
Entomopathogenic Fungi Biodiversity in the Soil of Three Provinces Located in Southwest China and First Approach to Evaluate Their Biocontrol Potential



Abstract
:1. Introduction
2. Materials and Methods
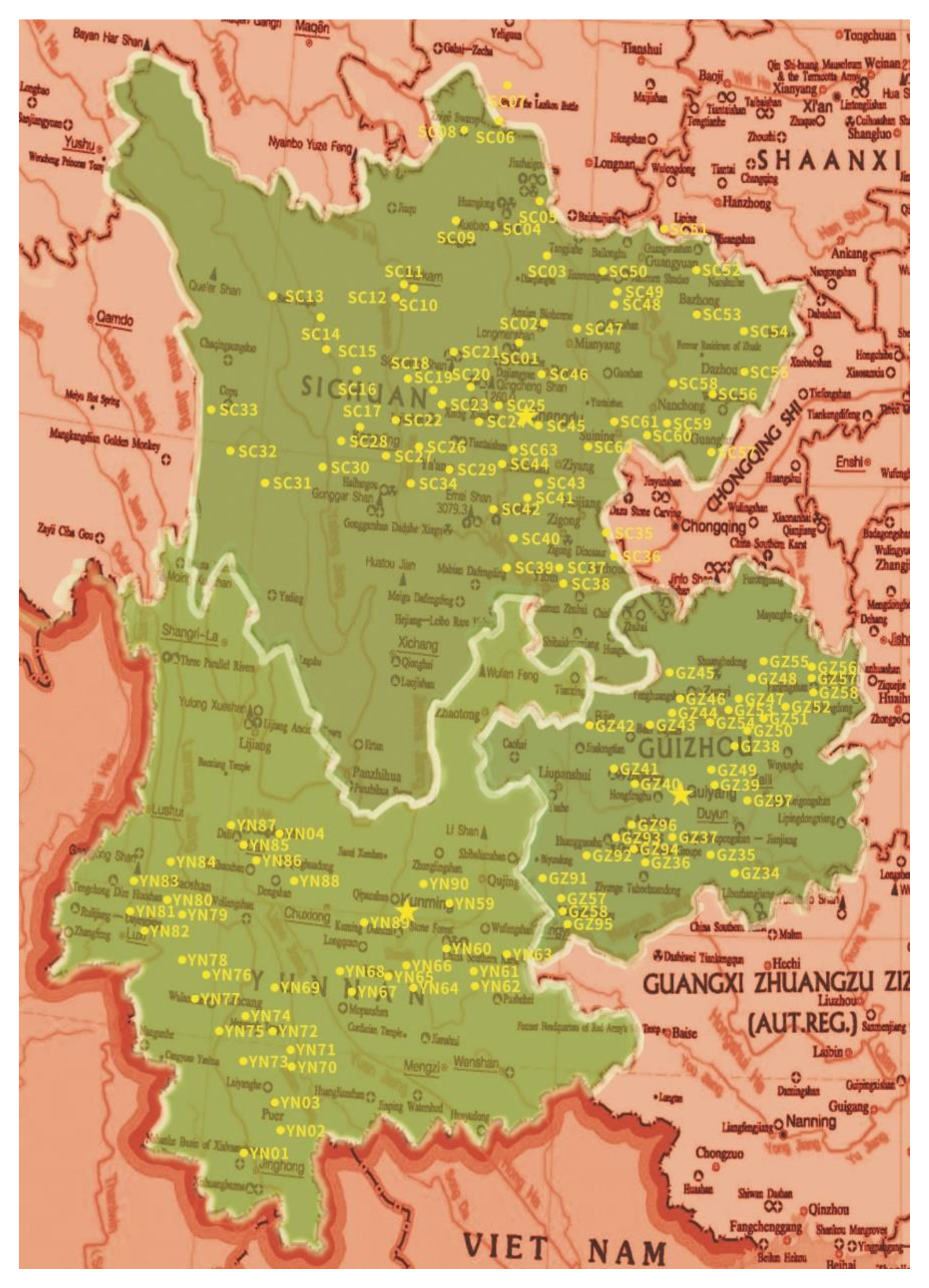
2.1. Soil Sample Collection
2.2. Isolation of Fungi from the Soil Samples
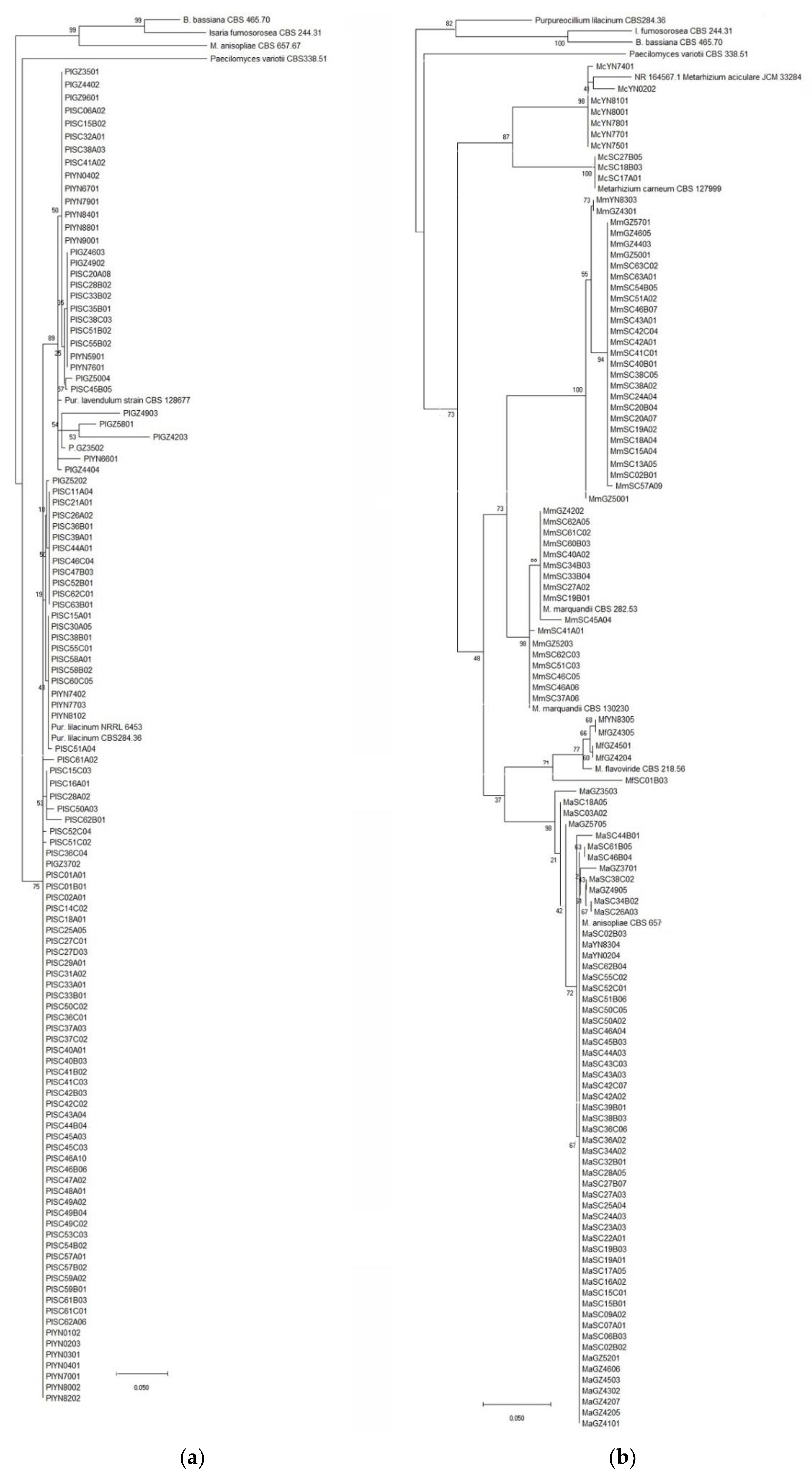
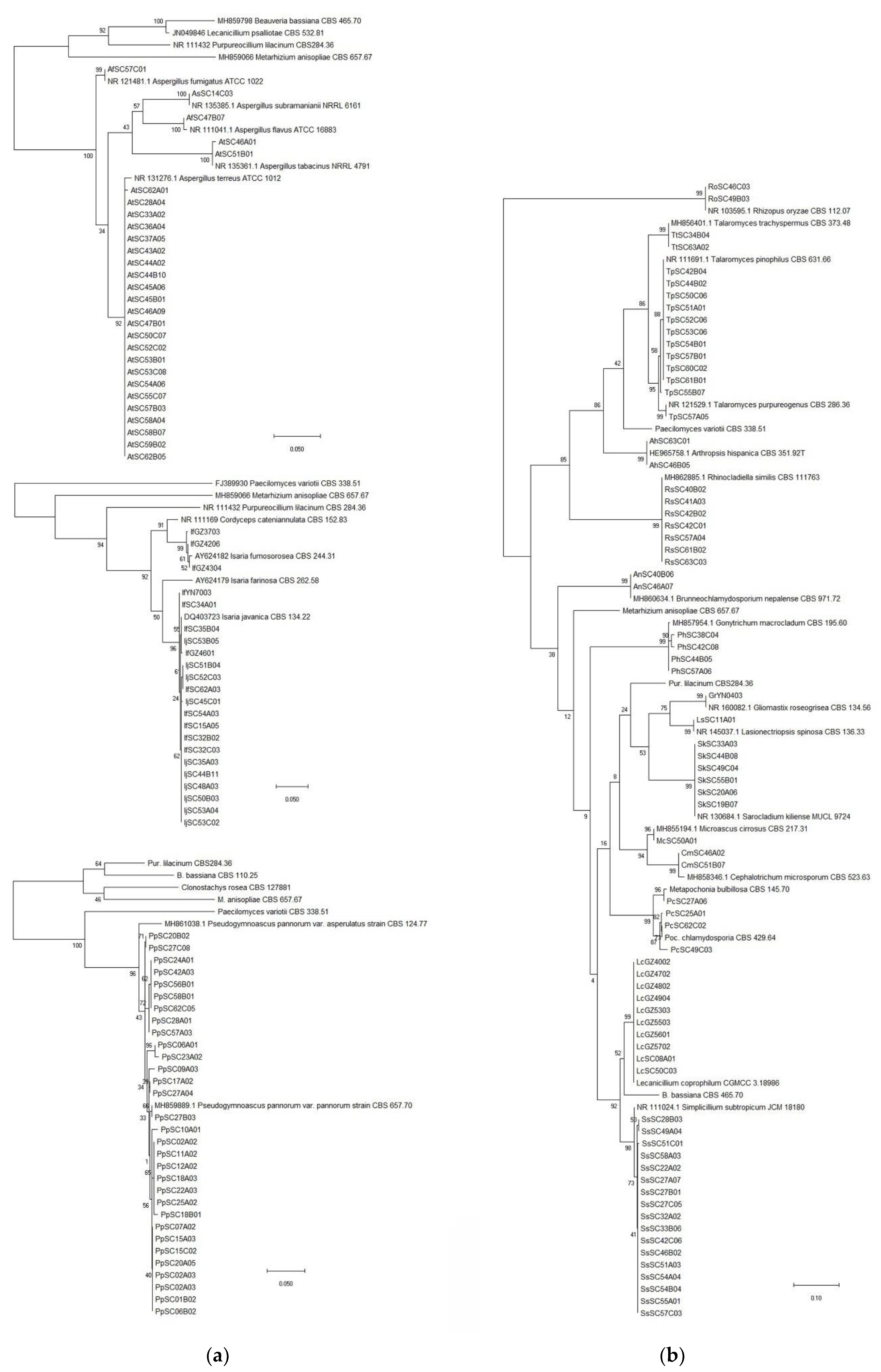
2.3. Identification of Fungal Isolates
2.4. Evaluation of Shannon–Wiener Index
2.5. Bioassay of Fungal Strains on B-Biotype Whitefly and Spodoptera litura
2.6. Statistical Analysis
3. Results
3.1. Entomopathogenic Fungi Species Diversity in Soils of Southwest China
3.2. The Distribution of Soil EF in Different Areas of Southwest China
3.3. The Biodiversity of EF in Different Soil Environments
3.4. The Pathogenicities of Fungal Isolates against B. tabaci and S. litura
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, C.; Wang, S. Insect Pathogenic Fungi: Genomics, Molecular Interactions, and Genetic Improvements. Annu. Rev. Entomol. 2017, 62, 73–90. [Google Scholar] [CrossRef]
- Arthurs, S.; Dara, S.K. Microbial biopesticides for invertebrate pests and their markets in the United States. J. Invertebr. Pathol. 2019, 165, 13–21. [Google Scholar] [CrossRef]
- Kumar, K.K.; Sridhar, J.; Murali-Baskaran, R.K.; Senthil-Nathan, S.; Kaushal, P.; Dara, S.K.; Arthurs, S. Microbial biopesticides for insect pest management in India: Current status and future prospects. J. Invertebr. Pathol. 2019, 165, 74–81. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Lopes, R.B.; Faria, M.; Delalibera, I., Jr.; Fernandes, E.K.K.; Luz, C. Current status and perspectives of fungal entomopathogens used for microbial control of arthropod pests in Brazil. J. Invertebr. Pathol. 2019, 165, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Saikia, S.; Bora, P.; Bora, L.C. Bioagent mediated management of citrus canker. Indian J. Agric. Sci. 2021, 91, 198–201. [Google Scholar]
- Xie, T.; Jiang, L.; Li, J.; Hong, B.; Wang, X.; Jia, Y. Effects of Lecanicillium lecanii strain JMC-01 on the physiology, biochemistry, and mortality of Bemisia tabaci Q-biotype nymphs. PeerJ 2019, 7, e7690. [Google Scholar] [CrossRef] [Green Version]
- Boomsma, J.J.; Jensen, A.B.; Meyling, N.V.; Eilenberg, J. Evolutionary interaction networks of insect pathogenic fungi. Annu. Rev. Entomol. 2014, 59, 467–485. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Gou, X.; Hillerislambers, J.; Deng, Y.; Manzanedo, R.D. Increasing climate sensitivity of subtropical conifers along an aridity gradient. For. Ecol. Manag. 2021, 482, 118841. [Google Scholar] [CrossRef]
