bibliographie/recolte-qualite-des-fruits/qualite-du-fruit.html (accessed on 10 November 2020).
- Harris, K.L.; Bobe, G.; Bourquin, L.D. Patulin Surveillance in Apple Cider and Juice Marketed in Michigan. J. Food Prot. 2009, 72, 1255–1261. [Google Scholar] [CrossRef]
- Zambounis, A.; Ganopoulos, I.; Tsaftaris, A.; Valasiadis, D.; Madesis, P. Metagenomics analysis of fungal communities associated with postharvest diseases in pear fruits under the effect of management practices. Arch. Microbiol. 2020, 202, 1–10. [Google Scholar] [CrossRef]
- Dachoupakan, C.; Ratomahenina, R.; Martinez, V.; Guiraud, J.; Baccou, J.; Schorr-Galindo, S. Study of the phenotypic and genotypic biodiversity of potentially ochratoxigenic black aspergilli isolated from grapes. Int. J. Food Microbiol. 2009, 132, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, H.; Strub, C.; Hilaire, F.; Schorr-Galindo, S. First report: Penicillium adametzioides, a potential biocontrol agent for ochratoxin-producing fungus in grapes, resulting from natural product pre-harvest treatment. Food Control 2015, 51, 23–30. [Google Scholar] [CrossRef]
- Macdonald, S.; Long, M.; Gilbert, J.; Felgueiras, I. Liquid chromatographic method for determination of patulin in clear and cloudy apple juices and apple puree: Collaborative study. J. Assoc. Off. Anal. Chem. 2000, 83, 1387–1394. [Google Scholar]
- Tannous, J.; Atoui, A.; El Khoury, A.; Kantar, S.; Chdid, N.; Oswald, I.P.; Puel, O.; Lteif, R. Development of a real-time PCR assay for Penicillium expansum quantification and patulin estimation in apples. Food Microbiol. 2015, 50, 28–37. [Google Scholar] [CrossRef]
- Vancov, T.; Keen, B. Amplification of soil fungal community DNA using the ITS86Fand ITS4 primers. FEMS Microbiol. Lett. 2009, 296, 91–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, R.H.; Larsson, K.-H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef] [PubMed]
- Wassermann, B.; Kusstatscher, P.; Berg, G. Microbiome response to hot water treatment and potential synergy with biological control on stored apples. Front. Microbiol. 2019, 10, 2502. [Google Scholar] [CrossRef]
- Shen, Y.; Nie, J.; Li, Z.; Li, H.; Wu, Y.; Dong, Y.; Zhang, J. Differentiated surface fungal communities at point of harvest on apple fruits from rural and peri-urban orchards. Sci. Rep. 2018, 8, 1–12. [Google Scholar]
- Abdelfattah, A.; Wisniewski, M.; Droby, S.; Schena, L. Spatial and compositional variation in the fungal communities of organic and conventionally grown apple fruit at the consumer point-of-purchase. Hortic. Res. 2016, 3, 16047. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Abdelfattah, A.; Norelli, J.; Burchard, E.; Schena, L.; Droby, S.; Wisniewski, M. Apple endophytic microbiota of different rootstock/scion combinations suggests a genotype-specific influence. Microbiome 2018, 6, 1–11. [Google Scholar] [CrossRef]
- Alonso, S.; Laca, A.; Rendueles, M.; Mayo, B.; Díaz, M. Cider Apple Native Microbiota Characterization by PCR-DGGE. J. Inst. Brew. 2015, 121, 287–289. [Google Scholar] [CrossRef]
- Misery, B.; Legendre, P.; Rue, O.; Bouchart, V.; Guichard, H.; Laplace, J.; Cretenet, M. Diversity and dynamics of bacterial and fungal communities in cider for distillation. Int. J. Food Microbiol. 2021, 339, 108987. [Google Scholar] [CrossRef]
- Leff, J.W.; Fierer, N. Bacterial Communities Associated with the Surfaces of Fresh Fruits and Vegetables. PLoS ONE 2013, 8, e59310. [Google Scholar] [CrossRef] [Green Version]
- Wisniewski, M.; Droby, S. The postharvest microbiome: The other half of sustainability. Biol. Control 2019, 137. [Google Scholar] [CrossRef]
- Tournas, V.; Memon, S.U. Internal contamination and spoilage of harvested apples by patulin-producing and other toxigenic fungi. Int. J. Food Microbiol. 2009, 133, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Qiao, N.; Zhao, J.; Zhang, H.; Tian, F.; Zhai, Q.; Chen, W. Postharvest control of Penicillium expansum in fruits: A review. Food Biosci. 2020, 36, 100633. [Google Scholar] [CrossRef]
- Jackson, L.S.; Beacham-Bowden, T.; Keller, S.E.; Adhikari, C.; Taylor, K.T.; Chirtel, S.J.; Merker, R.I. Apple Quality, Storage, and Washing Treatments Affect Patulin Levels in Apple Cider. J. Food Prot. 2003, 66, 618–624. [Google Scholar] [CrossRef]
- Baert, K.; Devlieghere, F.; Flyps, H.; Oosterlinck, M.; Ahmed, M.M.; Rajković, A.; Verlinden, B.; Nicolaï, B.; Debevere, J.; De Meulenaer, B. Influence of storage conditions of apples on growth and patulin production by Penicillium expansum. Int. J. Food Microbiol. 2007, 119, 170–181. [Google Scholar] [CrossRef] [PubMed]
- McCallum, J.L.; Tsao, R.; Zhou, T. Factors Affecting Patulin Production by Penicillium expansum. J. Food Prot. 2002, 65, 1937–1942. [Google Scholar] [CrossRef]
- Coates, L.; Johnson, G. Postharvest diseases of fruits and vegetables. Plant Pathog. Plant Dis. 1997, 33, 534–548. [Google Scholar]
- Konstantinou, S.; Karaoglanidis, G.S.; Bardas, G.A.; Minas, I.S.; Doukas, E.; Markoglou, A.N. Postharvest Fruit Rots of Apple in Greece: Pathogen Incidence and Relationships Between Fruit Quality Parameters, Cultivar Susceptibility, and Patulin Production. Plant. Dis. 2011, 95, 666–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, J.W.; Hewett, E.W.; Hertog, M.L.A.T.M. Postharvest softening of apple (Malus domestica) fruit: A review. N. Z. Crop. Hortic. Sci. 2002, 30, 145–160. [Google Scholar] [CrossRef]
- Damoglou, A.P.; Campbell, D.S. The effect of pH on the production of patulin in apple juice. Lett. Appl. Microbiol. 1986, 2, 9–11. [Google Scholar] [CrossRef]
- Castoria, R.; Mannina, L.; Durán-Patrón, R.; Maffei, F.; Sobolev, A.P.; De Felice, D.V.; Pinedo-Rivilla, C.; Ritieni, A.; Ferracane, R.; Wright, S.A.I. Conversion of the Mycotoxin Patulin to the Less Toxic Desoxypatulinic Acid by the Biocontrol Yeast Rhodosporidium kratochvilovae Strain LS11. J. Agric. Food Chem. 2011, 59, 11571–11578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinedo, C.; Wright, S.A.I.; Collado, I.G.; Goss, R.J.M.; Castoria, R.; Hrelia, P.; Maffei, F.; Durán-Patrón, R. Isotopic Labeling Studies Reveal the Patulin Detoxification Pathway by the Biocontrol Yeast Rhodotorula kratochvilovae LS11. J. Nat. Prod. 2018, 81, 2692–2699. [Google Scholar] [CrossRef]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T.; Kirk, P.M.; Lücking, R.; et al. A higher-level phylogenetic classification of the Fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef]
- Houbraken, J.; Kocsub, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.-C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.-A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 169, 5–169. [Google Scholar] [CrossRef] [PubMed]
- Félix, C.R.; Andrade, D.A.; Almeida, J.H.; Navarro, H.M.C.; Fell, J.W.; Landell, M.F. Vishniacozyma alagoana sp. nov. a tremellomycetes yeast associated with plants from dry and rainfall tropical forests. Int. J. Syst. Evol. Microbiol. 2020, 70, 3449–3454. [Google Scholar] [CrossRef] [PubMed]
- Vadkertiová, R.; Dudášová, H.; Stratilová, E.; Balaščáková, M. Diversity of yeasts in the soil adjacent to fruit trees of the Rosaceae family. Yeast 2019, 36, 617–631. [Google Scholar] [CrossRef] [PubMed]