- Meyling, N.V.; Eilenberg, J. Occurrence and distribution of soil borne entomopathogenic fungi within a single organic agroecosystem. Agric. Ecosyst. Environ. 2006, 113, 336–341. [Google Scholar] [CrossRef]
- Masoudi, A.; Koprowski, J.l.; Bhattarai, U.R.; Wang, D. Elevational distribution and morphological attributes of the entomopathogenic fungi from forests of the Qinling Mountains in China. Appl. Microbiol. Biotechnol. 2018, 102, 1483–1499. [Google Scholar] [CrossRef]
- Sumio, T.; Hiroaki, M.; Hidemi, K.; Oka, N.P.; Kwang-Shing, C. Variations in the occurrence patterns of male moths of the common cutworm, Spodoptera litura (Lepidoptera: Noctuidae) among Southeastern Asian countries, as detected by sex pheromone trapping. Appl. Entomol. Zool. 2008, 43, 569–576. [Google Scholar]
- Mound, L.A.; Halsey, S.H. Whitefly of the World. A Systematic Catalogue of the Aleyrodidae (Hemiptera) with Host Plant and Natural Enemy Data; British Musernu: London, UK, 1978; p. 340. [Google Scholar]
- Faria, M.; Wraight, S.P. Biological control of Bemisia tabaci with fungi. Crop. Prot. 2001, 20, 767–778. [Google Scholar] [CrossRef]
- Li, J.; Li, Q.; Wu, Y.; Ye, L.; Huang, X. Mountains act as museums and cradles for hemipteran insects in China: Evidence from patterns of richness and phylogenetic structure. Glob. Ecol. Biogeogr. 2021, 30, 1070–1085. [Google Scholar] [CrossRef]
- Liu, Y.; Shen, Z.; Wang, Q.; Su, X.; Zhang, W.; Shrestha, N.; Xu, X.; Wang, Z. Determinants of richness patterns differ between rare and common species: Implications for Gesneriaceae conservation in China. Divers. Distrib. 2017, 23, 235–246. [Google Scholar] [CrossRef]
- Hu, Q.B.; Ren, S.X.; Wu, J.H.; Chang, J.M.; Musa, P.D. Investigation of destruxin A and B from 80 Metarhizium strains in China, and the optimization of cultural conditions for the strain MaQ10. Toxicon 2006, 48, 491–498. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Chen, J.; Wu, J. Improvement of semi-artificial diet for Spodoptera litura. J. Northwest A F Univ. (Nat. Sci. Ed.) 2016, 44, 109–113. [Google Scholar]
- Boni, S.B.; Mwashimaha, R.A.; Mlowe, N.; Sotelo-Cardona, P.; Nordey, T. Efficacy of indigenous entomopathogenic fungi against the black aphid, Aphis fabae Scopoli under controlled conditions in Tanzania. Int. J. Trop. Insect Sci. 2021, 41, 1643–1651. [Google Scholar] [CrossRef]
- Renwick, J.; Daly, P.; Reeves, E.P.; Kavanagh, K. Susceptibility of larvae of Galleria mellonella to infection by Aspergillus fumigatus is dependent upon stage of conidial germination. Mycopathologia 2006, 161, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Akinci, H.A.; Ozman-Sullivan, S.K.; Diler, H.; Celik, N.; Sullivan, G.T.; Karaca, G. Entomopathogenic Fungi Isolated From Thaumetopoea Pityocampa and Their Efficacies against Its Larvae. Fresenius Environ. Bull. 2017, 26, 5251–5257. [Google Scholar]
- Giraldo, A.; Crous, P.W. Inside Plectosphaerellaceae. Stud. Mycol. 2019, 92, 227–286. [Google Scholar] [CrossRef]
- Singh, S.; Poornesha, B.; Sandhu, R.K.; Ramanujam, B. Natural occurrence of entomopathogenic fungus, Cladosporium cladosporioides on blow fly, Chrysomya megacephala (Fabricius) (Diptera: Calliphoridae) on ber in Punjab, India. J. Biol. Control 2020, 34, 140–143. [Google Scholar] [CrossRef]
- Iskender, N.A.; Ortucu, S.; Aksu, Y.; Saral, A. Isolation, characterization and pathogenicity of fungi from pristiphora abietina (hymenoptera: Tenthredinidae). Fresenius Environ. Bull. 2017, 26, 628–633. [Google Scholar]
- Han, P.; Zhang, X.; Xu, D.; Zhang, B.; Lai, D.; Zhou, L. Metabolites from Clonostachys Fungi and Their Biological Activities. J. Fungi 2020, 6, 229. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, A.A.; Younus, A.S.; Ali, A.N. Efficacy of Clonostachys rosea, as a promising entomopathogenic fungus, against coleopteran stored product insect pests under laboratory conditions. Egypt. J. Biol. Pest Control 2021, 31, 6. [Google Scholar] [CrossRef]
- Su, L.; Zhu, H.; Guo, Y.X.; Du, X.P.; Guo, J.G.; Zhang, L.; Qin, C. Lecanicillium coprophilum (Cordycipitaceae, Hypocreales), a new species of fungus from the feces of Marmota monax in China. Phytotaxa 2019, 387, 55–62. [Google Scholar] [CrossRef]
- Gielen, R.; Meister, H.; Tammaru, T.; Poldmaa, K. Fungi Recorded on Folivorous Lepidoptera: High Diversity Despite Moderate Prevalence. J. Fungi 2021, 7, 25. [Google Scholar] [CrossRef]
- Mongkolsamrit, S.; Khonsanit, A.; Thanakitpipattana, D.; Tasanathai, K.; Noisripoom, W.; Lamlertthon, S.; Himaman, W.; Houbraken, J.; Samson, R.A.; Luangsa-ard, J. Revisiting Metarhizium and the description of new species from Thailand. Stud. Mycol. 2020, 95, 171–251. [Google Scholar] [CrossRef] [PubMed]
- Linnakoski, R.; De Beer, Z.W.; Rousi, M.; Niemela, P.; Pappinen, A.; Wingfield, M.J. Fungi, including Ophiostoma karelicum sp nov., associated with Scolytus ratzeburgi infesting birch in Finland and Russia. Mycol. Res. 2008, 112, 1475–1488. [Google Scholar] [CrossRef]
- Taieb, K.H.; Gharsallah, H.; Ksentini, I.; Leclerque, A.; Schuster, C.; Cheffi, M.; Naayma, S.; Triki, M.A.; Ksantini, M. Screening of biological activities of fungi associated with pistachio bark beetle, Chaetoptelius vestitus (Coleoptera, Curculionidae), infesting pistachio cultivations in Tunisia. J. Appl. Microbiol. 2020, 128, 1472–1485. [Google Scholar] [CrossRef]