- Landi, L.; De Miccolis Angelini, R.M.; Pollastro, S.; Abate, D.; Faretra, F.; Romanazzi, G. Genome sequence of the brown rot fungal pathogen Monilinia fructigena 06 Biological Sciences 0604 Genetics 06 Biological Sciences 0607 Plant Biology. BMC Res. Notes 2018, 11, 10–12. [Google Scholar]
- Gao, L.L.; Zhang, Q.; Sun, X.Y.; Jiang, L.; Zhang, R.; Sun, G.Y.; Zha, Y.L.; Biggs, A.R. Etiology of Moldy Core, Core Browning, and Core Rot of Fuji Apple in China. Plant. Dis. 2013, 97, 510–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stošić, S.; Ristić, D.; Gašić, K.; Starović, M.; Grbić, M.L.; Vukojević, J.; Živković, S. Talaromyces minioluteus: New Postharvest Fungal Pathogen in Serbia. Plant. Dis. 2020, 104, 656–667. [Google Scholar] [CrossRef] [PubMed]
- Karaman, K.; Sagdic, O.; Yilmaz, M.T. Potential of natamycin to control growth of Zygosaccharomyces spp. in apple juice during storage. Int. J. Food Microbiol. 2020, 332, 108771. [Google Scholar] [CrossRef]
- Sui, Y.; Wisniewski, M.; Droby, S.; Piombo, E.; Wu, X.; Yue, J. Genome Sequence, Assembly, and Characterization of the Antagonistic Yeast Candida oleophila Used as a Biocontrol Agent Against Post-harvest Diseases. Front. Microbiol. 2020, 11, 295. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Peng, H.M.; Wang, X.; Li, B.Q.; Long, M.Y.; Tian, S.P. Biodegradation mechanisms of patulin in Candida guilliermondii: An iTRAQ-based proteomic analysis. Toxins 2017, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C. A critical review of producers of small lactone mycotoxins: Patulin, penicillic acid and moniliformin. Abstract 2018, 11, 73–100. [Google Scholar] [CrossRef]
- Deinhard, G.; Blanz, P.; Poralla, K.; Altan, E. Bacillus acidoterrestris sp. nov., a New Thermotolerant Acidophile Isolated from Different Soils. Syst. Appl. Microbiol. 1987, 10, 47–53. [Google Scholar] [CrossRef]
- Mehta, P.; Walia, A.; Kakkar, N.; Shirkot, C.K. Tricalcium phosphate solubilisation by new endophyte Bacillus methylotrophicus CKAM isolated from apple root endosphere and its plant growth-promoting activities. Acta Physiol. Plant. 2014, 36, 2033–2045. [Google Scholar] [CrossRef]
- Ju, R.; Zhao, Y.; Li, J.; Jiang, H.; Liu, P.; Yang, T.; Bao, Z.; Zhou, B.; Zhou, X.; Liu, X. Identification and evaluation of a potential biocontrol agent, Bacillus subtilis, against Fusarium sp. in apple seedlings. Ann. Microbiol. 2014, 64, 377–383. [Google Scholar] [CrossRef]
- Walsh, F.; Smith, D.P.; Owens, S.M.; Duffy, B.; Frey, J.E. Restricted streptomycin use in apple orchards did not adversely alter the soil bacteria communities. Front. Microbiol. 2014, 4, 383. [Google Scholar] [CrossRef]
- Rivas-Garcia, T.; Murillo-Amador, B.; Nieto-Garibay, A.; Chiquito-Contreras, R.G.; Rincon-Enriquez, G.; Hernandez-Montiel, L.G. Effect of Ulvan on the Biocontrol Activity of Debaryomyces hansenii and Stenotrophomonas rhizophila against Fruit Rot of Cucumis melo L. Agronomy 2018, 8, 273. [Google Scholar] [CrossRef] [Green Version]
- Bevardi, M.; Frece, J.; Mesarek, D.; Bošnir, J.; Mrvčić, J.; Delaš, F.; Markov, K. Antifungal and Antipatulin Activity of Gluconobacter oxydans Isolated from Apple Surface. Arch. Ind. Hyg. Toxicol. 2013, 64, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Mari, L.; Cleenwerck, I.; Accetto, T.; Vandamme, P.; Tr, J. From Apple Cider Vinegar. 2000, pp. 1–16. Available online: https://www.webmd.com/diet/supplement-guide-apple-cider-vinegar#1 (accessed on 16 February 2021).