- Hernandez-Trejo, A.; Estrada-Drouaillet, B.; Lopez-Santillan, J.A.; Rios-Velasco, C.; Varela-Fuentes, S.E.; Rodriguez-Herrera, R.; Osorio-Hernandez, E. In vitro evaluation of native entomopathogenic fungi and neem (Azadiractha indica) extracts on Spodoptera frugiperda. Phyton-Int. J. Exp. Bot. 2019, 88, 47–54. [Google Scholar]
- Da Costa, G.L.; de Moraes, A.M.; de Oliveira, P.C. Pathogenic action of Penicillium species on mosquito vectors of human tropical diseases. J. Basic Microbiol. 1998, 38, 337–341. [Google Scholar] [CrossRef]
- Dogan, S.; Ocak, I.; Hasenekoglu, I.; Sezek, F. First records of fungi in the families Caligonellidae, Cryptognathidae, Stigmaeidae and Tectocepheidae mites (Arachnida: Acari) from Turkey. Arch. Sci. 2003, 56, 137–142. [Google Scholar]
- Ferraz, C.M.; Soares, F.E.F.; Senna, C.C.; Silva, L.P.C.; Araujo, J.V.; Moreira, T.F.; Monteiro, F.C.; Braga, F.R. Interaction of the nematophagous fungus Pochonia chlamydosporia on eggs of Spartocera dentiventris (Berg) (Hemiptera: Coreidae) under laboratory conditions. Braz. J. Biol. 2021, 81, 1122–1124. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.D.; Liu, X.Z. Occurrence and diversity of insect-associated fungi in natural soils in China. Appl. Soil Ecol. 2008, 39, 100–108. [Google Scholar] [CrossRef]
- Sharma, L.; Oliveira, I.; Torres, L.; Marques, G. Entomopathogenic fungi in Portuguese vineyards soils: Suggesting a ‘Galleria-Tenebrio-bait method’ as bait-insects Galleria and Tenebrio significantly underestimate the respective recoveries of Metarhizium (robertsii) and Beauveria (bassiana). Mycokeys 2018, 38, 1–23. [Google Scholar] [CrossRef]
- Toledo-Hernandez, R.A.; Toledo, J.; Valle-Mora, J.; Holguin-Melendez, F.; Liedo, P.; Huerta-Palacios, G. Pathogenicity and virulence of Purpureocillium lilacinum (Hypocreales: Ophiocordycipitaceae) on Mexican fruit fly adults. Fla. Entomol. 2019, 102, 309–314. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.; Ghosh, S.K. Diversity of soil fungi in North 24 Parganas and their antagonistic potential against Leucinodes orbonalis Guen. (Shoot and fruit borer of brinjal). Environ. Monit. Assess. 2014, 186, 8707–8716. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Goncalves, F.; Oliveira, I.; Torres, L.; Marques, G. Insect-associated fungi from naturally mycosed vine mealybug Planococcus ficus (Signoret) (Hemiptera: Pseudococcidae). Biocontrol Sci. Technol. 2018, 28, 122–141. [Google Scholar] [CrossRef]
- Nonaka, K.; Kaifuchi, S.; Omura, S.; Masuma, R. Five new Simplicillium species (Cordycipitaceae) from soils in Tokyo, Japan. Mycoscience 2013, 54, 42–53. [Google Scholar] [CrossRef]
- Niu, X.; Xie, W.; Zhang, J.; Hu, Q. Biodiversity of Entomopathogenic Fungi in the Soils of South China. Microorganisms 2019, 7, 311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasambala, D.T.; Kasambala, D.T.; Meadow, R.; Meyling, N.V.; Klingen, I. Natural Occurrence of Entomopathogenic Fungi as Endophytes of Sugarcane (Saccharum officinarum) and in Soil of Sugarcane Fields. Insects 2021, 12, 160. [Google Scholar] [CrossRef] [PubMed]
- Ali-Shtayeh, M.S.; Mara’i, A.-B.B.M.; Jamous, R.M. Distribution, occurrence and characterization of entomopathogenic fungi in agricultural soil in the Palestinian area. Mycopathologia 2003, 156, 235–244. [Google Scholar] [CrossRef]
- Niewiadomska, A.; Sawinska, Z.; Wolna-Maruwka, A. Impact of seed dressings on microbiological activity of soil under winter triticale cultivation. Arch. Environ. Prot. 2012, 38, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Dung, J.K.S.; Kaur, N.; Walenta, D.L.; Alderman, S.C.; Frost, K.E.; Hamm, P.B. Reducing Claviceps purpurea sclerotia germination with soil-applied fungicides. Crop Prot. 2018, 106, 146–149. [Google Scholar] [CrossRef]
- Luangsa-ard, J.; Houbraken, J.; van Doorn, T.; Hong, S.-B.; Borman, A.M.; Hywel-Jones, N.L.; Samson, R.A. Purpureocillium, a new genus for the medically important Paecilomyces lilacinus. FEMS Microbiol. Lett. 2011, 321, 141–149. [Google Scholar] [CrossRef]
- Saghrouni, F.; Saidi, W.; Ben, S.Z.; Gheith, S.; Ben, S.M.; Ranque, S.; Denguezli, M. Cutaneous hyalohyphomycosis caused by Purpureocillium lilacinum in an immunocompetent patient: Case report and review. Med. Mycol. 2013, 51, 664–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baidoo, R.; Mengistu, T.; McSorley, R.; Stamps, R.H.; Brito, J.; Crow, W.T. Management of Root-knot Nematode (Meloidogyne incognita) on Pittosporum tobira under Greenhouse, Field, and On-farm Conditions in Florida. J. Nematol. 2017, 49, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Perdomo, H.; Cano, J.; Gene, J.; Garcia, D.; Hernandez, M.; Guarro, J. Polyphasic analysis of Purpureocillium lilacinum isolates from different origins and proposal of the new species Purpureocillium lavendulum. Mycologia 2013, 105, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, M.; Gomez Jimenez, M.I.; Ortiz, V.; Vega, F.E.; Kramer, M.; Parsa, S. Beauveria bassiana and Metarhizium anisopliae endophytically colonize cassava roots following soil drench inoculation. Biol. Control 2016, 95, 40–48. [Google Scholar] [CrossRef] [Green Version]
- Gallou, A.; Serna-Dominguez, M.G.; Berlanga-Padilla, A.M.; Ayala-Zermeno, M.A.; Mellin-Rosas, M.A.; Montesinos-Matias, R.; Arredondo-Bernal, H.C. Species clarification of Isaria isolates used as biocontrol agents against Diaphorina citri (Hemiptera: Liviidae) in Mexico. Fungal Biol. 2016, 120, 414–423. [Google Scholar] [CrossRef]