- Kim, Y.S.; Balaraju, K.; Jeon, Y. Effects of rhizobacteria Paenibacillus polymyxa APEC136 and Bacillus subtilis APEC170 on biocontrol of postharvest pathogens of apple fruits. J. Zhejiang Univ. Sci. B 2016, 17, 931–940. [Google Scholar] [CrossRef]
- Errampalli, D. Penicillium expansum (Blue Mold); Elsevier BV: Amsterdam, The Netherlands, 2014; pp. 189–231. [Google Scholar]
- Zhong, L.; Carere, J.; Lu, Z.; Lu, F.; Zhou, T. Patulin in Apples and Apple-Based Food Products: The Burdens and the Mitigation Strategies. Toxins 2018, 10, 475. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) |
|---|---|
| Pexp_patF_F | ATGAAATCCTCCCTGTGGGTTAGT |
| Pexp_patF_R | GAAGGATAATTTCCGGGGTAGTCATT |
| ITS86F | GTGAATCATCGAATCTTTGAA |
| ITS4R | TCCTCCGCTTATTGATATGC |
| 341F | CCTACGGGNGGCWGCAG a |
| 785R | GACTACHVGGGTATCTAATCC |
| Sampling Step | n 1 | Percentage of Positive Samples 2 | Patulin Content Range 3 (µg·kg−1 of Apple) | Tukey Grouping 4 |
|---|---|---|---|---|
| Orchard | 16 | 6 | 0–280 | a |
| Before transporting | 19 | 47 | 0–2943 | b |
| Unloading | 19 | 95 | 0–1169 | ab |
| Beginning of storage | 13 | 69 | 0–909 | ab |
| Sampling Steps | n 1 | Percentage of Positive Samples | Mean ± SE 2 (Log DNA P. expansum/g of Apple) |
|---|---|---|---|
| Orchard | 26 | 88.46 | 4.21 ± 2.0 |
| Before transporting | 19 | 84.21 | 4.26 ± 2.2 |
| Unloading | 19 | 84.21 | 3.99 ± 2.3 |
| Beginning of storage | 19 | 84.21 | 4.25 ± 2.4 |
| End of storage | 19 | 84.21 | 4.09 ± 2.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Riachy, R.; Strub, C.; Durand, N.; Guibert, B.; Guichard, H.; Constancias, F.; Chochois, V.; Lopez-Lauri, F.; Fontana, A.; Schorr-Galindo, S. Microbiome Status of Cider-Apples, from Orchard to Processing, with a Special Focus on Penicillium expansum Occurrence and Patulin Contamination. J. Fungi 2021, 7, 244. https://doi.org/10.3390/jof7040244
Al Riachy R, Strub C, Durand N, Guibert B, Guichard H, Constancias F, Chochois V, Lopez-Lauri F, Fontana A, Schorr-Galindo S. Microbiome Status of Cider-Apples, from Orchard to Processing, with a Special Focus on Penicillium expansum Occurrence and Patulin Contamination. Journal of Fungi. 2021; 7(4):244. https://doi.org/10.3390/jof7040244
Chicago/Turabian StyleAl Riachy, Reem, Caroline Strub, Noël Durand, Benjamin Guibert, Hugues Guichard, Florentin Constancias, Vincent Chochois, Félicie Lopez-Lauri, Angélique Fontana, and Sabine Schorr-Galindo. 2021. "Microbiome Status of Cider-Apples, from Orchard to Processing, with a Special Focus on Penicillium expansum Occurrence and Patulin Contamination" Journal of Fungi 7, no. 4: 244. https://doi.org/10.3390/jof7040244