- Zhang, B.; Zou, C.; Hu, Q. Effects of Isaria fumosorosea on TYLCV (Tomato Yellow Leaf Curl Virus) Accumulation and Transmitting Capacity of Bemisia tabaci. PLoS ONE 2016, 11, e0164356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagele, S.B.; Adhikari, M.; Gurung, S.K.; Lee, H.G.; Kim, S.W.; Kim, H.S.; Ju, H.J.; Gwon, B.H.; Kosol, S.; Lee, Y.S.; et al. New Records of Aspergillus allahabadii and Penicillium sizovae from Crop Field Soil in Korea. Mycobiology 2018, 46, 297–304. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Liu, X.; Guo, S.; Wang, W. Cordyceps industry in China. Mycology 2015, 6, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hu, Q.; Weng, Q. Secondary metabolites (SMs) of Isaria cicadae and Isaria tenuipes. RSC Adv. 2019, 9, 172–184. [Google Scholar] [CrossRef] [Green Version]
- Weng, Q.F.; Zhang, X.F.; Chen, W.; Hu, Q.B. Secondary Metabolites and the Risks of Isaria fumosorosea and Isaria farinosa. Molecules 2019, 24, 664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmaltz, S.; Aita, B.C.; Alves, E.A.; Fochi, A.; Bolson, V.F.; Navarro-Diaz, H.J.; Kuhn, R.C.; Mazutti, M.A. Ultrasound-assisted fermentation for production of beta-1,3-glucanase and chitinase by Beauveria bassiana. J. Chem. Technol. Biot. 2021, 96, 88–98. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Isolates | If Reported EF |
|---|---|---|
| Aspergillus flavus | 1 | Yes. Aphis fabae [19] |
| Aspergillus fumigatus | 1 | Yes. Galleria mellonella [20] |
| Arthropsis hispanica | 2 | No |
| Aspergillus subramanianii | 1 | No |
| Aspergillus tabacinus | 2 | No |
| Aspergillus terreus | 23 | Yes. Thaumetopoea pityocampa [21] |
| Beauveria bassiana | 12 | Yes |
| Brunneochlamydosporium nepalense | 2 | Yes. Myzus persicae [22] |
| Cephalotrichum microsporum | 2 | No |
| Cladosporium cladosporioides | 9 | Yes. Chrysomya megacephala [23] |
| Cladosporium halotolerans | 2 | No |
| Cladosporium sphaerospermum | 1 | Yes. Pristiphora abietina [24] |
| Clonostachys pityrodes | 1 | Yes. Caenothabditis elegans [25] |
| Clonostachys rosea | 15 | Yes. Trogoderma granarium [26] |
| Gonytrichum macrocladum | 4 | No |
| Gliomastix roseogrisea | 1 | Yes. Spodoptera litura |
| Isaria fumosorosea | 3 | Yes |
| Isaria javanica | 19 | Yes |
| Lasionectriopsis spinosa | 1 | No |
| Lecanicillium coprophilum | 10 | Yes. Diaphorina citri [27] |
| Metapochonia bulbillosa | 1 | Yes. Cabera pusaria [28] |
| Metarhizium aciculare | 7 | Yes. Nephotettix virescens [29] |
| Metarhizium anisopliae | 58 | Yes |
| Metarhizium carneum | 3 | Yes |
| Metarhizium flavoviride | 5 | Yes |
| Metarhizium marquandii | 44 | Yes |
| Microascus cirrosus | 1 | No |
| Penicillium brevicompactum | 2 | Yes. Scolytus ratzeburgi [30] |
| Penicillium chrysogenum | 1 | Yes. Chaetoptelius vestitus [31] |
| Penicillium citrinum | 37 | Yes. Spodoptera frugiperda [32] |
| Penicillium janthinellum | 11 | Yes. Aedes aegypti [33] |
| Penicillium manginii | 2 | No |
| Penicillium madriti | 4 | No |
| Penicillium raperi | 11 | Yes. Stigmaeidae mites [34] |
| Pochonia chlamydosporia | 3 | Yes. Spartocera dentiventris [35] |
| Pseudogymnoascus pannorum | 30 | Yes. Galleria mellonella [36] |
| Purpureocillium lavendulum | 30 | Yes. Galleria mellonella [37] |
| Purpureocillium lilacinum | 82 | Yes. Anastrepha ludens [38] |
| Rhinocladiella similis | 7 | No |
| Rhizopus oryzae | 2 | Yes. Leucinodes orbonalis [39] |
| Sarocladium kiliense | 6 | Yes. Vine mealybug [40] |
| Simplicillium subtropicum | 17 | Yes. Hepiaua [41] |
| Talaromyces purpureogenus | 1 | No |
| Talaromyces pinophilum | 11 | Yes. Chaetoptelius vestitus [31] |
| Talaromyces trachyspermus | 2 | No |
| Region | Sample Number | Isolation Rate (%) | Isolate Number | EF Species | Shannon Wiener Index (SWI) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Total | Fungi | EF | Fungi | EF | Total | EF | |||
| Sichuan | 124 | 120 | 120 | 96.77 | 96.77 | 409 | 379 | 29 | 2.98 |
| Yunnan | 40 | 27 | 27 | 67.50 | 67.50 | 36 | 35 | 10 | 1.89 |
| Guizhou | 48 | 35 | 34 | 72.92 | 70.83 | 52 | 45 | 11 | 2.14 |
| Total | 212 | 182 | 181 | 85.85 | 85.38 | 497 | 459 | 32 | - |
| Sample Environment | Sample Numbers | Isolation Rate (%) | Isolate Number | EF Species | Shannon Wiener Index | ||||
|---|---|---|---|---|---|---|---|---|---|
| Total | Fungi | EF | Fungi | EF | Total | EF | |||
| Crop | 63 | 55 | 54 | 87.30 | 85.71 | 162 | 151 | 24 | 2.88 |
| Forest | 41 | 30 | 30 | 73.17 | 73.17 | 71 | 67 | 18 | 2.74 |
| Grassy | 66 | 58 | 58 | 87.88 | 87.88 | 173 | 155 | 24 | 3.05 |
| Orchard | 20 | 19 | 19 | 95.00 | 95.00 | 44 | 42 | 16 | 2.39 |
| Fallow | 22 | 20 | 20 | 90.91 | 90.91 | 47 | 44 | 19 | 2.47 |
| Total | 212 | 182 | 181 | 85.85 | 85.38 | 497 | 459 | 32 | - |
| Isolates | Species | Corrected Mortality (%) (7 d Post-Treatment) | |
|---|---|---|---|
| B. tabaci | S. litura | ||
| AnSC46A07 | Acremonium nepalense | 11.86 ± 1.20 fg | 10.53 ± 2.89 e |
| AhSC46B05 | Arthropsis hispanica | 15.56 ± 2.65 e | 1.76 ± 1.67 h |
| AfSC47B07 | Aspergillus flavus | 12.22 ± 0.58 fg | 7.02 ± 1.67 ef |
| AfSC57C01 | Aspergillus fumigatus | 17.78 ± 3.22 de | 10.53 ± 2.89 e |
| AsSC14C03 | Aspergillus subramanianii | 19.26 ± 0.88 d | 1.76 ± 3.33 h |
| AtSC51B01 | Aspergillus tabacinus | 10.37 ± 0.88 gh | −1.76 ± 1.67 i |
| BbSC09A01 | Beauveria bassiana | 37.78 ± 2.31 b | 36.31 ± 0.00 c |
| CmSC51B07 | Cephalotrichum microsporum | 10.37 ± 2.19 gh | 15.79 ± 2.89 d |
| ChSC55C04 | Cladosporium halotolerans | 7.04 ± 2.73 h | 36.84 ± 2.89 c |
| CrSC40B04 | Clonostachys rossmaniae | −4.07 ± 0.33 k | 8.77 ± 3.33 ef |
| GmSC57A06 | Gonytrichum macrocladum | 10.74 ± 4.91 gh | −1.76 ± 3.33 i |
| IjSC62A03 | Isaria javanica | 57.78 ± 6.93 a | 63.16 ± 5.78 b |
| IfGZ4206 | Isaria fumosorosea | 29.63 ± 4.41 c | 78.95 ± 5.00 a |
| LsSC11A01 | Lasionectriopsis spinosa | 4.44 ± 2.00 i | 1.76 ± 1.67 h |
| LcSC08A01 | Lecanicillium coprophilum | 17.41 ± 0.33 de | 5.26 ± 2.89 f |
| McSC50A01 | Microascus cirrosus | 20.37 ± 4.98 d | 5.26 ± 0.00 f |
| PmSC55C05 | Penicillium madriti | 12.96 ± 1.67 fg | 5.26 ± 2.89 f |
| PmSC62C04 | Penicillium manginii | 8.52 ± 1.20 h | 8.77 ± 1.67 ef |
| PrSC58B03 | Penicillium raperi | 14.44 ± 1.53 ef | 5.26 ± 2.89 f |
| PbSC53C07 | Penicillium brevicompactum | 11.85 ± 1.76 d | 5.26 ± 2.89 f |
| PcSC62C02 | Pochonia chlamydosporia | 10.74 ± 0.33 gh | 7.02 ± 1.67 ef |
| PpSC42A03 | Pseudogymnoascus pannorum | 17.41 ± 0.33 de | −1.76 ± 1.67 i |
| PlSC29A01 | Purpureocillium lilacinum | 28.15 ± 2.91 c | 7.02 ± 4.41 ef |
| RsSC61B02 | Rhinocladiella similis | 12.96 ± 0.33 fg | 5.26 ± 1.67 f |
| SkSC33A03 | Sarocladium kiliense | 6.67 ± 20.8 h | 7.02 ± 1.67 ef |
| SsSC54A04 | Simplicillium subtropicum | 16.30 ± 3.28 e | 10.53 ± 2.89 e |
| TpSC42B04 | Talaromyces pinophilum | 17.04 ± 6.68 de | 10.53 ± 5.78 e |
| TpSC57A05 | Talaromyces purpureogenus | 13.33 ± 3.01 fg | 3.51 ± 1.67 fg |
| TtSC34B04 | Talaromyces trachyspermus | 20.00 ± 3.08 d | 10.53 ± 2.89 e |
| CK | - | 1.67 ± 1.67 j | 3.33 ± 1.67 fg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, W.; Xie, W.; Cai, W.; Thaochan, N.; Hu, Q. Entomopathogenic Fungi Biodiversity in the Soil of Three Provinces Located in Southwest China and First Approach to Evaluate Their Biocontrol Potential. J. Fungi 2021, 7, 984. https://doi.org/10.3390/jof7110984
Chen W, Xie W, Cai W, Thaochan N, Hu Q. Entomopathogenic Fungi Biodiversity in the Soil of Three Provinces Located in Southwest China and First Approach to Evaluate Their Biocontrol Potential. Journal of Fungi. 2021; 7(11):984. https://doi.org/10.3390/jof7110984
Chicago/Turabian StyleChen, Wei, Weiwen Xie, Wei Cai, Narit Thaochan, and Qiongbo Hu. 2021. "Entomopathogenic Fungi Biodiversity in the Soil of Three Provinces Located in Southwest China and First Approach to Evaluate Their Biocontrol Potential" Journal of Fungi 7, no. 11: 984. https://doi.org/10.3390/jof7110984